Contrasting Signals of the Westerly Index and North Atlantic Oscillation over the Drought Sensitivity of Tree-Ring Chronologies from the Mediterranean Basin




Abstract
1. Introduction
2. Material and Methods
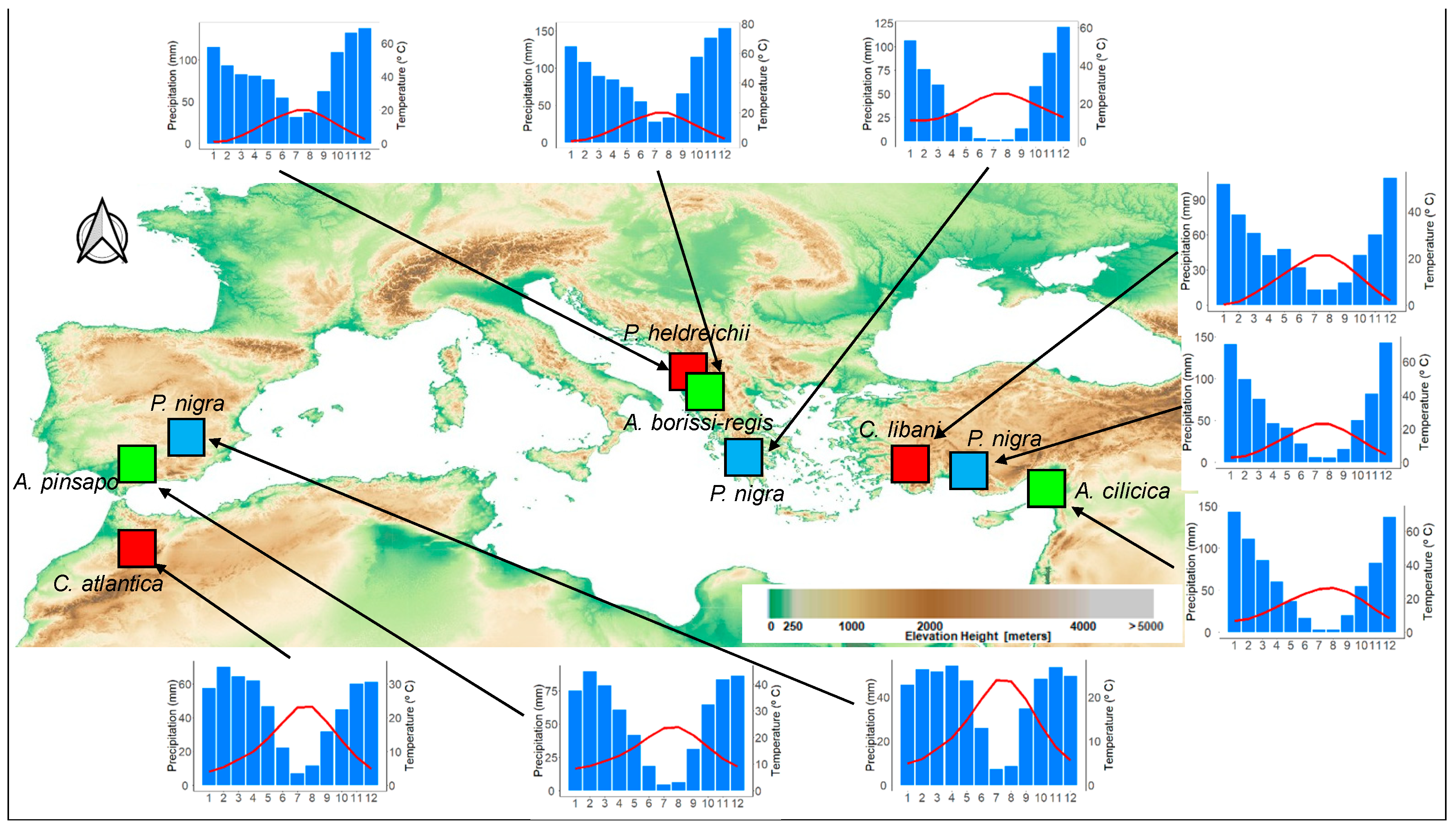
2.1. Study Area and Tree Species
2.2. Dendrochronological Dataset and Chronology Building
2.3. Climate Data, Drought Index, and Large-Scale Atmospheric Oscillations
2.4. Statistical Analysis
3. Results
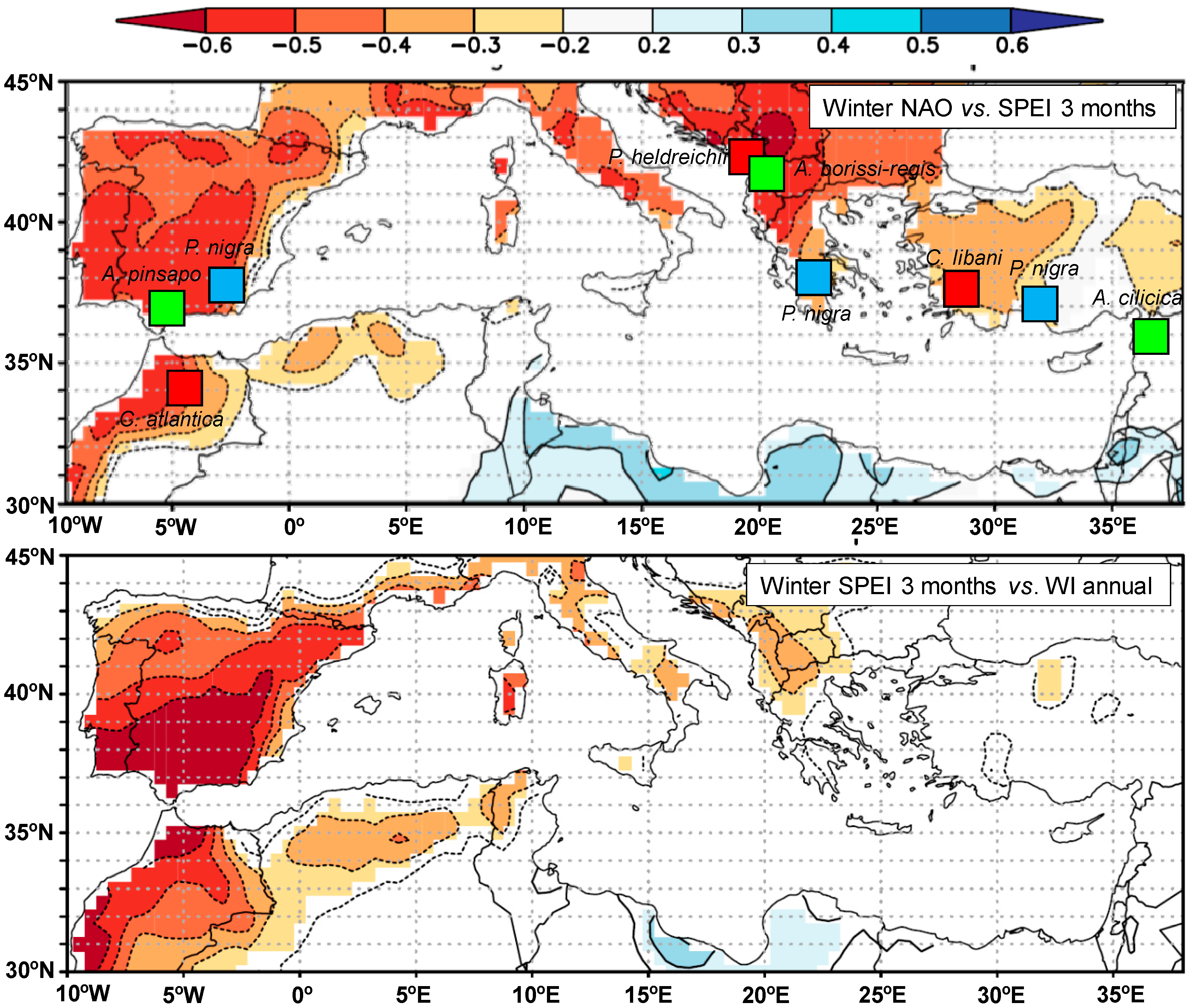
3.1. Atmospheric Circulation Indices and Drought
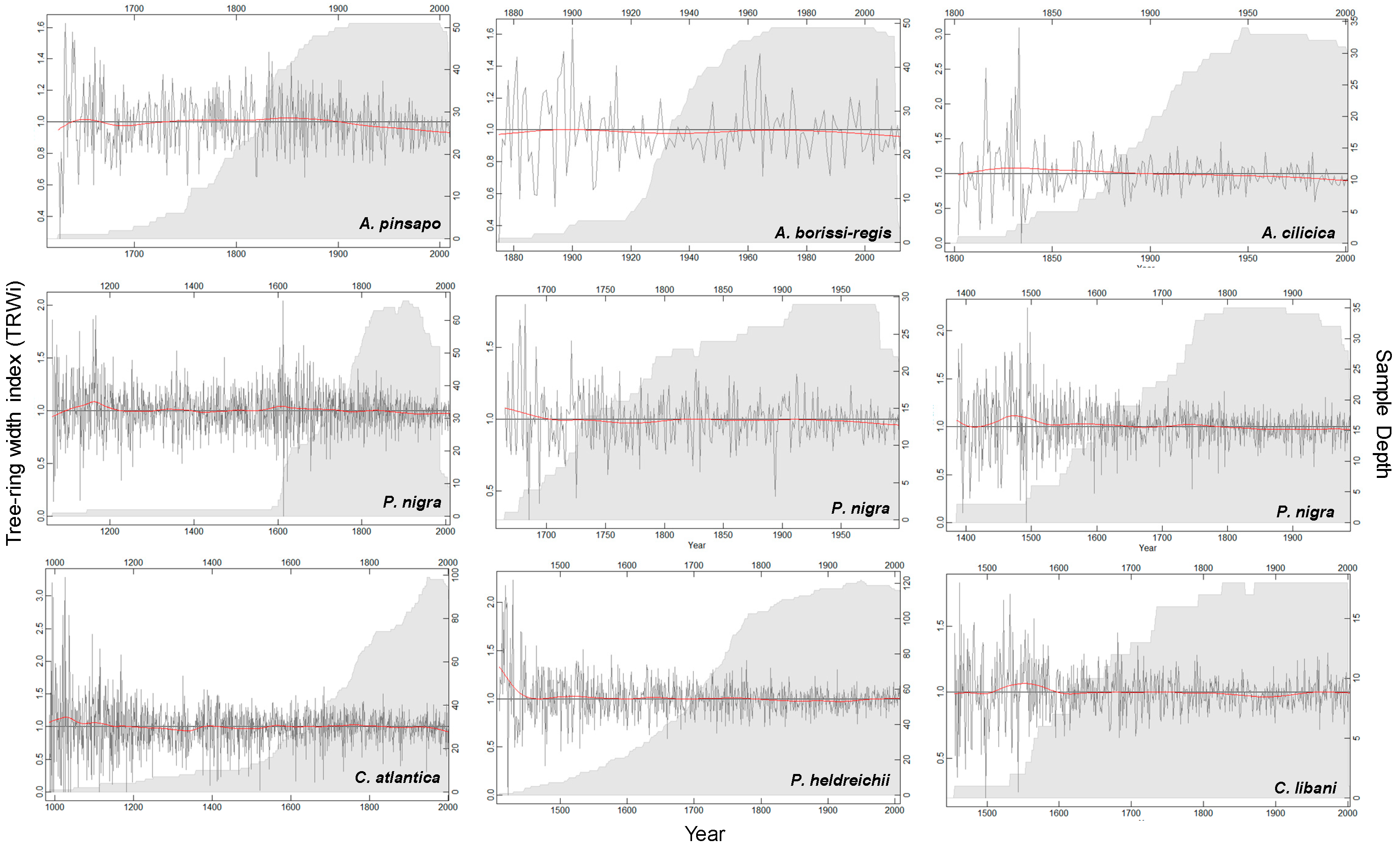
3.2. Chronology Characteristics
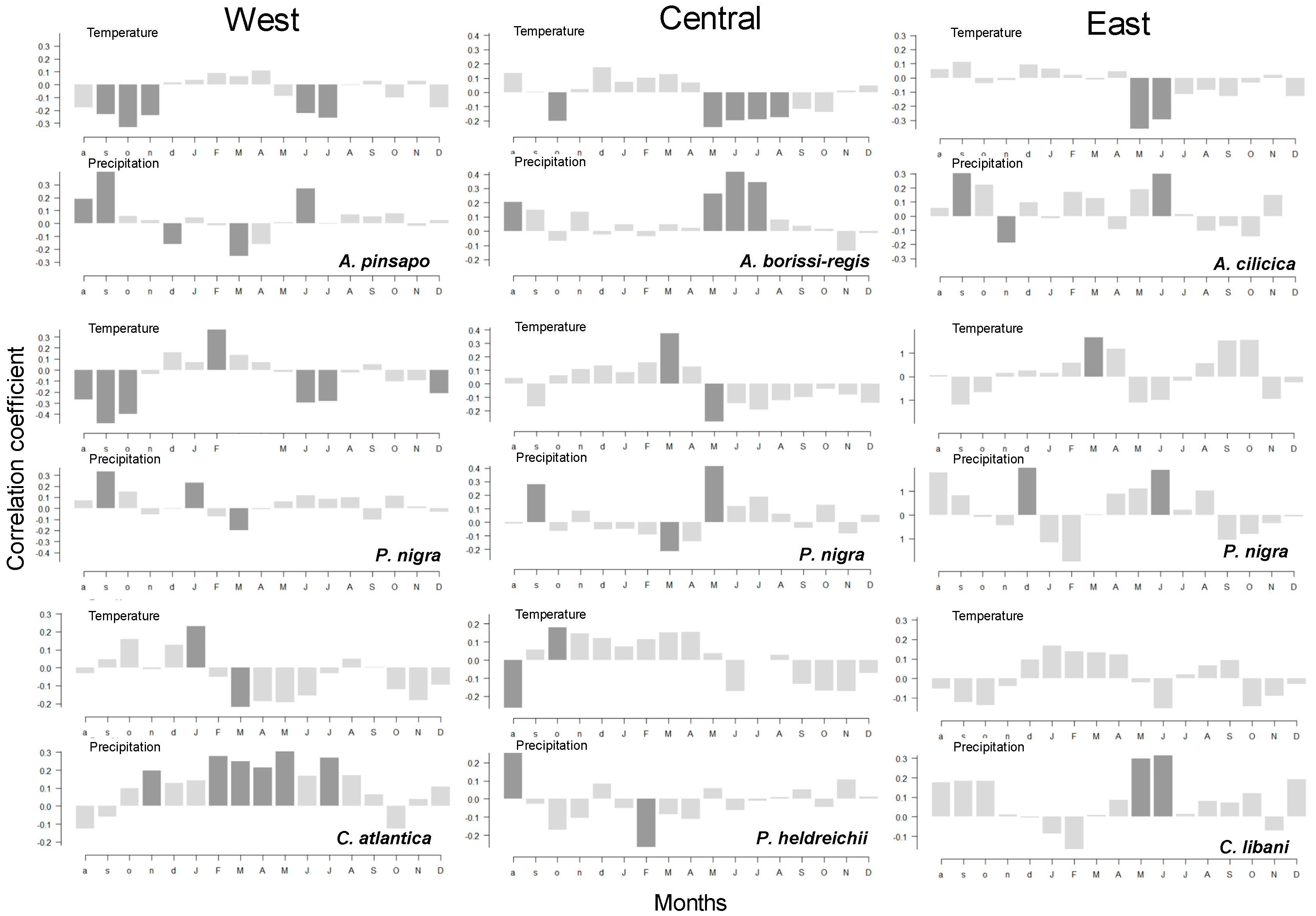
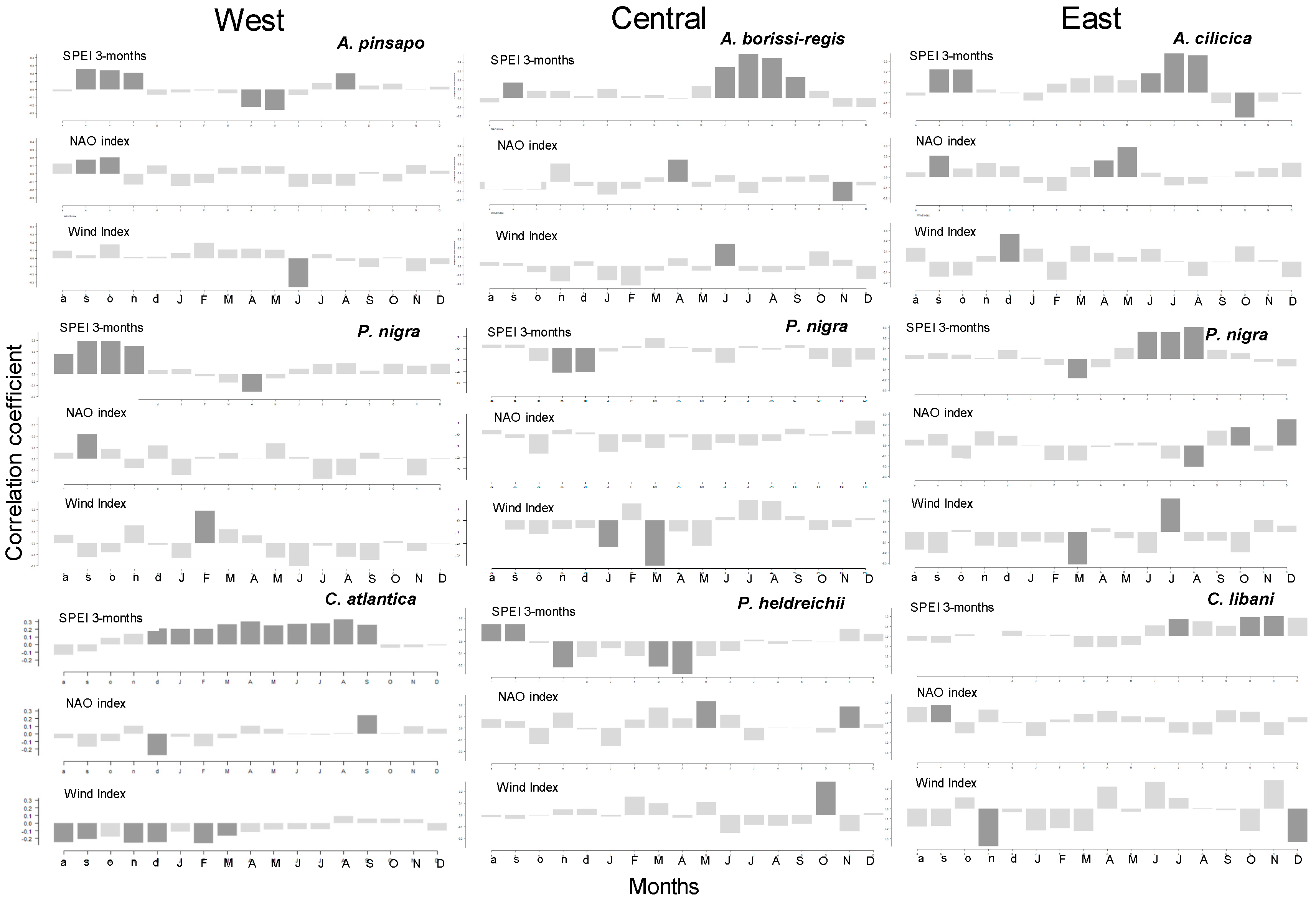
3.3. Climate–Growth Relationships
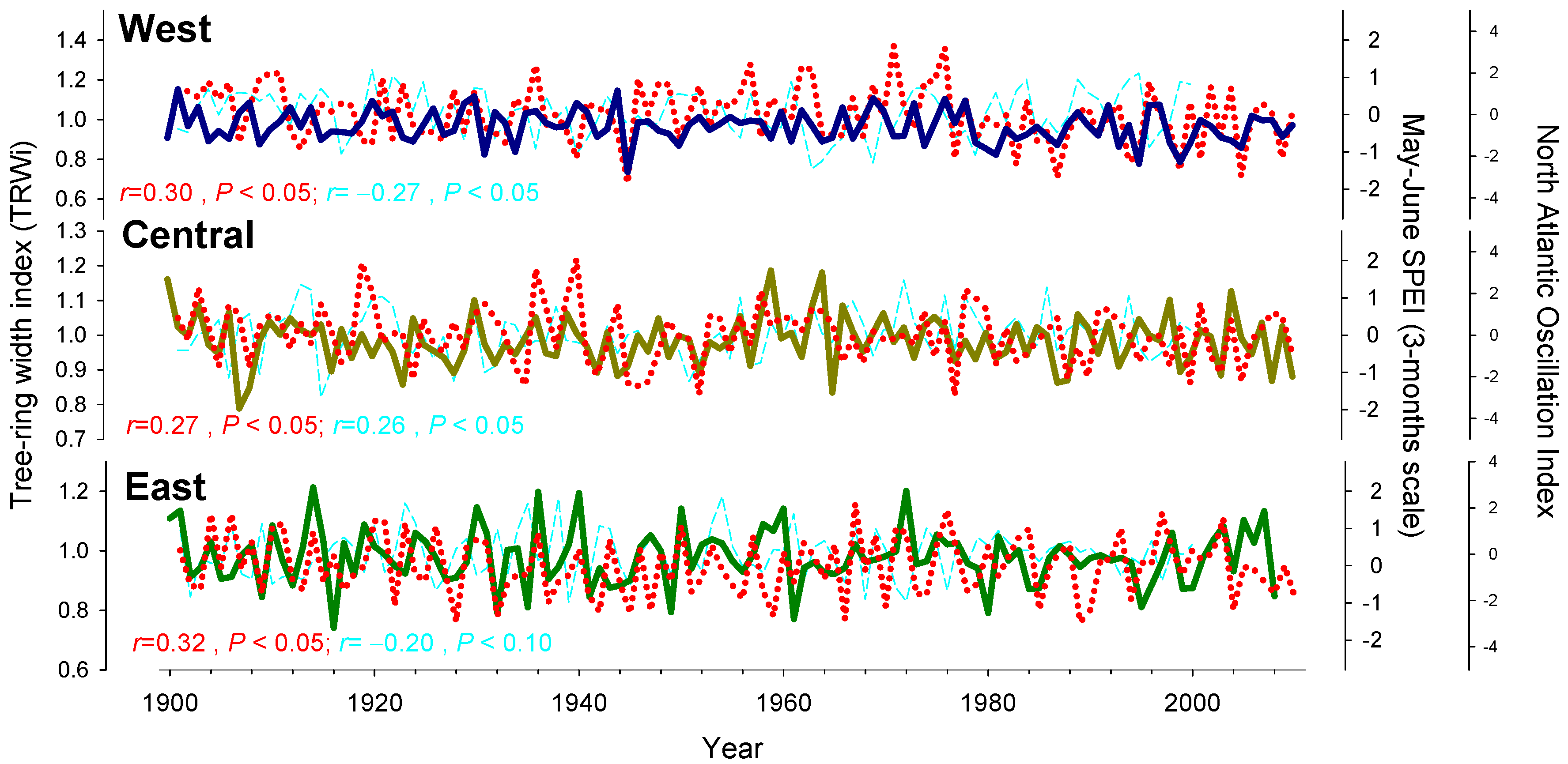
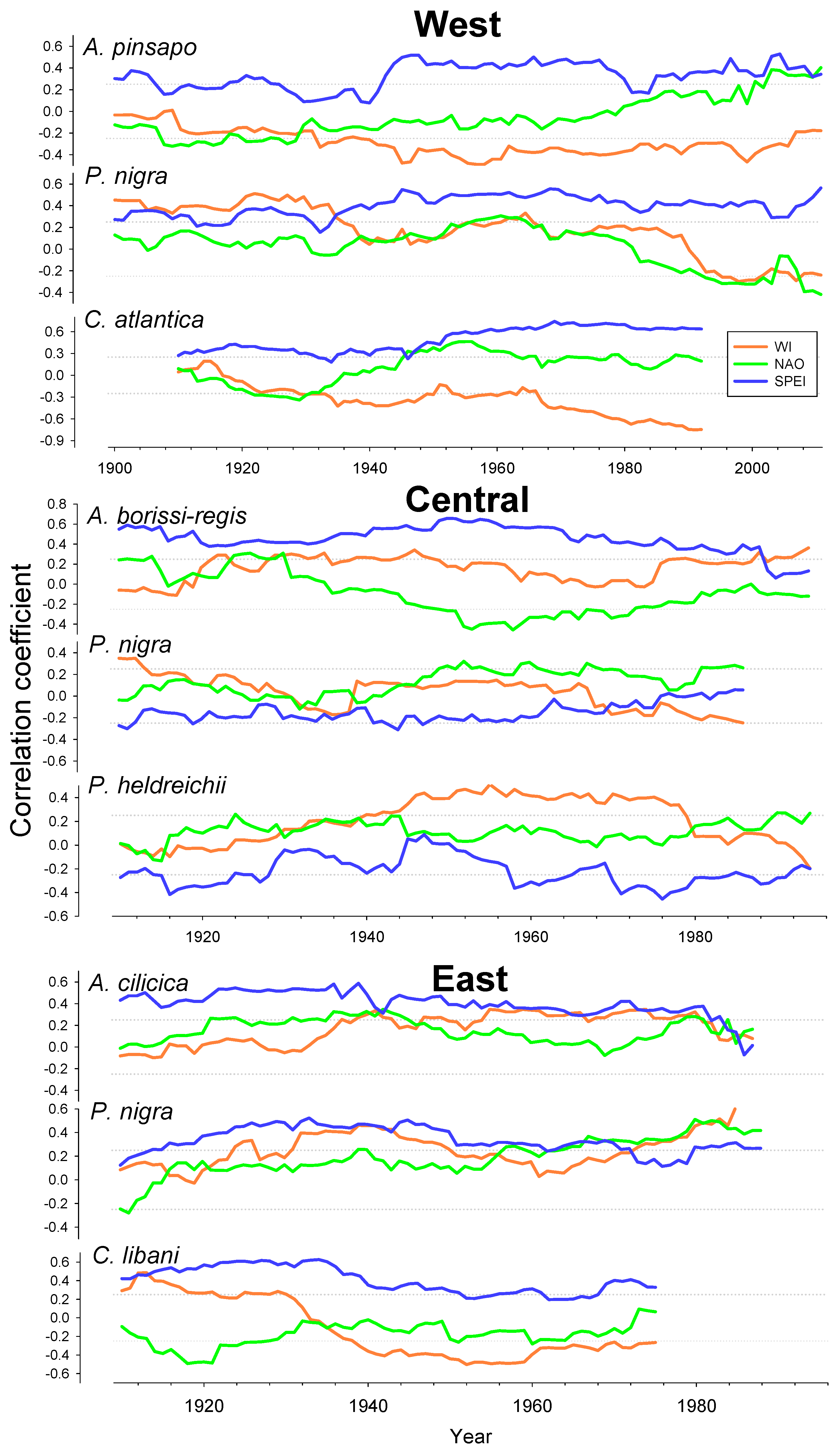
3.4. Relationships between Growth SPEI, NAO, and WI
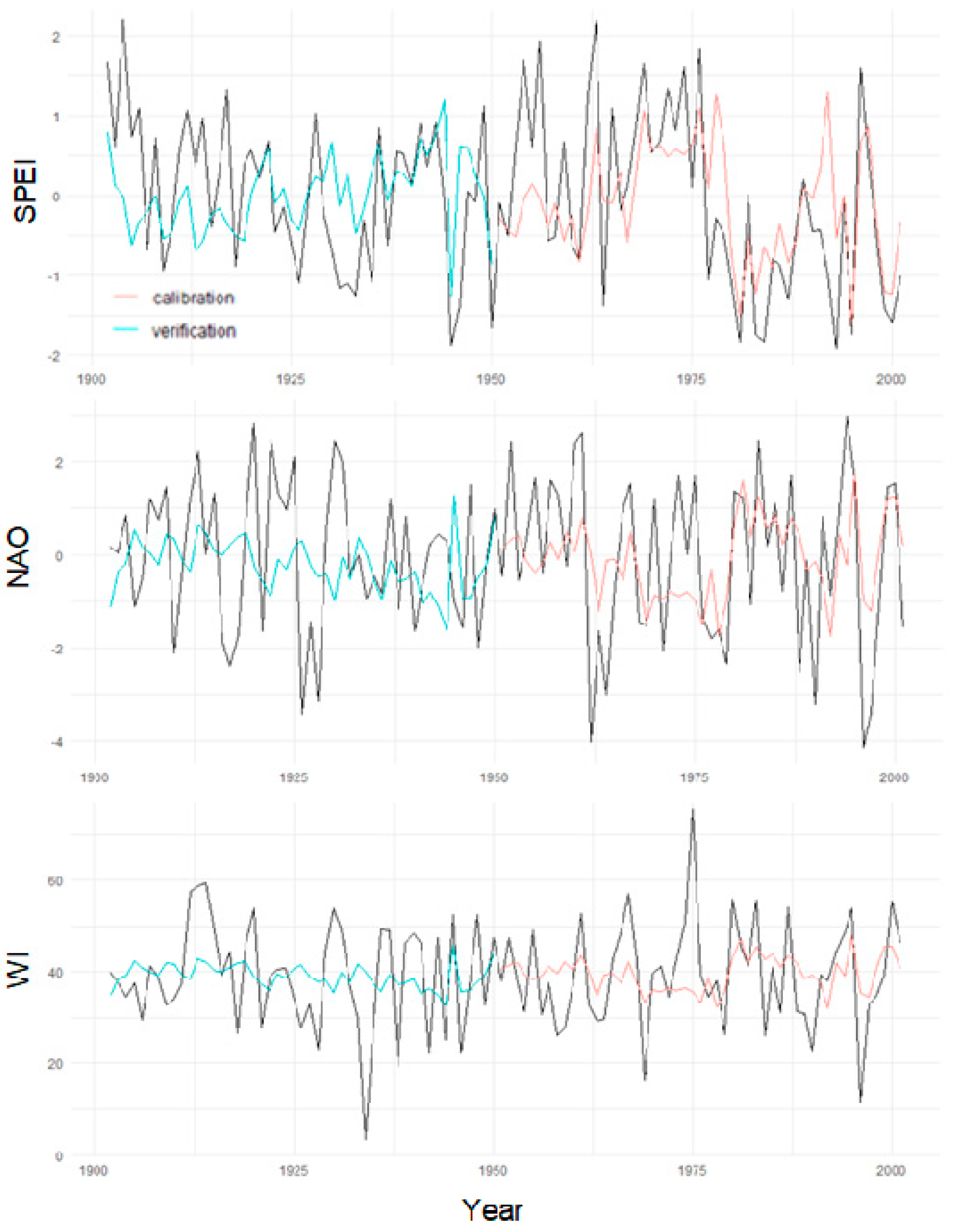
3.5. Exploring WI Reconstructions from Tree Rings
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | West | Central | East | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ABPI | CEAT | PINI | ABBR | PIHE | PINI | ABCI | CELI | PINI | |
| WI | Jun | Ndt−1 | Novt−1 | Jun | Oct | Mar | Dect−1 | Novt−1 | Jul |
| NAO | Octt−1 | Dect−1 | Sept−1 | Apr | May | Octt−1 | May | Sept−1 | Aug |
| SPEI | Aut-1 | Su | Aut−1 | Su | Apr | Wit−1 | Su | Jul | Su |
References
- IPCC. Climate Change 2013—The Physical Science Basis: Working Group I Contribution to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Intergovernmental Panel on Climate Change, Ed.; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781107415324. [Google Scholar]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Chang. 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Hurrell, J.W. Decadal trends in the North Atlantic oscillation: Regional temperatures and precipitation. Science 1995, 269, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Trigo, R.M.; Osborn, T.J.; Corte-Real, J.M. The North Atlantic Oscillation influence on Europe: Climate impacts and associated physical mechanisms. Clim. Res. 2002, 20, 9–17. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Camarero, J.J.; Zabalza, J.; Sangüesa-Barreda, G.; López-Moreno, J.I.; Tague, C.L. Evapotranspiration deficit controls net primary production and growth of silver fir: Implications for Circum-Mediterranean forests under forecasted warmer and drier conditions. Agric. For. Meteorol. 2015, 206, 45–54. [Google Scholar] [CrossRef]
- Madrigal-González, J.; Ballesteros-Cánovas, J.A.; Herrero, A.; Ruiz-Benito, P.; Stoffel, M.; Lucas-Borja, M.E.; Andivia, E.; Sancho-García, C.; Zavala, M.A. Forest productivity in southwestern Europe is controlled by coupled North Atlantic and Atlantic Multidecadal Oscillations. Nat. Commun. 2017, 8, 1–8. [Google Scholar] [CrossRef]
- Luterbacher, J.; García-Herrera, R.; Akcer-On, S.; Allan, R.; Alvarez-Castro, M.C.; Benito, G.; Booth, J.; Büntgen, U.; Cagatay, N.; Colombaroli, D.; et al. A Review of 2000 Years of Paleoclimatic Evidence in the Mediterranean, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2012; ISBN 9780124160422. [Google Scholar]
- Dünkeloh, A.; Jacobeit, J. Circulation dynamics of Mediterranean precipitation variability 1948-98. Int. J. Climatol. 2003, 23, 1843–1866. [Google Scholar] [CrossRef]
- Xoplaki, E.; González-Rouco, J.F.; Luterbacher, J.; Wanner, H. Mediterranean summer air temperature variability and its connection to the large-scale atmospheric circulation and SSTs. Clim. Dyn. 2003, 20, 723–739. [Google Scholar] [CrossRef]
- Seim, A.; Treydte, K.; Trouet, V.; Frank, D.; Fonti, P.; Tegel, W.; Panayotov, M.; Fernández-Donado, L.; Krusic, P.; Büntgen, U. Climate sensitivity of Mediterranean pine growth reveals distinct east-west dipole. Int. J. Climatol. 2014, 35, 2503–2513. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Camarero, J.J.; Sánchez-Salguero, R.; Gutiérrez, E.; Linares, J.C.; Génova, M.; Ribas, M.; Tíscar, P.A.; López-Sáez, J.A. Droughts and climate warming desynchronize Black pine growth across the Mediterranean Basin. Sci. Total Environ. 2019, 697. [Google Scholar] [CrossRef]
