Fermentation of Blackberry with L. plantarum JBMI F5 Enhance the Protection Effect on UVB-Mediated Photoaging in Human Foreskin Fibroblast and Hairless Mice through Regulation of MAPK/NF-κB Signaling


Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Antibodies
2.2. Preparation of FBB
2.3. Biochemical Analysis
2.4. Cell Culture
2.5. Cell Viability Assay
2.6. Animals
2.7. UVB Induced Skin Damage Model in Hairless Mice
2.8. Wrinkle Measurement
2.9. Measurement of Type I Procollagen and MMP-1
2.10. Determination of the Antioxidant Activity
2.11. Immunoblotting
2.12. Histological Analysis
2.13. Statistical Analysis
3. Results
3.1. Biochemical Changes by Fermentation
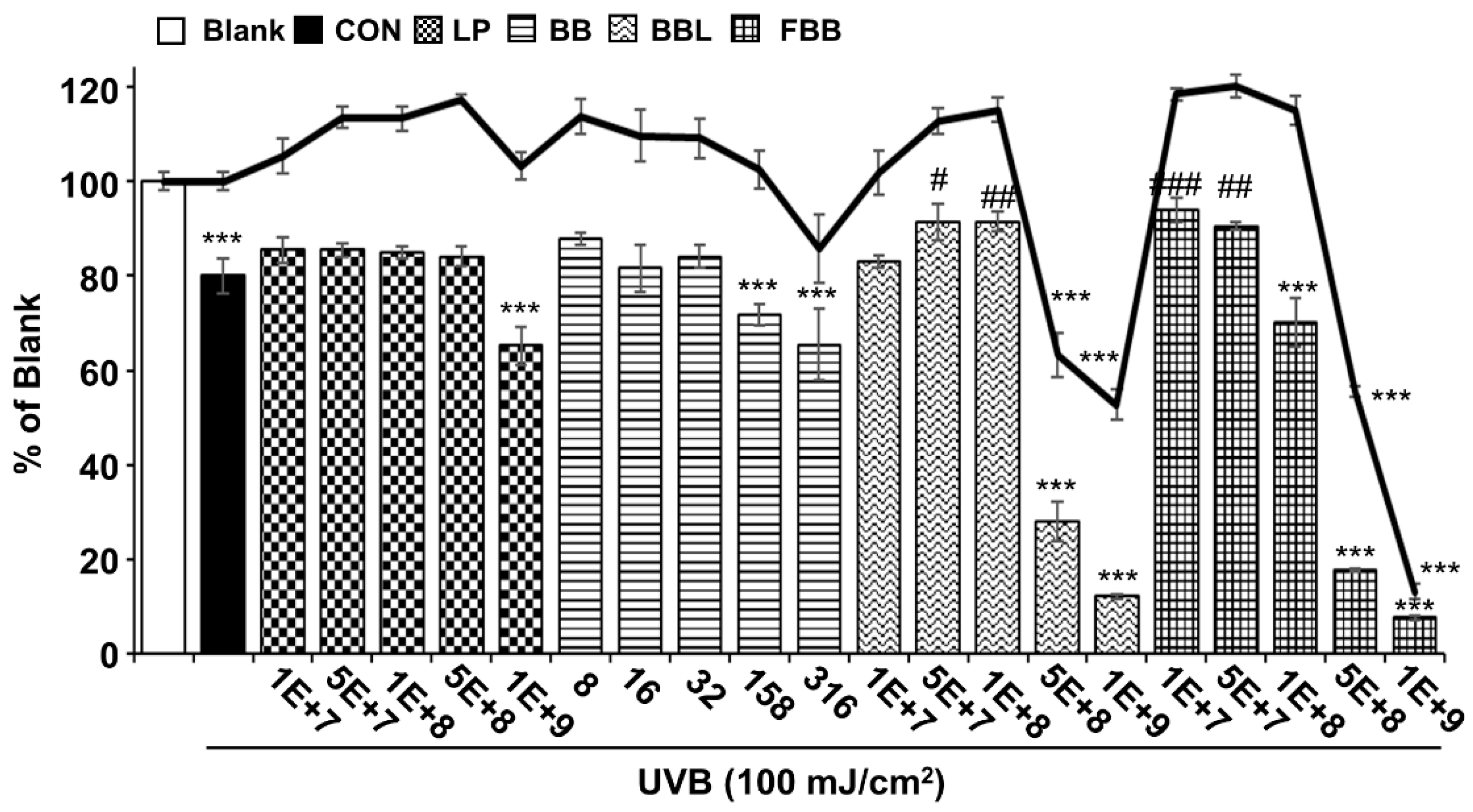
3.2. Protective Effects of FBB on UVB-Induced Damage in Hs68 Cells
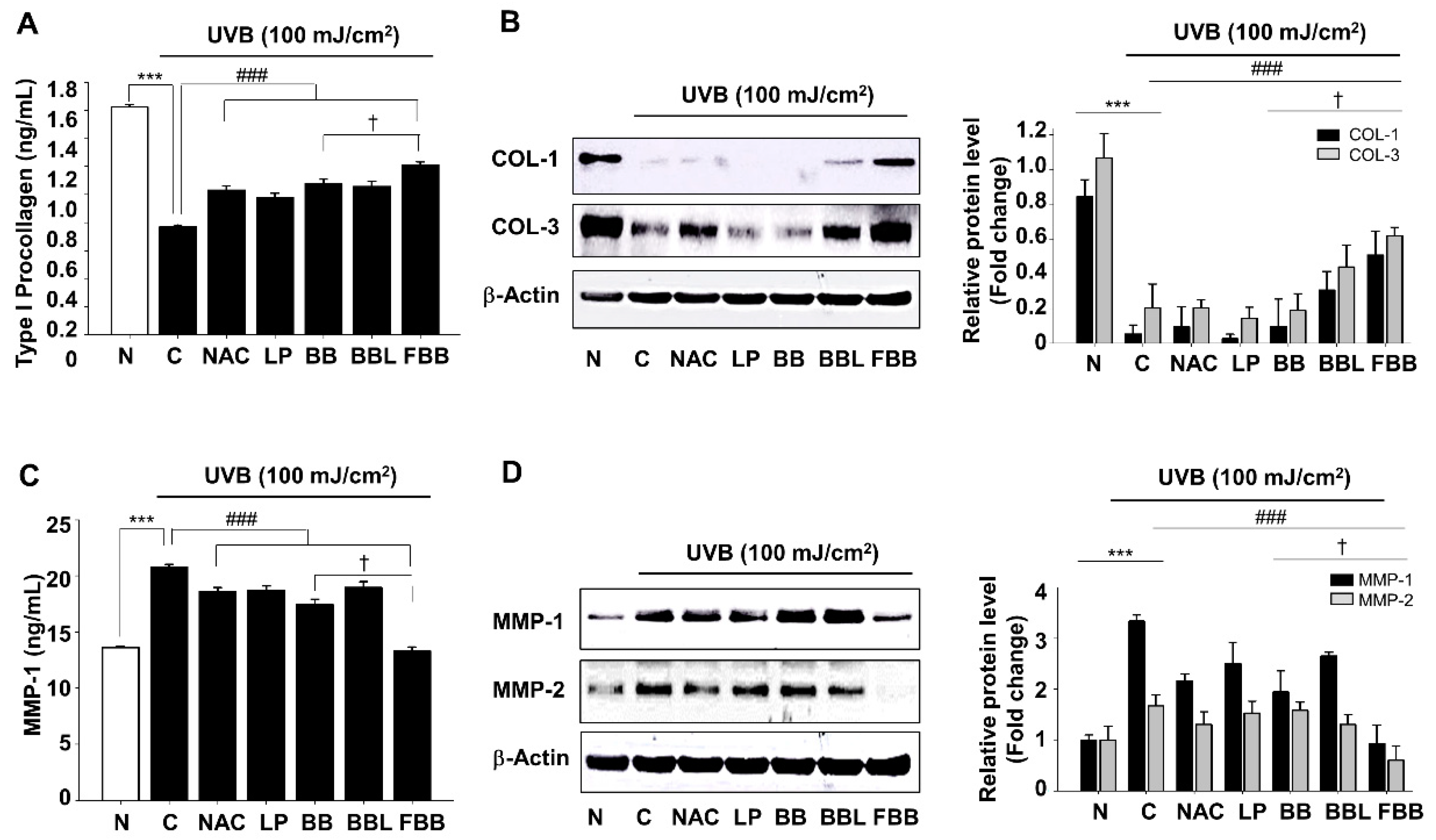
3.3. Regulation of UVB-Induced Collagens (COLs) Degradation and MMPs Expression by FBB in Hs68 Cells
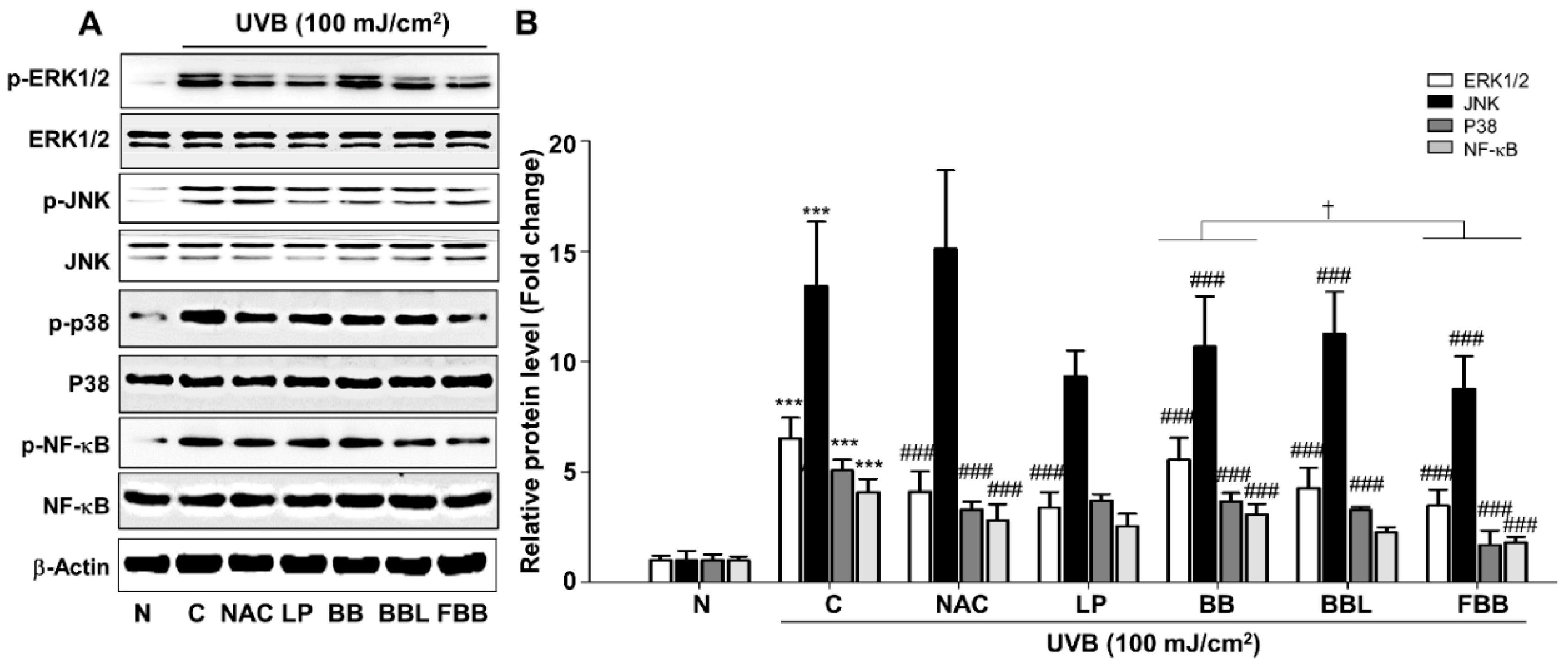
3.4. FBB Suppresses the MAPK and NF-κB Signaling in UVB-Exposed Hs68 Fibroblasts
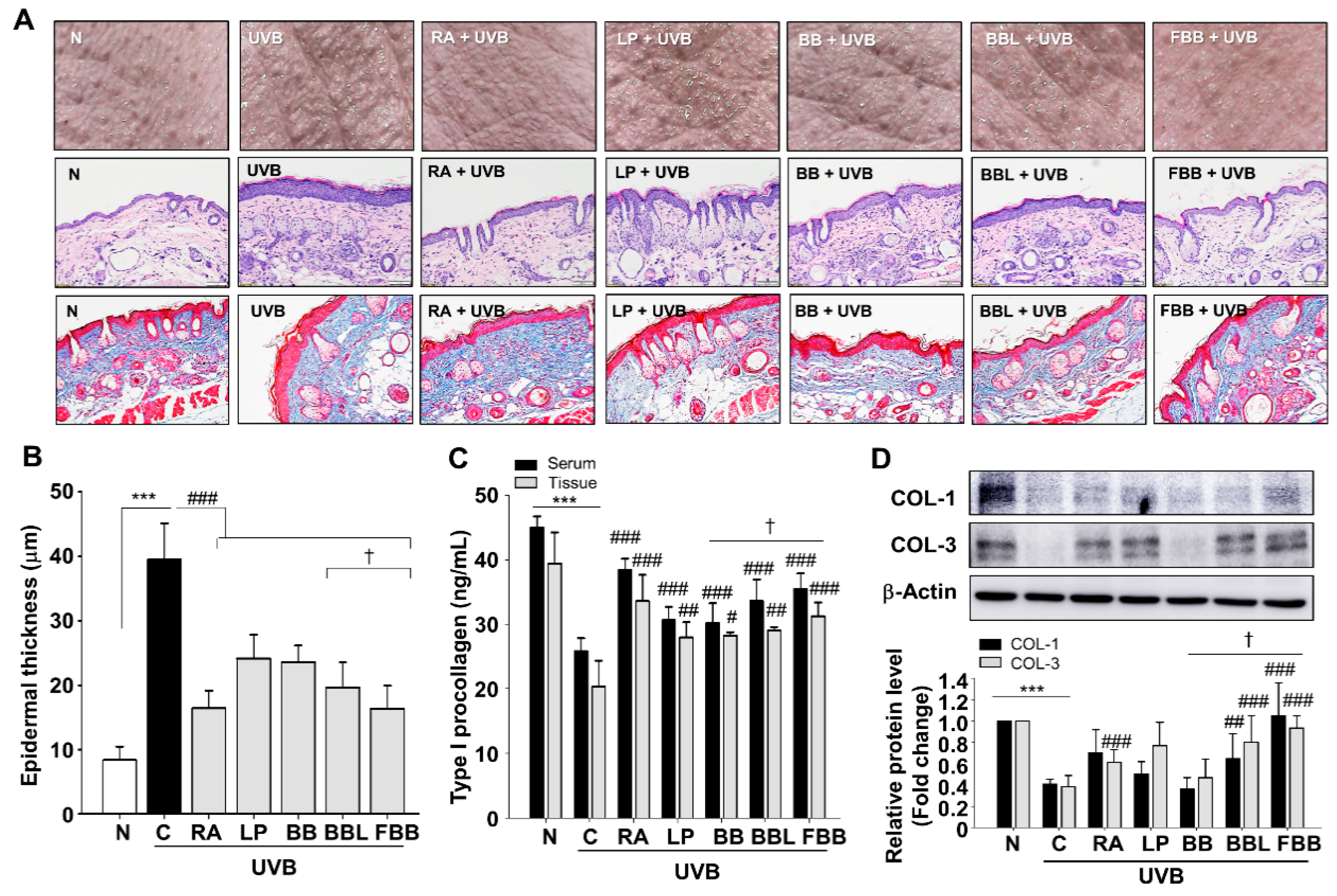
3.5. FBB Inhibits UVB-Induced Wrinkle Formation and COLs Degradation in SKH-1 Mice
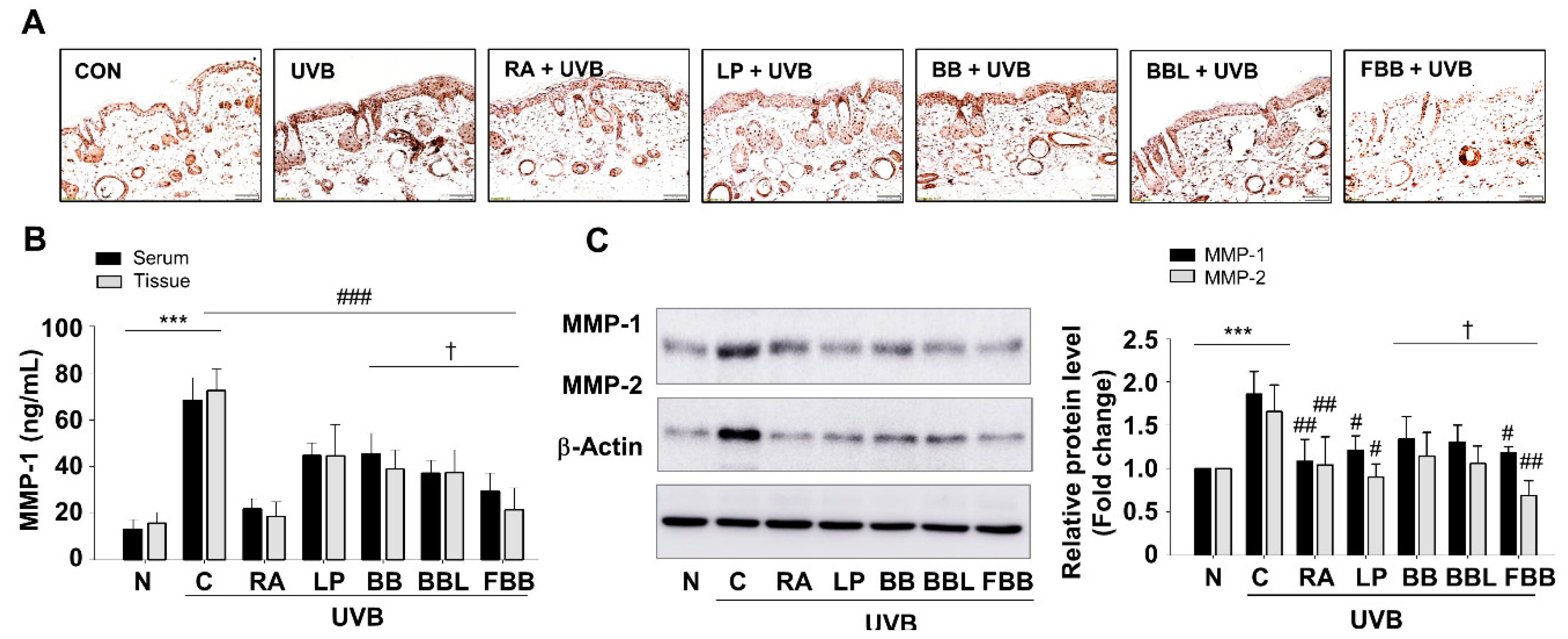
3.6. FBB Prevents UVB-Induced MMPs Production in SKH-1 Mice
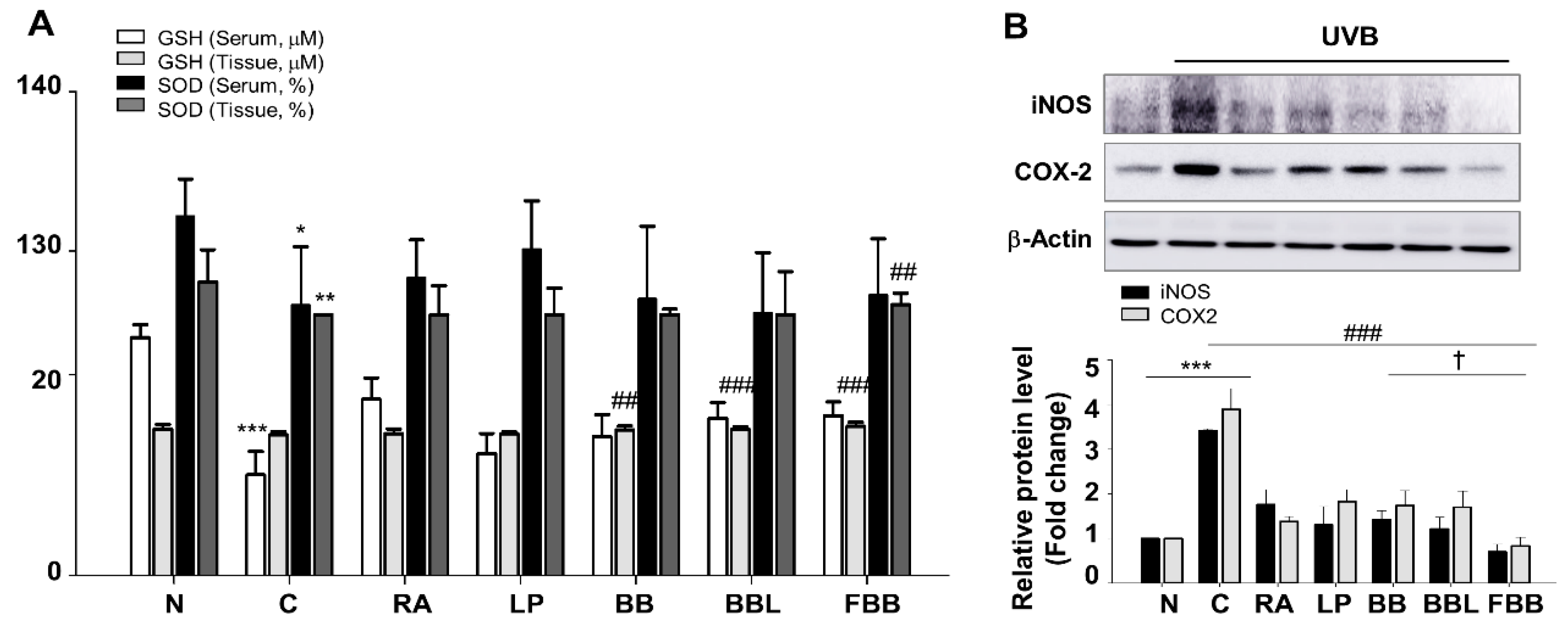
3.7. FBB Increases the Activity of Antioxidant Effcts
