
Detection of Aflatoxin B1 in Single Peanut Kernels by Combining Hyperspectral and Microscopic Imaging Technologies


 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Instruments
2.3. Data Analysis
3. Results and Discussions
3.1. Statistical Analyses of AFB1 Accumulation
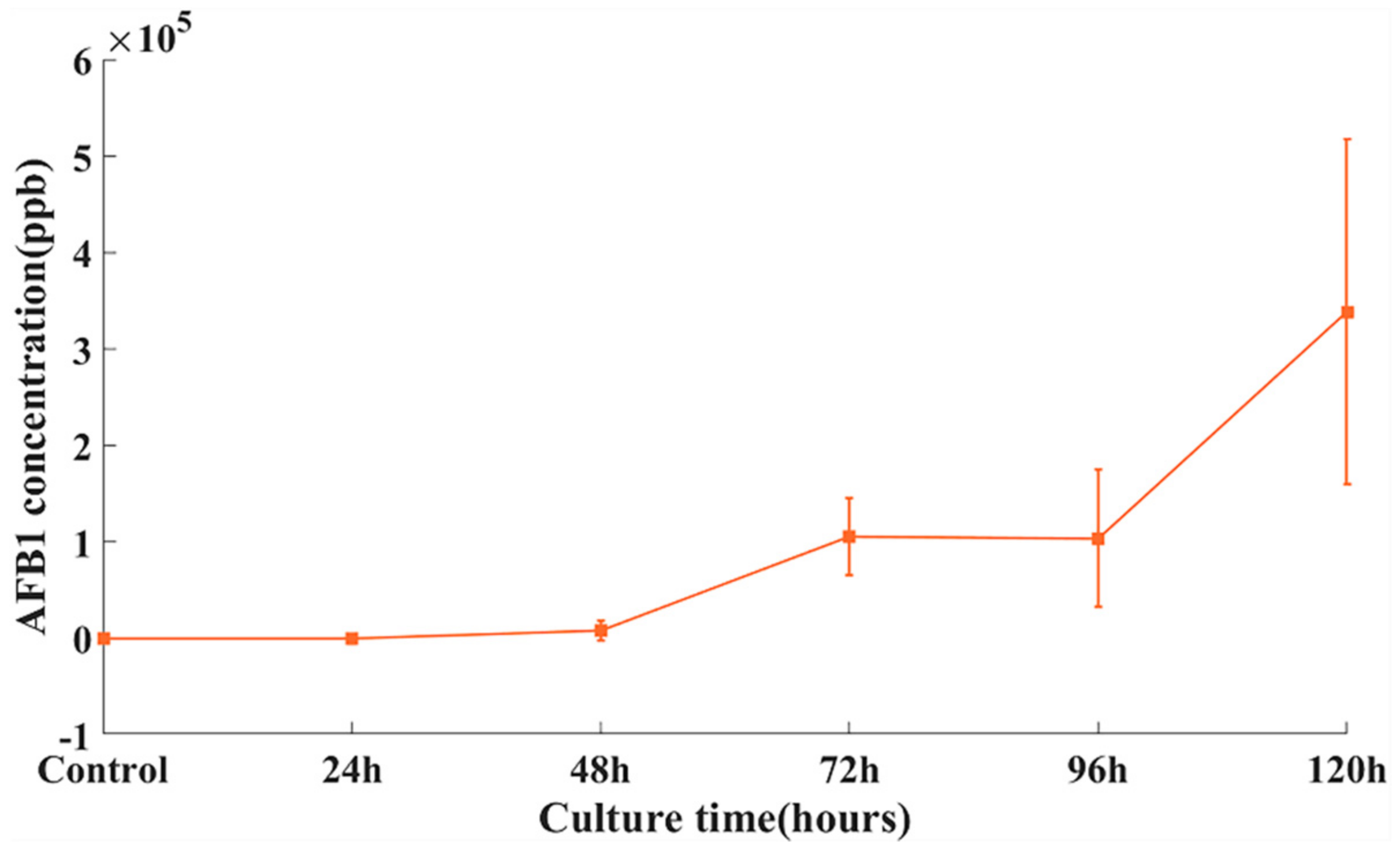
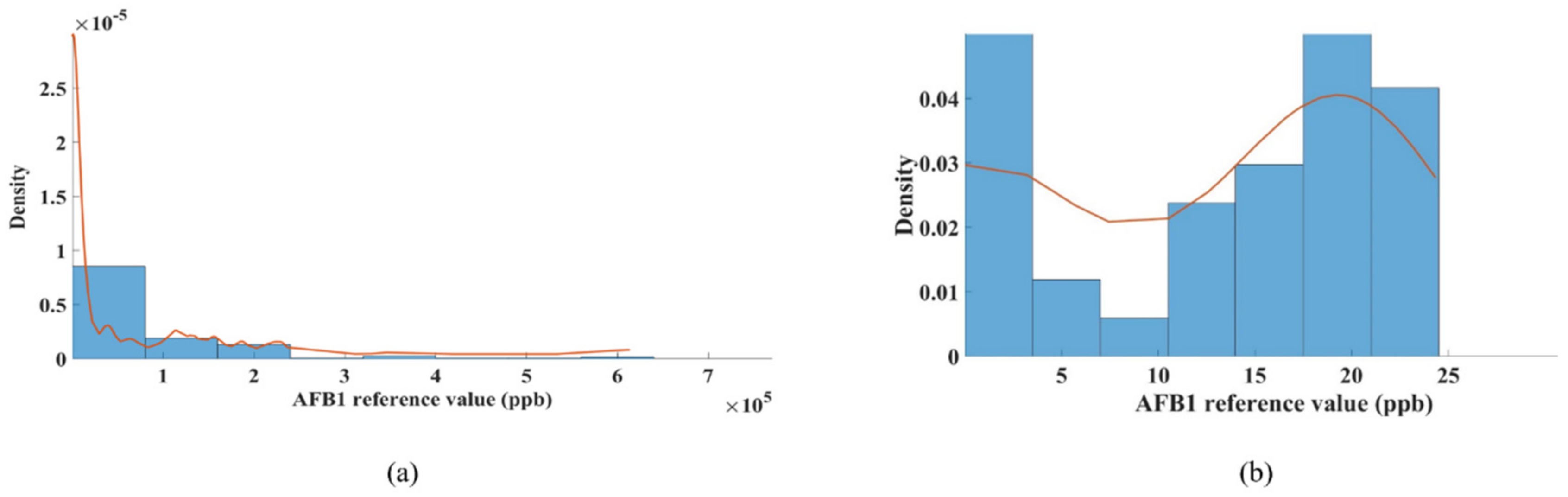
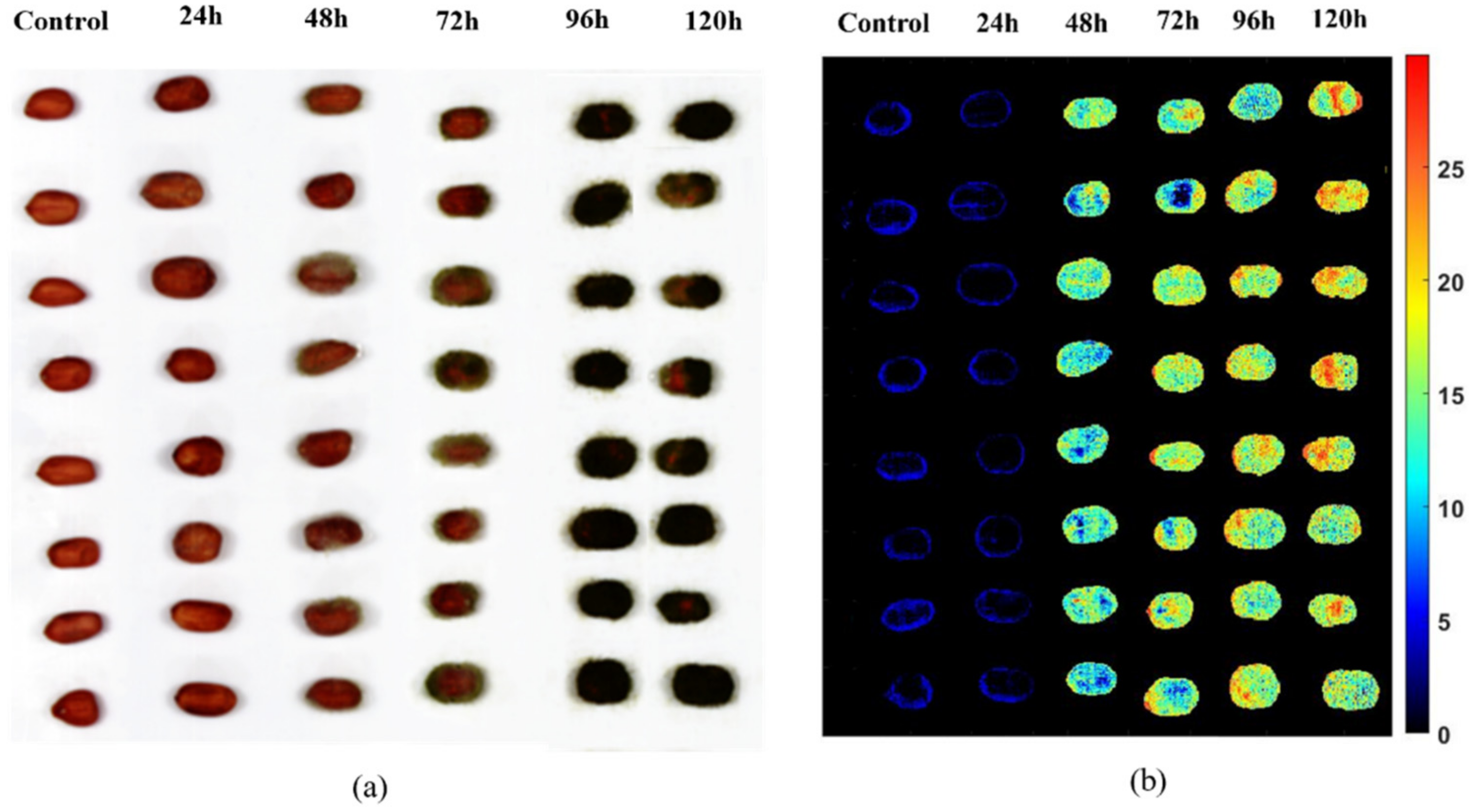
3.1.1. AFB1 Distribution in Peanut Samples
3.1.2. Spectral Information Extraction
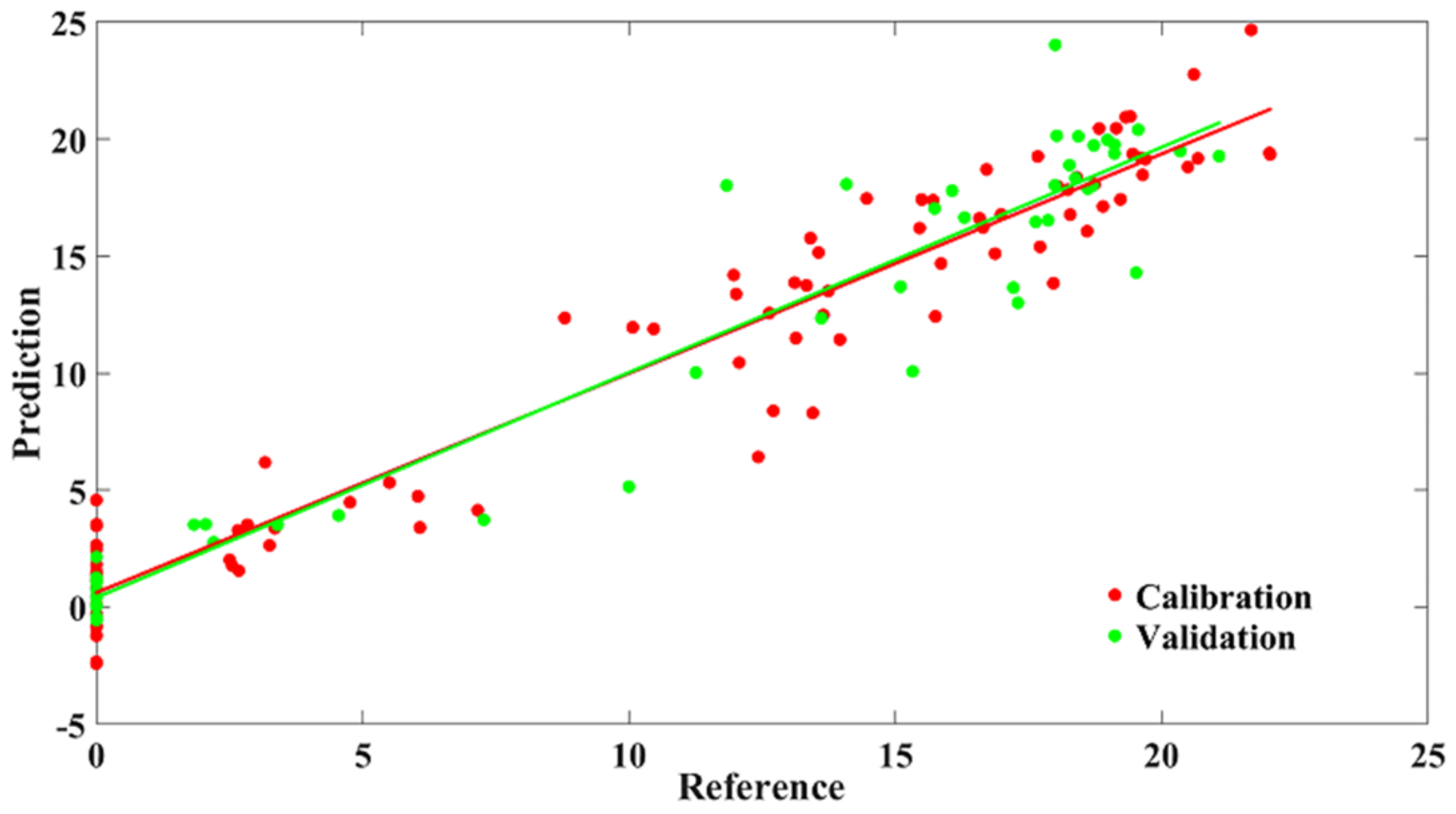
3.1.3. Quantification Analysis Based on Full Wavelengths
3.1.4. Modeling Results Based on Selected Wavelengths
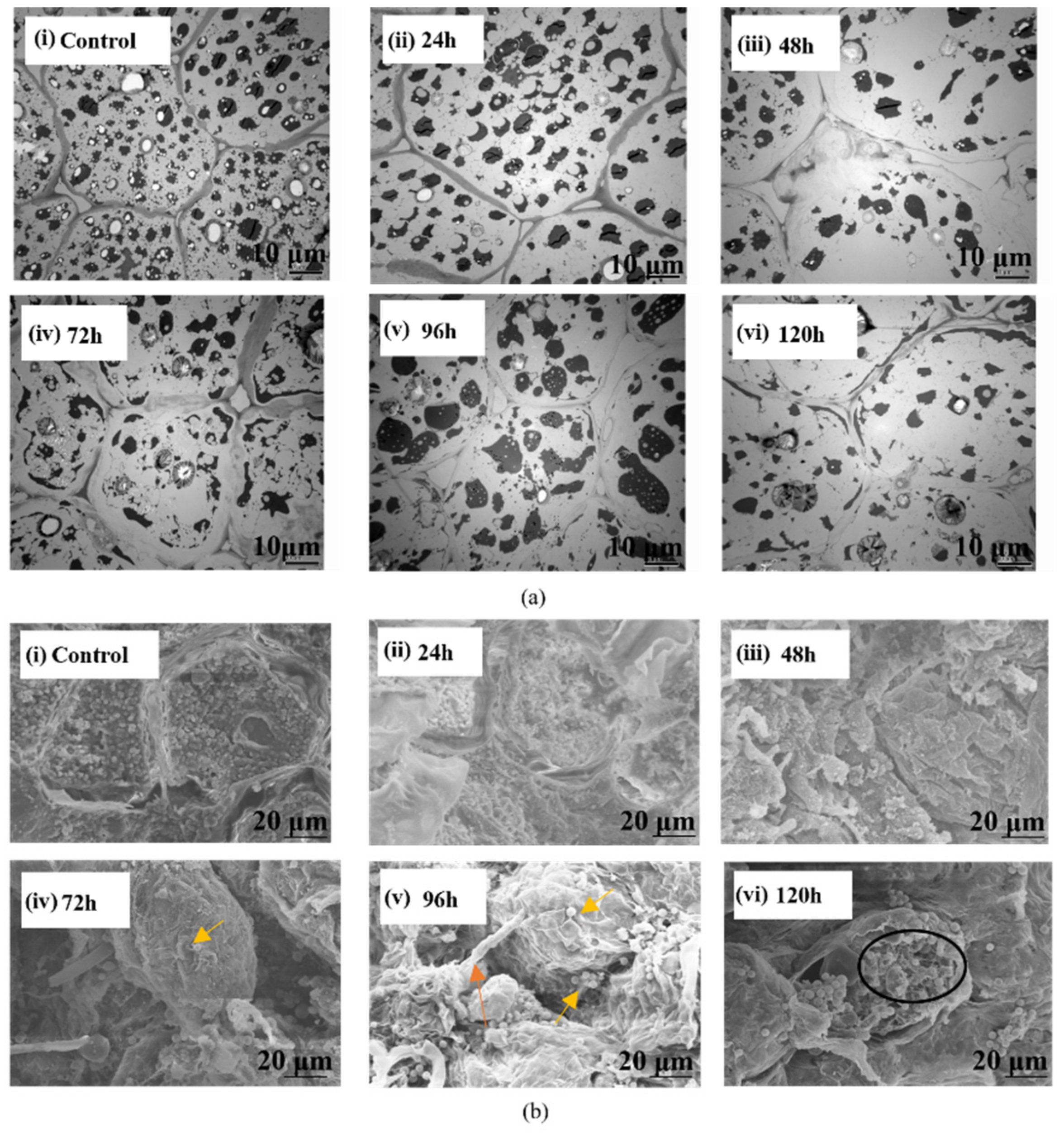
3.2. Micro-Scale Examination of the Complex of A. Flavus and Kernel
3.2.1. SR-FTIR Results and Analysis
3.2.2. Micro-Scale Inter- and Intra-Cellular Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shah, H.U.; Simpson, T.J.; Alam, S.; Khattak, K.F.; Perveen, S. Mould incidence and mycotoxin contamination in maize kernels from Swat Valley, North West Frontier Province of Pakistan. Food Chem. Toxicol. 2010, 48, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, N.J.; Bowers, E.; Hurburgh, C.; Wu, F. Potential economic losses to the US corn industry from aflatoxin contamination. Food Addit. Contam. Part A 2016, 33, 540–550. [Google Scholar] [CrossRef] [PubMed]