- Camarero, J.J. Direct and Indirect Effects of the North Atlantic Oscillation on Tree Growth and Forest Decline in Northeastern Spain. In Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2011; Volume 46, pp. 129–152. [Google Scholar] [CrossRef]
- Camarero, J.J.; Manzanedo, R.D.; Sanchez-Salguero, R.; Navarro-Cerrillo, R.M. Growth response to climate and drought change along an aridity gradient in the southernmost Pinus nigra relict forests. Ann. For. Sci. 2013, 70, 769–780. [Google Scholar] [CrossRef]
- Dorado-Liñán, I.; Zorita, E.; Martínez-Sancho, E.; Gea-Izquierdo, G.; Filippo, A.D.; Gutiérrez, E.; Levanic, T.; Piovesan, G.; Vacchiano, G.; Zang, C.; et al. Large-scale atmospheric circulation enhances the Mediterranean East-West tree growth contrast at rear-edge deciduous forests. Agric. For. Meteorol. 2017, 239, 86–95. [Google Scholar] [CrossRef]
- Rozas, V.; Camarero, J.J.; Sangüesa-Barreda, G.; Souto, M.; García-González, I. Summer drought and ENSO-related cloudiness distinctly drive Fagus sylvatica growth near the species rear-edge in northern Spain. Agric. For. Meteorol. 2015, 201, 153–164. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Lopez-Moreno, J.I.; Beguería, S.; Lorenzo-Lacruz, J.; Sanchez-Lorenzo, A.; García-Ruiz, J.M.; Azorin-Molina, C.; Morán-Tejeda, E.; Revuelto, J.; Trigo, R.; et al. Evidence of increasing drought severity caused by temperature rise in southern Europe. Environ. Res. Lett. 2014, 9, 1–9. [Google Scholar] [CrossRef]
- Hurrell, J.W.; Kushnir, Y.; Visbeck, M. The North Atlantic oscillation. Science 2001, 291, 603–605. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Castro, M. Westerly Index: Un Nuevo Índice de Vientos del Oeste en el Canal de la Mancha a Partir de Observaciones en los Diarios de Navegación de la Royal Navy (1685–2008); Universidad Pablo de Olavide: Sevilla, Spain, 2014. [Google Scholar]
- Glueck, M.F.; Stockton, C.W. Reconstruction of the North Atlantic Oscillation, 1429–1983. Int. J. Climatol. 2001, 21, 1453–1465. [Google Scholar] [CrossRef]
