Training Petting Zoo Sheep to Act Like Petting Zoo Sheep: An Empirical Evaluation of Response-Independent Schedules and Shaping with Negative Reinforcement


Abstract
Simple Summary
Abstract
1. Introduction
2. General Methods
2.1. Subjects, Setting, and Material
2.2. Procedure
Response Definitions
3. Experiment 1
3.1. Materials and Methods
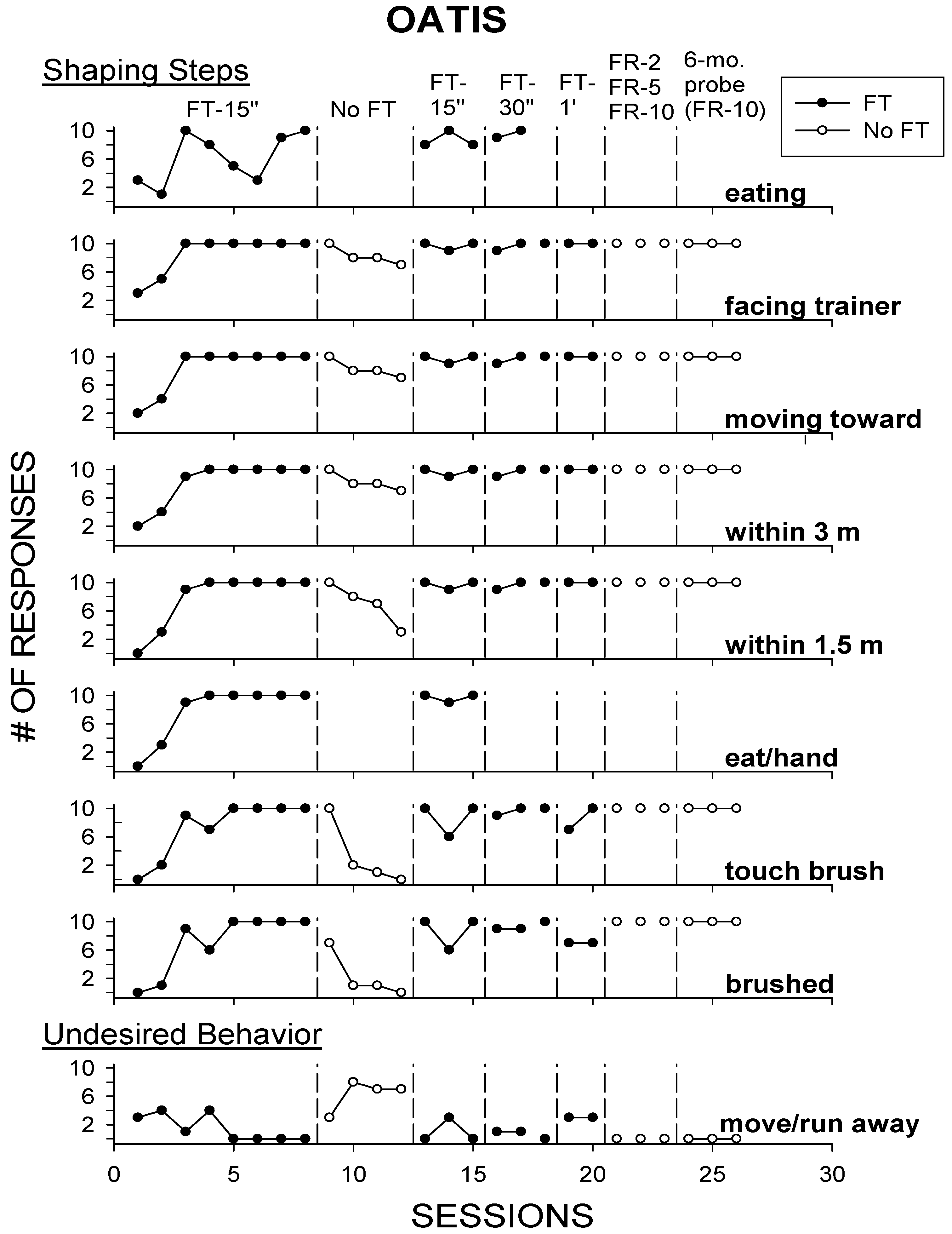
3.1.1. Baseline/Fixed-Time Food Schedule of 15 s
3.1.2. No Fixed-Time Schedule
3.1.3. Return to Fixed-Time Schedule (15 s; 30 s; 1 min)
3.1.4. Fixed-Ratio Schedule (FR-2; FR-5; FR-10)
3.1.5. 6-Month Probe/Fixed-Ratio Schedule (FR-10)
3.2. Results and Discussion
4. Experiment 2
4.1. Materials and Methods
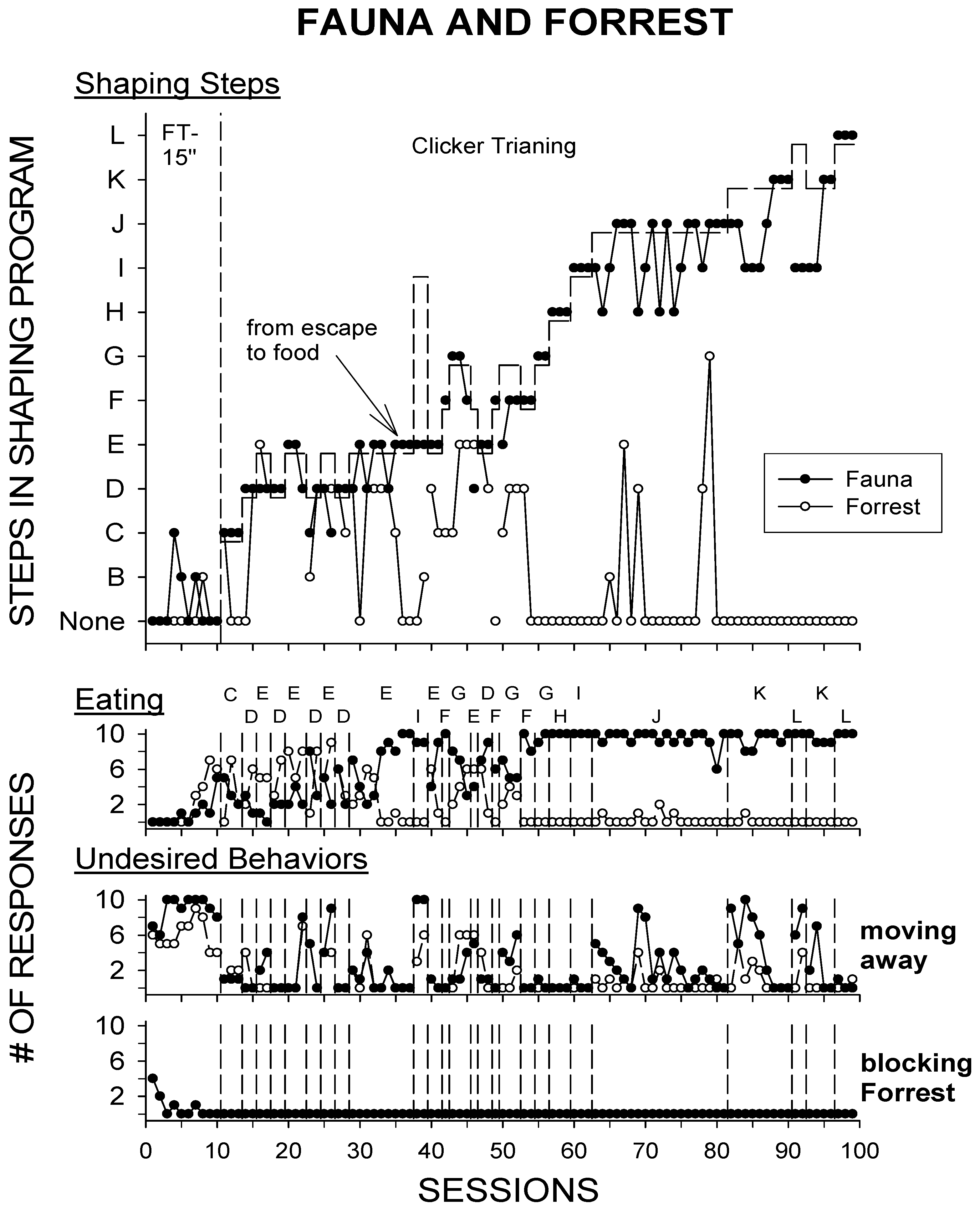
4.1.1. Baseline/FT-15 s
4.1.2. Clicker Training-Negative Reinforcement/Escape (CT R−)
4.1.3. Clicker Training–Positive Reinforcement/Food (CT R+)
4.2. Results and Discussion
5. General Discussion
5.1. Response-Independent Schedules and Approach/Contact Behavior
5.2. Shaping with Negative Reinforcement
5.3. Negative Verse Positive Reinforcement and Aversive Control
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Pryor, K. Don’t shoot the dog! In The New Art of Teaching and Training, 1st ed.; Bantam Books: New York, NY, USA, 1999. [Google Scholar]
- Skinner, B.F. Science and Human Behavior, 1st ed.; Macmillan: New York, NY, USA, 1953. [Google Scholar]
- Skinner, B.F. How to teach animals. Sci. Am. 1951, 185, 26–29. [Google Scholar] [CrossRef]
- Peterson, G.B. A day of great illumination: B. F. Skinner’s discovery of shaping. J. Exp. Anal. Behav. 2004, 82, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Breland, K.; Breland, M. A field of applied animal psychology. Am. Psychol. 1951, 6, 202–204. [Google Scholar] [CrossRef] [PubMed]
- Desmond, T.; Laule, G. Use of positive reinforcement training in the management of species for reproduction. Zoo Biol. 1994, 13, 471–477. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Mutual benefits of research collaborations between zoos and academic institutions. Zoo Biol. 2008, 27, 470–487. [Google Scholar] [CrossRef]
- Forthman, D.L.; Ogden, J.J. The role of applied behavior analysis in zoo management: Today and tomorrow. J. Appl. Behav. Anal. 1992, 25, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, H. Engineering environments for behavioral opportunities in the zoo. Behav. Anal. 1978, 1, 34–47. [Google Scholar] [CrossRef] [PubMed]