- Tao, F.; Yao, H.; Hruska, Z.; Liu, Y.; Rajasekaran, K.; Bhatnagar, D. Use of Visible-Near-Infrared (Vis-NIR) Spectroscopy to Detect Aflatoxin B1 on Peanut Kernels. Appl. Spectrosc. 2019, 73, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Heitschmidt, G.W.; Ni, X.; Windham, W.R.; Hawkins, S.; Chu, X. Identification of aflatoxin B1 on maize kernel surfaces using hyperspectral imaging. Food Control 2014, 42, 78–86. [Google Scholar] [CrossRef]
- Yu, P.; Block, H.; Niu, Z.; Doiron, K. Rapid characterization of molecular chemistry, nutrient make-up and microlocation of internal seed tissue. J. Synchrotron Radiat. 2007, 14, 382–390. [Google Scholar] [CrossRef]
- Kumar Chaudhari, A.; Singh, A.; Kumar Singh, V.; Kumar Dwivedy, A.; Das, S.; Grace Ramsdam, M.; Dkhar, M.S.; Kayang, H.; Kishore Dubey, N. Assessment of chitosan biopolymer encapsulated α-Terpineol against fungal, aflatoxin B1 (AFB1) and free radicals mediated deterioration of stored maize and possible mode of action. Food Chem. 2020, 311, 126010. [Google Scholar] [CrossRef]
