Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities


Abstract
:1. Introduction
2. Results
2.1. HPCPC Purification
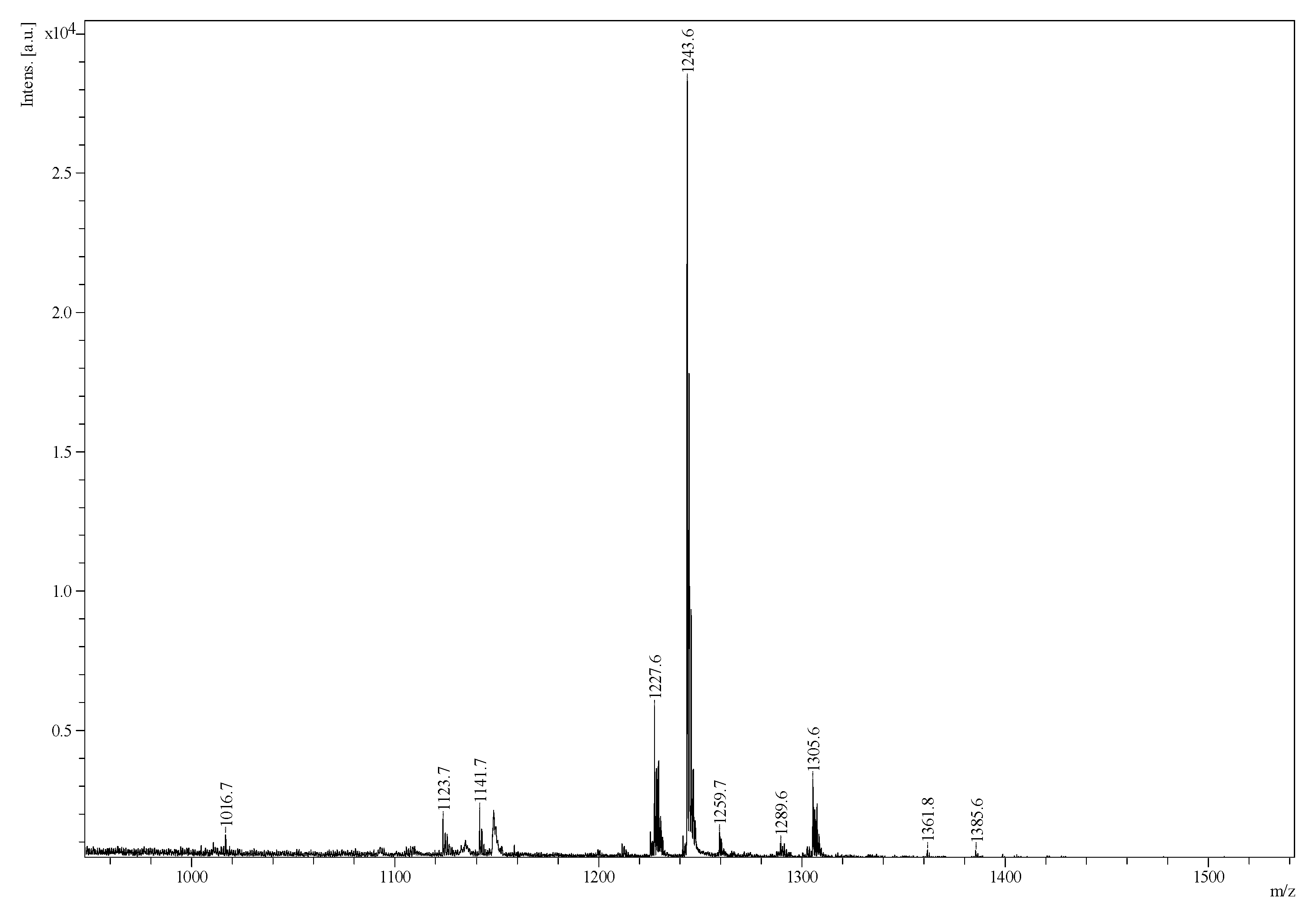
