
Conifer Needle Phyllosphere as a Potential Passive Monitor of Bioaerosolised Antibiotic Resistance Genes

 ,
,
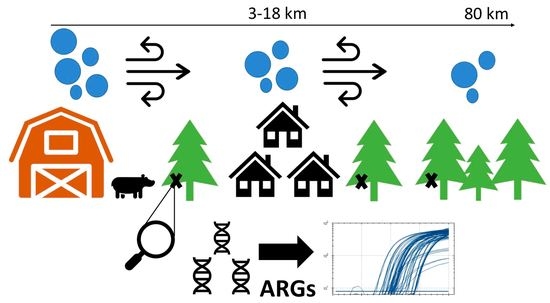
Abstract
:
{kind=link}
{kind=link}
{kind=link}
1. Introduction
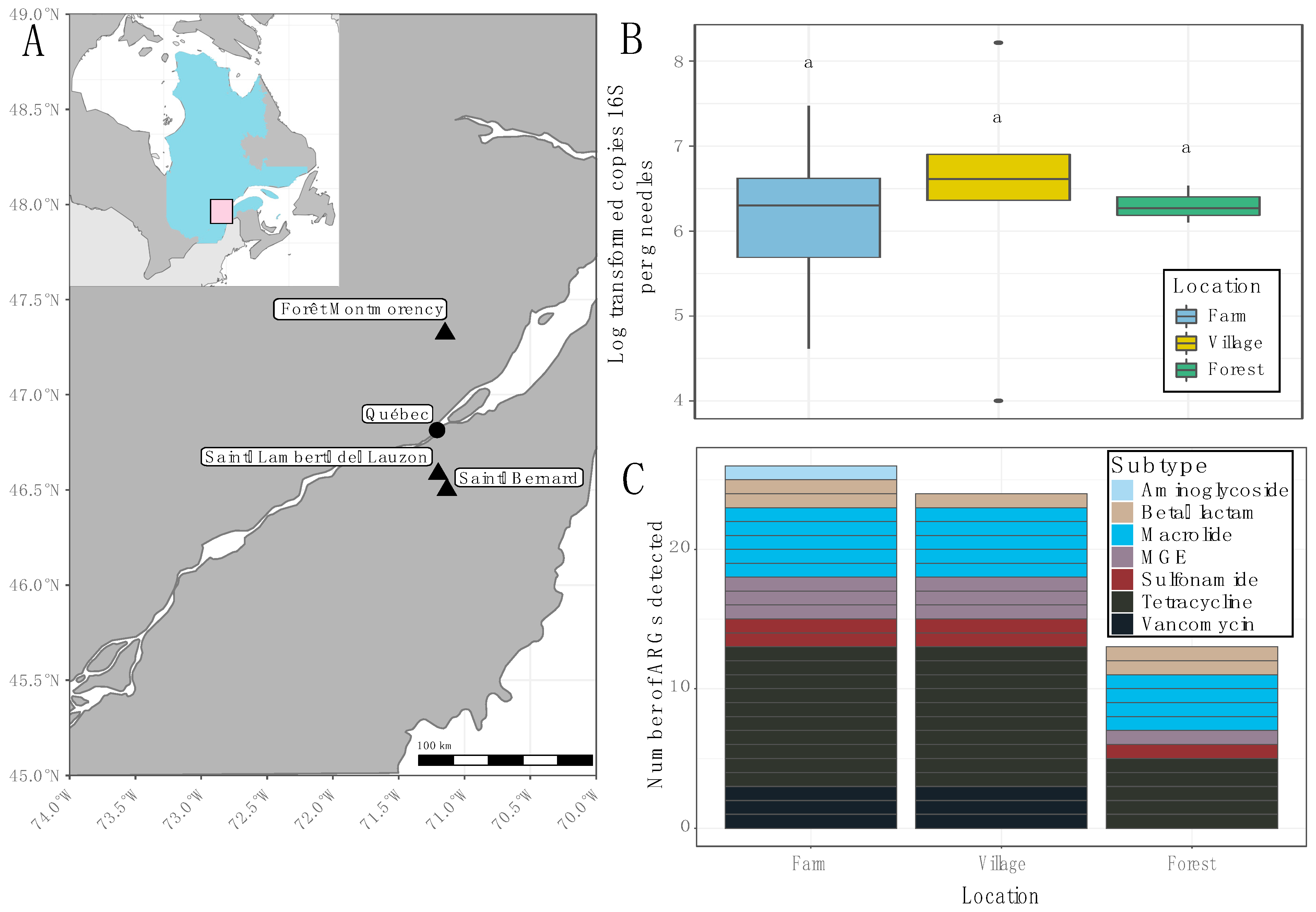
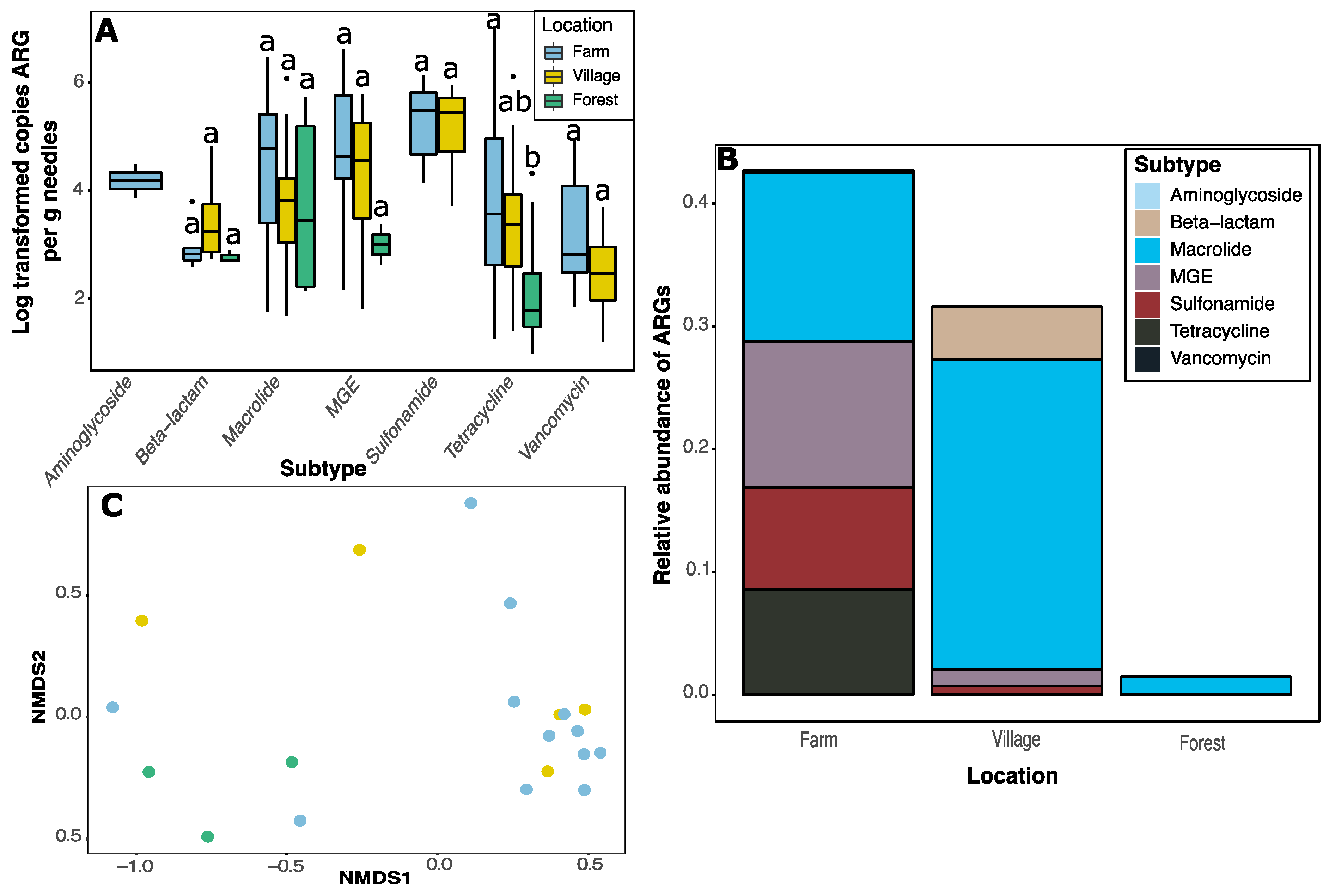
2. Results
3. Discussion
4. Materials & Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Destoumieux-Garzón, D. The One Health Concept: 10 Years Old and a Long Road Ahead. Front. Vet. Sci. 2018, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Robinson, T.P.; Bu, D.P.; Carrique-Mas, J.; Fèvre, E.M.; Gilbert, M.; Grace, D.; Hay, S.I.; Jiwakanon, J.; Kakkar, M.; Kariuki, S.; et al. Antibiotic Resistance Is the Quintessential One Health Issue. Trans. R. Soc. Trop. Med. Hyg. 2016, 110, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of Antimicrobial Resistance in the Environment and Its Relevance to Environmental Regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global Trends in Antimicrobial Use in Food Animals. Proc. Natl. Acad. Sci USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [Green Version]
- He, Y.; Yuan, Q.; Mathieu, J.; Stadler, L.; Senehi, N.; Sun, R.; Alvarez, P.J.J. Antibiotic Resistance Genes from Livestock Waste: Occurrence, Dissemination, and Treatment. NPJ Clean Water 2020, 3, 4. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-Y.; Wang, Y.; Walsh, T.R.; Yi, L.-X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of Plasmid-Mediated Colistin Resistance Mechanism MCR-1 in Animals and Human Beings in China: A Microbiological and Molecular Biological Study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Rhouma, M.; Thériault, W.; Rabhi, N.; Duchaine, C.; Quessy, S.; Fravalo, P. First Identification of Mcr-1/Mcr-2 Genes in the Fecal Microbiota of Canadian Commercial Pigs during the Growing and Finishing Period. VMRR 2019, 10, 65–67. [Google Scholar] [CrossRef] [Green Version]