- Falsafi, S.R.; Rostamabadi, H.; Assadpour, E.; Jafari, S.M. Morphology and microstructural analysis of bioactive-loaded micro/nanocarriers via microscopy techniques; CLSM/SEM/TEM/AFM. Adv. Colloid Interface Sci. 2020, 280, 102166. [Google Scholar] [CrossRef]
- Zhu, L.-J.; Dogan, H.; Gajula, H.; Gu, M.-H.; Liu, Q.-Q.; Shi, Y.-C. Study of kernel structure of high-amylose and wild-type rice by X-ray microtomography and SEM. J. Cereal Sci. 2012, 55, 1–5. [Google Scholar] [CrossRef]
- Jäger, W. Chapter Two—Advanced and in situ transmission electron microscopy of diamond: A review. In Semiconductors and Semimetals; Nebel, C.E., Aharonovich, I., Mizuochi, N., Hatano, M., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 104, pp. 31–104. [Google Scholar]
- Esmail, S.M.; Omara, R.I.; Abdelaal, K.A.A.; Hafez, Y.M. Histological and biochemical aspects of compatible and incompatible wheat- Puccinia striiformis interactions. Physiol. Mol. Plant Pathol. 2019, 106, 120–128. [Google Scholar] [CrossRef]
- Zhou, X.; Zhong, J.; Dong, J.; Kong, L.; Liu, G.; Tang, Y. The BL01B1 infrared beamline at Shanghai Synchrotron Radiation Facility. Infrared Phys. Technol. 2018, 94, 250–254. [Google Scholar] [CrossRef]
- Wang, W.; Ni, X.; Lawrence, K.C.; Yoon, S.-C.; Heitschmidt, G.W.; Feldner, P. Feasibility of detecting Aflatoxin B1 in single maize kernels using hyperspectral imaging. J. Food Eng. 2015, 166, 182–192. [Google Scholar] [CrossRef]