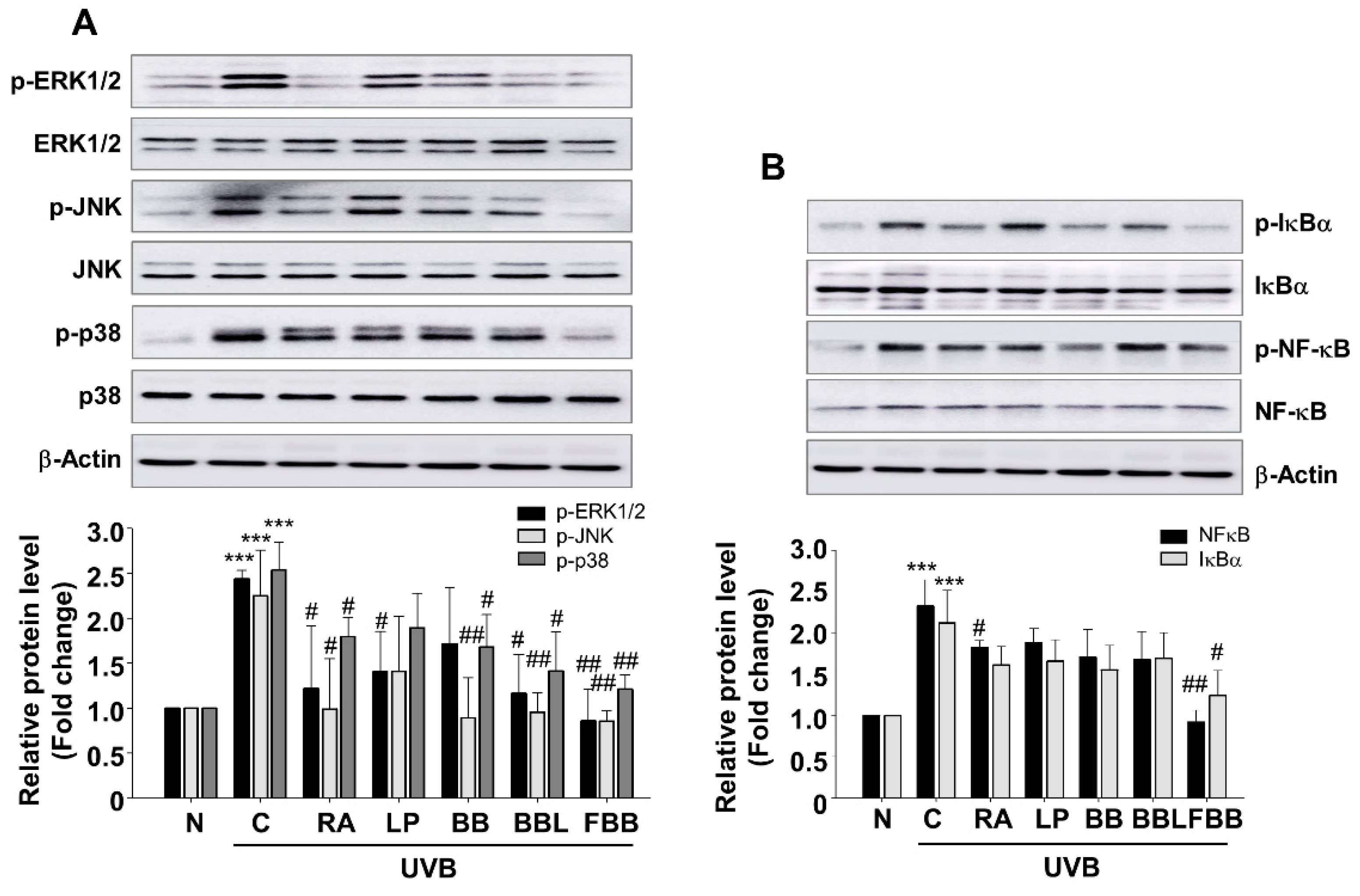
3.8. Effect of FBB on UVB-Mediated MAPK and NF-κB Signaling in SKH-1 Mice
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Naylor, E.C.; Watson, R.E.; Sherratt, M.J. Molecular aspects of skin ageing. Maturitas 2011, 69, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Quan, T.; Qin, Z.; Xia, W.; Shao, Y.; Voorhees, J.J.; Fisher, G.J. Matrix-degrading metalloproteinases in photoaging. J. Investig. Dermatol. Symp. Proc. 2009, 14, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Rittié, L. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002, 1, 705–720. [Google Scholar] [PubMed]
- Yamamoto, Y. Role of active oxygen species and antioxidants in photoaging. J. Dermatol. Sci. 2001, 27, 1–4. [Google Scholar] [CrossRef]
- Bickers, D.R.; Athar, M. Oxidative Stress in the Pathogenesis of Skin Disease. J. Investig. Dermatol. 2006, 126, 2565–2575. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Isolauri, E. The development of gut immune responses and gut microbiota: Effects of probiotics in prevention and treatment of allergic disease. Curr. Issues Intestig. Microbiol. 2002, 3, 15–22. [Google Scholar]
- Parvez, S.; Malik, K.; Kang, S.A.; Kim, H.-Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef] [PubMed]
