1. Introduction
Fish oil supplements are one of the most commonly used non-vitamin/non-mineral natural products consumed in the United States and are used for several putative health benefits, including for their atheroprotective benefits on heart disease [
1]. The main active ingredient in fish oils appears to be n-3 polyunsaturated fatty acids (PUFA), namely eicosapentaenoic acid (EPA; 20:5 n-3) and docosahexaenoic acid (DHA; 22:6 n-3). N-3 PUFA can modify a variety of cellular processes associated with lipid metabolism, inflammation, thrombosis, and atherosclerosis [
2]. Results from various epidemiological and clinical studies have also demonstrated the important role of n-3 PUFA in decreasing triglycerides (TG) [
3].
Although TG lowering is a consistent observation, the effect of dietary n-3 fish oil on other cardiovascular disease (CVD) risk variables are less consistent. In fact, several large clinical trials, as well as a recent meta-analysis involving more than 77,000 individuals, have failed to show a cardiovascular benefit for fish oil supplementation containing various amounts of EPA and DHA [
4,
5]. It is possible that the inconsistencies of fish oil studies derive from differences in their n-3 PUFA dose and study design, but also due to the differential effect of EPA and DHA on metabolic pathways related to cardiovascular disease. Regarding lipid metabolism, both complementary and divergent effects have been described for EPA and DHA. Generally, DHA, but not EPA, tends to increase plasma levels of low-density lipoprotein cholesterol (LDL-C) and high-density lipoprotein cholesterol (HDL-C) [
6]. The mechanism for this difference between EPA and DHA is not fully understood, but it has been suggested to be related to their different effect on lipoprotein subfractions. The main lipoproteins, VLDL, LDL and HDL, exist in many different size subfractions, which vary in their relationship to cardiovascular risk [
7,
8]. Some human trials show that DHA increased LDL particle size and that EPA and DHA had different effects on HDL subfractions [
9]. When purified EPA and DHA were directly compared, DHA shows a stronger effect in increasing larger HDL2 particle levels, whereas EPA decreases smaller HDL3 particle levels to a greater extent than DHA [
9].
Most n-3 fish oil clinical trials have used a mixture of EPA and DHA with various ratios and have not carefully examined the potential differential impact of the two types of fatty acids (FAs) on lipids and lipoproteins [
5,
10]. Thus, it remains largely unknown whether fish oil supplements with different ratios of EPA to DHA may possibly differ in their cardiovascular benefit. Natural sources of fish oils can substantially vary in the relative amounts of EPA and DHA that they contain. For example, tuna oil contains more DHA than EPA, whereas anchovy and sardine oil contain more EPA than DHA [
11,
12]. Consequently, the molar ratio of EPA to DHA in over-the-counter fish oil supplements can vary from approximately 0.3 to 3 [
13], depending on the source of fish used to extract the oil. In the current study, we used two different fish oil formulations, representing a wide range of EPA/DHA ratios (0.3 and 2.3), and compared their effect on lipid and lipoprotein levels, as well as other CVD risk factors, in healthy normolipidemic adults.
5. Discussion
This study addressed whether fish oil supplements with varied EPA/DHA ratios have different effects on lipoprotein metabolism in healthy normolipidemic adults. We chose two fish oil supplements from natural sources that nearly spans the range for EPA/DHA that can be found in over-the-counter fish oil supplements. Our data indicate that 12 g/day of EPA- or DHA-rich fish oil supplement (~4.8 g/d total EPA+DHA) were similarly well tolerated in normolipidemic subjects, and that both fish oil supplements similarly decreased plasma TG levels, as well as overall particle numbers of VLDL and TG-rich lipoprotein subfractions.
Although fish oils have some other clinical indications, such as for reducing the risk of acute pancreatitis from hypertriglyceridemia [
20], the main driver for their use by the public has been for their perceived benefit in reducing cardiovascular disease [
2]. The most recent recommendations from the American Heart Association concluded that n-3 PUFA at high doses of 4 g/d (>3 g/d total EPA+DHA) is effective in reducing TG in hyperlipidemic individuals [
21]. It has not been clearly demonstrated, however, that lowering TG by other drugs like fibrates can decrease cardiovascular events [
22]. The REDUCE-IT trial, which used 4 g of purified EPA in patients with modest hypertriglyceridemia, did show a substantial further reduction in cardiovascular events when used on top of statins, but it is still uncertain whether it was due to TG lowering or by some other mechanism [
23]. Recently, the STRENTH trial of using 4 g of a mixture of EPA and DHA (EPA to DHA ratio: 2.75) has been halted because of a lack of an apparent effect in reducing CVD outcomes in patients with mixed dyslipidaemia [
24], raising the question of the efficacy of fish oil supplements with different ratios of EPA to DHA on CVD risk. Purified EPA is only available as a prescription medication, whereas all the over-the-counter forms of fish oil supplements contain a varying mixture of EPA and DHA.