- Li, H.; Liang, Y.; Xu, Q.; Cao, D. Key wavelengths screening using competitive adaptive reweighted sampling method for multivariate calibration. Anal. Chim. Acta 2009, 648, 77–84. [Google Scholar] [CrossRef]
- Liu, K.; Chen, X.; Li, L.; Chen, H.; Ruan, X.; Liu, W. A consensus successive projections algorithm—Multiple linear regression method for analyzing near infrared spectra. Anal. Chim. Acta 2015, 858, 16–23. [Google Scholar] [CrossRef]
- Guo, Q.; Wu, W.; Massart, D.L.; Boucon, C.; de Jong, S. Feature selection in principal component analysis of analytical data. Chemom. Intell. Lab. Syst. 2002, 61, 123–132. [Google Scholar] [CrossRef]
- Barnes, R.J.; Dhanoa, M.S.; Lister, S.J. Standard Normal Variate Transformation and De-trending of Near-Infrared Diffuse Reflectance Spectra. Appl. Spectrosc. 1989, 43, 772–777. [Google Scholar] [CrossRef]
- Nicolaï, B.M.; Theron, K.I.; Lammertyn, J. Kernel PLS regression on wavelet transformed NIR spectra for prediction of sugar content of apple. Chemom. Intell. Lab. Syst. 2007, 85, 243–252. [Google Scholar] [CrossRef]
- Box, G.E.P.; Cox, D.R. An Analysis of Transformations. J. R. Stat. Soc. Ser. B (Methodol.) 1964, 26, 211–243. [Google Scholar] [CrossRef]
