
Evaluating the Spectral and Physiological Responses of Grapevines (Vitis vinifera L.) to Heat and Water Stresses under Different Vineyard Cooling and Irrigation Strategies

 ,
,  ,
,  ,
,  ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
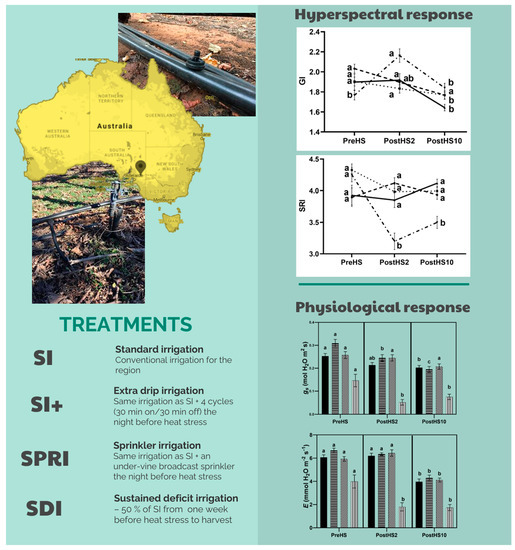
2.1. Study Area and Experimental Design
- Standard drip irrigation (SI)—conventional irrigation for the region, growers applied 4 h of additional irrigation during the day preceding HS. Irrigation was applied using a single dripline with pressure-compensating emitters spaced 0.3 m apart, each with a flow rate of 1 L h−1. This spacing and flow rate delivered approximately 6 L vine−1 h−1, 1.11 mm h−1.
- Extra drip irrigation (SI+)—same irrigation as SI and, in addition, four cycles of 30 min on/30 min off were triggered at night before HS. The treatment consisted of a separated irrigation line with two drippers per vine (flow rate: 13.5 L h−1). The target flow rate was 54 L vine−1 night−1. The system was controlled with a Galcon G.S.I. DC power wireless solenoid controller.
- Sprinkler irrigation (SPRI)—the treatment consisted of the same irrigation as SI, and, in addition, an under-vine broadcast sprinkler covering both the under-vine and inter-row regions. Timing and volume of water were the same as SI+. The system was controlled with a Galcon G.S.I. DC power wireless solenoid controller.
- Sustained deficit irrigation (SDI)—50% of SI from approximatively one week before HS (approximatively two weeks post véraison) to harvest.
2.2. Meteorological Data
2.3. Physiological Measurements
2.4. Hyperspectral Measurements
2.5. Data Analysis
2.5.1. Evaluation of the Treatments
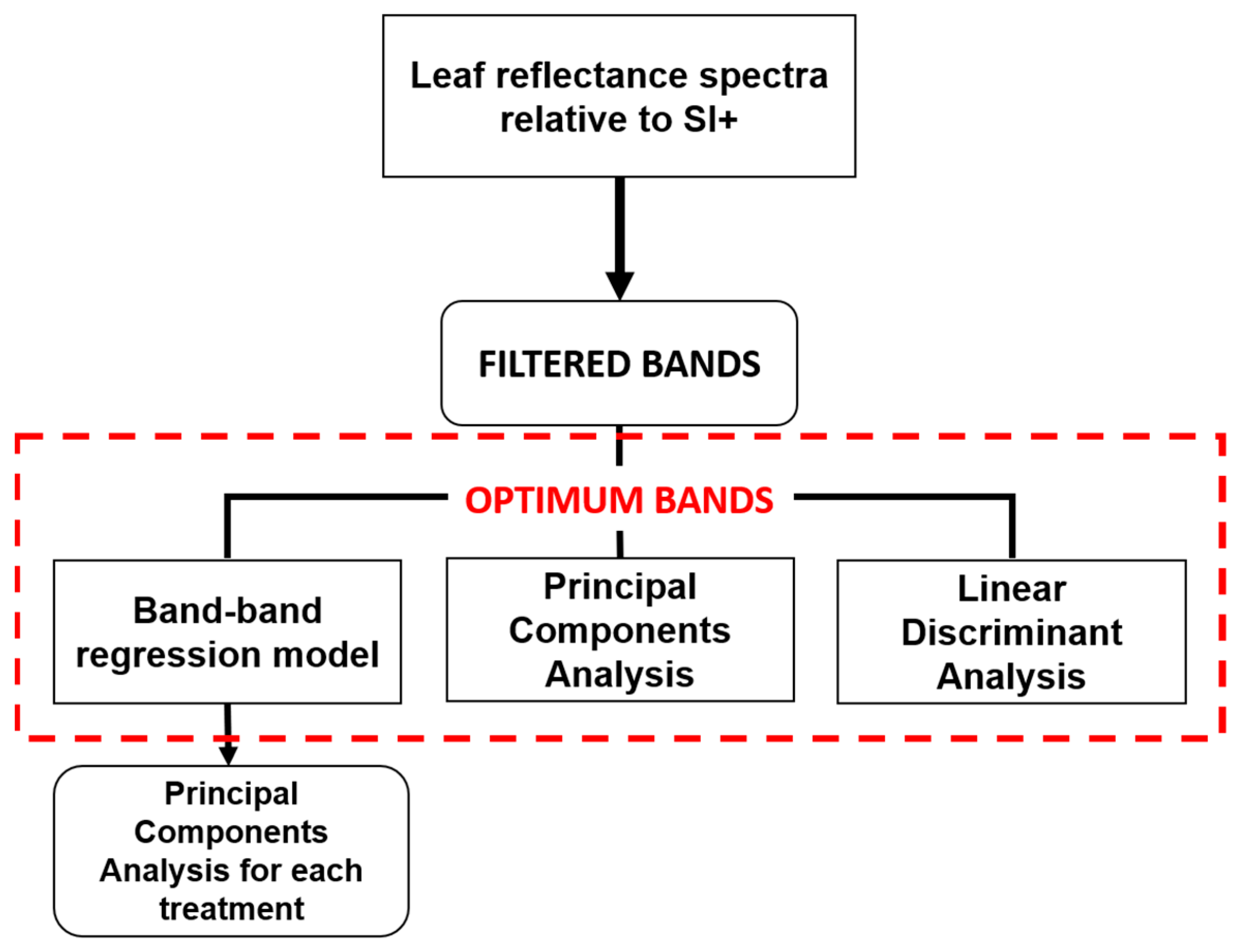
2.5.2. Optimum Hyperspectral Reflectance Bands Selection
3. Results
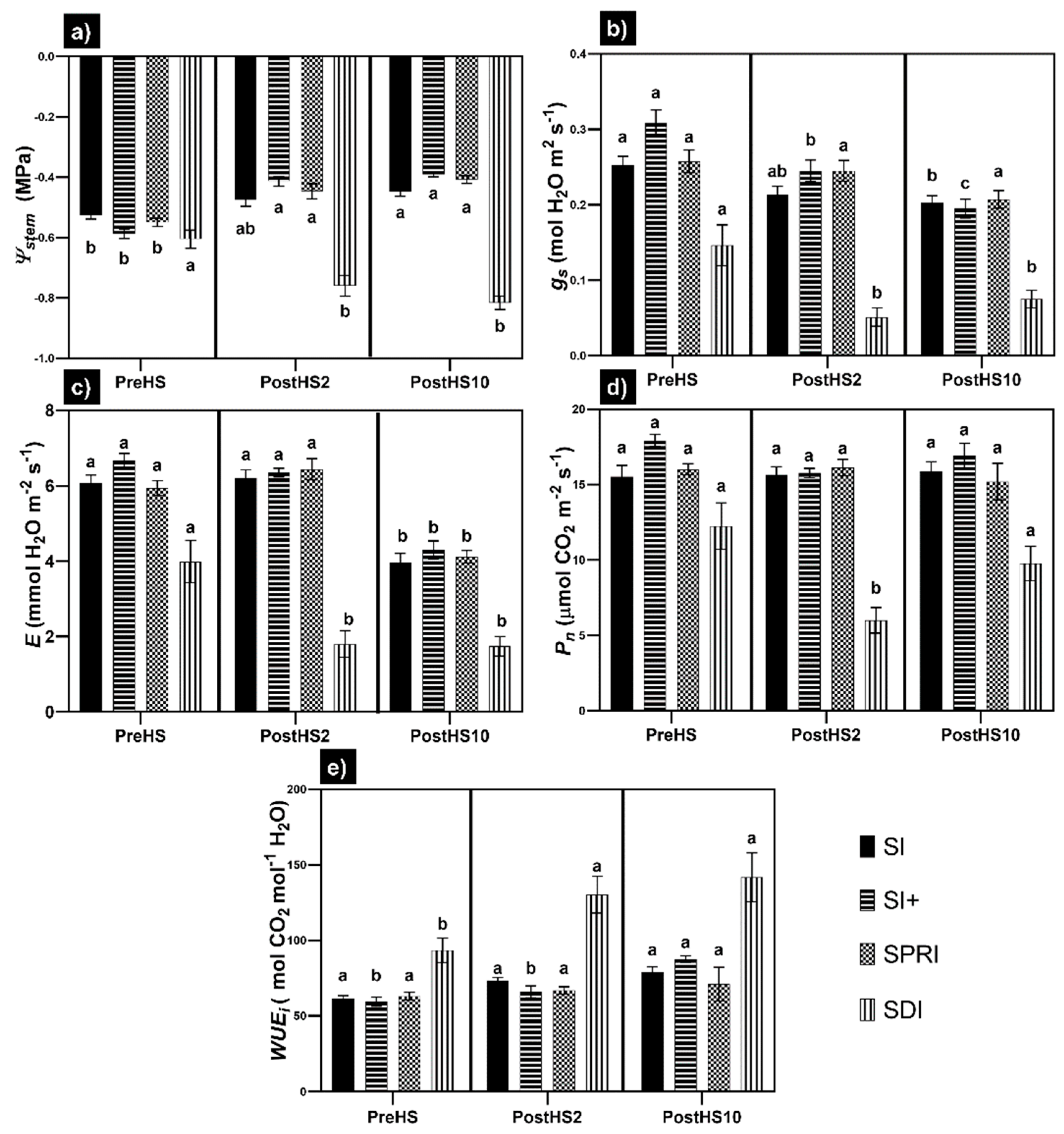
3.1. Grapevine Physiological Status: Water Relations and Gas Exchange
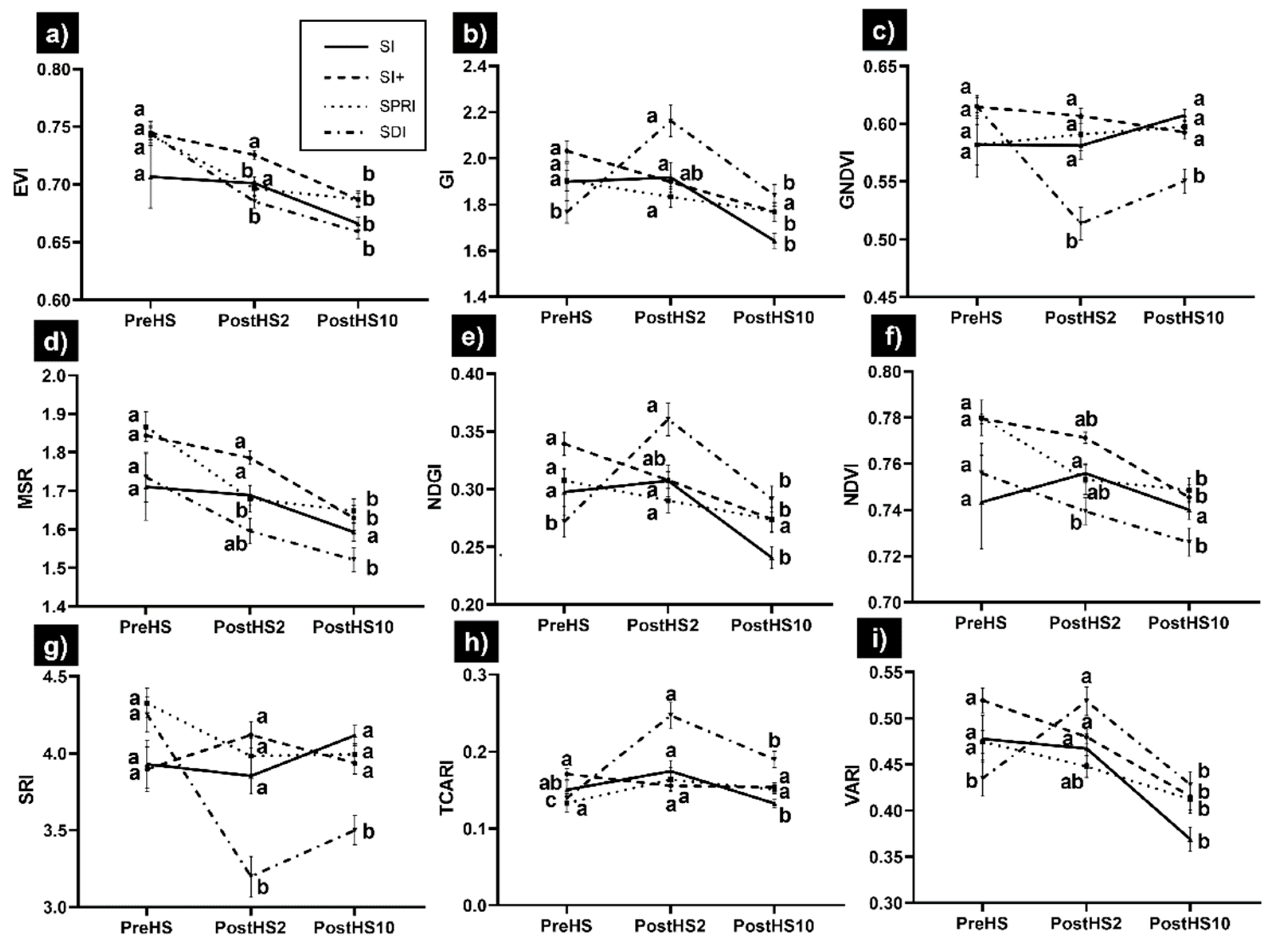
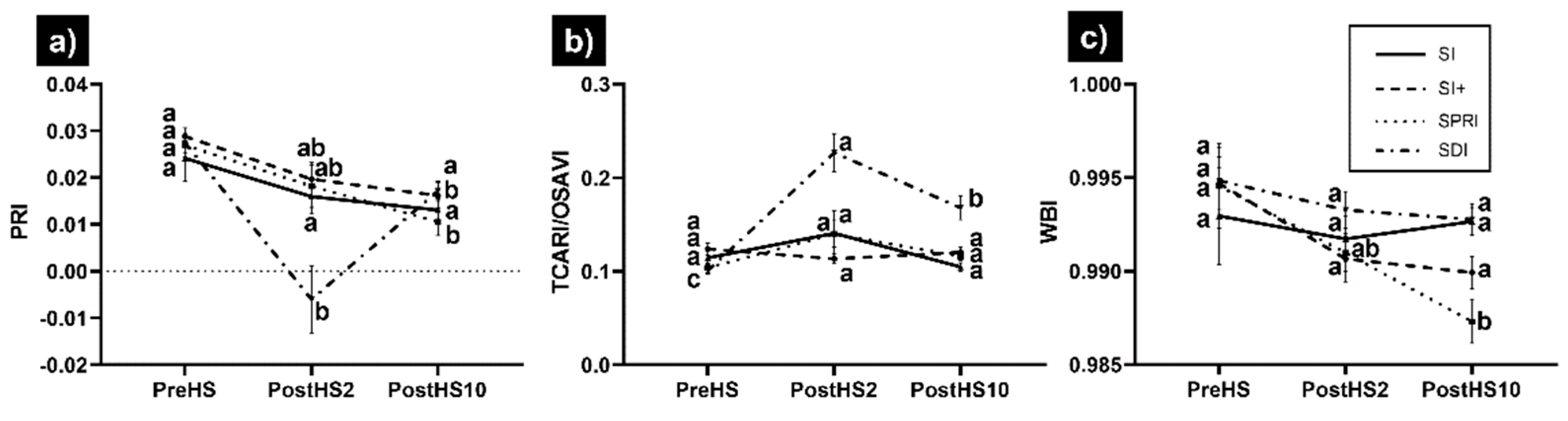
3.2. Hyperspectral-Derived VIs
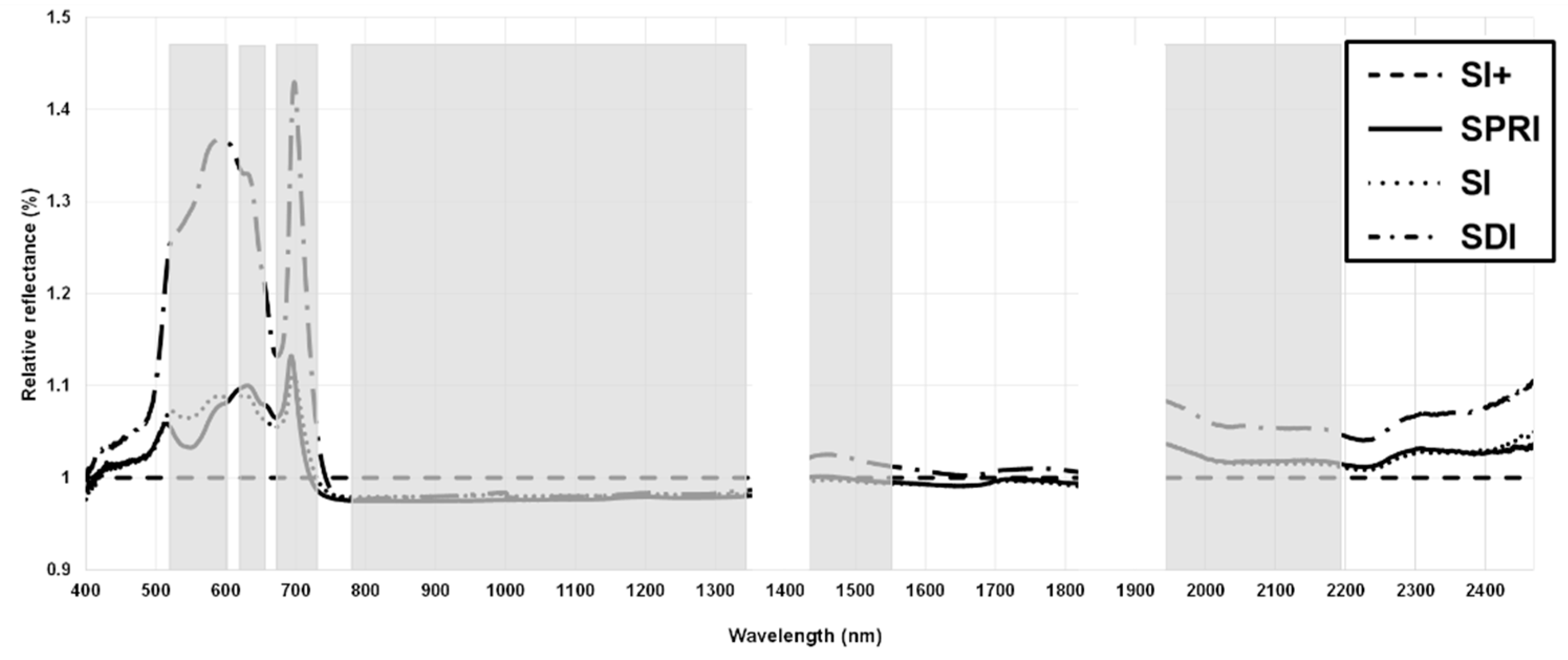
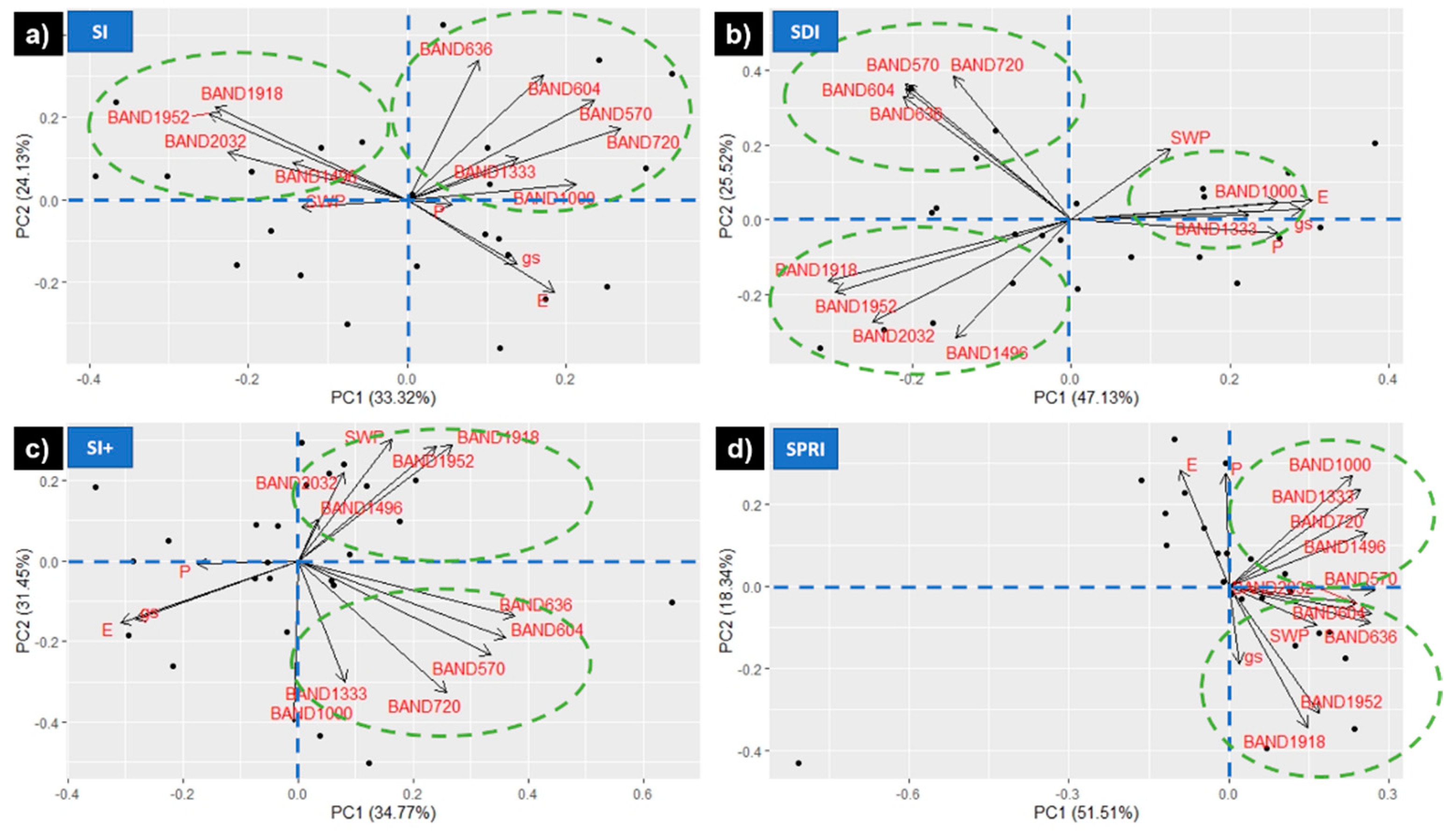
3.3. Optimum Bands Selection
4. Discussion
4.1. Which Cooling Was the Most Effective in Mitigating HS?
4.2. Which Is the Spectral Behavior of the Vines under Combined WS and HS?
5. Conclusions
- Combined HS and WS led to unsatisfactory Ψstem, gs, Pn, E, and WUEi values, which did not recover within 10 days.
