Anti-Inflammatory and Antioxidant Effects of Anthocyanins of Trifolium pratense (Red Clover) in Lipopolysaccharide-Stimulated RAW-267.4 Macrophages



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Anthocyanin Fractions from Red Clover
2.2. Cell Culture
2.3. Measurement of Intracellular ROS
2.4. Quantitative Polymerase Chain Reaction (qPCR) Analysis
2.5. Enzyme-Linked Immunosorbent Assay (ELISA) for TNFα
2.6. Western Blot Analysis
2.7. Identification and Quantification of Red Clover Polyphenols Using HPLC/ESI-MS Analysis
2.8. Statistical Analysis
3. Results
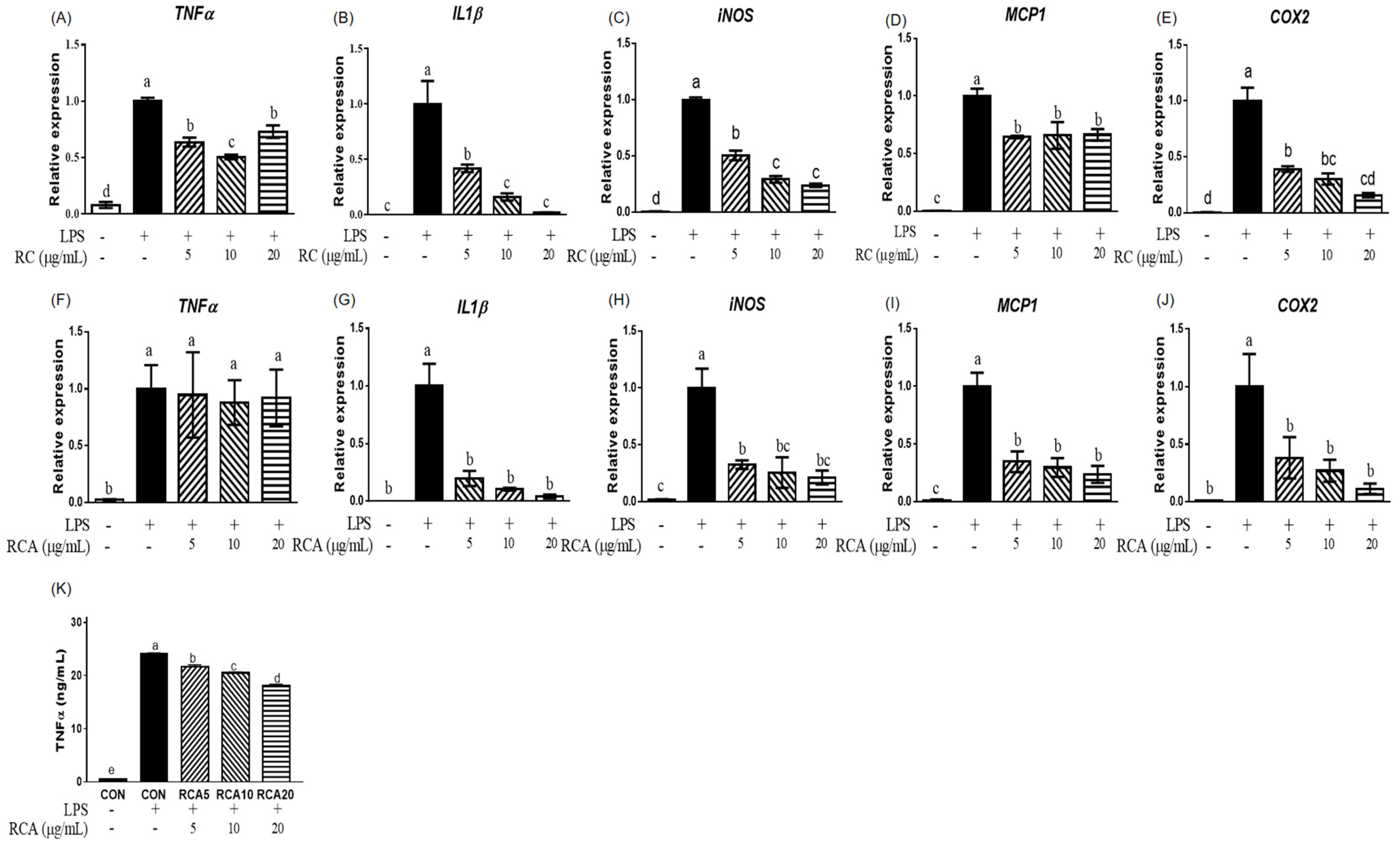
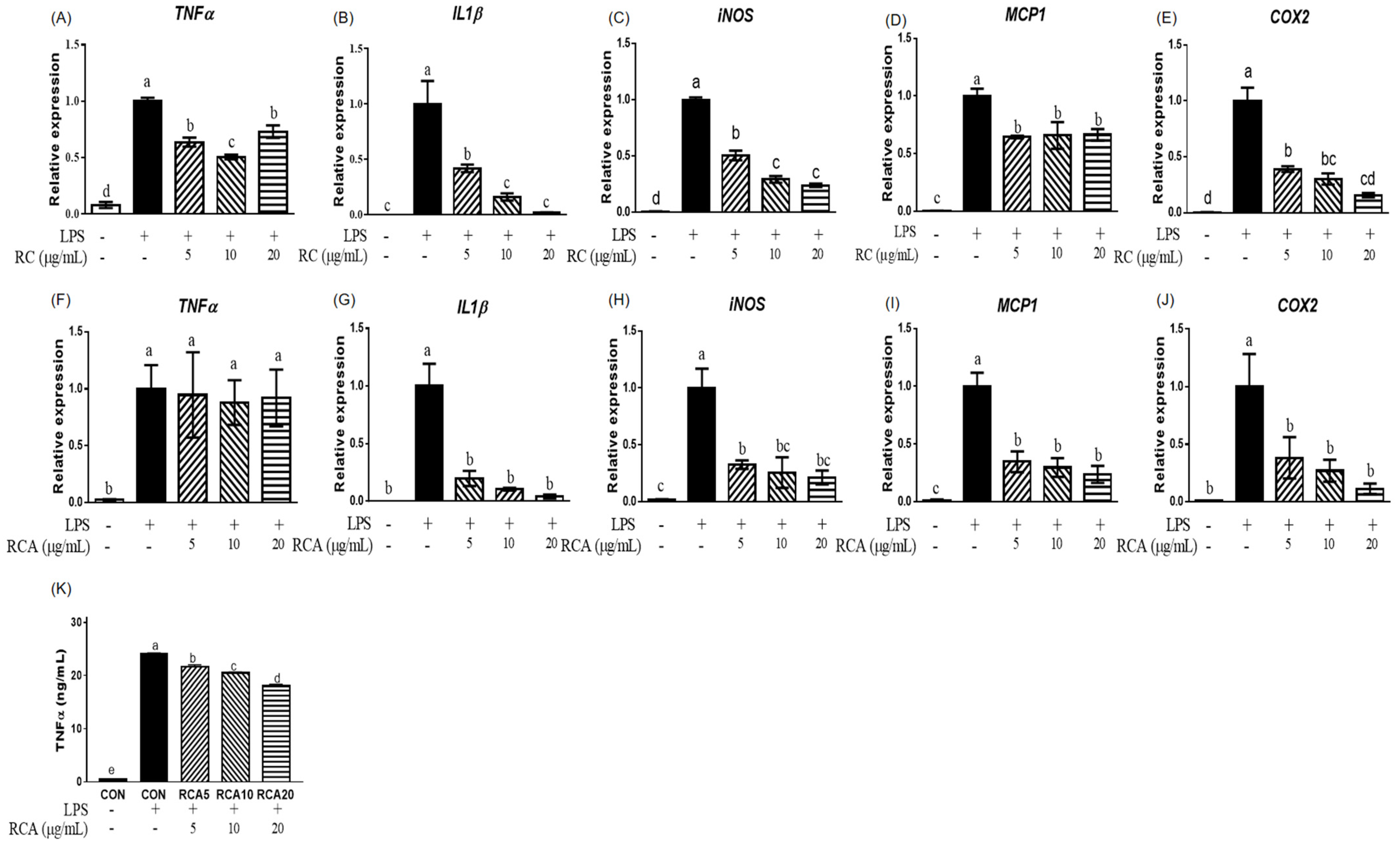
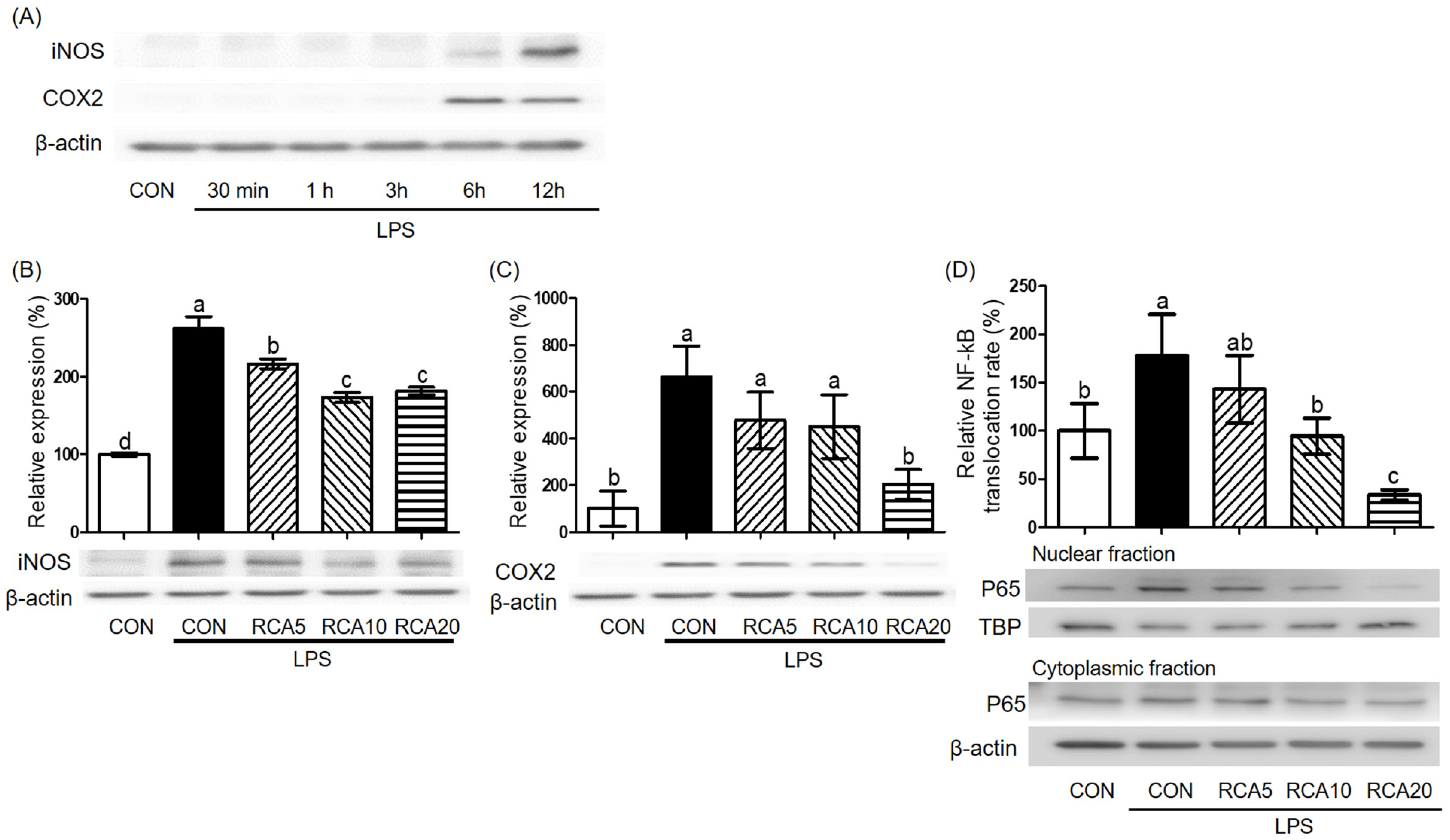
3.1. RC and RCA Decreased the Expression of Genes Related to Pro-Inflammatory Markers
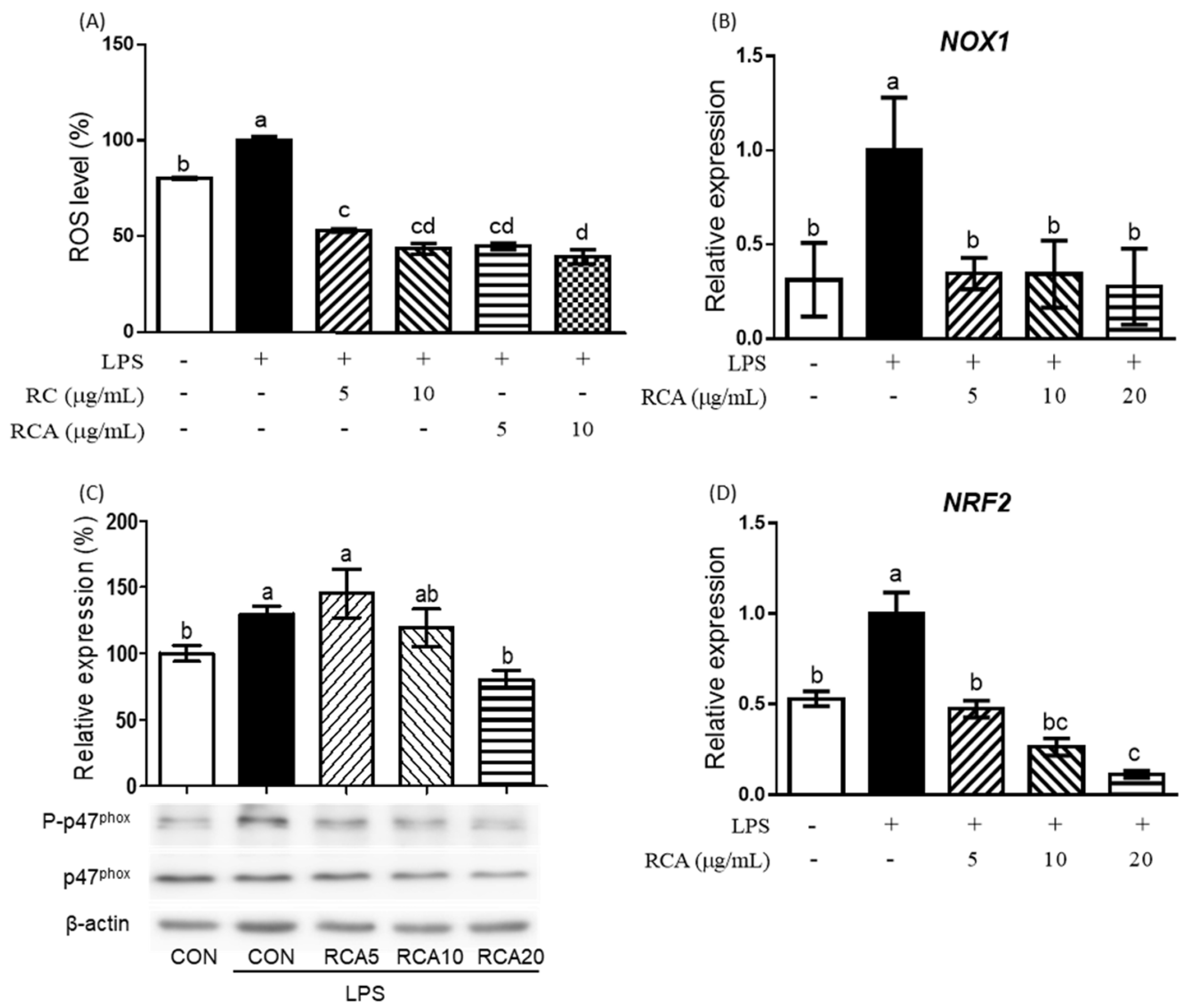
3.2. RC and RCA Inhibited LPS-Induced ROS Production
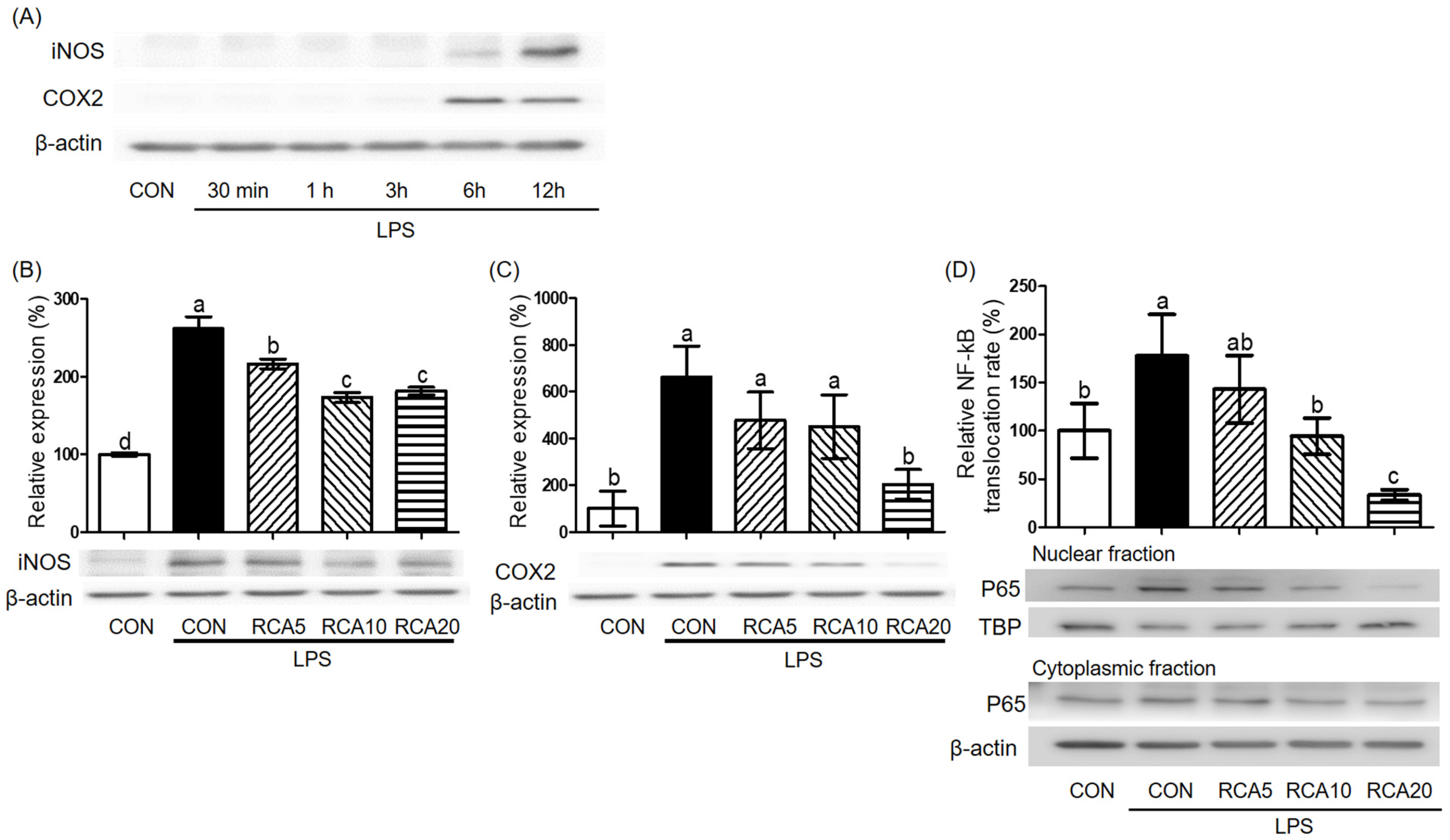
3.3. RCA Inhibited the Activation of NF-kB in LPS-Induced RAW 264.7 Cells
3.4. Identification and Quantification of Red Clover Polyphenols
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Coates, P.M.; Blackman, M.; Betz, J.; Cragg, G.M.; Levine, M.; Moss, J.; White, J.D. Encyclopedia of Dietary Supplements; CRC Press: New York, NY, USA, 2010; pp. 665–679. [Google Scholar]
- Booth, N.L.; Piersen, C.E.; Banuvar, S.; Geller, S.E.; Shulman, L.P.; Farnsworth, N.R. Clinical studies of red clover (Trifolium pratense) dietary supplements in menopause: A literature review. Menopause 2006, 13, 251–264. [Google Scholar] [CrossRef] [PubMed]
- Van de Weijer, P.H.; Barentsen, R. Isoflavones from red clover (Promensil) significantly reduce menopausal hot flush symptoms compared with placebo. Maturitas 2002, 42, 187–193. [Google Scholar] [CrossRef]