- Bouilly-Gauthier, D.; Jeannes, C.; Maubert, Y.; Duteil, L.; Queille-Roussel, C.; Piccardi, N.; Montastier, C.; Manissier, P.; Piérard, G.; Ortonne, J.-P.; et al. Clinical evidence of benefits of a dietary supplement containing probiotic and carotenoids on ultraviolet-induced skin damage. Br. J. Dermatol. 2010, 163, 536–543. [Google Scholar] [CrossRef]
- Kim, H.M.; Lee, D.E.; Park, S.D.; Kim, Y.-T.; Kim, Y.J.; Jeong, J.W.; Jang, S.S.; Ahn, Y.-T.; Sim, J.-H.; Huh, C.-S.; et al. Oral administration of Lactobacillus plantarum HY7714 protects hairless mouse against ultraviolet B-induced photoaging. J. Microbiol. Biotechnol. 2014, 24, 1583–1591. [Google Scholar] [CrossRef]
- ElAzab, N.; Mendy, A.; Gasana, J.; Vieira, E.R.; Quizon, A.; Forno, E. Probiotic Administration in Early Life, Atopy, and Asthma: A Meta-analysis of Clinical Trials. Pediatrics 2013, 132, 666–676. [Google Scholar] [CrossRef]
- Murapa, P.; Dai, J.; Chung, M.; Mumper, R.J.; D’Orazio, J. Anthocyanin-rich Fractions of Blackberry Extracts Reduce UV-induced Free Radicals and Oxidative Damage in Keratinocytes. Phytother. Res. 2012, 26, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Divya, S.P.; Wang, X.; Pratheeshkumar, P.; Son, Y.-O.; Roy, R.V.; Kim, D.; Dai, J.; Hitron, J.A.; Wang, L.; Asha, P.; et al. Blackberry extract inhibits UVB-induced oxidative damage and inflammation through MAP kinases and NF-κB signaling pathways in SKH-1 mice skin. Toxicol. Appl. Pharmacol. 