- Markowitz, H.; Aday, C.; Gavazzi, A. Effectiveness of acoustic “prey”: Environmental enrichment for a captive African leopard (Panthera pardus). Zoo Biol. 1995, 14, 371–379. [Google Scholar] [CrossRef]
- Laule, G.E.; Thurston, R.H.; Alford, P.L.; Bloomsmith, M.A. Training to reliably obtain blood and urine samples from a diabetic chimpanzee (Pan troglodytes). Zoo Biol. 1996, 15, 587–591. [Google Scholar] [CrossRef]
- Bloomsmith, M.A.; Laule, G.E.; Alford, P.L.; Thurston, R.H. Using training to moderate chimpanzee aggression during feeding. Zoo Biol. 1994, 13, 557–566. [Google Scholar] [CrossRef]
- Bloomsmith, M.A.; Stone, A.M.; Laule, G.E. Positive reinforcement training to enhance the voluntary movement of group–housed chimpanzees within their enclosures. Zoo Biol. 1998, 17, 333–341. [Google Scholar] [CrossRef]
- Hellmuth, H.; Augustine, L.; Watkins, B.; Hope, K. Using operant conditioning and desensitization to facilitate veterinary care with captive reptiles. Vet. Clin. Exot. Anim. Pr. 2012, 15, 425–443. [Google Scholar] [CrossRef] [PubMed]
- Joyce-Zuniga, N.M.; Newberry, R.C.; Robbins, C.T.; Ware, J.V.; Jansen, H.T.; Nelson, O.L. Positive reinforcement training for blood collection in grizzly bears (Ursus arctos horribilis) results in undetectable elevations in serum cortisol levels: A preliminary investigation. J. Appl. Anim. Welf. Sci. 2016, 19, 210–215. [Google Scholar] [CrossRef]
- Grandin, T.; Rooney, M.B.; Phillips, M.; Cambre, R.C.; Irlbeck, N.A.; Graffam, W. Conditioning of Nyala (Tragelaphus angasi) to blood sampling in a crate with positive reinforcement. Zoo Biol. 1995, 14, 261–273. [Google Scholar] [CrossRef]
- Phillips, M.; Grandin, T.; Graffam, W.; Irlbeck, N.A.; Cambre, R.C. Crate conditioning of bongo (Tragelaphus eurycerus) for Veterinary and husbandry procedures at the Denver Zoological Gardens. Zoo Biol. 1998, 17, 25–32. [Google Scholar] [CrossRef]
- Galbicka, G. Shaping in the 21st century: Moving percentile schedules into applied settings. J. Appl. Behav. Anal. 1994, 27, 739–760. [Google Scholar] [CrossRef]
- Pear, J.J.; Legris, J.A. Shaping by automated tracking of an arbitrary operant response. J. Exp. Anal. Behav. 1987, 47, 241–247. [Google Scholar] [CrossRef]
- Ferguson, D.L.; Rosales-Ruiz, J. Loading the problem loader: The effects of target training and shaping on trailer-loading behavior of horses. J. Appl. Behav. Anal. 2001, 34, 409–424. [Google Scholar] [CrossRef]
- Fernandez, E.J. ORCA: A new kind of lab. Clicker J. Mag. Anim. Train. 2001, 51, 18–23. [Google Scholar]