- Lipovac, M.; Chedraui, P.; Gruenhut, C.; Gocan, A.; Stammler, M.; Imhof, M. Improvement of postmenopausal depressive and anxiety symptoms after treatment with isoflavones derived from red clover extracts. Maturitas 2010, 65, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, L.A.; Chedraui, P.A.; Morocho, N.; Ross, S.; San Miguel, G. The effect of red clover isoflavones on menopausal symptoms, lipids and vaginal cytology in menopausal women: A randomized, double-blind, placebo-controlled study. Gynecol. Endocrinol. 2005, 21, 257–264. [Google Scholar] [CrossRef]
- Castaneda-Ovando, A.; de Lourdes Pacheco-Hernández, M.; Páez-Hernández, M.E.; RodrÃguez, J.A.; Galán-Vidal, C.A. Chemical studies of anthocyanins: A review. Food Chem. 2009, 113, 859–871. [Google Scholar] [CrossRef]
- Yousuf, B.; Gul, K.; Wani, A.A.; Singh, P. Health benefits of anthocyanins and their encapsulation for potential use in food systems: A review. Crit. Rev. Food Sci. Nutr. 2016, 56, 2223–2230. [Google Scholar] [CrossRef]
- Zhu, Y.; Ling, W.; Guo, H.; Song, F.; Ye, Q.; Zou, T.; Li, D.; Zhang, Y.; Li, G.; Xiao, Y.; et al. Anti-inflammatory effect of purified dietary anthocyanin in adults with hypercholesterolemia: A randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 843–849. [Google Scholar] [CrossRef]
- Selvaraju, V.; Ayine, P.; Fadamiro, M.; Babu, J.R.; Brown, M.; Geetha, T. Urinary biomarkers of inflammation and oxidative stress are elevated in obese children and correlate with a marker of endothelial dysfunction. Oxid. Med. Cell Longev. 2019, 2019, 9604740. [Google Scholar] [CrossRef]