- Jaya, S.; Chari, V.K.; Konda Naganathan, G.; Jeyam, S. Sensing of Moisture Content of In-Shell Peanuts by NIR Reflectance Spectroscopy. J. Sens. Technol. 2012, 2, 1–7. [Google Scholar]
- Sundaram, J.; Kandala, C.V.; Butts, C.L. Application of near infrared spectroscopy to peanut grading and quality analysis: Overview. Sens. Instrum. Food Qual. Saf. 2009, 3, 156–164. [Google Scholar] [CrossRef]
- Nigam, S.; Waliyar, F.; Aruna, R.; Reddy, S.; Kumar, L.; Craufurd, P.; Diallo, A.; Ntare, B.R.; Upadhyaya, H. Breeding Peanut for Resistance to Aflatoxin Contamination at ICRISAT. Peanut Sci. 2009, 36, 42–49. [Google Scholar] [CrossRef]
- Socrates, G. Infrared and Raman characteristic group frequencies table and charts. Int. J. Food Microbiol. 2005, 108, 1–347. [Google Scholar]
- Valasi, L.; Georgiadou, M.; Tarantilis, P.A.; Yanniotis, S.; Pappas, C.S. Rapid screening on aflatoxins’ presence in Pistachia vera nuts using diffuse reflectance infrared Fourier transform spectroscopy and chemometrics. J. Food Sci. Technol. 2021, 58, 356–365. [Google Scholar] [CrossRef]
- Kaya-Celiker, H.; Mallikarjunan, P.K.; Kaaya, A. Mid-infrared spectroscopy for discrimination and classification of Aspergillus spp. contamination in peanuts. Food Control 2015, 52, 103–111. [Google Scholar] [CrossRef]