- Barriopedro, D.; Gallego, D.; Alvarez-Castro, M.C.; García-Herrera, R.; Wheeler, D.; Peña-Ortiz, C.; Barbosa, S.M. Witnessing North Atlantic westerlies variability from ships’ logbooks (1685–2008). Clim. Dyn. 2014, 43, 939–955. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Manzano, A.; Clemente, M.A.; Morata, A.; Luna, M.Y.; Beguería, S.; Vicente-Serrano, S.M.; Martín, M.L. Analysis of the atmospheric circulation pattern effects over SPEI drought index in Spain. Atmos. Res. 2019, 230, 104630. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; García-Herrera, R.; Barriopedro, D.; Azorin-Molina, C.; López-Moreno, J.I.; Martín-Hernández, N.; Tomás-Burguera, M.; Gimeno, L.; Nieto, R. The Westerly Index as complementary indicator of the North Atlantic oscillation in explaining drought variability across Europe. Clim. Dyn. 2016, 47, 845–863. [Google Scholar] [CrossRef]
- Till, C.; Guiot, J. Reconstruction of precipitation in Morocco since 1100 A.D. Based on Cedrus atlantica tree-ring widths. Quat. Res. 1990, 33, 337–351. [Google Scholar] [CrossRef]
- Cook, E.R.; D’Arrigo, R.D.; Briffa, K.R. A reconstruction of the North Atlantic Oscillation using tree-ring chronologies from North America and Europe. Holocene 1998, 8, 9–17. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Blackburn Press: Caldwell, NJ, USA, 2001. [Google Scholar]
- Matskovsky, V.; Dolgova, E.; Lomakin, N.; Matveev, S. Dendroclimatology and historical climatology of Voronezh region, European Russia, since 1790s. Int. J. Climatol. 2017, 37, 3057–3066. [Google Scholar] [CrossRef]
- Tejedor, E.; Saz, M.A.; Esper, J.; Cuadrat, J.M.; de Luis, M. Summer drought reconstruction in northeastern Spain inferred from a tree ring latewood network since 1734. Geophys. Res. Lett. 2017, 44, 8492–8500. [Google Scholar] [CrossRef]
- Luterbacher, J.; Xoplaki, E.; Dietrich, D.; Rickli, R.; Jacobeit, J.; Beck, C.; Gyalistras, D.; Schmutz, C.; Wanner, H. Reconstruction of sea level pressure fields over the Eastern North Atlantic and Europe back to 1500. Clim. Dyn. 2002, 18, 545–562. [Google Scholar] [CrossRef]
- Grissino-Mayer, H.D.; Fritts, H.C. The International Tree-Ring Data Bank: An enhanced global database serving the global scientific community. Holocene 1997, 7, 235–238. Available online: http://www.ncdc.noaa.gov/paleo/treering.html (accessed on 13 June 2018). [CrossRef]