- Tsuda, T. Dietary anthocyanin-rich plants: Biochemical basis and recent progress in health benefits studies. Mol. Nutr. Food Res. 2012, 56, 159–170. [Google Scholar] [CrossRef]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clin. Invest. 2001, 107, 7–11. [Google Scholar] [CrossRef]
- Huang, W.Y.; Liu, Y.M.; Wang, J.; Wang, X.N.; Li, C.Y. Anti-inflammatory effect of the blueberry anthocyanins malvidin-3-glucoside and malvidin-3-galactoside in endothelial cells. Molecules 2014, 19, 12827–12841. [Google Scholar] [CrossRef] [Green Version]
- Decendit, A.; Mamani-Matsuda, M.; Aumont, V.; Waffo-Teguo, P.; Moynet, D.; Boniface, K.; Richard, E.; Krisa, S.; Rambert, J.; Merillon, J.M.; et al. Malvidin-3-O-beta glucoside, major grape anthocyanin, inhibits human macrophage-derived inflammatory mediators and decreases clinical scores in arthritic rats. Biochem. Pharmacol. 2013, 86, 1461–1467. [Google Scholar] [CrossRef]
- Serra, D.; Paixao, J.; Nunes, C.; Dinis, T.C.; Almeida, L.M. Cyanidin-3-glucoside suppresses cytokine-induced inflammatory response in human intestinal cells: Comparison with 5-aminosalicylic acid. PLoS ONE 2013, 8, e73001. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Zhang, M.; Chen, H.; Peng, J.; Jiang, F.; Shi, X.; Bai, Y.; Jian, M.; Jia, Y. Anthocyanins from black peanut skin protect against UV-B induced keratinocyte cell and skin oxidative damage through activating Nrf 2 signaling. Food Funct. 2019, 10, 6815–6828. [Google Scholar] [CrossRef]
- Krenn, L.; Paper, D.H. Inhibition of angiogenesis and inflammation by an extract of red clover (Trifolium pratense L.). Phytomedicine Int. J. Phytother. Phytopharm. 2009, 16, 1083–1088. [Google Scholar] [CrossRef]