In the current study, plasma TG levels were decreased by 14.4% on average due to fish oil supplements containing a total for 4.8 g of EPA plus DHA, which agrees with previous finding showing that ≥4 g/d of n-3 PUFA results in 9%–26% reduction in circulating TG in normolipidemic to borderline hyperlipidemic individuals [
3]. When comparing the effectiveness of the two fish oils used in this study, no differences in TG-lowering effect were observed, which is also in line with previous studies [
25,
26]. The mechanism related to TG lowering by fish oil supplementation is thought to be due to suppression of hepatic VLDL production, one of the main lipoprotein carriers of TG [
27]. When we examined both VLDL-P and size and TRL-P and size by NMR, we observed that both fish oil supplements equally lowered larger particles, which are more enriched in TG.
Neither fish oil supplement in this study changed plasma levels of LDL-C as determined by the Friedewald equation or by a homogenous direct assay (Denka). Pro-atherogenic sdLDL-C also did not change, as well as plasma PCSK9, which regulates circulating LDL-C levels [
28]. Both fish oil supplements, however, showed a similar small effect in raising apoB between 5.4%–6.8%, which given the greater predictive value of apoB over LDL-C on the impact of a therapy on cardiovascular outcomes suggests that neither fish oil treatment used in this study may be beneficial [
29]. There was also a trend toward increased LDL-P for both fish oils consistent with changes in apoB, but it did not reach statistical significance. When LDL subfractions were individually analyzed there was a statistically significant increase in small over large LDL for both fish oil supplements, which would be expected to increase CVD risk [
30]. When comparing on-treatment values, there was a small but significantly greater increase in overall LDL-P levels for DHA-rich oil compared to EPA-rich oil. Several previous studies have shown that the effect of DHA on increasing LDL-C and LDL particle size was greater than for EPA [
9,
10], whereas other studies showed no change in LDL-C with either EPA or DHA. It has been suggested that the effect of n-3 fish oils on LDL-C is less consistent compared with their TG-lowering effect, and depends on multiple factors, such as study period and population, as well as n-3 PUFA dose and types. The differences observed in this study in LDL-C and LDL subclass profile on the EPA vs. DHA-rich fish oil supplements need to be examined in subjects with elevated LDL-C and other types of dyslipidemias, which could reveal greater differences in these two types of fish oil supplements.
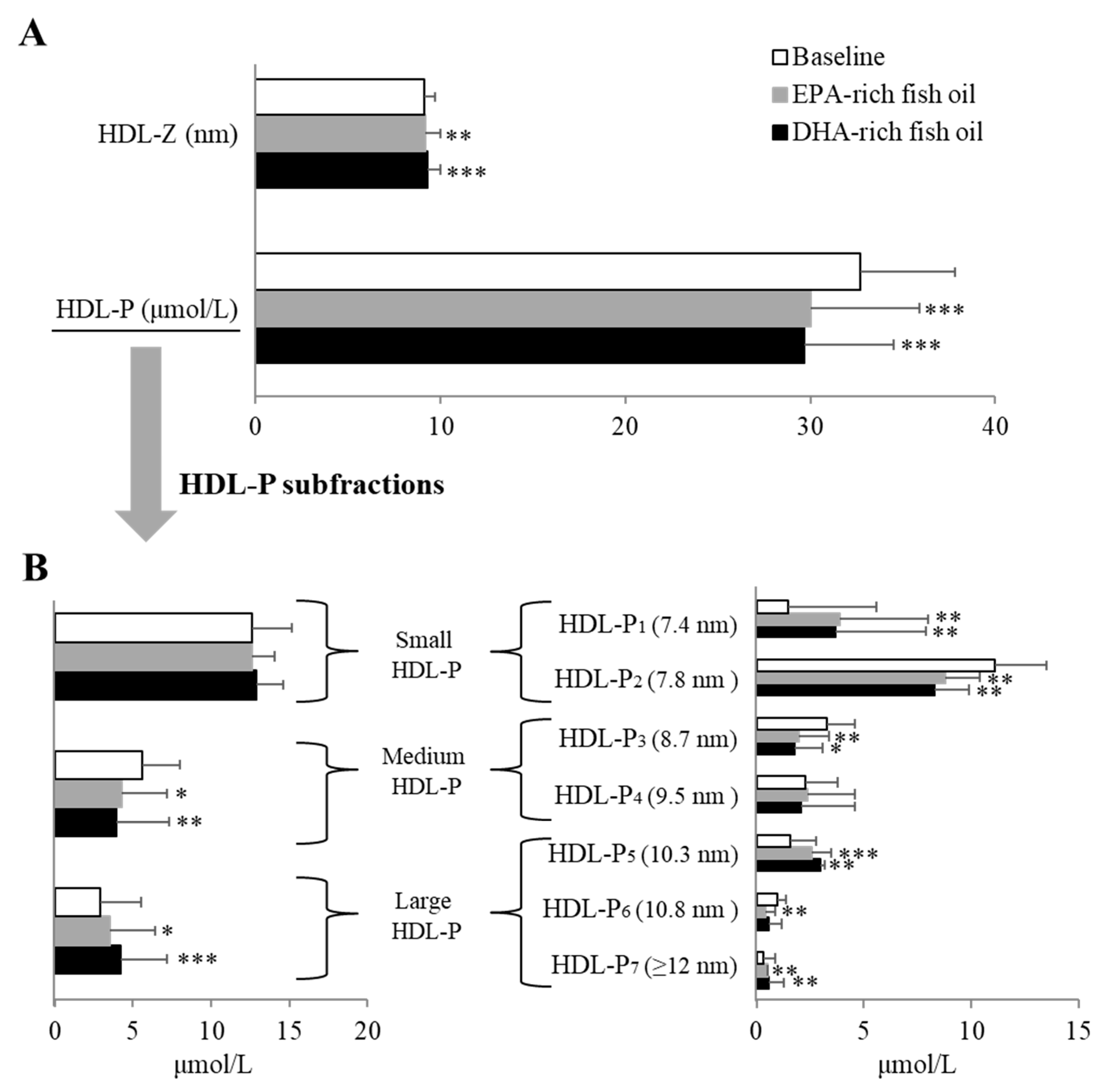
HDL, which is generally considered to be atheroprotective [
31], has not been well studied in regard to the effect of fish oil supplementation on its composition and function. By meta-analysis, n-3 PUFA consumption appears to moderately increased HDL-C levels, but the effect is generally small, especially when study participants are healthy [
32]. In the currently study, neither fish oil supplement resulted in changes in HDL-C or in apoE-HDL, a minor HDL subclass thought to be more cardioprotective [
33]. Given the multitude of potential functions of HDL and its complicated subtraction distribution, HDL-C may be a poor metric of its function, as has been shown in the case of cholesterol efflux [
31,
34]. We, therefore, also examined its subfractions by NMR and observed again that the two fish oils showed nearly identical effects in causing a decrease in HDL-P. The smallest subfraction increased on both oils and this would, in general, be considered to be pro-atherogenic but another larger subfraction increased, making it difficult to predict the overall impact on CVD risk. The various proteins in HDL are also known to influence the functions of HDL [
