Chikungunya Virus Transmission at Low Temperature by Aedes albopictus Mosquitoes

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Results
2.1. Survival Rates
2.2. Effect of Temperature and Genotype on Infection, Dissemination and Transmission of CHIKV
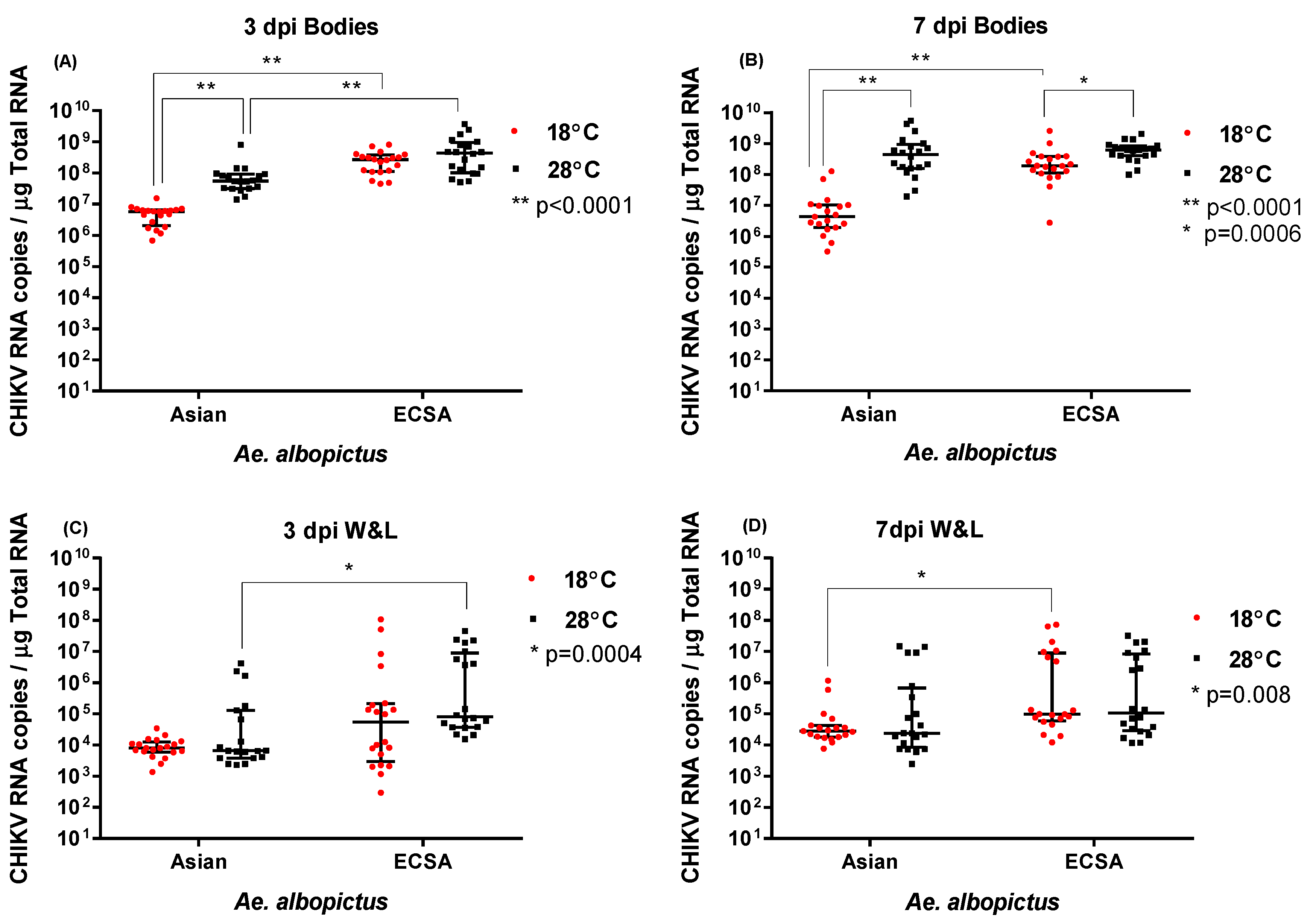
2.3. Effect of temperature and genotype on CHIKV RNA levels
3. Discussion
4. Materials and Methods
4.1. Mosquitoes
4.2. Virus Strains
4.3. Mosquito Oral Challenge
4.4. Nucleic Acid Extraction
4.5. Quantitative RT-PCR to Detect CHIKV
4.6. Cytopathic Effect in Cell Culture
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fischer, D.; Thomas, S.M.; Suk, J.E.; Sudre, B.; Hess, A.; Tjaden, N.B.; Beierkuhnlein, C.; Semenza, J.C. Climate change effects on Chikungunya transmission in Europe: Geospatial analysis of vector’s climatic suitability and virus’ temperature requirements. Int. J. Health Geogr. 2013, 12, 51. [Google Scholar] [CrossRef] [PubMed]
- Manimunda, S.P.; Vijayachari, P.; Uppoor, R.; Sugunan, A.P.; Singh, S.S.; Rai, S.K.; Sudeep, A.B.; Muruganandam, N.; Chaitanya, I.K.; Guruprasad, D.R. Clinical progression of Chikungunya fever during acute and chronic arthritic stages and the changes in joint morphology as revealed by imaging. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Powers, A.M.; Brault, A.C.; Tesh, R.B.; Weaver, S.C. Re-emergence of Chikungunya and O’nyong-nyong viruses: Evidence for distinct geographical lineages and distant evolutionary relationships. J. Gen. Virol. 2000, 81, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Lumsden, W.H.R. An epidemic of virus disease in Southern Province, Tanganyika Territory, in 1952–1953. I. Clinical features. Trans. R. Soc. Trop. Med. Hyg. 1955, 49, 28–32. [Google Scholar] [CrossRef]
- Rezza, G. Chikungunya is back in Italy: 2007–2017. J. Travel Med. 2018, 25. [Google Scholar] [CrossRef] [PubMed]
- European Centre for Disease Prevention and Control. Cluster of Autochtonous Chikungunya Cases in France; ECDC: Stockholm, Sweden, 23 August 2017.
- Reiter, P.; Fontenille, D.; Paupy, C. Aedes albopictus as an epidemic vector of Chikungunya virus: Another emerging problem? Lancet Infect. Dis. 2006, 6, 463–464. [Google Scholar] [CrossRef]
- Borgherini, G.; Poubeau, P.; Staikowsky, F.; Lory, M.; Moullec, N.L.; Becquart, J.P.; Wengling, C.; Michault, A.; Paganin, F. Outbreak of Chikungunya on Reunion island: Early clinical and laboratory features in 157 adult patients. Clin. Infect. Dis. 2007, 44, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Schuffenecker, I.; Iteman, I.; Michault, A.; Murri, S.; Frangeul, L.; Vaney, M.C.; Lavenir, R.; Pardigon, N.; Reynes, J.M.; Pettinelli, F.; et al. Genome microevolution of Chikungunya viruses causing the Indian Ocean outbreak. PLoS Med. 2006, 3, e263. [Google Scholar] [CrossRef]