- Mueller, M.; Hobiger, S.; Jungbauer, A. Red clover extract: A source for substances that activate peroxisome proliferator-activated receptor Î ± and ameliorate the cytokine secretion profile of lipopolysaccharide-stimulated macrophages. Menopause 2010, 17, 379–387. [Google Scholar] [CrossRef]
- Lee, S.G.; Parks, J.S.; Kang, H.W. Quercetin, a functional compound of onion peel, remodels white adipocytes to brown-like adipocytes. J. Nutr. Biochem. 2017, 42, 62–71. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.K.; Rasmussen, H.E.; Ehlers, S.J.; Blobaum, K.R.; Lu, F.; Schlegal, V.L.; Carr, T.P.; Lee, J.Y. Repression of proinflammatory gene expression by lipid extract of Nostoc commune var sphaeroides Kutzing, a blue-green alga, via inhibition of nuclear factor-kappaB in RAW 264.7 macrophages. Nutr. Res. (New York, NY.) 2008, 28, 83–91. [Google Scholar] [CrossRef] [Green Version]
- Cho, M.J.; Howard, L.R.; Prior, R.L.; Clark, J.R. Flavonoid glycosides and antioxidant capacity of various blackberry, blueberry and red grape genotypes determined by high-performance liquid chromatography/mass spectrometry. J. Sci. Food Agric. 2004, 84, 1771–1782. [Google Scholar] [CrossRef]
- Fan, J.; Heller, N.M.; Gorospe, M.; Atasoy, U.; Stellato, C. The role of post-transcriptional regulation in chemokine gene expression in inflammation and allergy. Eur. Respir. J. 2005, 26, 933–947. [Google Scholar] [CrossRef]
- Kontoyiannis, D.; Pasparakis, M.; Pizarro, T.T.; Cominelli, F.; Kollias, G. Impaired on/off regulation of TNF biosynthesis in mice lacking TNF AU-rich elements: Implications for joint and gut-associated immunopathologies. Immunity 1999, 10, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Surh, Y.J.; Chun, K.S.; Cha, H.H.; Han, S.S.; Keum, Y.S.; Park, K.K.; Lee, S.S. Molecular mechanisms underlying