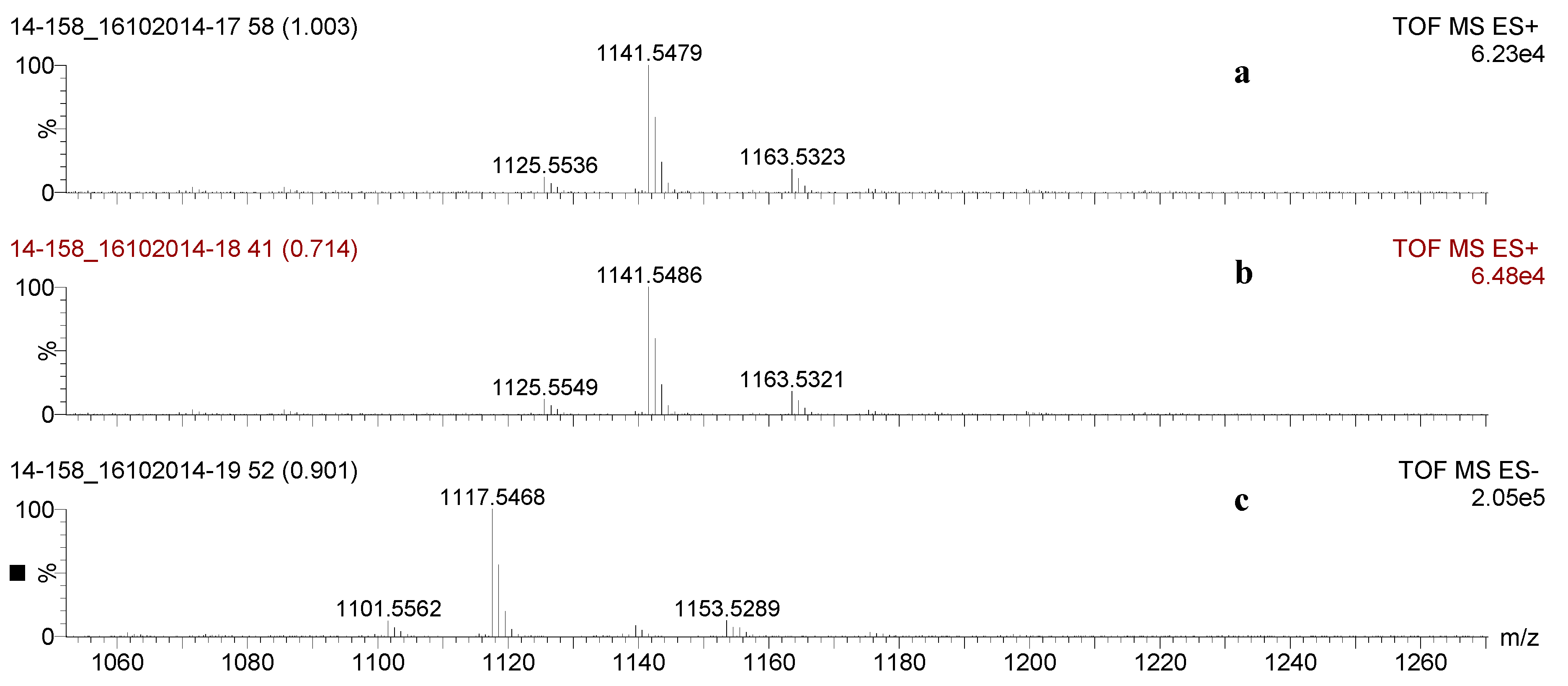
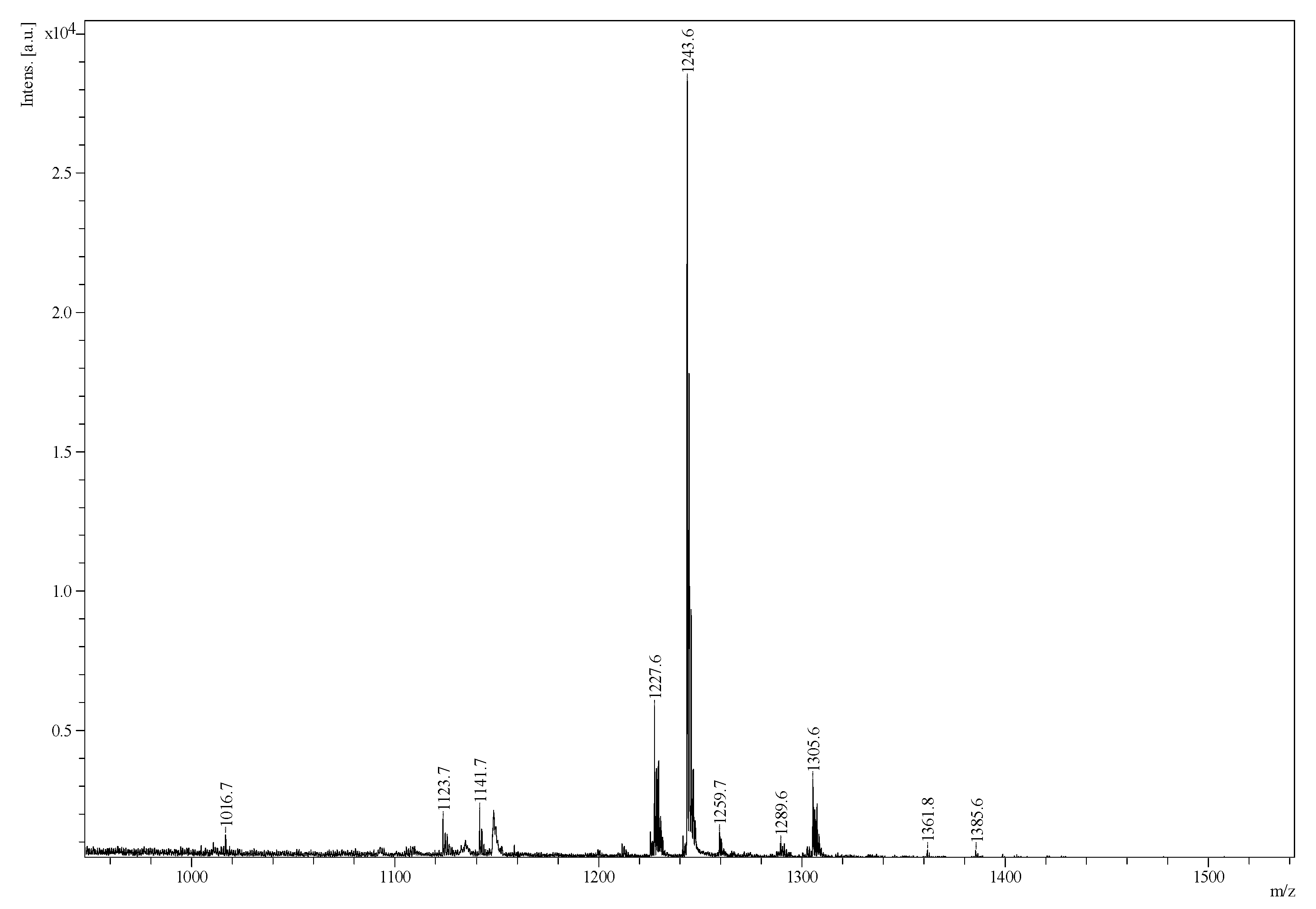