- Kuniholm, P.I. Long tree-ring chronologies for the Eastern Mediterranean, Archaeometry 1994. In Proceedings of the 29th International Symposium on Archaeometry, Ankara, Turkey; 1994; pp. 401–409. [Google Scholar]
- Touchan, R.; Xoplaki, E.; Funkhouser, G.; Luterbacher, J.; Hughes, M.K.; Erkan, N.; Akkemik, Ü.; Stephan, J. Reconstructions of spring/summer precipitation for the Eastern Mediterranean from tree-ring widths and its connection to large-scale atmospheric circulation. Clim. Dyn. 2005, 25, 75–98. [Google Scholar] [CrossRef]
- Seim, A.; Buentgen, U.; Fonti, P.; Haska, H.; Herzig, F.; Tegel, W.; Trouet, V.; Treydte, K. Climate sensitivity of a millennium-long pine chronology from Albania. Clim. Res. 2012, 51, 217–228. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.J.; Carrer, M.; Gutiérrez, E.; Alla, A.Q.; Andreu-Hayles, L.; Hevia, A.; Koutavas, A.; Martínez-Sancho, E.; Nola, P.; et al. Climate extremes and predicted warming threaten Mediterranean Holocene firs forests refugia. Proc. Natl. Acad. Sci. USA 2017, 114, E10142–E10150. [Google Scholar] [CrossRef]
- Kuniholm, P.I.; Groneman, C. Pinus Nigra–Taygetos forest, Greece; National Climatic Data Center, NESDIS, NOAA, U.S. Department of Commerce: Boulder, CO, USA, 2005. [Google Scholar]
- Linares, J.C.; Camarero, J.J.; Carreira, J.A. Interacting effects of climate and forest-cover changes on mortality and growth of the southernmost European fir forests. Glob. Ecol. Biogeogr. 2009, 18, 485–497. [Google Scholar] [CrossRef]
- Shestakova, T.A.; Voltas, J.; Saurer, M.; Berninger, F.; Esper, J.; Andreu-Hayles, L.; Daux, V.; Helle, G.; Leuenberger, M.; Loader, N.; et al. Spatio-temporal patterns of tree growth as related to carbon isotope fractionation in European forests under changing climate. Glob. Ecol. Biogeogr. 2019, 28, 1295–1309. [Google Scholar] [CrossRef]
- Esper, J.; Frank, D.; Büntgen, U.; Verstege, A.; Luterbacher, J.; Xoplaki, E. Long-term drought severity variations in Morocco. Geophys. Res. Lett. 2007, 34, L17702. [Google Scholar] [CrossRef]