2015, 284, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Calvo-Castro, L.A.; Syed, D.N.; Chamcheu, J.C.; Vilela, F.M.P.; Perez, A.M.; Vaillant, F.; Rojas, M.; Mukhtar, H. Protective effect of tropical highland blackberry juice (Rubus adenotrichos Schltdl.) against UVB-mediated damage in human epidermal keratinocytes and in a reconstituted skin equivalent model. Photochem. Photobiol. 2013, 89, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Gul, R.; Rah, S.-Y.; Kim, S.H.; Park, S.K.; Im, M.-J.; Kwon, H.J.; Kim, U.-H. Molecular mechanism of ADP-ribosyl cyclase activation in angiotensin II signaling in murine mesangial cells. Am. J. Physiol. Physiol. 2008, 294, F982–F989. [Google Scholar] [CrossRef]
- Wei, X.; Luo, M.; Xu, L.; Zhang, Y.; Lin, X.; Kong, P.; Liu, H. Production of Fibrinolytic Enzyme from Bacillus amyloliquefaciens by Fermentation of Chickpeas, with the Evaluation of the Anticoagulant and Antioxidant Properties of Chickpeas. J. Agric. Food Chem. 2011, 59, 3957–3963. [Google Scholar] [CrossRef]
- Saliou, C.; Kitazawa, M.; McLaughlin, L.; Yang, J.-P.; Lodge, J.K.; Tetsuka, T.; Iwasaki, K.; Cillard, J.; Okamoto, T.; Packer, L. Antioxidants modulate acute solar ultraviolet radiation-induced NF-kappa-B activation in a human keratinocyte cell line. Free Radic. Boil. Med. 1999, 26, 174–183. [Google Scholar] [CrossRef]