- The cooling systems evaluated in the present study were efficacious in mitigating the adverse effects of HS. Specifically, SI+ and SPRI exhibited higher Ψstem after HS. Moreover, in SI+, Pn was not affected by HS in cooled vines, and in SPRI, both Pn and gs were unaffected.
- The spectral VIs showed that SI+, SPRI, and SI were rapidly able to recover the greenness and vigor, as shown by GI, NDGI, and VARI.
- The vine physiological function did not completely recover even 10 days after HS with SRI, TCARI, and TCARI/OSAVI significantly different than their values before HS. The lack of full recovery may indicate that the VIs were sensitive to changes in gs.
- The spectral regions more sensitive to HS were NIR (770–1340 nm), water absorption bands (1941–2200 nm), and the transition region between the green and red bands (600–604 nm), with NIR having the ability to discriminate between SDI and the cooling treatments.
- The single wavebands most sensitive to HS were 604, 720, and 1333–1340 nm.
- The hyperspectral data were consistent with physiological data, identifying SDI as the worst-performing treatment under HS, and SI+ and SPRI as effective cooling strategies to cope with HS.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Arneth, A.; Barbosa, H.; Benton, T.; Calvin, K.; Calvo, E.; Connors, S. Summary for policymakers. In Climate Change and Land: 602 an Ipcc Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems; IPCC: Geneva, Swizerland, 2019. [Google Scholar]
- Perkins-Kirkpatrick, S.; Pitman, A. Extreme events in the context of climate change. Public Health Res. Pract. 2018, 28, 2–5. [Google Scholar] [CrossRef]
- Cogato, A.; Pagay, V.; Marinello, F.; Meggio, F.; Grace, P.; Migliorati, M.D.A. Assessing the feasibility of using Sentinel-2 imagery to quantify the impact of heatwaves on irrigated vineyards. Remote Sens. 2019, 11, 2869. [Google Scholar] [CrossRef]
- Bucur, G.M.; Babes, A.C. Research on trends in extreme weather conditions and their effects on grapevine in Romanian viticulture. Bull. UASVM Hortic. 2016, 73, 126–134. [Google Scholar]
- Duchêne, E.; Huard, F.; Dumas, V.; Schneider, C.; Merdinoglu, D. The challenge of adapting grapevine varieties to climate change. Clim. Res. 2010, 41, 193–204. [Google Scholar] [CrossRef]
- Carvalho, L.; Coito, J.L.; Gonçalves, E.M.F.; Chaves, M.M.; Amâncio, S. Differential physiological response of the grapevine varieties Touriga Nacional and Trincadeira to combined heat, drought and light stresses. Plant Biol. 2015, 18, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Greer, D.H.; Weedon, M.M. The impact of high temperatures on vitis vinifera cv. semillon grapevine performance and berry ripening. Front. Plant Sci. 2013, 4, 1–9. [Google Scholar] [CrossRef]