- Kaya-Celiker, H.; Mallikarjunan, P.K.; Kaaya, A. Characterization of Invasion of Genus Aspergillus on Peanut Seeds Using FTIR-PAS. Food Anal. Methods 2016, 9, 105–113. [Google Scholar] [CrossRef]
- Kos, G.; Lohninger, H.; Krska, R. Fourier transform mid-infrared spectroscopy with attenuated total reflection (FT-IR/ATR) as a tool for the detection of Fusarium fungi on maize. Vib. Spectrosc. 2002, 29, 115–119. [Google Scholar] [CrossRef]
- Bothast, R.J. Fungal deterioration and related phenomena in cereals, legumes and oilseeds. Int. J. Food Microbiol. 2007, 116, 210–243. [Google Scholar]


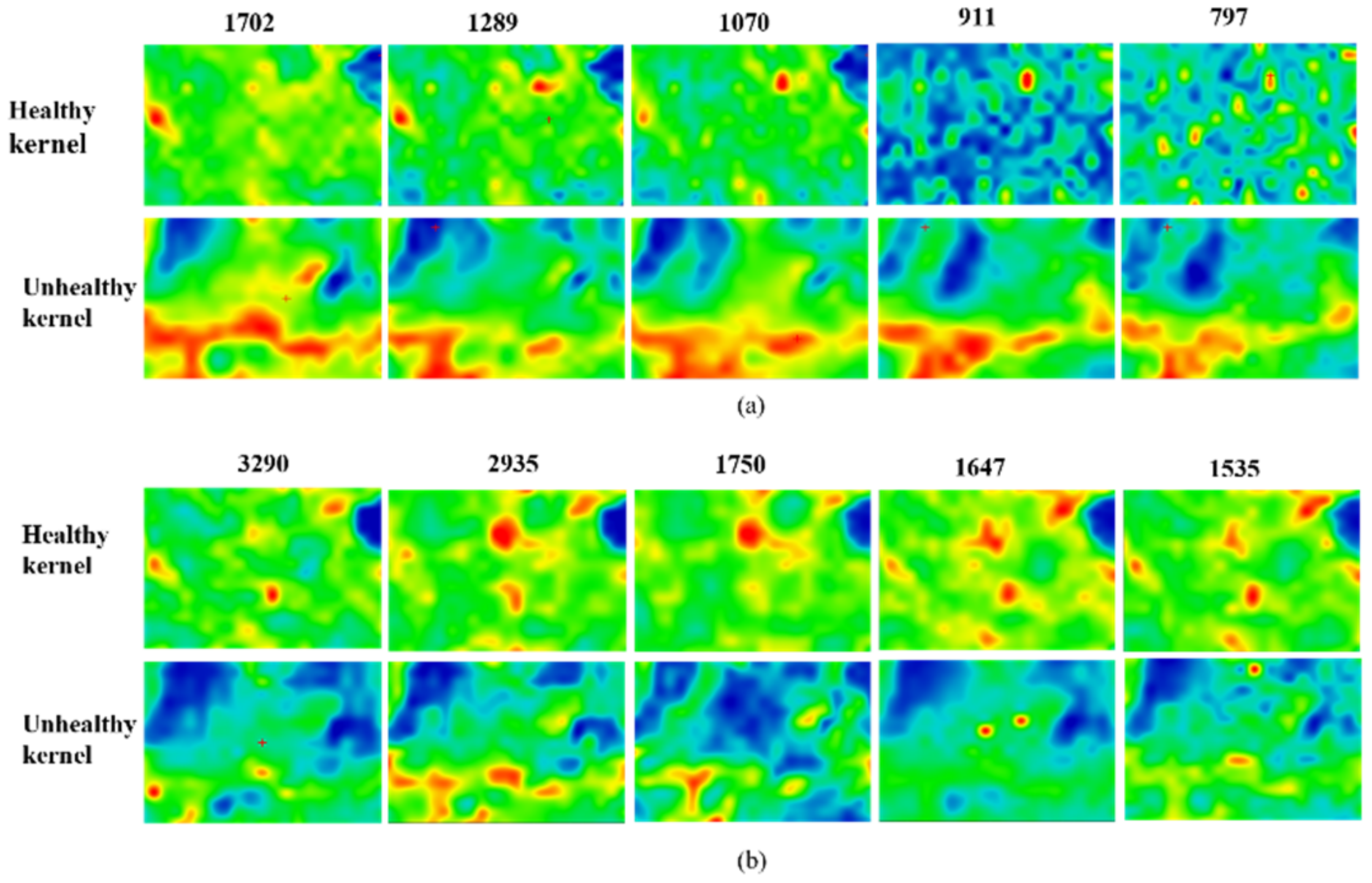


{kind=link}
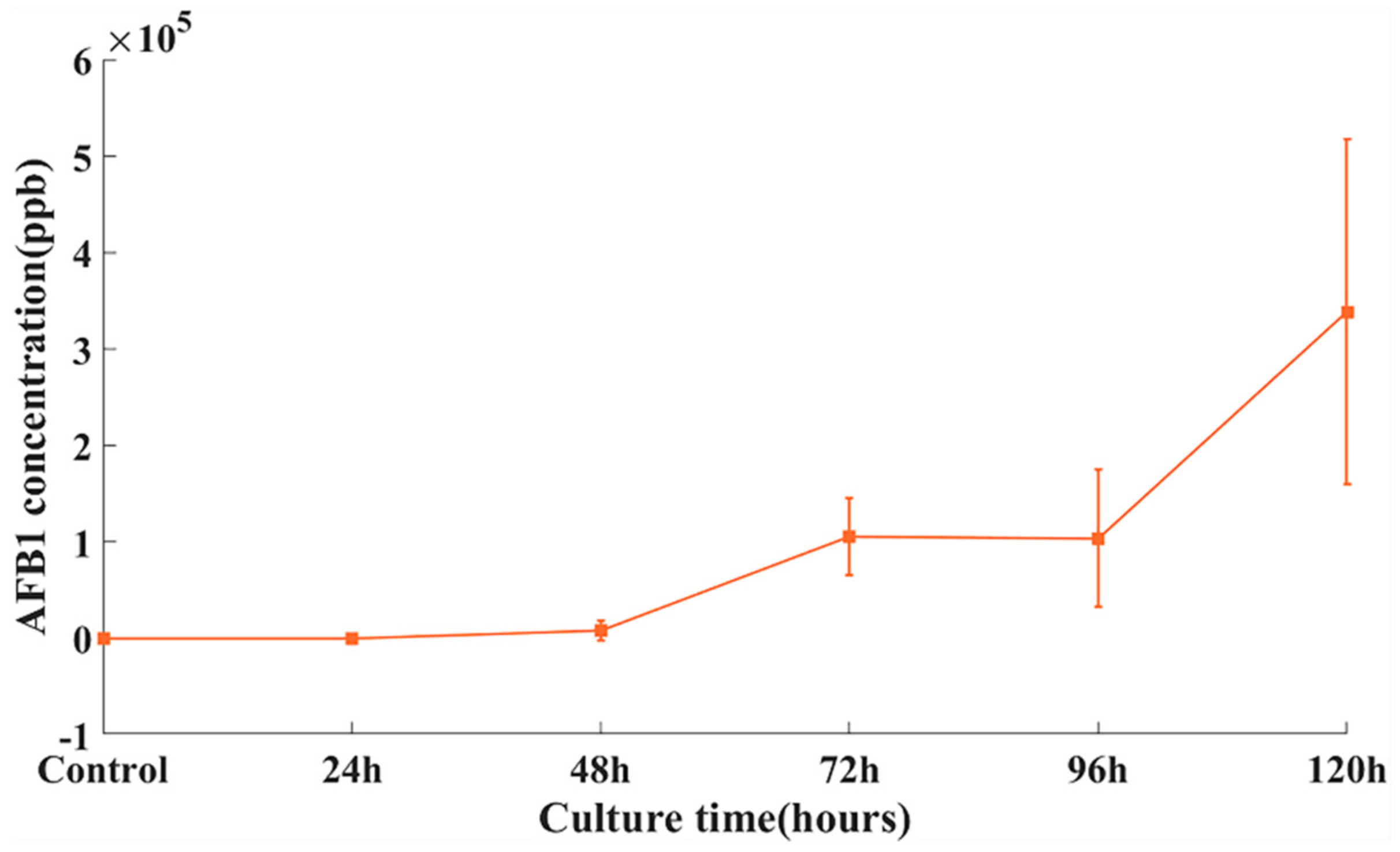
{kind=link}
{kind=link}
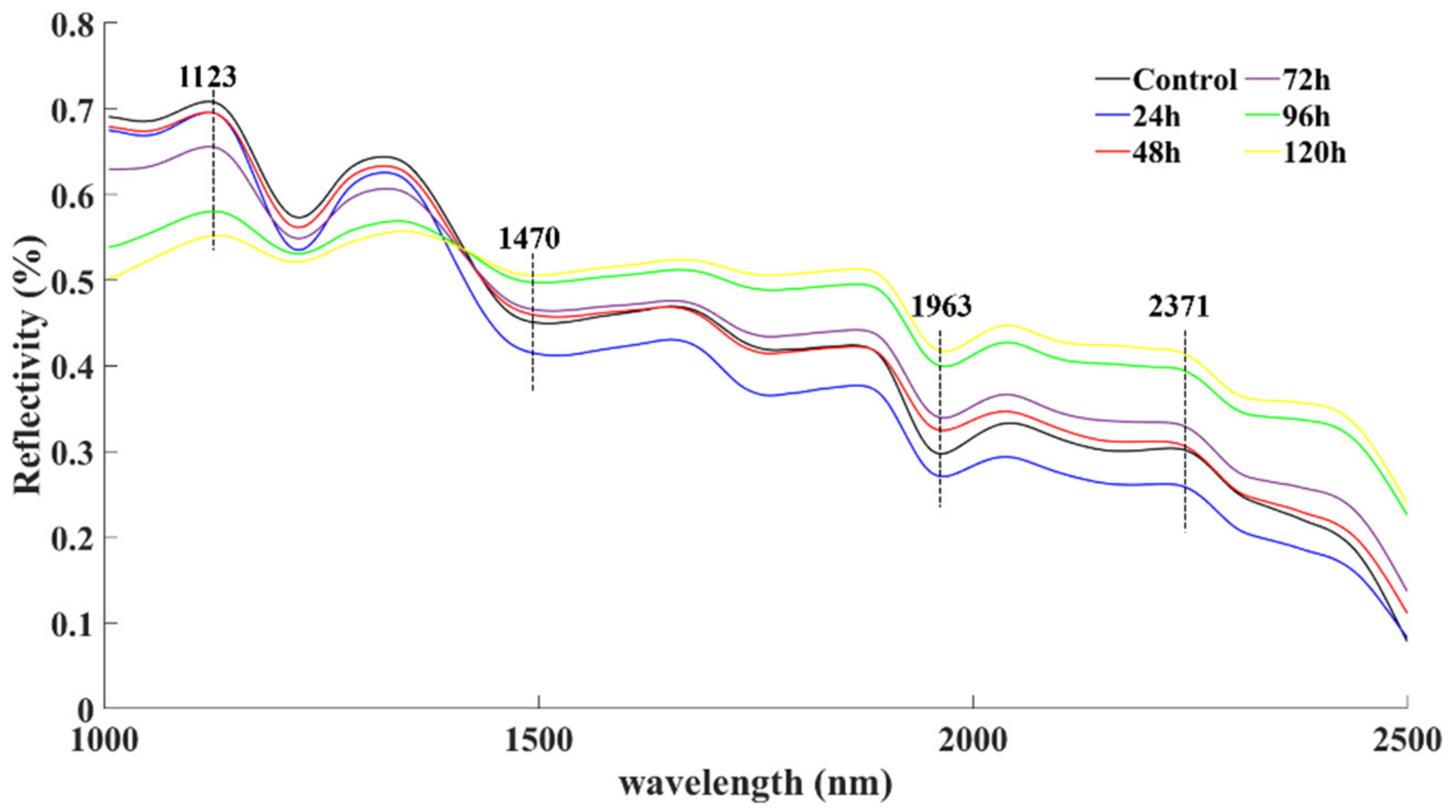
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modeling Method | Pre-Processing | LVs | RMSEc | RMSEv | RPD | ||
|---|---|---|---|---|---|---|---|
| PLSR | Reflectance | 7 | 0.65 | 81424 | 0.39 | 126674 | 1.11 |
| SNV | 9 | 0.69 | 95208 | 0.56 | 86061 | 1.30 | |
| 1st Deriv. | 5 | 0.59 | 97577 | 0.35 | 99381 | 1.05 | |
| 2nd Deriv. | 5 | 0.68 | 87665 | 0.42 | 121903 | 1.20 | |
| SVR | Reflectance | \ | 0.54 | 109734 | 0.15 | 147624 | 0.54 |