- Tsetsarkin, K.A.; Vanlandingham, D.; McGee, C.E.; Higgs, S. A single mutation in chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, e201. [Google Scholar] [CrossRef]
- Wimalasiri-Yapa, B.M.C.R.; Stassen, L.; Huang, X.; Hafner, L.M.; Hu, W.; Devine, G.J.; Yakob, L.; Jansen, C.C.; Faddy, H.M.; Viennet, E.; et al. Chikungunya virus in Asia-Pacific: A systematic review. Emerg. Microbes Infect. 2019, 8, 70–79. [Google Scholar] [CrossRef]
- Huang, X.; Hu, W.; Yakob, L.; Devine, G.J.; McGraw, E.A.; Jansen, C.C.; Faddy, H.M.; Frentiu, F.D. El Niño southern oscillation, overseas arrivals and imported Chikungunya cases in Australia: A time series analysis. PLoS Negl. Trop. Dis. 2019, 13, 0007376. [Google Scholar] [CrossRef] [PubMed]
- Viennet, E.; Knope, K.; Faddy, H.M.; Williams, C.R.; Harley, D. Assessing the threat of Chikungunya virus emergence in Australia. Commun. Dis. Intell. Q Rep. 2013, 37, E136–E143. [Google Scholar] [PubMed]
- Hill, M.P.; Axford, J.K.; Hoffmann, A.A. Predicting the spread of Aedes albopictus in Australia under current and future climates: Multiple approaches and datasets to incorporate potential evolutionary divergence. Austral Ecol. 2014, 39, 469–478. [Google Scholar] [CrossRef]
- Van den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Smith, G.A.; Mackenzie, J.S. Vector competence of Australian mosquitoes for Chikungunya virus. Vector Borne Zoonotic Dis. 2010, 10, 489–495. [Google Scholar] [CrossRef] [PubMed]
- Dupont-Rouzeyrol, M.; Caro, V.; Guillaumot, L.; Vazeille, M.; D’Ortenzio, E.; Thiberge, J.-M.; Baroux, N.; Gourinat, A.-C.; Grandadam, M.; Failloux, A.-B. Chikungunya virus and the mosquito vector Aedes aegypti in New Caledonia (south pacific region). Vector Borne Zoonotic Dis. 2012, 12, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Christofferson, R.C.; Chisenhall, D.M.; Wearing, H.J.; Mores, C.N. Chikungunya viral fitness measures within the vector and subsequent transmission potential. PLoS ONE 2014, 9, e110538. [Google Scholar] [CrossRef] [PubMed]
- Coffey, L.L.; Failloux, A.B.; Weaver, S.C. Chikungunya virus-vector interactions. Viruses 2014, 6, 4628–4663. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Aedes aegypti and Aedes albopictus and vectorial capacity for Chikungunya virus. J. Infect. Dis. 2016, 214, S453–S458. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Stige, L.C.; Chan, K.