- Cook, E.R.; Peters, K. Calculating unbiased tree-ring indices for the study of climatic and environmental change. Holocene 1997, 7, 361–370. [Google Scholar] [CrossRef]
- Cook, E. A Time Series Analysis Approach to Tree Ring Standardization; The University of Arizona: Tucson, AZ, USA, 1985. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D.; Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Clim. Appl. Meteorol. 1984, 2, 201–213. [Google Scholar] [CrossRef]
- Mitchell, T.D.; Jones, P.D. An improved method of constructing a database of monthly climate observations and associated high-resolution grids. Int. J. Climatol. 2005, 25, 693–712. Available online: https://crudata.uea.ac.uk/cru/data/pci.htm (accessed on 13 June 2018). [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M. SPEI: Calculation of the Standardised Precipitation-Evapotranspiration Index, R package version 1.7. 2017. Available online: https://rdrr.io/cran/SPEI/ (accessed on 2 June 2020).
- Stenseth, N.C.; Ottersen, G.; Hurrell, J.W.; Mysterud, A.; Lima, M.; Chan, K.S.; Yoccoz, N.G.; Ådlandsvik, B. Studying climate effects on ecology through the use of climate indices: The North Atlantic Oscillation, El Niño Southern Oscillation and beyond. Proc. R. Soc. B Biol. Sci. 2003, 270, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- KNMI Climate Explorer. Available online: http://climexp.knmi.nl/ (accessed on 15 June 2018).
- Vicente-Serrano, S.M.; López-Moreno, J.I.; Lorenzo-Lacruz, J.; Kenawy, A.E.; Azorin-Molina, C.; Morán-Tejeda, E.; Pasho, E.; Zabalza, J.; Beguería, S.; Angulo-Martínez, M. The NAO Impact on Droughts in the Mediterranean Region. In Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2011; Volume 46, pp. 23–40. [Google Scholar] [CrossRef]
- Meko, D.; Graybill, D.A. Tree-ring reconstruction of Upper Gila Rwer Discharge. J. Am. Water Resour. Assoc. 1995, 31, 605–616. [Google Scholar] [CrossRef]
- Zang, C.; Biondi, F. treeclim: An R package for the numerical calibration of proxy-climate relationships. Ecography (Cop.) 2015, 38, 431–436. [Google Scholar] [CrossRef]
- Bunn, A.G. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020.