- Lee, Y.-R.; Noh, E.-M.; Jeong, E.-Y.; Yun, S.-K.; Jeong, Y.-J.; Kim, J.-H.; Kwon, K.-B.; Kim, B.-S.; Lee, S.-H.; Park, C.-S.; et al. Cordycepin inhibits UVB-induced matrix metalloproteinase expression by suppressing the NF-κB pathway in human dermal fibroblasts. Exp. Mol. Med. 2009, 41, 548. [Google Scholar] [CrossRef]
- Tanaka, K.; Hasegawa, J.; Asamitsu, K.; Okamoto, T. Prevention of the Ultraviolet B-Mediated Skin Photoaging by a Nuclear Factor κB Inhibitor, Parthenolide. J. Pharmacol. Exp. Ther. 2005, 315, 624–630. [Google Scholar] [CrossRef]
- Matsumura, Y.; Ananthaswamy, H.N. Toxic effects of ultraviolet radiation on the skin. Toxicol. Appl. Pharmacol. 2004, 195, 298–308. [Google Scholar] [CrossRef]
- Gum, R.; Wang, H.; Lengyel, E.; Juarez, J.; Boyd, D. Regulation of 92 kDa type IV collagenase expression by the jun aminoterminal kinase- and the extracellular signal-regulated kinase-dependent signaling cascades. Oncogene 1997, 14, 1481–1493. [Google Scholar] [CrossRef] [PubMed]
- Van de Velde, F.; Esposito, D.; Grace, M.H.; Pirovani, M.E.; Lila, M.A. Anti-inflammatory and wound healing properties of polyphenolic extracts from strawberry and blackberry fruits. Food Res. Int. 2019, 121, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Şanlier, N.; Gökcen, B.B.; Sezgin, A.C. Health benefits of fermented foods. Crit. Rev. Food Sci. Nutr. 2019, 59, 506–527. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.; Lee, Y.; Huang, Y.-H.; Lim, H.-W.; Jang, K.; Kim, D.-D.; Lim, C.-J. Probiotic fermentation augments the skin anti-photoaging properties of Agastache rugosa through up-regulating antioxidant components in UV-B-irradiated HaCaT keratinocytes. BMC Complement. Altern. Med. 2018, 18, 196. [Google Scholar] [CrossRef] [PubMed]
- Curiel, J.A.; Pinto, D.; Marzani, B.; Filannino, P.; Farris, G.A.; Gobbetti, M.; Rizzello, C.G. Lactic acid fermentation as a tool to enhance the antioxidant properties of Myrtus communis berries. Microb. Cell Factories 2015, 14, 67. [Google Scholar] [CrossRef] [PubMed]
- Fisher, G.J.; Kang, S.; Varani, J.; Bata-Csorgo, Z.; Wan, Y.; Datta, S.; Voorhees, J.J. Mechanisms of photoaging and chronological skin aging. Arch. Dermatol. 2002, 138, 1462–1470. [Google Scholar] [CrossRef]
- Bigot, N.; Beauchef, G.; Hervieu, M.; Oddos, T.; Demoor, M.; Boumediene, K.; Galéra, P. NF-κB Accumulation Associated with COL1A1 Trans activators Defects during Chronological Aging Represses Type I Collagen Expression through a −112/−61-bp Region of the COL1A1 Promoter in Human Skin Fibroblasts. J. Investig. Dermatol. 2012, 132, 2360–2367. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Asamitsu, K.; Uranishi, H.; Iddamalgoda, A.; Ito, K.; Kojima, H.; Okamoto, T. Protecting skin photoaging by NF-kappaB inhibitor. Curr. Drug Metab. 2010, 11, 431–435. [Google Scholar] [CrossRef]
- Brenneisen, P.; Sies, H.; Scharffetter-Kochanek, K.; Scharffetter-Kochanek, K. Ultraviolet-B Irradiation and Matrix Metalloproteinases. Ann. N. Y. Acad. Sci. 2002, 973, 31–43. [Google Scholar] [CrossRef]
- Romana-Souza, B.; Monte-Alto-Costa, A. Olive oil inhibits ageing signs induced by chronic stress in ex vivo human skin via inhibition of extracellular-signal-related kinase 1/2 and c- JUN pathways. Int. J. Cosmet. Sci. 2019, 41, 156–163. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TPC (mg/g) | TFC (mg/g) | VC (mg/100 g) | DPPH Radical Scavenging Activity (%) | ABTS Radical Scavenging Activity (%) | |
|---|---|---|---|---|---|
| BB | 18.96 ± 0.25 | 0.24 ± 0.09 | 51.05 ± 3.36 | 5.17 ± 0.98 | 6.06 ± 0.45 |
| FBB | 28.86 ± 0.25 *** | 0.38 ± 0.02 * | 282.56 ± 34.63 *** | 14.19 ± 0.32 *** | 33.06 ± 0.88 *** |
| AA | NA | NA | NA | 94.24 ± 0.13 | NA |
| Trolox | NA | NA | NA | NA | 94.62 ± 0.08 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-R.; Jeong, D.-H.; Kim, S.; Lee, S.-W.; Sin, H.-S.; Yu, K.-Y.; Jeong, S.-I.; Kim, S.