- Fournier, C.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-Producing Enterobacterales in Pigs in Portugal: Evidence of Direct Links with Antibiotic Selective Pressure. Int. J. Antimicrob. Agents 2020, 55, 105802. [Google Scholar] [CrossRef]
- Nehme, B.; Létourneau, V.; Forster, R.J.; Veillette, M.; Duchaine, C. Culture-Independent Approach of the Bacterial Bioaerosol Diversity in the Standard Swine Confinement Buildings, and Assessment of the Seasonal Effect. Environ. Microbiol. 2008, 10, 665–675. [Google Scholar] [CrossRef]
- Kristiansen, A.; Saunders, A.M.; Hansen, A.A.; Nielsen, P.H.; Nielsen, J.L. Community Structure of Bacteria and Fungi in Aerosols of a Pig Confinement Building. FEMS Microbiol. Ecol. 2012, 80, 390–401. [Google Scholar] [CrossRef]
- Mbareche, H.; Veillette, M.; Pilote, J.; Létourneau, V.; Duchaine, C. Bioaerosols Play a Major Role in the Nasopharyngeal Microbiota Content in Agricultural Environment. Int. J. Environ. Res. Public Health 2019, 16, 1375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galès, A.; Latrille, E.; Wéry, N.; Steyer, J.-P.; Godon, J.-J. Needles of Pinus Halepensis as Biomonitors of Bioaerosol Emissions. PLoS ONE 2014, 9, e112182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manibusan, S.; Mainelis, G. Passive Bioaerosol Samplers: A Complementary Tool for Bioaerosol Research. A Review. J. Aerosol Sci. 2022, 163, 105992. [Google Scholar] [CrossRef]
- Di Guardo, A.; Zaccara, S.; Cerabolini, B.; Acciarri, M.; Terzaghi, G.; Calamari, D. Conifer Needles as Passive Biomonitors of the Spatial and Temporal Distribution of DDT from a Point Source. Chemosphere 2003, 52, 789–797. [Google Scholar] [CrossRef]
- Lin, X.-J.; Barrington, S.; Nicell, J.; Choinière, D.; Vézina, A. Influence of Windbreaks on Livestock Odour Dispersion Plume in the Field. Agric. Ecosyst. Environ. 2006, 116, 263–272. [Google Scholar] [CrossRef]