- Jones, P.D.; Briffa, K.R.; Osborn, T.J.; Lough, J.M.; Van Ommen, T.D.; Vinther, B.M.; Luterbacher, J.; Wahl, E.R.; Zwiers, F.W.; Mann, M.E.; et al. High-resolution palaeoclimatology of the last millennium: A review of current status and future prospects. Holocene 2009, 19, 3–49. [Google Scholar] [CrossRef]
- Lionello, P.; Malanotte-Rizzoli, P.; Boscolo, R. The Mediterranean Climate: An Overview of the Main Characteristics and Issues. Dev. Earth Environ. Sci. 2006, 4, 1–26. [Google Scholar]
- Luterbacher, J.; Xoplaki, E.; Casty, C.; Wanner, H.; Kiittel, M.; Rutishauser, T.; Bronnimann, S.; Fischer, E.; Fleitmann, D.; Gonzalez-rouco, F.J.; et al. Mediterranean Climate Variability over the Last Centuries: A Review. Dev. Earth Environ. Sci. 2006, 4, 27–148. [Google Scholar] [CrossRef]
- Lionello, P.; Abrantes, F.; Gacic, M.; Planton, S.; Trigo, R.; Ulbrich, U. The climate of the Mediterranean region: Research progress and climate change impacts. Reg. Environ. Chang. 2014, 14, 1679–1684. [Google Scholar] [CrossRef]
- Lionello, P. The Climate of the Mediterranean Region: From the Past to the Future; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780123914774. [Google Scholar]
- Jalut, G.; Dedoubat, J.J.; Fontugne, M.; Otto, T. Holocene circum-Mediterranean vegetation changes: Climate forcing and human impact. Quat. Int. 2009, 200, 4–18. [Google Scholar] [CrossRef]
- Mensing, S.A.; Tunno, I.; Sagnotti, L.; Florindo, F.; Noble, P.; Archer, C.; Zimmerman, S.; Pavón-Carrasco, F.J.; Cifani, G.; Passigli, S.; et al. 2700 years of Mediterranean environmental change in central Italy: Asynthesis of sedimentary and cultural records to interpret past impacts of climate on society. Quat. Sci. Rev. 2015, 116, 72–94. [Google Scholar] [CrossRef]
- Ulbrich, U.; Lionello, P.; Belušić, D.; Jacobeit, J.; Knippertz, P.; Kuglitsch, F.G.; Leckebusch, G.C.; Luterbacher, J.; Maugeri, M.; Maheras, P.; et al. Climate of the Mediterranean: Synoptic Patterns, Temperature, Precipitation, Winds, and Their Extremes; Elsevier: Amsterdam, The Netherlands, 2012; ISBN 9780124160422. [Google Scholar]
- Nissen, K.M.; Leckebusch, G.C.; Pinto, J.G.; Renggli, D.; Ulbrich, S.; Ulbrich, U. Cyclones causing wind storms in the Mediterranean: Characteristics, trends and links to large-scale patterns. Nat. Hazards Earth Syst. Sci. 2010, 10, 1379–1391. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- García-Alix, A.; Jiménez-Espejo, F.J.; Toney, J.L.; Jiménez-Moreno, G.; Ramos-Román, M.J.; Anderson, R.S.; Ruano, P.; Queralt, I.; Delgado Huertas, A.; Kuroda, J. Alpine bogs of southern Spain show human-induced environmental change superimposed on long-term natural variations. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Xoplaki, E.; Trigo, R.M.; García-Herrera, R.; Barriopedro, D.; D’Andrea, F.; Fischer, E.M.; Gimeno, L.; Gouveia, C.; Hernández, E.; Kuglitsch, F.G.; et al. Large-Scale Atmospheric Circulation Driving Extreme Climate Events in the Mediterranean and Its Related Impacts; Elsevier: Oxford, UK, 2012; ISBN 9780124160422. [Google Scholar]
- Linares, J.C.; Covelo, F.; Carreira, J.A.; Merino, J.Á. Phenological and water-use patterns underlying maximum growing season length at the highest elevations: Implications under climate change. Tree Physiol. 2012, 32, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Bastos, A.; Janssens, I.A.; Gouveia, C.M.; Trigo, R.M.; Ciais, P.; Chevallier, F.; Peñuelas, J.; Rödenbeck, C.; Piao, S.; Friedlingstein, P.; et al. European land CO2 sink influenced by NAO and East-Atlantic Pattern coupling. Nat. Commun. 2016, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Babst, F.; Bodesheim, P.; Charney, N.; Friend, A.D.; Girardin, M.P.; Klesse, S.; Moore, D.J.P.; Seftigen, K.; Björklund, J.; Bouriaud, O.; et al. When tree rings go global: Challenges and opportunities for retro- and prospective insight. Quat. Sci. Rev. 2018, 197, 1–20. [Google Scholar] [CrossRef]
- Esper, J.; George, S.S.; Anchukaitis, K.; D’Arrigo, R.; Ljungqvist, F.C.; Luterbacher, J.; Schneider, L.; Stoffel, M.; Wilson, R.; Büntgen, U. Large-scale, millennial-length temperature reconstructions from tree-rings. Dendrochronologia 2018, 50, 81–90. [Google Scholar] [CrossRef]
- Touchan, R.; Anchukaitis, K.J.; Meko, D.M.; Attalah, S.; Baisan, C.; Aloui, A. Long term context for recent drought in northwestern Africa. Geophys. Res. Lett. 2008, 35, 1–5. [Google Scholar] [CrossRef]
- Manrique, E.; Fernandez-Cancio, A. Extreme climatic events in dendroclimatic reconstructions from Spain. Clim. Chang. 2000, 44, 123–138. [Google Scholar]
- Akkemik, Ü.; Aras, A. Reconstruction (1689–1994 AD) of April-August precipitation in the southern part of central Turkey. Int. J. Climatol. 2005, 25, 537–548. [Google Scholar] [CrossRef]
- Pozo-Vazquez, D.; Santos-Alamillos, F.J.; Lara-Fanego, V.; Ruiz-Arias, J.A.; Tovar-Pescador, J. Hydrological, Socioeconomic and Ecological Impacts of the North Atlantic Oscillation in the Mediterranean Region; Springer: Dordrecht, The Netherlands, 2011; Volume 46, pp. 213–231. [Google Scholar]







| Variables | West | Central | East | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ABPI | CEAT | PINI | ABBR | PIHE | PINI | ABCI | CELI | PINI | |