- Liang, L.; Sun, Q.; Luo, X.; Wang, J.; Zhang, L.; Deng, M.; Di, L.; Liu, Z. Long-term spatial and temporal variations of vegetative drought based on vegetation condition index in China. Ecosphere 2017, 8, e01919. [Google Scholar] [CrossRef]
- Cowan, T.; Purich, A.; Perkins-Kirkpatrick, S.; Pezza, A.; Boschat, G.; Sadler, K. More Frequent, Longer, and Hotter Heat Waves for Australia in the Twenty-First Century. J. Clim. 2014, 27, 5851–5871. [Google Scholar] [CrossRef]
- Schoetter, R.; Cattiaux, J.; Douville, H. Changes of western European heat wave characteristics projected by the CMIP5 ensemble. Clim. Dyn. 2014, 45, 1601–1616. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef]
- Bonada, M.; Sadras, V.; Fuentes, S. Effect of elevated temperature on the onset and rate of mesocarp cell death in berries of Shiraz and Chardonnay and its relationship with berry shrivel. Aust. J. Grape Wine Res. 2013, 19, 87–94. [Google Scholar] [CrossRef]
- Xiao, Z.; Liao, S.; Rogiers, S.; Sadras, V.; Tyerman, S. Effect of water stress and elevated temperature on hypoxia and cell death in the mesocarp of Shiraz berries. Aust. J. Grape Wine Res. 2018, 24, 487–497. [Google Scholar] [CrossRef]
- Sadras, V.; Moran, M.; Bonada, M. Effects of elevated temperature in grapevine. I Berry sensory traits. Aust. J. Grape Wine Res. 2012, 19, 95–106. [Google Scholar] [CrossRef]
- Zhang, P.; Howell, K.; Krstic, M.; Herderich, M.; Barlow, E.W.R.; Fuentes, S. Environmental factors and seasonality affect the concentration of rotundone in Vitis vinifera L. cv. Shiraz wine. PLoS ONE 2015, 10, e0133137. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.A.; Andersen, M.N.; Wollenweber, B.; Kørup, K.; Zhang, X.; Olesen, J.E. Impact of heat-wave at high and low VPD on photosynthetic components of wheat and their recovery. Environ. Exp. Bot. 2018, 147, 138–146. [Google Scholar] [CrossRef]
- Bhusal, N.; Han, S.-G.; Yoon, T.-M. Impact of drought stress on photosynthetic response, leaf water potential, and stem sap flow in two cultivars of bi-leader apple trees (Malus × domestica Borkh.). Sci. Hortic. 2019, 246, 535–543. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Herrera, J.C.; Dayer, S.; Feng, Q.; Hochberg, U.; Castellarin, S.D. The physiology of drought stress in grapevine: Towards an integrative definition of drought tolerance. J. Exp. Bot. 2020, 71, 4658–4676. [Google Scholar] [CrossRef]