-Y. Fermentation of Blackberry with L. plantarum JBMI F5 Enhance the Protection Effect on UVB-Mediated Photoaging in Human Foreskin Fibroblast and Hairless Mice through Regulation of MAPK/NF-κB Signaling. Nutrients 2019, 11, 2429. https://doi.org/10.3390/nu11102429
Kim H-R, Jeong D-H, Kim S, Lee S-W, Sin H-S, Yu K-Y, Jeong S-I, Kim S-Y. Fermentation of Blackberry with L. plantarum JBMI F5 Enhance the Protection Effect on UVB-Mediated Photoaging in Human Foreskin Fibroblast and Hairless Mice through Regulation of MAPK/NF-κB Signaling. Nutrients. 2019; 11(10):2429. https://doi.org/10.3390/nu11102429
Chicago/Turabian StyleKim, Ha-Rim, Da-Hye Jeong, Sol Kim, Sang-Wang Lee, Hong-Sig Sin, Kang-Yeol Yu, Seung-Il Jeong, and Seon-Young Kim. 2019. "Fermentation of Blackberry with L. plantarum JBMI F5 Enhance the Protection Effect on UVB-Mediated Photoaging in Human Foreskin Fibroblast and Hairless Mice through Regulation of MAPK/NF-κB Signaling" Nutrients 11, no. 10: 2429. https://doi.org/10.3390/nu11102429
APA StyleKim, H.-R., Jeong, D.-H., Kim, S., Lee, S.-W., Sin, H.-S., Yu, K.-Y., Jeong, S.-I., & Kim, S.-Y. (2019). Fermentation of Blackberry with L. plantarum JBMI F5 Enhance the Protection Effect on UVB-Mediated Photoaging in Human Foreskin Fibroblast and Hairless Mice through Regulation of MAPK/NF-κB Signaling. Nutrients, 11(10), 2429. https://doi.org/10.3390/nu11102429