- Adrizal, A.; Patterson, P.H.; Hulet, R.M.; Bates, R.M.; Myers, C.A.B.; Martin, G.P.; Shockey, R.L.; van der Grinten, M.; Anderson, D.A.; Thompson, J.R. Vegetative Buffers for Fan Emissions from Poultry Farms: 2. Ammonia, Dust and Foliar Nitrogen. J. Environ. Sci. Health Part B 2008, 43, 96–103. [Google Scholar] [CrossRef] [Green Version]
- Larsson, D.G.J.; Flach, C.-F. Antibiotic Resistance in the Environment. Nat. Rev. Microbiol. 2021, 20, 257–269. [Google Scholar] [CrossRef]
- Public Health Agency of Canada. Canadian Antimicrobial Resistance Surveillance System Report—Update 2020. Ottawa, Canada. 2020. Available online: https://www.canada.ca/content/dam/hc-sc/documents/services/drugs-health-products/canadian-antimicrobial-resistance-surveillance-system-2020-report/CARSS-2020-report-2020-eng.pdf (accessed on 6 December 2021).
- Lekagul, A.; Tangcharoensathien, V.; Yeung, S. Patterns of Antibiotic Use in Global Pig Production: A Systematic Review. Vet. Anim. Sci. 2019, 7, 100058. [Google Scholar] [CrossRef]
- Létourneau, V.; Nehmé, B.; Mériaux, A.; Massé, D.; Cormier, Y.; Duchaine, C. Human Pathogens and Tetracycline-Resistant Bacteria in Bioaerosols of Swine Confinement Buildings and in Nasal Flora of Hog Producers. Int. J. Hyg. Environ. Health 2010, 213, 444–449. [Google Scholar] [CrossRef]
- Hong, P.-Y.; Li, X.; Yang, X.; Shinkai, T.; Zhang, Y.; Wang, X.; Mackie, R.I. Monitoring Airborne Biotic Contaminants in the Indoor Environment of Pig and Poultry Confinement Buildings: Biotic Contaminants of Animal Confinement Buildings. Environ. Microbiol. 2012, 14, 1420–1431. [Google Scholar] [CrossRef]
- Wu, N.; Qiao, M.; Zhang, B.; Cheng, W.-D.; Zhu, Y.-G. Abundance and Diversity of Tetracycline Resistance Genes in Soils Adjacent to Representative Swine Feedlots in China. Environ. Sci. Technol. 2010, 44, 6933–6939. [Google Scholar] [CrossRef] [PubMed]