35], and the HDL proteome has been reported to respond to dietary lipid composition [
36]. Another important function of HDL is its anti-inflammatory abilities that may contribute to its anti-atherogenic properties [
37]. Fish oil-derived n-3 PUFAs can possibly influence inflammatory processes by a variety of mechanisms, such as shared and different roles of EPA and DHA in producing bioactive metabolites [
38]. In the current study, we found the down-regulation of several proteins involved in inflammation by both fish oil supplements, but to a greater extent with EPA-rich fish oil. When compared to DHA-rich oil, EPA-rich oil significantly decreased the abundance of gelsolin, a multifunctional protein involved in multiple biological process, including modulation of inflammatory response [
39]. EPA-rich oil also resulted in a greater increase in the abundance of apoM in HDL fractions compared with DHA-rich oil. This was consistent with also the greater increase in plasma levels of apoM after EPA-rich oil supplementation. ApoM has several potential anti-atherosclerotic functions [
40]. ApoM forms a complex with sphingosine-1-phosphate (S1P), a potent bioactive lipid [
41], and delivers it to endothelial cells where it may decrease vascular permeability. Increased S1P on HDL has been shown to be inversely related to CVD events and treatment of mice with stable analogues of S1P have been shown to reduce atherosclerosis [
42]. ApoM has also been reported to enhance reverse cholesterol transport by promoting the formation of pre–β-HDL [
43]. The transcription and secretion of apoM are regulated by a series of transcription factors, such as liver X receptor and forkhead box A2, that are known to be modulated by dietary nutrients, including omega-3 fish oils [
44]. Previous animal studies have also showed that dietary n-3 PUFA-rich fish oil can enhance reverse cholesterol transport [
45], but at least for the first step of this process (cellular cholesterol efflux) [
46], we observed no difference in plasma CEC compared to baseline after either fish oil supplement. The limited and mixed findings of clinical studies on n-3 PUFAs containing different ratios of EPA and DHA warrant further investigation on the individual effect of EPA and DHA on lipoprotein subfractions, proteome, and other cardiovascular biomarkers besides those related to lipids.
Study Strengths and Limitations
A strength of this study is its double-blind, randomized, crossover design, which maximized our ability to detect differences between the two fish oil supplements. The two fish oil supplements also had a similar taste and appearance and were well matched in their FA profile except for having different EPA/DHA ratios. In addition, no major changes were found in lifestyle and dietary factors of the participant during the course of the study. Our study does have the following limitations: the sample size was relatively small, the supplement duration was short, participants were relatively healthy although they are generally overweight, it was a single dose design, and we did not collect red blood cells to measure membrane incorporation of DHA and EPA before and after treatment. In addition, we compared a large number of variables simultaneously therefore the multiple testing problem is likely to result in some significant results simply due to chance alone. Another limitation is that more research is needed to understand the clinical relevance in regard to atherogenesis for the differences we did observe in the effect of the two supplements on the NMR lipoprotein profile and the apoM content of HDL. Furthermore, no effects on inflammation and glucose homeostasis were observed with either fish oil supplement, but further studies are still warranted to examine EPA/DHA supplementation for specific subgroups of subjects with inflammation and insulin resistance.

 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}