chemopreventive activities of anti-inflammatory phytochemicals: Down-regulation of COX-2 and iNOS through suppression of NF-kappa B activation. Mutat. Res. 2001, 480, 243–268. [Google Scholar] [CrossRef]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef]
- Bedard, K.; Krause, K.H. The NOX family of ROS-generating NADPH oxidases: Physiology and pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef]
- El-Benna, J.; Dang, P.M.; Gougerot-Pocidalo, M.A.; Marie, J.C.; Braut-Boucher, F. p47phox, the phagocyte NADPH oxidase/NOX2 organizer: Structure, phosphorylation and implication in diseases. Exp. Mol. Med. 2009, 41, 217–225. [Google Scholar] [CrossRef] [Green Version]
- O′Leary, D.P.; Bhatt, L.; Woolley, J.F.; Gough, D.R.; Wang, J.H.; Cotter, T.G.; Redmond, H.P. TLR-4 signalling accelerates colon cancer cell adhesion via NF-kappaB mediated transcriptional up-regulation of Nox-1. PLoS ONE 2012, 7, e44176. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.G.; Kim, B.; Yang, Y.; Pham, T.X.; Park, Y.K.; Manatou, J.; Koo, S.I.; Chun, O.K.; Lee, J.Y. Berry anthocyanins suppress the expression and secretion of proinflammatory mediators in macrophages by inhibiting nuclear translocation of NF-kappaB independent of NRF2-mediated mechanism. J. Nutr. Biochem. 2014, 25, 404–411. [Google Scholar] [CrossRef]
- Khorasani Esmaeili, A.; Mat Taha, R.; Mohajer, S.; Banisalam, B. Antioxidant activity and total phenolic and flavonoid content of various solvent extracts from in vivo and in vitro grown Trifolium pratense L. (Red Clover). Biomed Res. Int. 2015, 2015, 643285. [Google Scholar] [CrossRef] [Green Version]