- Min, Z.; Li, R.; Chen, L.; Zhang, Y.; Li, Z.; Liu, M.; Ju, Y.; Fang, Y. Alleviation of drought stress in grapevine by foliar-applied strigolactones. Plant Physiol. Biochem. 2019, 135, 99–110. [Google Scholar] [CrossRef]
- Mittler, R. Abiotic stress, the field environment and stress combination. Trends Plant Sci. 2006, 11, 15–19. [Google Scholar] [CrossRef]
- Jing, B.; Shah, F.; Xiao, E.; Coulter, J.A.; Wu, W. Sprinkler irrigation increases grain yield of sunflower without enhancing the risk of root lodging in a dry semi-humid region. Agric. Water Manag. 2020, 239, 106270. [Google Scholar] [CrossRef]
- Gilbert, D.E.; Meyer, J.L.; Kissler, J.J.; La Vine, P.D.; Carlson, C.V. Evaporation cooling of vineyards. Calif. Agric. 1970, 24, 12–14. [Google Scholar] [CrossRef][Green Version]
- Pagay, V.; Tyerman, S.; Jeffery, D.; Muhlack, R.; McCarthy, M.; Boss, P. Using in-Canopy Misters to Mitigate the Negative Effects of Heatwaves in Grapevines; Final Report to Wine Australia; 2018; Available online: https://www.wineaustralia.com/research/projects/using-in-canopy-misters-to-mitigate-the (accessed on 10 September 2021).
- Edwards, E.; Smithson, L.; Graham, D.; Clingeleffer, P. Grapevine canopy response to a high-temperature event during deficit irrigation. Aust. J. Grape Wine Res. 2011, 17, 153–161. [Google Scholar] [CrossRef]
- Sousa, T.A.; Oliveira, M.T.; Moutinho-Pereira, J. Physiological indicators of plant water status of irrigated and non-irrigated grapevines grown in a low rainfall area of portugal. Plant Soil 2006, 282, 127–134. [Google Scholar] [CrossRef]
- Girona, J.; Mata, M.; del Campo, J.; Arbonés, A.; Bartra, E.; Marsal, J. The use of midday leaf water potential for scheduling deficit irrigation in vineyards. Irrig. Sci. 2006, 24, 115–127. [Google Scholar] [CrossRef]
- Cogato, A.; Meggio, F.; Collins, C.; Marinello, F. Medium-resolution multispectral data from Sentinel-2 to assess the damage and the recovery time of late frost on vineyards. Remote Sens. 2020, 12, 1896. [Google Scholar] [CrossRef]
- Poblete, T.; Ortega-Farías, S.; Moreno, M.A.; Bardeen, M. Artificial neural network to predict vine water status spatial variability using multispectral information obtained from an unmanned aerial vehicle (UAV). Sensors 2017, 17, 2488. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Ustin, S.; Whiting, M.L. Temporal and spatial relationships between within-field yield variability in cotton and high-spatial hyperspectral remote sensing imagery. Agron. J. 2005, 97, 641–653. [Google Scholar] [CrossRef]