- Fernandez, E.J.; Dorey, N.R. An Examination of Shaping with an African Crested Porcupine (Hystrix cristata). J. Appl. Anim. Welf. Sci. 2020, 1–7. [Google Scholar] [CrossRef]
- Ghaemmaghami, M.; Hanley, G.P.; Jessel, J.; Landa, R. Shaping complex functional communication responses. J. Appl. Behav. Anal. 2018, 51, 502–520. [Google Scholar] [CrossRef] [PubMed]
- Midgley, M.; Lea, S.E.G.; Kirby, R.M. Algorithmic shaping and misbehavior in the acquisition of token deposit by rats. J. Exp. Anal. Behav. 1989, 52, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Osborne, M.L.; Himadi, B. Evaluation of a shaping procedure with the changing-criterion design. Behav. Interv. 1990, 5, 75–81. [Google Scholar] [CrossRef]
- Slater, C.; Dymond, S. Using differential reinforcement to improve equine welfare: Shaping appropriate truck loading and feet handling. Behav. Process. 2011, 86, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Iwata, B.A. Negative reinforcement in applied behavior analysis: An emerging technology. J. Appl. Behav. Anal. 1987, 20, 361–378. [Google Scholar] [CrossRef] [PubMed]
- Pierce, W.D.; Cheney, C.D. Behavior Analysis and Learning, 5th ed.; Psychology Press: London, UK, 2013. [Google Scholar]
- McLean, A.N. The positive aspects of correct negative reinforcement. Anthrozoos 2005, 18, 245–254. [Google Scholar] [CrossRef]
- Warren-Smith, A.K.; McGreevy, P.D. The use of blended positive and negative reinforcement in shaping the halt response of horses (Equus caballus). Anim. Welf. 2007, 16, 481–488. [Google Scholar]
- Rachlin, H.; Hineline, P.N. Training and maintenance of keypecking in the pigeon by negative reinforcement. Science 1967, 157, 954–955. [Google Scholar] [CrossRef]
- Kurland, A. Shaping mental states—Or can you shape love. Am. Anim. Train. Mag. 2000, 1, 14–17. [Google Scholar]
- Delprato, D.J. An animal analogue to systematic desensitization and elimination of avoidance. Behav. Res. Ther. 1973, 11, 49–55. [Google Scholar] [CrossRef]
- Wilson, G.T.; Davison, G.C. Processes of fear reduction in systematic desensitization: Animal studies. Psychol. Bull. 1971, 76, 1–14. [Google Scholar] [CrossRef]
- Wolpe, J. Psychotherapy by reciprocal inhibition. Cond. Reflex 1968, 3, 234–240. [Google Scholar] [PubMed]