- Booth, N.L.; Overk, C.R.; Yao, P.; Burdette, J.E.; Nikolic, D.; Chen, S.N.; Bolton, J.L.; Van Breemen, R.B.; Pauli, G.F.; Farnsworth, N.R. The chemical and biologic profile of a red clover (Trifolium pratense L.) phase II clinical extract. J. Altern. Complement. Med. 2006, 12, 133–139. [Google Scholar] [CrossRef] [Green Version]
- Dedio, W.; Clark, K.W. Influence of cytokinins on isoflavone and anthocyanin synthesis in red clover seedlings. Pestic. Sci. 1971, 2, 65–68. [Google Scholar] [CrossRef]
- Wu, X.; Prior, R.L. Systematic identification and characterization of anthocyanins by HPLC-ESI-MS/MS in common foods in the United States: Fruits and berries. J. Agric. Food Chem. 2005, 53, 2589–2599. [Google Scholar] [CrossRef]
- Jabri, M.A.; Rtibi, K.; Ben-Said, A.; Aouadhi, C.; Hosni, K.; Sakly, M.; Sebai, H. Antidiarrhoeal, antimicrobial and antioxidant effects of myrtle berries (Myrtus communis L.) seeds extract. J. Pharm. Pharmacol. 2016, 68, 264–274. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| COX2 | GCCTACTACAAGTGTTTCTTTTTGCA | CATTTTGTTTGATTGTTCACACCAT |
| GAPDH | GGTGGTCTCCTCTGACTTCAACA | GTTGCTGTAGCCAAATTCGTTGT |
| IL1β | GTCACAAGAAACCATGGCACAT | GCCCATCAGAGGCAAGGA |
| iNOS | AATCTTGGAGCGAGTTGTGG | CAGGAAGTAGGTGAGGGCTTG |
| MCP1 | CTTCTGGGCCTGCTGTTCA | CCAGCCTACTCATTGGGATCA |
| NOX1 | TTCACAGTTATTCATATCATTGC | AGAGAACAGAAGCGAGAG |
| NRF2 | CTCGCTGGAAAAAGAAGTG | CCGTCCAGGAGTTCAGAGG |
| TNFα | GGCTGCCCCGACTACGT | ACTTTCTCCTGGTATGAGATAGCAAAT |
| Number | RT (min) | Isoflavone Derivatives | Amount (μg/g dw) | [M-H]-m/z |
|---|---|---|---|---|
| 1 | 27.1 | Luteolin 7-O-β-D-glucoside | 368.5 ± 14.7 | 449, 287 |
| 2 | 28.7 | unknown tetrahydroxyflavone glucoside | 2394.3 ± 28.0 | 449, 287 |
| 3 | 29.2 | unknown tetrahydroxyflavone glucoside | 1356.2 ± 18.0 | 449, 287 |