| SNV | \ | 0.52 | 119822 | 0.74 | 28938 | 1.84 | |
| 1st Deriv. | \ | 0.53 | 113386 | 0.52 | 80050 | 0.89 | |
| 2nd Deriv. | \ | 0.86 | 59764 | 0.34 | 111908 | 1.02 |
| Modeling Method | Pre-Processing | LVs | RMSEc | RMSEv | RPD | ||
|---|---|---|---|---|---|---|---|
| PLSR | Origin | 9 | 0.96 | 1.751 | 0.85 | 3.582 | 2.61 |
| SNV | 7 | 0.93 | 2.433 | 0.89 | 2.650 | 3.32 | |
| 1st Deriv. | 8 | 0.95 | 2.055 | 0.87 | 2.856 | 2.48 | |
| 2nd Deriv. | 10 | 0.97 | 1.658 | 0.87 | 2.957 | 2.63 | |
| SVR | Origin | \ | 0.96 | 1.772 | 0.72 | 4.916 | 1.85 |
| SNV | \ | 0.95 | 2.044 | 0.88 | 3.205 | 2.85 | |
| 1st Deriv. | \ | 0.97 | 1.375 | 0.91 | 2.265 | 3.24 | |
| 2nd Deriv. | \ | 0.99 | 0.409 | 0.89 | 2.798 | 2.74 |
| Method | Number | Wavelength (nm) |
|---|---|---|
| SPA | 7 | 1173, 1580, 1918, 2007, 2338, 2405, 2471 |
| CARS | 10 | 1547, 1630, 1680, 1791, 2067, 2123, 2184, 2316, 2361, 2405 |
| WCS | 8 | 1117, 1307, 1530, 1763, 1946, 2029, 2161, 2422 |
| Modeling Method | Variable Selection Method | LVs | RMSEc | RMSEv | RPD | ||
|---|---|---|---|---|---|---|---|
| PLSR | SPA | 5 | 0.91 | 2.914 | 0.90 | 2.740 | 2.75 |
| CARS | 6 | 0.91 | 2.483 | 0.90 | 3.012 | 3.14 | |
| WCS | 6 | 0.81 | 3.446 | 0.79 | 3.842 | 1.70 | |
| SVR | SPA | \ | 0.89 | 3.187 | 0.87 | 2.980 | 2.32 |