- Cogato, A.; Pezzuolo, A.; Sørensen, C.G.; De Bei, R.; Sozzi, M.; Marinello, F. A GIS-based multicriteria index to evaluate the mechanisability potential of Italian vineyard area. Land 2020, 9, 469. [Google Scholar] [CrossRef]
- Mirás-Avalos, J.M.; Pérez-Sarmiento, F.; Alcobendas, R.; Alarcón, J.J.; Mounzer, O.; Nicolás, E. Using midday stem water potential for scheduling deficit irrigation in mid–late maturing peach trees under Mediterranean conditions. Irrig. Sci. 2016, 34, 161–173. [Google Scholar] [CrossRef]
- Choné, X.; Van Leeuwen, C.; Dubourdieu, D.; Gaudillère, J.P. Stem water potential is a sensitive indicator of grapevine water status. Ann. Bot. 2001, 87, 477–483. [Google Scholar] [CrossRef]
- Prieto, J.A.; Lebon, É.; Ojeda, H. Stomatal behavior of different grapevine cultivars in response to soil water status and air water vapor pressure deficit. J. Int. Sci. Vigne Vin 2010, 44, 9–20. [Google Scholar] [CrossRef]
- Santesteban, L.G.; Miranda, C.; Royo, J.B. Effect of water deficit and rewatering on leaf gas exchange and transpiration decline of excised leaves of four grapevine (Vitis vinifera L.) cultivars. Sci. Hortic. 2009, 121, 434–439. [Google Scholar] [CrossRef]
- Tomás, M.; Medrano, H.; Escalona, J.M.; Martorell, S.; Pou, A.; Ribas-Carbo, M.; Flexas, J. Variability of water use efficiency in grapevines. Environ. Exp. Bot. 2014, 103, 148–157. [Google Scholar] [CrossRef]
- Greer, D.H.; Weston, C. Heat stress affects flowering, berry growth, sugar accumulation and photosynthesis of Vitis vinifera cv. Semillon grapevines grown in a controlled environment. Funct. Plant Biol. 2010, 37, 206–214. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Berjón, A.; López-Lozano, R.; Miller, J.R.; Martín, P.; Cachorro, V.; González, M.R.; De Frutos, A. Assessing vineyard condition with hyperspectral indices: Leaf and canopy reflectance simulation in a row-structured discontinuous canopy. Remote Sens. Environ. 2005, 99, 271–287. [Google Scholar] [CrossRef]
- Rouse, J.; Haas, R.; Schell, J.; Deering, D.; Harlan, J. Monitoring the Vernal Advancement and Retrogradation (Greenwave Effect) of Natural Vegetation; Type III Final Report; NASA/GSFC: Greenbelt, MD, USA, 1974; p. 371.