| 4 | 31.5 | Isoquercitrin-6″-O-malonate | 1677.9 ± 20.9 | 551, 303 |
| 5 | 32.3 | Pratensein-7-O-β-D-glucoside | 682.1 ± 6.3 | 463, 301 |
| 6 | 33.4 | Genistin | 12892.3 ± 35.6 | 433, 271, 153 |
| 7 | 33.9 | Hyperoside | 3103.0 ± 20.9 | 465, 303 |
| 8 | 34.3 | Isoquercetriin | 8940.1 ± 16.6 | 465, 303 |
| 9 | 35.0 | Apigenin-7-O-β-D-glucoside | 512.1 ± 1.7 | 433, 271 |
| 10 | 36.1 | Pseudobaptigenin | 5809.6 ± 14.0 | 283 |
| 11 | 37.0 | unknown tetrahydroxyflavone glucoside | 24560.7 ± 60.7 | 449, 287 |
| 12 | 38.4 | Kaempferol or Luteolin | 5022.7 ± 37.8 | 287 |
| 13 | 38.9 | Gemosteom-7-O-β-D-glucoside-6″-O-malonate | 17933.8 ± 123.6 | 519, 271 |
| 14 | 39.3 | unknown tetrahydroxyflavone glucoside | 2691.2 ± 13.6 | 449, 287 |
| 15 | 40.2 | 3-methylquercetin-7-O-β-D-glucoside | 7838.7 ± 7.6 | 479, 317 |
| 16 | 42.7 | Kaempferol or Luteolin | 12876.9 ± 16.1 | 287 |
| 17 | 44.3 | Pratensein-7-O-β-D-glucoside-6″-malonate | 2437.5 ± 8.8 | 549, 317 |
| 18 | 44.9 | Pseudobaptigenin-7-O-β-D-glucoside | 2280.1 ± 8.4 | 445, 283 |
| 19 | 45.5 | Kaempferol or Luteolin | 2213.7 ± 9.1 | 287 |
| 20 | 46.7 | Glycitein | 312.1 ± 15.1 | 285, 167 |
| 21 | 47.5 | Pseudobaptigenin-7-O-β-D-glucoside-6″-O-malonate | 2361.1 ± 47.3 | 445, 283, 137 |
| 22 | 52.2 | Formononetin-7-O-β-D-glucoside-6″-O-malonate | 2231.1 ± 19.0 | 517, 269 |
| 23 | 53.7 | Calysosin-Glucoside-Malonate | 213.7 ± 15.9 | 533, 285, 137 |
| 24 | 57.0 | Prunetin-4′-O-β-D-glucoside-6″-O-malonate | 2089.3 ± 38.1 | 533, 285 |
| 25 | 58.7 | Formononetin | 497.3 ± 13.6 | 269, 137 |
| 26 | 60.2 | Biochanin A-7-O-β-D-glucoside-6″-O-malonate | 3713.8 ± 3.1 | 533, 285 |
| 27 | 66.8 | Biochanin A | 1059.8 ± 14.8 | 285 |
| Total | 128069.5 ± 628.9 |