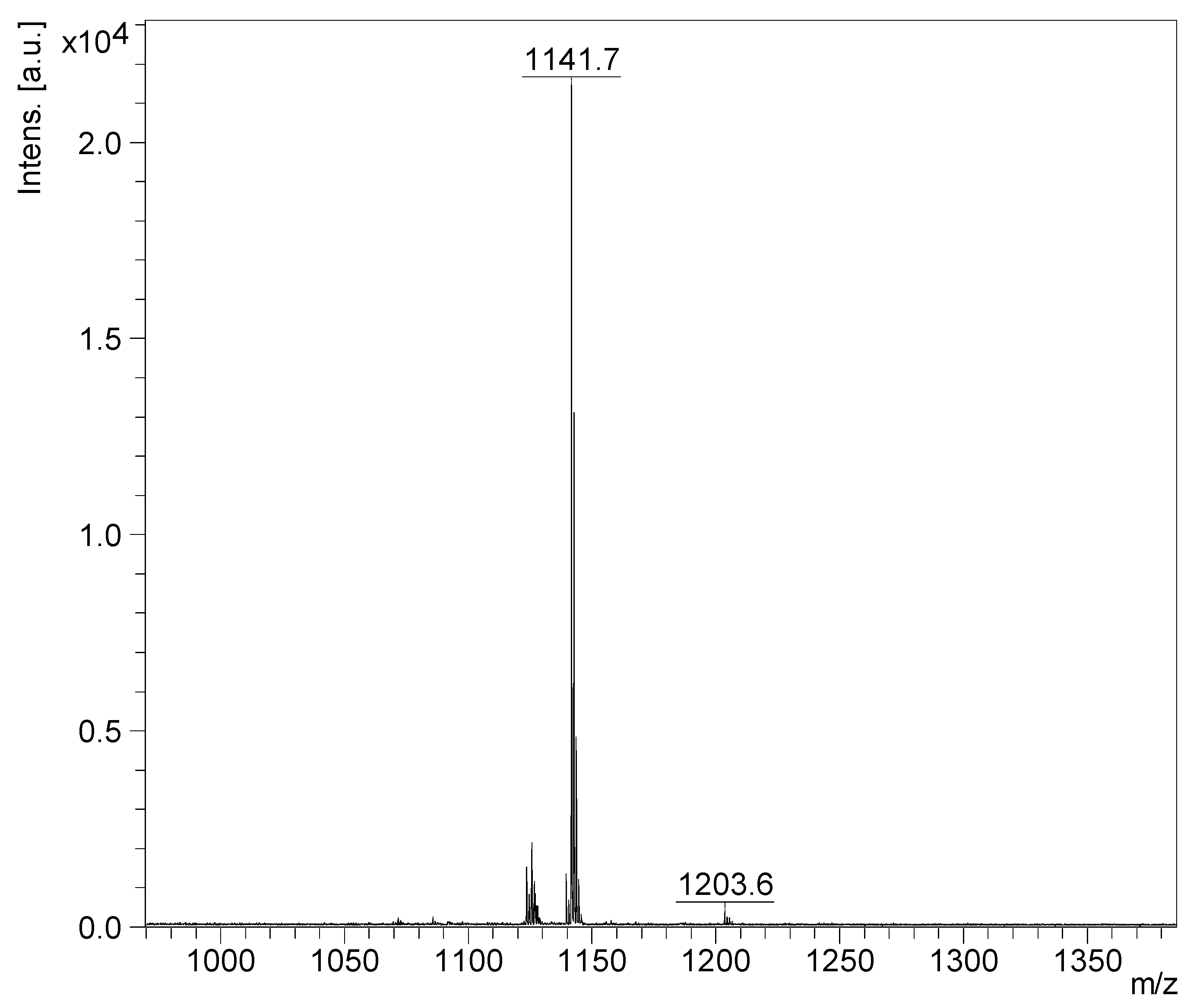
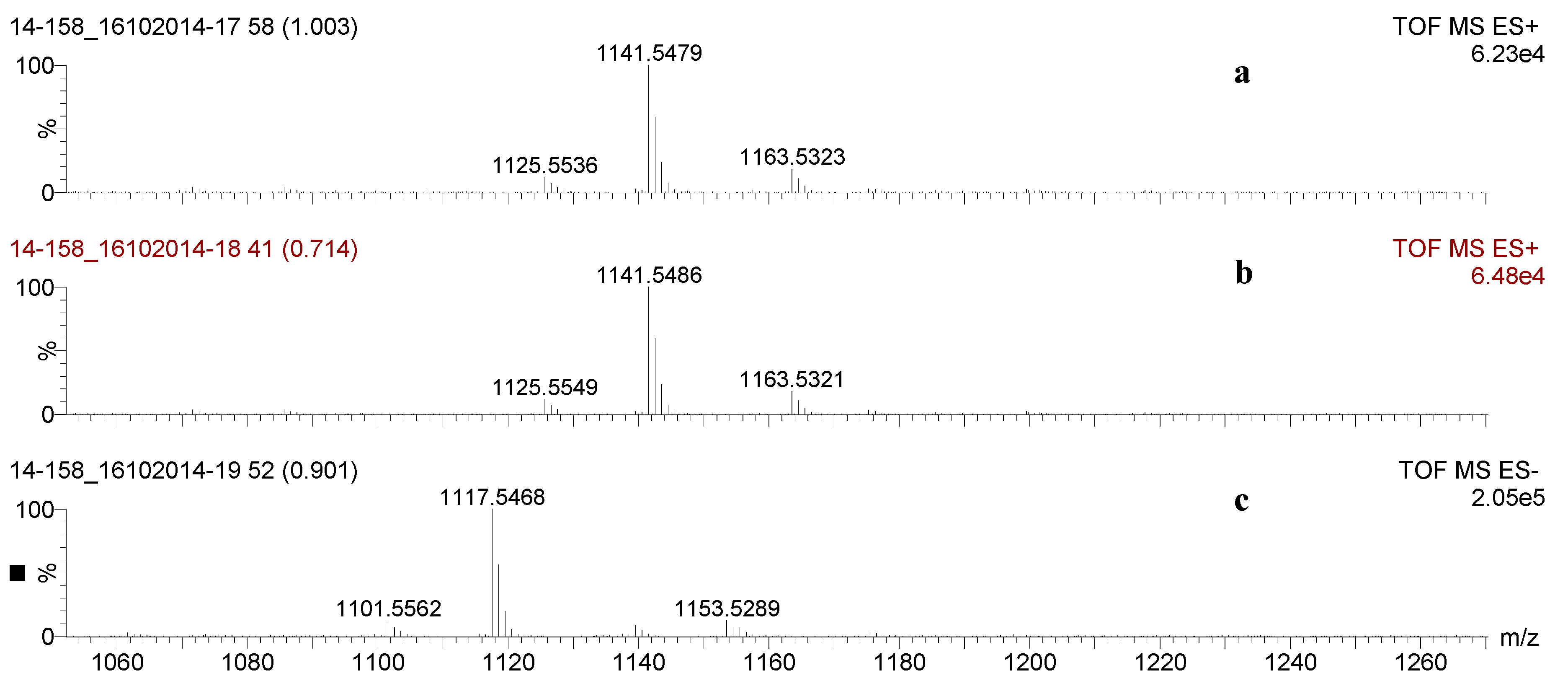
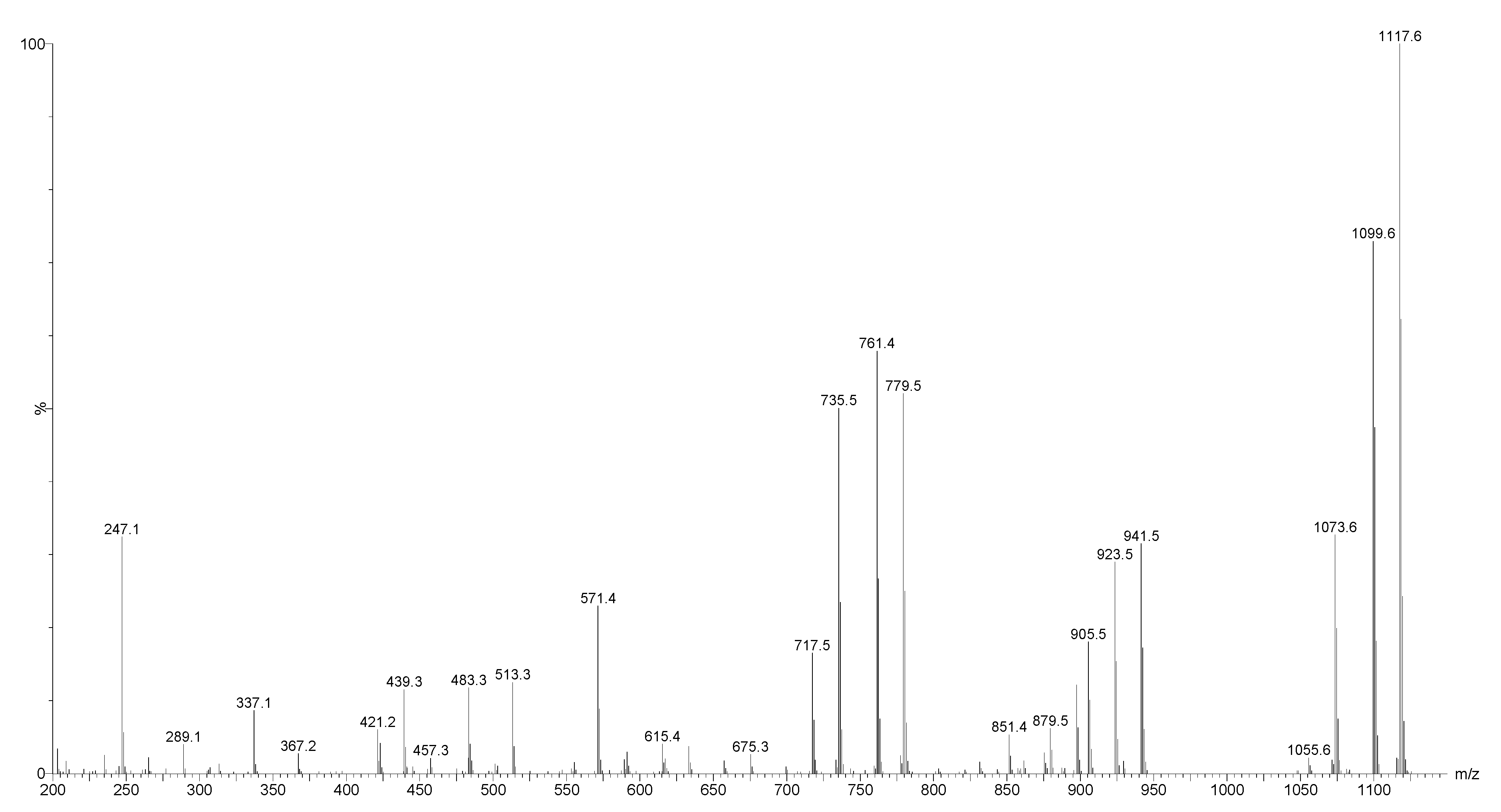