- Acevedo-Opazo, C.; Tisseyre, B.; Guillaume, S.; Ojeda, H. Test of NDVI information for a relevant vineyard zoning related to vine water status. In Proceedings of the VI European Conference on Precision Agriculture (ECPA), Skiathos, Greece, 3–6 June 2007; pp. 547–554. [Google Scholar]
- Acevedo-Opazo, C.; Tisseyre, B.; Guillaume, S.; Ojeda, H. The potential of high spatial resolution information to define within-vineyard zones related to vine water status. Precis. Agric. 2008, 9, 285–302. [Google Scholar] [CrossRef]
- Baluja, J.; Diago, M.P.; Balda, P.; Zorer, R.; Meggio, F.; Morales, F.; Tardaguila, J. Assessment of vineyard water status variability by thermal and multispectral imagery using an unmanned aerial vehicle (UAV). Irrig. Sci. 2012, 30, 511–522. [Google Scholar] [CrossRef]
- Espinoza, C.Z.; Khot, L.R.; Sankaran, S.; Jacoby, P.W. High resolution multispectral and thermal remote sensing-based water stress assessment in subsurface irrigated grapevines. Remote Sens. 2017, 9, 961. [Google Scholar] [CrossRef]
- Serrano, L.; González-Flor, C.; Gorchs, G. Assessing vineyard water status using the reflectance based Water Index. Agric. Ecosyst. Environ. 2010, 139, 490–499. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Gonzalez-Dugo, V.; Williams, L.; Suárez, L.; Jimenez-Berni, J.A.; Goldhamer, D.; Fereres, E. A PRI-based water stress index combining structural and chlorophyll effects: Assessment using diurnal narrow-band airborne imagery and the CWSI thermal index. Remote Sens. Environ. 2013, 138, 38–50. [Google Scholar] [CrossRef]
- Di Gennaro, S.F.; Matese, A.; Gioli, B.; Toscano, P.; Zaldei, A.; Palliotti, A.; Genesio, L. Multisensor approach to assess vineyard thermal dynamics combining high-resolution unmanned aerial vehicle (UAV) remote sensing and wireless sensor network (WSN) proximal sensing. Sci. Hortic. 2017, 221, 83–87. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Viña, A.; Ciganda, V.; Rundquist, D.C.; Arkebauer, T.J. Remote estimation of canopy chlorophyll content in crops. Geophys. Res. Lett. 2005, 32, 1–4. [Google Scholar] [CrossRef]
- Chen, J.M. Evaluation of vegetation indices and a modified simple ratio for boreal applications. Can. J. Remote Sens. 1996, 22, 229–242. [Google Scholar] [CrossRef]
- Haboudane, D.; Miller, J.R.; Tremblay, N.; Zarco-Tejada, P.J.; Dextraze, L. Integrated narrow-band vegetation indices for prediction of crop chlorophyll content for application to precision agriculture. Remote Sens. Environ. 2002, 81, 416–426. [Google Scholar] [CrossRef]
- Courel, M.-F.; Chamard, P.; Guenegou, M.J.; Lerhun, J.; Levasseur, M.; Togola, M. Utilisation des bandes spectrales du vert et du rouge pour une meilleure évaluation des formations végétales actives. In Proceedings of the Congrès AUPELF-UREF, Sherbrooke, QC, Canada, 21–23 October 1991; pp. 203–210. [Google Scholar]
- Jordan, C.F. Derivation of leaf-area index from quality of light on the forest floor. Ecology 1969, 50, 663–666. [Google Scholar] [CrossRef]
- Gitelson, A.A.; Kaufman, Y.J.; Stark, R.; Rundquist, D. Novel algorithms for remote estimation of vegetation fraction. Remote Sens. Environ. 2002, 80, 76–87. [Google Scholar] [CrossRef]
- Pôças, I.; Rodrigues, A.; Gonçalves, S.; Costa, P.M.; Gonçalves, I.; Pereira, L.S.; Cunha, M. Predicting grapevine water status based on hyperspectral reflectance vegetation indices. Remote Sens. 2015, 7, 16460–16479. [Google Scholar] [CrossRef]
- Gamon, J.A.; Peñuelas, J.; Field, C.B. A narrow-waveband spectral index that tracks diurnal changes in photosynthetic efficiency. Remote Sens. Environ. 1992, 41, 35–44. [Google Scholar] [CrossRef]
- Liu, H.Q.; Huete, A. A feedback based modification of the NDVI to minimize canopy background and atmospheric noise. IEEE Trans. Geosci. Remote Sens. 1995, 33, 457–465. [Google Scholar] [CrossRef]
- Cheng, Y.-B.; Zarco-Tejada, P.J.; Riano, D.; Rueda, C.A.; Ustin, S. Estimating vegetation water content with hyperspectral data for different canopy scenarios: Relationships between AVIRIS and MODIS indexes. Remote Sens. Environ. 2006, 105, 354–366. [Google Scholar] [CrossRef]
- Dold, C.; Heitman, J.; Giese, G.; Howard, A.; Havlin, J.; Sauer, T. Upscaling Evapotranspiration with parsimonious models in a North Carolina vineyard. Agronomy 2019, 9, 152. [Google Scholar] [CrossRef]
- Penuelas, J.; Filella, I.; Biel, C.; Serrano, L.; Savé, R. The reflectance at the 950–970 nm region as an indicator of plant water status. Int. J. Remote Sens. 1993, 14, 1887–1905. [Google Scholar] [CrossRef]
- Fórián, T.; Nagy, A.; Riczu, P.; Mézes, L.; Tamás, J. Vineyards characteristic by using GIS and refl ectance measurements on the Nagy-Eged hill in Hungary. Int. J. Hortic. Sci. 2016, 18, 57–60. [Google Scholar] [CrossRef]
- Thenkabail, P.S.; Enclona, E.A.; Ashton, M.S.; Van Der Meer, B. Accuracy assessments of hyperspectral waveband performance for vegetation analysis applications. Remote Sens. Environ. 2004, 91, 354–376. [Google Scholar] [CrossRef]
- Ray, S.S.; Singh, J.P.; Panigraphy, S. Use of hyperstectralremote senings data for crop stress detection: Ground-based studies. Int. Arch. Photogramm. Remote. Sens. Spat. Inf. Sci. 2010, 38, 562–570. [Google Scholar]
- Mutanga, O.; Skidmore, A.; Prins, H. Predicting in situ pasture quality in the Kruger National Park, South Africa, using continuum-removed absorption features. Remote Sens. Environ. 2004, 89, 393–408. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; van Leeuwen, W.J.D. The use of vegetation indices in forested regions: Issues of linearity and saturation. Int. Geosci. Remote Sens. Symp. 1997, 4, 1966–1968. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses (Versión 1.0.5). Available online: https://cran.r-project.org/package=factoextra (accessed on 1 March 2021).
- Ruiz, E.; Jackson, S.; Cimentada, J. Corrr: Correlations in R. Available online: https://cran.r-project.org/web/packages/corrr/index.htm (accessed on 1 March 2021).
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Yang, J.; Yang, J.Y. Why can LDA be performed in PCA transformed space? Pattern Recognit. 2003, 36, 563–566. [Google Scholar] [CrossRef]
- Roever, C.; Raabe, N.; Luebke, K.; Ligges, U.; Szepannek, G.; Zentgraf, M. klaR: Classification and visualization. Available online: https://cran.r-project.org/package=klaR (accessed on 1 March 2021).