| CARS | \ | 0.95 | 1.950 | 0.93 | 2.637 | 3.32 | |
| WCS | 0.91 | 2.337 | 0.88 | 2.936 | 2.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Jia, B.; Lu, Y.; Yoon, S.-C.; Ni, X.; Zhuang, H.; Guo, X.; Le, W.; Wang, W. Detection of Aflatoxin B1 in Single Peanut Kernels by Combining Hyperspectral and Microscopic Imaging Technologies. Sensors 2022, 22, 4864. https://doi.org/10.3390/s22134864
Zhang H, Jia B, Lu Y, Yoon S-C, Ni X, Zhuang H, Guo X, Le W, Wang W. Detection of Aflatoxin B1 in Single Peanut Kernels by Combining Hyperspectral and Microscopic Imaging Technologies. Sensors. 2022; 22(13):4864. https://doi.org/10.3390/s22134864
Chicago/Turabian StyleZhang, Haicheng, Beibei Jia, Yao Lu, Seung-Chul Yoon, Xinzhi Ni, Hong Zhuang, Xiaohuan Guo, Wenxin Le, and Wei Wang. 2022. "Detection of Aflatoxin B1 in Single Peanut Kernels by Combining Hyperspectral and Microscopic Imaging Technologies" Sensors 22, no. 13: 4864. https://doi.org/10.3390/s22134864
APA StyleZhang, H., Jia, B., Lu, Y., Yoon, S.-C., Ni, X., Zhuang, H., Guo, X., Le, W., & Wang, W. (2022). Detection of Aflatoxin B1 in Single Peanut Kernels by Combining Hyperspectral and Microscopic Imaging Technologies. Sensors, 22(13), 4864. https://doi.org/10.3390/s22134864