| Latitude (N) | 35°10′33″ | 32°58′36″ | 37°49′12″ | 40°21′00″ | 40°42′00″ | 36°55′12″ | 35°34′12″ | 36°40′12″ | 36°27′00″ |
| Longitude (W/E) | 5°11′58″ W | 5°3′35″ W | 2°57′00″ W | 20°23′24″ E | 20°7′58″ E | 22°21′00″ E | 36°12′00″ E | 29°55′12″ E | 32°31′12″ E |
| Elevation (m a.s.l.) | 1800 | 2200 | 1800 | 1144 | 2000 | 1400 | 1450 | 1800 | 1580 |
| Aspect | N | N | E | N | W | NW | W | NW | W |
| MAP | 560 | 510 | 483 | 973 | 1007 | 565 | 721 | 769 | 691 |
| MAT | 16.7 | 13.4 | 12.1 | 10.6 | 10.1 | 17.6 | 17.3 | 14.7 | 13.1 |
| Nºof trees/Nº of cores | 26/52 | 50/100 | 35/69 | 49/49 | 124/124 | 30/30 | 34/34 | 36/36 | 19/19 |
| Mean/Max Age (years) | 211/397 | 309/1024 | 272/947 | 94/122 | 310/604 | 232/343 | 111/207 | 348/619 | 332/557 |
| Title | SPEI | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| West | Central | East | ||||||||
| Season | NAO | ABPI | CEAT | PINI | ABBR | PIHE | PINI | ABCI | CELI | PINI |
| WIan | 0.49 | −0.39 | −0.14 | −0.22 | −0.03 | −0.19 | −0.01 | −0.26 | −0.02 | −0.04 |
| WIwi | 0.79 | −0.61 | −0.36 | −0.53 | −0.10 | −0.10 | 0.02 | −0.11 | −0.04 | −0.08 |
| Wisp | 0.50 | −0.41 | 0.06 | −0.09 | −0.12 | −0.25 | −0.20 | −0.14 | −0.23 | −0.08 |
| WIsu | 0.34 | 0.03 | −0.01 | −0.08 | −0.02 | −0.13 | −0.06 | −0.29 | −0.02 | −0.14 |
| WIau | 0.56 | −0.30 | −0.17 | −0.30 | −0.12 | −0.21 | 0.09 | −0.12 | 0.03 | 0.03 |
| Variables | West | Central | East | ||||||
|---|---|---|---|---|---|---|---|---|---|
| ABPI | CEAT | PINI | ABBR | PIHE | PINI | ABCI | CELI | PINI | |
| Tree-ring width (mm) ± SD | 1.07 ± 0.36 | 0.76 ± 0.23 | 0.56 ± 0.12 | 2.67 ± 0.63 | 0.80 ± 0.15 | 0.93 ± 0.28 | 1.02 ± 0.36 | 1.29 ± 0.22 | 0.58 ± 0.11 |
| Time span | 1614–2011 | 977–2001 | 1050–2010 | 1874–2012 | 1405–2008 | 1657–1999 | 1795–2001 | 1370–1988 | 1444–2003 |
| AC | 0.832 | 0.791 | 0.842 | 0.759 | 0.815 | 0.807 | 0.810 | 0.802 | 0.844 |
| MS | 0.237 | 0.311 | 0.248 | 0.243 | 0.213 | 0.244 | 0.221 | 0.284 | 0.221 |
| Rbt | 0.533 | 0.756 | 0.590 | 0.464 | 0.570 | 0.496 | 0.474 | 0.575 | 0.460 |
| SNR | 4.399 | 3.639 | 7.207 | 5.621 | 2.192 | 9.095 | 6.703 | 11.93 | 5.372 |
| PC1 (%) | 37.22 | 58.09 | 39.30 | 33.05 | 36.05 | 36.56 | 30.50 | 35.05 | 26.35 |
| EPS 1930–2010 | 0.925 | 0.992 | 0.976 | 0.951 | 0.992 | 0.952 | 0.971 | 0.905 | 0.943 |
| Month | SPEI 3 | West | Central | East | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| NAO | WI | ABPI | CEAT | PINI | ABBR | PIHE | PINI | ABCI | CELI | PINI | |
| January | 0.06 | 0.04 | −0.02 | −0.10 | −0.07 | −0.17 | −0.17 | 0.01 | 0.07 | 0.01 | −0.03 |
| February | 0.20 | −0.01 | −0.08 | −0.10 | −0.11 | −0.18 | −0.14 | 0.01 | −0.05 | 0.01 | −0.10 |
| March | 0.24 | 0.16 | −0.16 | −0.08 | −0.15 | −0.09 | −0.08 | −0.05 | −0.10 | −0.08 | −0.10 |
| April | −0.16 | −0.08 | −0.12 | −0.11 | −0.19 | −0.01 | 0.00 | −0.07 | −0.04 | −0.08 | −0.09 |
| May | 0.02 | 0.07 | −0.15 | −0.17 | −0.21 | −0.03 | −0.10 | −0.01 | −0.01 | −0.01 | −0.12 |
| June | −0.10 | −0.09 | −0.10 | −0.16 | −0.23 | 0.01 | −0.06 | 0.01 | 0.07 | −0.01 | 0.06 |
| July | 0.00 | 0.03 | −0.18 | −0.11 | −0.17 | 0.02 | −0.13 | 0.07 | 0.01 | 0.05 | −0.01 |
| August | 0.02 | −0.07 | −0.13 | 0.07 | −0.09 | 0.02 | −0.06 | 0.05 | 0.06 | 0.06 | 0.02 |
| September | −0.12 | −0.09 | 0.03 | 0.15 | 0.10 | 0.01 | −0.01 | 0.15 | −0.05 | 0.20 | 0.09 |
| October | 0.03 | 0.07 | 0.09 | 0.16 | 0.05 | −0.08 | −0.01 | 0.18 | 0.03 | 0.20 | 0.03 |
| November | 0.06 | 0.08 | 0.07 | 0.07 | 0.05 | −0.16 | −0.07 | 0.13 | 0.08 | 0.15 | 0.04 |
| December | −0.07 | −0.08 | 0.09 | −0.02 | 0.00 | −0.16 | −0.16 | 0.07 | 0.09 | 0.07 | 0.00 |
| West | 1903–1953 | 1954–2001 | 1903–2001 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| RE | CE | DW | r | RE | CE | DW | R | Intercept | Slope | r | |
| SPEIsu | 0.175 | 0.502 | 1.516 | 0.58 | 0.353 | 1.049 | 1.517 | 0.62 | −1.975 | 1.979 | 0.52 |
| NAOdt-1 | 0.243 | 0.515 | 1.232 | 0.26 | 0.218 | 0.223 | 1.974 | 0.42 | 1.601 | −1.666 | 0.29 |
| WIndt-1 | 0.025 | 0.020 | 1.714 | 0.33 | 0.188 | 0.191 | 1.294 | 0.30 | 50.136 | −10.973 | 0.28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Casas-Gómez, P.; Sánchez-Salguero, R.; Ribera, P.; Linares, J.C. Contrasting Signals of the Westerly Index and North Atlantic Oscillation over the Drought Sensitivity of Tree-Ring Chronologies from the Mediterranean Basin. Atmosphere 2020, 11, 644. https://doi.org/10.3390/atmos11060644
Casas-Gómez P, Sánchez-Salguero R, Ribera P, Linares JC. Contrasting Signals of the Westerly Index and North Atlantic Oscillation over the Drought Sensitivity of Tree-Ring Chronologies from the Mediterranean Basin. Atmosphere. 2020; 11(6):644. https://doi.org/10.3390/atmos11060644
Chicago/Turabian StyleCasas-Gómez, Pablo, Raúl Sánchez-Salguero, Pedro Ribera, and Juan C. Linares. 2020. "Contrasting Signals of the Westerly Index and North Atlantic Oscillation over the Drought Sensitivity of Tree-Ring Chronologies from the Mediterranean Basin" Atmosphere 11, no. 6: 644. https://doi.org/10.3390/atmos11060644
APA StyleCasas-Gómez, P., Sánchez-Salguero, R., Ribera, P., & Linares, J. C. (2020). Contrasting Signals of the Westerly Index and North Atlantic Oscillation over the Drought Sensitivity of Tree-Ring Chronologies from the Mediterranean Basin. Atmosphere, 11(6), 644. https://doi.org/10.3390/atmos11060644