- Greer, D.H.; Weedon, M. Modelling photosynthetic responses to temperature of grapevine (Vitis vinifera cv. Semillon) leaves on vines grown in a hot climate. Plant Cell Environ. 2011, 35, 1050–1064. [Google Scholar] [CrossRef]
- Zsófi, Z.; Gál, L.; Szilágyi, Z.; Szűcs, E.; Marschall, M.; Nagy, Z.; Bálo, B. Use of stomatal conductance and pre-dawn water potential to classify terroir for the grape variety Kékfrankos. Aust. J. Grape Wine Res. 2009, 15, 36–47. [Google Scholar] [CrossRef]
- Pagay, V.; Canela, F.; Bennet, C. How Does Phenological Stage Influence Grapevine Water Requirements for Shiraz and Chardonnay in the Riverland? Final Report to Wine Australia; 2021; Available online: https://www.growag.com/listings/research-project/incubator-initiative-how-does-phenological-stage-influence-grapevine-water-requirements-for-shiraz-and-chardonnay-in-the-riverland (accessed on 10 September 2021).
- Acevedo-Opazo, C.; Ortega-Farias, S.; Fuentes, S. Effects of grapevine (Vitis vinifera L.) water status on water consumption, vegetative growth and grape quality: An irrigation scheduling application to achieve regulated deficit irrigation. Agric. Water Manag. 2010, 97, 956–964. [Google Scholar] [CrossRef]
- Pierantozzi, P.; Torres, M.; Bodoira, R.; Maestri, D. Water relations, biochemical–physiological and yield responses of olive trees (Olea europaea L. cvs. Arbequina and Manzanilla) under drought stress during the pre-flowering and flowering period. Agric. Water Manag. 2013, 125, 13–25. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Lee, H.; Adhikari, A.; Han, A.R.; Kim, H.S. Evaluation of morphological, physiological, and biochemical traits for assessing drought resistance in eleven tree species. Sci. Total. Environ. 2021, 779, 146466. [Google Scholar] [CrossRef] [PubMed]
- Brito, C.; Dinis, L.-T.; Moutinho-Pereira, J.; Correia, C.M.; Pereira, M. Drought stress effects and olive tree acclimation under a changing climate. Plants 2019, 8, 232. [Google Scholar] [CrossRef]
- Patakas, A.; Noitsakis, B.; Chouzouri, A. Optimization of irrigation water use in grapevines using the relationship between transpiration and plant water status. Agric. Ecosyst. Environ. 2005, 106, 253–259. [Google Scholar] [CrossRef]
- Patakas, A.; Noitsakis, B.; Stavrakas, D. Adaptation of leaves of Vitis vinifera L. to seasonal drought as affected by leaf age. Vitis 1997, 36, 11–14. [Google Scholar]
- Rapaport, T.; Hochberg, U.; Shoshany, M.; Karnieli, A.; Rachmilevitch, S. Combining leaf physiology, hyperspectral imaging and partial least squares-regression (PLS-R) for grapevine water status assessment. ISPRS J. Photogramm. Remote Sens. 2015, 109, 88–97. [Google Scholar] [CrossRef]
- Palliotti, A.; Poni, S. Grapevine under light and heat stresses. In Grapevine in a Changing Environment; Wiley: Hoboken, NJ, USA, 2015; pp. 148–178. [Google Scholar]
- Urban, J.; Ingwers, M.W.; McGuire, M.A.; Teskey, R.O. Increase in leaf temperature opens stomata and decouples net photosynthesis from stomatal conductance in Pinus taeda and Populus deltoides x nigra. J. Exp. Bot. 2017, 68, 1757–1767. [Google Scholar] [CrossRef]
- Flexas, J.; Galmés, J.; Gallé, A.; Gulías, J.; Pou, A.; Ribas-Carbo, M.; Tomàs, M.; Medrano, H. Improving water use efficiency in grapevines: Potential physiological targets for biotechnological improvement. Aust. J. Grape Wine Res. 2010, 16, 106–121. [Google Scholar] [CrossRef]
- Bchir, A.; Escalona, J.M.; Gallé, A.; Hernández-Montes, E.; Tortosa, I.; Braham, M.; Medrano, H. Carbon isotope discrimination (δ13C) as an indicator of vine water status and water use efficiency (WUE): Looking for the most representative sample and sampling time. Agric. Water Manag. 2016, 167, 11–20. [Google Scholar] [CrossRef]
- Luo, H.-B.; Ma, L.; Xi, H.-F.; Duan, W.; Li, S.-H.; Loescher, W.; Wang, J.-F.; Wang, L.-J. Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (Vitis amurensis L.) leaves. PLoS ONE 2011, 6, e23033. [Google Scholar] [CrossRef]
- Bauer, H. Photosynthesis of Ivy Leaves (Hedera helix) after Heat Stress I. CO2-Gas Exchange and diffusion resistances. Physiol. Plant 1978, 44, 400–406. [Google Scholar] [CrossRef]
- Peñuelas, J.; Filella, L. Visible and near-infrared reflectance techniques for diagnosing plant physiological status. Trends Plant Sci. 1998, 3, 151–156. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Ismail, M.R.; Malek, M.A.; Latif, M.A.; Oladosu, Y. Heat shock proteins: Functions and response against heat stress in plants. Int. J. Sci. Technol. Res. 2014, 3, 204–218. [Google Scholar]
- Wang, L.-J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.-J.; Cheng, J.-S.; Luo, H.-B.; Li, S.-H. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef]
- Zarco-Tejada, P.J.; Jimenez-Berni, J.A.; Suárez, L.; Sepulcre-Cantó, G.; Morales, F.; Miller, J.R. Imaging chlorophyll fluorescence with an airborne narrow-band multispectral camera for vegetation stress detection. Remote Sens. Environ. 2009, 113, 1262–1275. [Google Scholar] [CrossRef]
- Perry, E.M.; Roberts, D.A. Sensitivity of narrow-band and broad-band indices for assessing nitrogen availability and water stress in an annual crop. Agron. J. 2008, 100, 1211–1219. [Google Scholar] [CrossRef]
- Romero, M.; Luo, Y.; Su, B.; Fuentes, S. Vineyard water status estimation using multispectral imagery from an UAV platform and machine learning algorithms for irrigation scheduling management. Comput. Electron. Agric. 2018, 147, 109–117. [Google Scholar] [CrossRef]
- Xiao, F.; Yang, Z.Q.; Lee, K.W. Photosynthetic and physiological responses to high temperature in grapevine (Vitis vinifera L.) leaves during the seedling stage. J. Hortic. Sci. Biotechnol. 2016, 92, 2–10. [Google Scholar] [CrossRef]