-S.; Zhou, J.; Yang, J.; Sang, S.; Wang, M.; Yang, Z.; Yan, Z.; Jiang, T.; et al. Climate variation drives Dengue dynamics. Proc. Natl. Acad. Sci. USA 2017, 114, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.; Ritchie, S.A.; Russell, R.C.; Zalucki, M.P.; Van Den Hurk, A.F. Ability for Aedes albopictus (Diptera: Culicidae) to survive at the climatic limits of its potential range in eastern Australia. J. Med. Entomol. 2014, 51, 948–957. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Lourenço-de-Oliveira, R.; Mousson, L.; Vazeille, M.; Fuchs, S.; Yébakima, A.; Gustave, J.; Girod, R.; Dusfour, I.; Leparc-Goffart, I.; et al. Chikungunya virus transmission potential by local Aedes mosquitoes in the Americas and Europe. PLoS Negl. Trop. Dis. 2015, 9, e0003780. [Google Scholar] [CrossRef] [PubMed]
- Honório, N.A.; Wiggins, K.; Câmara, D.C.P.; Eastmond, B.; Alto, B.W. Chikungunya virus vector competency of Brazilian and Florida mosquito vectors. PLoS Negl. Trop. Dis. 2018, 12, e0006521. [Google Scholar] [CrossRef] [PubMed]
- Severini, F.; Boccolini, D.; Fortuna, C.; Di Luca, M.; Toma, L.; Amendola, A.; Benedetti, E.; Minelli, G.; Romi, R.; Venturi, G.; et al. Vector competence of Italian Aedes albopictus populations for the Chikungunya virus (E1-226V). PLoS Negl. Trop. Dis. 2018, 12, e0006435. [Google Scholar] [CrossRef] [PubMed]
- Zouache, K.; Fontaine, A.; Vega-Rua, A.; Mousson, L.; Thiberge, J.M.; De-Oliveira, R.L.; Caro, V.; Lambrechts, L.; Failloux, A.B. Three-way interactions between mosquito population, viral strain and temperature underlying Chikungunya virus transmission potential. Proc. Biol. Sci. 2014, 281. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Velez, D.; Lounibos, L.P.; Lord, C.C. Transmission risk of two Chikungunya lineages by invasive mosquito vectors from Florida and the Dominican Republic. PLoS Negl. Trop. Dis. 2017, 11, e0005724. [Google Scholar] [CrossRef]
- Sam, I.C.; Loong, S.-K.; Michael, J.C.; Chua, C.-L.; Wan, S.W.Y.; Vythilingam, I.; Chan, S.-Y.; Chiam, C.-W.; Yeong, Y.-S.; AbuBakar, S.; et al. Genotypic and phenotypic characterization of Chikungunya virus of different genotypes from Malaysia. PLoS ONE 2012, 7, e50476. [Google Scholar] [CrossRef]
- Vega-Rúa, A.; Zouache, K.; Girod, R.; Failloux, A.-B.; Lourenço-de-Oliveira, R. High level of vector competence of Aedes aegypti and Aedes albopictus from ten American countries as a crucial factor in the spread of Chikungunya virus. J. Virol. 2014, 88, 6294–6306. [Google Scholar] [CrossRef]
- Furuya-Kanamori, L.; Liang, S.; Milinovich, G.; Magalhaes, R.J.; Clements, A.C.; Hu, W.; Brasil, P.; Frentiu, F.D.; Dunning, R.; Yakob, L. Co-distribution and co-infection of chikungunya and dengue viruses. BMC Infect. Dis. 2016, 16, 84. [Google Scholar] [CrossRef]


{kind=link}
| dpi | Temperature | % Survival a | |
|---|---|---|---|
| °C | Asian | ECSA | |
| 3 | 18 | 100 (20/20) | 100 (20/20) |