- Jia, S.; He, X.; Bu, Y.; Shi, P.; Miao, Y.; Zhou, H.; Shan, Z.; Zhang, X.-X. Environmental Fate of Tetracycline Resistance Genes Originating from Swine Feedlots in River Water. J. Environ. Sci. Health Part B 2014, 49, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C.; Schwarz, S. Tetracycline and Phenicol Resistance Genes and Mechanisms: Importance for Agriculture, the Environment, and Humans. J. Environ. Qual. 2016, 45, 576–592. [Google Scholar] [CrossRef] [PubMed]
- Hegstad, K.; Mikalsen, T.; Coque, T.M.; Werner, G.; Sundsfjord, A. Mobile Genetic Elements and Their Contribution to the Emergence of Antimicrobial Resistant Enterococcus Faecalis and Enterococcus Faecium. Clin. Microbiol. Infect. 2010, 16, 541–554. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, A.Z.; Cen, T.; Li, X.; Li, D.; Chen, J. Petrol and Diesel Exhaust Particles Accelerate the Horizontal Transfer of Plasmid-Mediated Antimicrobial Resistance Genes. Environ. Int. 2018, 114, 280–287. [Google Scholar] [CrossRef]
- Mazhar, S.H.; Li, X.; Rashid, A.; Su, J.; Xu, J.; Brejnrod, A.D.; Su, J.-Q.; Wu, Y.; Zhu, Y.-G.; Zhou, S.G.; et al. Co-Selection of Antibiotic Resistance Genes, and Mobile Genetic Elements in the Presence of Heavy Metals in Poultry Farm Environments. Sci. Total Environ. 2021, 755, 142702. [Google Scholar] [CrossRef]
- Vorholt, J.A. Microbial Life in the Phyllosphere. Nat. Rev. Microbiol. 2012, 10, 828–840. [Google Scholar] [CrossRef]
- Xiang, Q.; Zhu, D.; Giles, M.; Neilson, R.; Yang, X.-R.; Qiao, M.; Chen, Q.-L. Agricultural Activities Affect the Pattern of the Resistome within the Phyllosphere Microbiome in Peri-Urban Environments. J. Hazard. Mater. 2020, 382, 121068. [Google Scholar] [CrossRef]
- R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 5 May 2018).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package; R Package Version 2.5-7; 2020; Available online: https://CRAN.R-project.org/package=vegan (accessed on 5 May 2018).