- Baer, D.M.; Peterson, R.F.; Sherman, J.A. The development of imitation by reinforcing behavioral similarity to a model. J. Exp. Anal. Behav. 1967, 10, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Thompson, R.H.; Iwata, B.A. A review of reinforcement control procedures. J. Appl. Behav. Anal. 2005, 38, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Carr, J.E.; Coriaty, S.; Wilder, D.A.; Gaunt, B.T.; Dozier, C.L.; Britton, L.N.; Avina, C.; Reed, C.L. A review of “noncontingent” reinforcement as treatment for the aberrant behavior of individuals with developmental disabilities. Res. Dev. Disabil. 2000, 21, 377–391. [Google Scholar] [CrossRef]
- Vollmer, T.R.; Sloman, K.N. The historical context of noncontingent reinforcement as a behavioral treatment. Eur. J. Behav. Anal. 2005, 6, 9–19. [Google Scholar] [CrossRef]
- Andrews, N.L.; Ha, J.C. The effects of automated scatter feeders on captive grizzly bear activity budgets. J. Appl. Anim. Welf. Sci. 2014, 17, 148–156. [Google Scholar] [CrossRef]
- Fernandez, E.J. Stereotypies and Foraging: Appetitive Search Behaviors and Stereotypies in Captive Animals; VDM Publishing: Düsseldorf, Germany, 2010. [Google Scholar]
- Davison, G.C. Systematic desensitization as a counterconditioning process. J. Abnorm. Psychol. 1968, 73, 91. [Google Scholar] [CrossRef]
- Wolpe, J.; Plaud, J.J. Pavlov’s contributions to behavior therapy: The obvious and the not so obvious. Am. Psychol. 1997, 52, 966. [Google Scholar] [CrossRef]
- Liddell, H.S. A comparative approach to the dynamics of experimental neuroses. Ann. N. Y. Acad. Sci. 1953, 56, 164–170. [Google Scholar] [CrossRef]
- Fernandez, E.J.; Timberlake, W. Superstition revisited: Sex, species, and adventitious reinforcement. Behav. Process. 2020, 170, 103979. [Google Scholar] [CrossRef] [PubMed]
- Timberlake, W.; Lucas, G.A. The basis of superstitious behavior: Chance contingency, stimulus substitution, or appetitive behavior? J. Exp. Anal. Behav. 1985, 44, 279–299. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, J. Dogs Are from Neptune, 1st ed.; Dogwise: Wenatchee, WA, USA, 2009. [Google Scholar]
- Hineline, P.N.; Rosales–Ruiz, J. Behavior in relation to aversive events: Punishment and negative reinforcement. In APA Handbooks in Psychology®. APA Handbook of Behavior Analysis Methods and Principles; Madden, G.J., Dube, W.V., Hackenberg, T.D., Hanley, G.P., Lattal, K.A., Eds.; American Psychological Association: Washington, DC, USA, 2013; Volume 1, pp. 483–512. [Google Scholar]
- Rentfro, A.D. Fearful to Friendly (F2F): A Constructional Fear Treatment for Domestic Cats Using a Negative Reinforcement Shaping Procedure in a Home Setting. Master’s Thesis, University of North Texas, Denton, TX, USA, 2012. [Google Scholar]