- Dobrowski, S.; Pushnik, J.; Zarco-Tejada, P.J.; Ustin, S. Simple reflectance indices track heat and water stress-induced changes in steady-state chlorophyll fluorescence at the canopy scale. Remote Sens. Environ. 2005, 97, 403–414. [Google Scholar] [CrossRef]
- Sonmez, N.K.; Emekli, Y.; Sarı, M.; Bastug, R.; Sari, M. Relationship between spectral reflectance and water stress conditions of Bermuda grass (Cynodon dactylon L.). N. Z. J. Agric. Res. 2008, 51, 223–233. [Google Scholar] [CrossRef]
- Houborg, R.; Soegaard, H.; Boegh, E. Combining vegetation index and model inversion methods for the extraction of key vegetation biophysical parameters using Terra and Aqua MODIS reflectance data. Remote Sens. Environ. 2007, 106, 39–58. [Google Scholar] [CrossRef]
- Rodríguez-Pérez, J.R.; Riaño, D.; Carlisle, E.; Ustin, S.; Smart, D.R. Evaluation of hyperspectral reflectance indexes to detect grapevine water status in vineyards. Am. J. Enol. Vitic. 2007, 58, 302–317. [Google Scholar]
- De Jong, S.; Addink, E.; Hoogenboom, P.; Nijland, W. The spectral response of Buxus sempervirens to different types of environmental stress—A laboratory experiment. ISPRS J. Photogramm. Remote Sens. 2012, 74, 56–65. [Google Scholar] [CrossRef]
- Wang, J.; Xu, R.; Yang, S. Estimation of plant water content by spectral absorption features centered at 1450 nm and 1940 nm regions. Environ. Monit. Assess. 2008, 157, 459–469. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Formula | Stress | Cultivar | Reference |
|---|---|---|---|---|
| Normalized Difference Vegetation Index [38] | NDVI = | WS | Vitis vinifera L. cv. Muscat, Carignan, Grenache Noir, Shiraz, Mourvedre, Petit Verdot | [39,40] |
| WS | Vitis vinifera L. cv. Tempranillo | [41] | ||
| WS | Vitis vinifera L. cv. Cabernet Sauvignon | [42] | ||
| WS | Vitis vinifera L. cv. Carménère | [28] | ||
| WS | Vitis vinifera L. cv. Chardonnay | [43] | ||
| WS | Vitis vinifera L. cv. Thompson Seedless | [44] | ||
| HS | Vitis vinifera L. cv. Sangiovese | [45] | ||
| Green Normalized Difference Vegetation Index [46] | GNDVI = | WS | Vitis vinifera L. cv. Carménère | [28] |
| WS | Vitis vinifera L. cv. Tempranillo | [41] | ||
| WS | Vitis vinifera L. cv. Cabernet Sauvignon | [42] | ||
| Modified Simple Ratio [47] | MSR = | WS | Vitis vinifera L. cv. Tempranillo | [41] |
| WS | Vitis vinifera L. cv. Carménère | [28] | ||
| Transformed Chlorophyll Absorption Ratio Index [48] | TCARI = ) | WS | Vitis vinifera L. cv. Tempranillo | [41] |
| HS | Vitis vinifera L. several cultivars | [49] | ||
| TCARI/Optimized Soil-Adjusted Vegetation Index [48] | TCARI/OSAVI = | WS | Vitis vinifera L. cv. Tempranillo | [41] |
| WS | Vitis vinifera L. cv. Thompson Seedless | [44] | ||
| Green Index [49] | GI = | WS | Vitis vinifera L. cv. Tempranillo | [41] |
| Simple Ratio Index 800/550 [50] | SRI = | WS | Vitis vinifera L. cv. Tempranillo | [41] |
| Visible Atmospherically Resistant Index [51] | VARI = | WS | Vitis vinifera L. cv. Touriga Nacional | [52] |
| Normalized Difference Greenness Vegetation Index [49] | NDGI = | WS | Vitis vinifera L. cv. Touriga Nacional | [52] |
| Photochemical Reflectance Index [53] | PRI = | WS | Vitis vinifera L. cv. Thompson Seedless | [44] |
| Enhanced Vegetation Index [54] | EVI = 2.5 ∗ | HS | Vitis vinifera L., several cultivars | [3] |
| WS | Several crops (e.g., cotton, creosote bush, spruce) | [55] | ||
| WS | Vitis vinifera L. cv. Chardonnay | [56] | ||
| Water Band Index [57] | WBI = | WS | Vitis vinifera L., several cultivars | [58] |
| Date | Avg Daily T a (°C) | Max Daily T a (°C) | Avg Daily RH a (%) | Min Daily RH a (%) | Avg Daily Soil T a,b (°C) | Max Daily Soil T a,b (°C) | Max Daily VPD a (kPa) | Avg SR a (W m−2) | Max SR a (W m−2) |
|---|---|---|---|---|---|---|---|---|---|
| 14 January 2020 | 26.2 | 35.4 | 38.1 | 12.6 | 43.5 | 80.6 | 2.1 | 371 | 1078 |
| 15 January 2020 | 27.0 | 39.0 | 47.3 | 12.6 | 49.6 | 89.9 | 1.9 | 339 | 1045 |
| 16 January 2020 | 21.0 | 33.3 | 54.9 | 0.0 | 46.2 | 80.6 | 1.12 | 356 | 1090 |
| Bands | R2 |
|---|---|
| 570~2032 | 0.182 |
| 636~1000 | 0.184 |
| 720~1948 | 0.082 |
| 1333~1952 | 0.001 |
| 1496~604 | 0.203 |
| Bands with Highest Factor Loading | Variability Explained (%) | ||||
|---|---|---|---|---|---|
| PC1 | PC2 | PC3 | PC1 | PC2 | PC3 |
| 1340, 1339, 1338, 1337, 1336 | 1969, 1970, 1968, 1967, 1971 | 604, 603, 601, 600, 602 | 62.62 | 27.00 | 7.88 |
| 1340, 1550, 2199, 720 | 1969, 1440, 883, 610 | 604, 1924, 2010, 1338 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cogato, A.; Wu, L.; Jewan, S.Y.Y.; Meggio, F.; Marinello, F.; Sozzi, M.; Pagay, V. Evaluating the Spectral and Physiological Responses of Grapevines (Vitis vinifera L.) to Heat and Water Stresses under Different Vineyard Cooling and Irrigation Strategies. Agronomy 2021, 11, 1940. https://doi.org/10.3390/agronomy11101940
Cogato A, Wu L, Jewan SYY, Meggio F, Marinello F, Sozzi M, Pagay V. Evaluating the Spectral and Physiological Responses of Grapevines (Vitis vinifera L.) to Heat and Water Stresses under Different Vineyard Cooling and Irrigation Strategies. Agronomy. 2021; 11(10):1940. https://doi.org/10.3390/agronomy11101940
Chicago/Turabian StyleCogato, Alessia, Lihua Wu, Shaikh Yassir Yousouf Jewan, Franco Meggio, Francesco Marinello, Marco Sozzi, and Vinay Pagay. 2021. "Evaluating the Spectral and Physiological Responses of Grapevines (Vitis vinifera L.) to Heat and Water Stresses under Different Vineyard Cooling and Irrigation Strategies" Agronomy 11, no. 10: 1940. https://doi.org/10.3390/agronomy11101940
APA StyleCogato, A., Wu, L., Jewan, S. Y. Y., Meggio, F., Marinello, F., Sozzi, M., & Pagay, V. (2021). Evaluating the Spectral and Physiological Responses of Grapevines (Vitis vinifera L.) to Heat and Water Stresses under Different Vineyard Cooling and Irrigation Strategies. Agronomy, 11(10), 1940. https://doi.org/10.3390/agronomy11101940