- Bach, H.-J.; Tomanova, J.; Schloter, M.; Munch, J.C. Enumeration of Total Bacteria and Bacteria with Genes for Proteolytic Activity in Pure Cultures and in Environmental Samples by Quantitative PCR Mediated Amplification. J. Microbiol. Methods 2002, 49, 235–245. [Google Scholar] [CrossRef]
- Stedtfeld, R.D.; Guo, X.; Stedtfeld, T.M.; Sheng, H.; Williams, M.R.; Hauschild, K.; Gunturu, S.; Tift, L.; Wang, F.; Howe, A.; et al. Primer Set 2.0 for Highly Parallel QPCR Array Targeting Antibiotic Resistance Genes and Mobile Genetic Elements. FEMS Microbiol. Ecol. 2018, 94, fiy130. [Google Scholar] [CrossRef]
- Roschanski, N.; Fischer, J.; Guerra, B.; Roesler, U. Development of a Multiplex Real-Time PCR for the Rapid Detection of the Predominant Beta-Lactamase Genes CTX-M, SHV, TEM and CIT-Type AmpCs in Enterobacteriaceae. PLoS ONE 2014, 9, e100956. [Google Scholar] [CrossRef]
- Nijuis, R.H.T.; Veldman, K.T.; Schelfaut, J.; Van Essen-Zandbergen, A.; Wessels, E.; Claas, E.C.J.; Gooskens, J. Detection of the Plasmid-Mediated Colistin-Resistance Gene Mcr-1 in Clinical Isolates and Stool Specimens Obtained from Hospitalized Patients Using a Newly Developed Real-Time PCR Assay. J. Antimicrob. Chemother. 2016, 71, 2344–2346. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
George, P.B.L.; Leclerc, S.; Turgeon, N.; Veillette, M.; Duchaine, C. Conifer Needle Phyllosphere as a Potential Passive Monitor of Bioaerosolised Antibiotic Resistance Genes. Antibiotics 2022, 11, 907. https://doi.org/10.3390/antibiotics11070907
George PBL, Leclerc S, Turgeon N, Veillette M, Duchaine C. Conifer Needle Phyllosphere as a Potential Passive Monitor of Bioaerosolised Antibiotic Resistance Genes. Antibiotics. 2022; 11(7):907. https://doi.org/10.3390/antibiotics11070907
Chicago/Turabian StyleGeorge, Paul B. L., Samantha Leclerc, Nathalie Turgeon, Marc Veillette, and Caroline Duchaine. 2022. "Conifer Needle Phyllosphere as a Potential Passive Monitor of Bioaerosolised Antibiotic Resistance Genes" Antibiotics 11, no. 7: 907. https://doi.org/10.3390/antibiotics11070907
APA StyleGeorge, P. B. L., Leclerc, S., Turgeon, N., Veillette, M., & Duchaine, C. (2022). Conifer Needle Phyllosphere as a Potential Passive Monitor of Bioaerosolised Antibiotic Resistance Genes. Antibiotics, 11(7), 907. https://doi.org/10.3390/antibiotics11070907