| 28 | 95 (19/20) | 100 (20/20) | |
| 7 | 18 | 95 (19/20) | 100 (20/20) |
| 28 | 100(20/20) | 100 (20/20) | |
| dpi | Temperature | % Infection a | % Dissemination b | % Transmissibility c | |||
|---|---|---|---|---|---|---|---|
| °C | Asian | ECSA | Asian | ECSA | Asian | ECSA | |
| 3 | 18 | 100 (20/20) | 100 (20/20) | 100 (20/20) | 100 (20/20) | 0 (0/20) | 0 (0/20) |
| 28 | 100 (19/19) | 100 (20/20) | 100 (19/19) | 100 (20/20) | 10.53 (2/19) | 25 (5/20) 1 | |
| 7 | 18 | 100 (19/19) | 100 (20/20) | 100 (19/19) | 100 (20/20) | 0 (0/19) | 25 (5/20) 2,3 |
| 28 | 100 (20/20) | 100 (20/20) | 100 (20/20) | 100 (20/20) | 20 (4/20) | 20 (4/20) | |
| dpi | Genotype | 18 °C | 28 °C | |||
|---|---|---|---|---|---|---|
| Median | IQR | Median | IQR | |||
| Bodies | 3 | Asian | 5.76 × 106 | 2.07 × 106–6.49 × 106 | 5.46 × 107 | 3.14 × 107–9.25 × 107 |
| ECSA | 2.67 × 108 | 1.12 × 108–3.78 × 108 | 4.39 × 108 | 1.03 × 108–9.52 × 108 | ||
| 7 | Asian | 4.31 × 106 | 1.93 × 106–1.03 × 107 | 4.41 × 108 | 1.60 × 108–9.60 × 108 | |
| ECSA | 1.91 × 108 | 1.13 × 108–3.87 × 108 | 6.20 × 108 | 4.07 × 108–8.42 × 108 | ||
| Wings and Legs | 3 | Asian | 7.88 × 103 | 5.88 × 103–1.25 × 104 | 6.55 × 103 | 3.86 × 103–1.28 × 105 |
| ECSA | 5.44 × 104 | 2.94 × 103–2.11 × 105 | 8.08 × 104 | 3.70 × 104–8.91 × 106 | ||
| 7 | Asian | 2.81 × 104 | 1.83 × 104–4.25 × 104 | 2.38 × 104 | 8.52 × 103–6.65 × 105 | |
| ECSA | 9.74 × 104 | 5.98 × 104–8.99 × 106 | 1.05 × 105 | 2.84 × 104–8.28 × 106 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wimalasiri-Yapa, B.M.C.R.; Stassen, L.; Hu, W.; Yakob, L.; McGraw, E.A.; Pyke, A.T.; Jansen, C.C.; Devine, G.J.; Frentiu, F.D. Chikungunya Virus Transmission at Low Temperature by Aedes albopictus Mosquitoes. Pathogens 2019, 8, 149. https://doi.org/10.3390/pathogens8030149
Wimalasiri-Yapa BMCR, Stassen L, Hu W, Yakob L, McGraw EA, Pyke AT, Jansen CC, Devine GJ, Frentiu FD. Chikungunya Virus Transmission at Low Temperature by Aedes albopictus Mosquitoes. Pathogens. 2019; 8(3):149. https://doi.org/10.3390/pathogens8030149
Chicago/Turabian StyleWimalasiri-Yapa, B. M. C. Randika, Liesel Stassen, Wenbiao Hu, Laith Yakob, Elizabeth A. McGraw, Alyssa T. Pyke, Cassie C. Jansen, Gregor J. Devine, and Francesca D. Frentiu. 2019. "Chikungunya Virus Transmission at Low Temperature by Aedes albopictus Mosquitoes" Pathogens 8, no. 3: 149. https://doi.org/10.3390/pathogens8030149
APA StyleWimalasiri-Yapa, B. M. C. R., Stassen, L., Hu, W., Yakob, L., McGraw, E. A., Pyke, A. T., Jansen, C. C., Devine, G. J., & Frentiu, F. D. (2019). Chikungunya Virus Transmission at Low Temperature by Aedes albopictus Mosquitoes. Pathogens, 8(3), 149. https://doi.org/10.3390/pathogens8030149