| Number | RT (min) | Anthocyanins | Amount (μg/g dw) | [M-H]-m/z |
|---|---|---|---|---|
| 1 | 27.5 | Delphinidin-3, 5-O-diglucoside | 139.0 ± 1.1 | 627, 465, 303 |
| 2 | 29.9 | Cyanidin-3-O-galactoside | 380.3 ± 2.2 | 449,287 |
| 3 | 31.5 | Cyanidin-3-O-glucoside | 47.7 ± 3.3 | 449,287 |
| 4 | 33.5 | Petunidin-3-O-galactoside | 145.5 ± 4.4 | 479,317 |
| 5 | 34.4 | Peonidin-3-O-galactoside | 633.6 ± 5.5 | 463,301 |
| 6 | 36.3 | Malvidin-3-O-galactoside | 2129.4 ± 6.6 | 493,331 |
| 7 | 37.3 | Petunidin-3-O-rutinoside | 123.9 ± 7.7 | 625,479,317 |
| Total | 3599.5 ± 10.4 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.G.; Brownmiller, C.R.; Lee, S.-O.; Kang, H.W. Anti-Inflammatory and Antioxidant Effects of Anthocyanins of Trifolium pratense (Red Clover) in Lipopolysaccharide-Stimulated RAW-267.4 Macrophages. Nutrients 2020, 12, 1089. https://doi.org/10.3390/nu12041089
Lee SG, Brownmiller CR, Lee S-O, Kang HW. Anti-Inflammatory and Antioxidant Effects of Anthocyanins of Trifolium pratense (Red Clover) in Lipopolysaccharide-Stimulated RAW-267.4 Macrophages. Nutrients. 2020; 12(4):1089. https://doi.org/10.3390/nu12041089
Chicago/Turabian StyleLee, Sang Gil, Cindi R. Brownmiller, Sun-Ok Lee, and Hye Won Kang. 2020. "Anti-Inflammatory and Antioxidant Effects of Anthocyanins of Trifolium pratense (Red Clover) in Lipopolysaccharide-Stimulated RAW-267.4 Macrophages" Nutrients 12, no. 4: 1089. https://doi.org/10.3390/nu12041089
APA StyleLee, S. G., Brownmiller, C. R., Lee, S.-O., & Kang, H. W. (2020). Anti-Inflammatory and Antioxidant Effects of Anthocyanins of Trifolium pratense (Red Clover) in Lipopolysaccharide-Stimulated RAW-267.4 Macrophages. Nutrients, 12(4), 1089. https://doi.org/10.3390/nu12041089