- Snider, K.S. A Constructional Canine Aggression Treatment: Using a Negative Reinforcement Shaping Procedure with Dogs in Home and Community Settings. Master’s Thesis, University of North Texas, Denton, TX, USA, 2007. [Google Scholar]
- Snider, K. Turning Fierce Dogs Friendly: Using Constructional Aggression Treatment to Rehabilitate Aggressive and Reactive Dogs, 1st ed.; Companion House Books: Mount Joy, PA, USA, 2018. [Google Scholar]
- Stewart, G. Behavior Adjustment Training: BAT for Fear, Frustration, and Aggression in Dogs, 1st ed.; Direct Book Service: Seattle, WA, USA, 2011. [Google Scholar]
- Stewart, G. Behavior Adjustment Training 2.0: New Practical Techniques for Fear, Frustration, and Aggression in Dogs, 1st ed.; Dogwise: Wenatchee, WA, USA, 2016. [Google Scholar]
- Tyner, S.; Brewer, A.; Helman, M.; Leon, Y.; Pritchard, J.; Schlund, M. Nice doggie! Contact desensitization plus reinforcement decreases dog phobias for children with autism. Behav. Anal. Pr. 2016, 9, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Sidman, M. Coercion and Its Fallout, 1st ed.; Authors Cooperative: Boston, MA, USA, 1989. [Google Scholar]
- Skinner, B.F. Beyond Freedom and Dignity, 1st ed.; Bantam Books: New York, NY, USA, 1971. [Google Scholar]
- Baron, A.; Galizio, M. Positive and negative reinforcement: Should the distinction be preserved? Behav. Anal. 2005, 28, 85–98. [Google Scholar] [CrossRef]
- Michael, J. Positive and negative reinforcement, a distinction that is no longer necessary; or a better way to talk about bad things. Behaviorism 1975, 3, 33–44. [Google Scholar] [CrossRef]
- Sidman, M. The distinction between positive and negative reinforcement: Some additional considerations. Behav. Anal. 2006, 29, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.G. What’s wrong with this picture? Effectiveness is not enough. Good Bird Mag. 2008, 4, 12–18. [Google Scholar]
- Critchfield, T.S. Skeptic’s Corner: Punishment—Destructive Force or Valuable Social “Adhesive”? Behav. Anal. Pr. 2014, 7, 36–44. [Google Scholar] [CrossRef]
- Perone, M. Negative effects of positive reinforcement. Behav. Anal. 2003, 26, 1–14. [Google Scholar] [CrossRef]
- Johnston, J.M.; Sherman, R.A. Applying the least restrictive alternative principle to treatment decisions: A legal and behavioral analysis. Behav. Anal. 1993, 16, 103–115. [Google Scholar] [CrossRef]
- Vollmer, T.R.; Hagopian, L.P.; Bailey, J.S.; Dorsey, M.F.; Hanley, G.P.; Lennox, D.; Riordan, M.M.; Spreat, S. The association for behavior analysis international position statement on restraint and seclusion. Behav. Anal. 2011, 34, 103–110. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Behavior/ Step | Definitions | Sessions Exp. 1 | Sessions Exp. 2 |
|---|---|---|---|
| Eating (thrown food) | Any contact of a sheep’s mouth with food thrown to the sheep at the start of a trial. (Exp. 1 did not always start trial w/food). | 1–8; 13–17 (13) | 1–99 (99) |
| Facing Trainer (B) | A sheep’s head faces the direction of the trainer. | N/A | N/A |
| Moving Toward (C) | The sheep moves in the direction of the trainer. | N/A | 11–13 (3) |
| Standing Within (x) (D-H) | The sheep moves within ‘x’ meters of the trainer. D = 3 m, E = 1.5 m, F = 1 m, G = 0.5 m, H = 0.25 m (Exp. 1 only measured 3 and 1.5 m) | N/A | 14–37; 40–59 (44) |
| Eat/Hand (I) | Any contact of a sheep’s mouth with food being held by the trainer. | N/A | 38–39; 60–62 (5) |
| Contact Head/Back (J) | The trainer contacts the sheep’s head/back without the sheep moving away. (Exp. 2 only) | N/A | 63–81 (19) |
| Touch Brush (K) | The sheep voluntarily contacts its nose/mouth to the brush. | N/A | 82–90; 93–96 (13) |
| Brushed (L) | The sheep allows at least 3 front-to-back brushes with the brush across its head/back. | N/A | 91–92; 97–99 (5) |
| Trial-Terminating (Undesired) Responses | |||
| Moving Away | Any movement of more than 0.25 m by a sheep away from the trainer. | N/A | N/A |
| Blocking Forrest | Any movement made by Fauna where ¾ or more of her body is between Forrest and the trainer. (Exp. 2 only) | N/A | N/A |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez, E.J. Training Petting Zoo Sheep to Act Like Petting Zoo Sheep: An Empirical Evaluation of Response-Independent Schedules and Shaping with Negative Reinforcement. Animals 2020, 10, 1122. https://doi.org/10.3390/ani10071122
Fernandez EJ. Training Petting Zoo Sheep to Act Like Petting Zoo Sheep: An Empirical Evaluation of Response-Independent Schedules and Shaping with Negative Reinforcement. Animals. 2020; 10(7):1122. https://doi.org/10.3390/ani10071122
Chicago/Turabian StyleFernandez, Eduardo J. 2020. "Training Petting Zoo Sheep to Act Like Petting Zoo Sheep: An Empirical Evaluation of Response-Independent Schedules and Shaping with Negative Reinforcement" Animals 10, no. 7: 1122. https://doi.org/10.3390/ani10071122
APA StyleFernandez, E. J. (2020). Training Petting Zoo Sheep to Act Like Petting Zoo Sheep: An Empirical Evaluation of Response-Independent Schedules and Shaping with Negative Reinforcement. Animals, 10(7), 1122. https://doi.org/10.3390/ani10071122