2.2. Mass Spectrometry Analysis of Saponins
MALDI-MS and ESI-MS Analyses of Saponins from the Body Wall of H. lessoni
2.3. Saponin Profiles by Negative-Ion ESI-MS
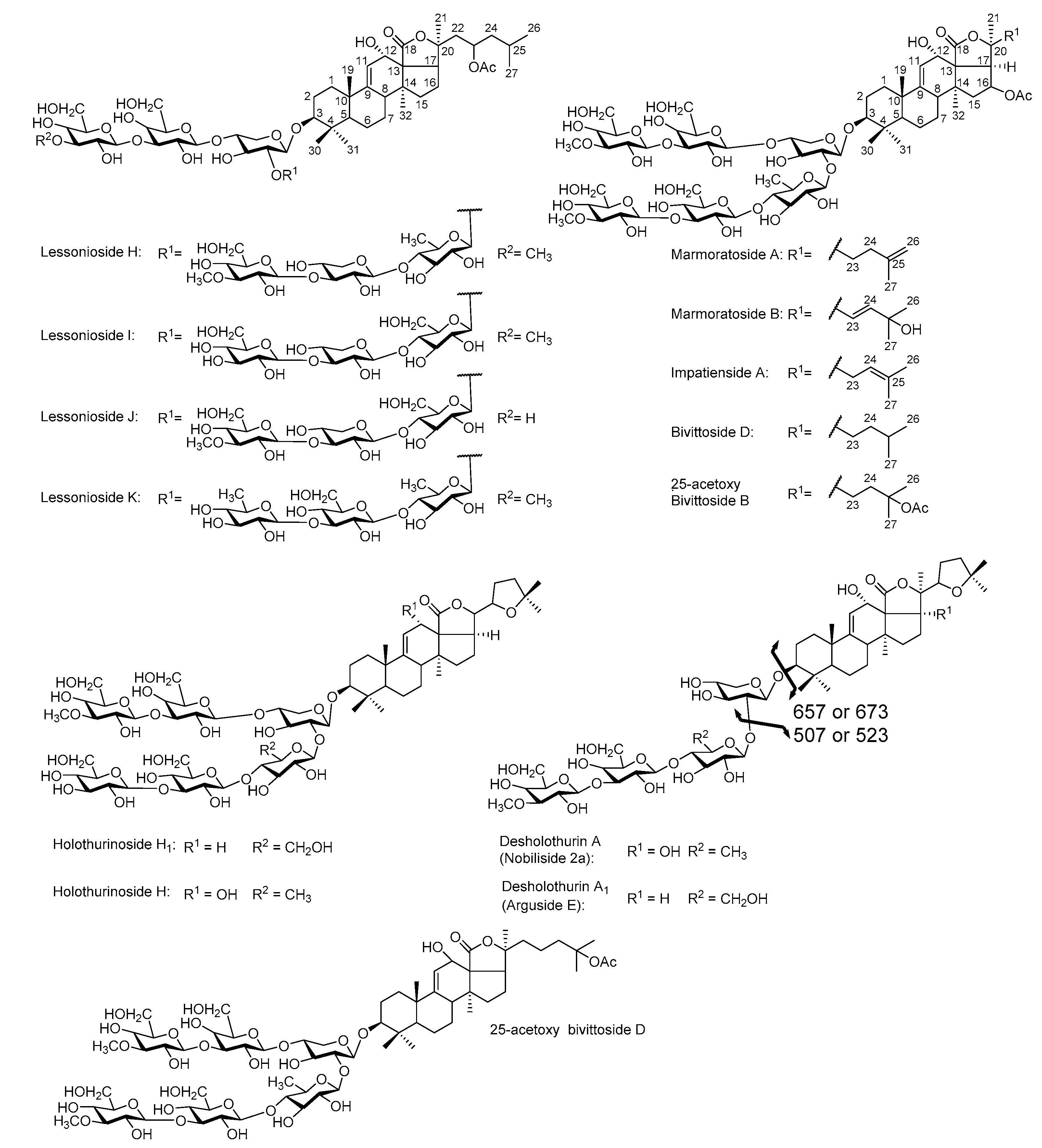
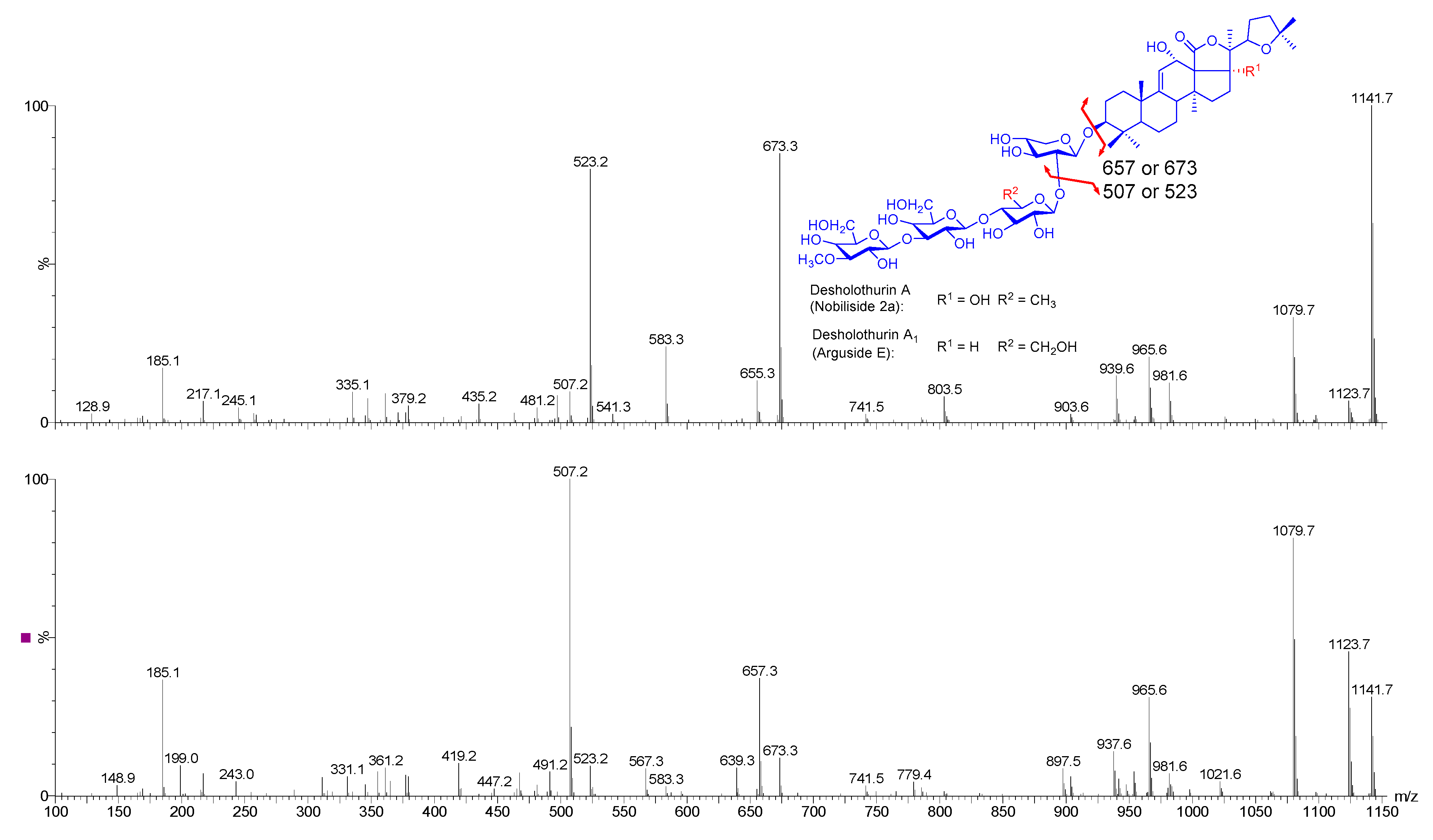
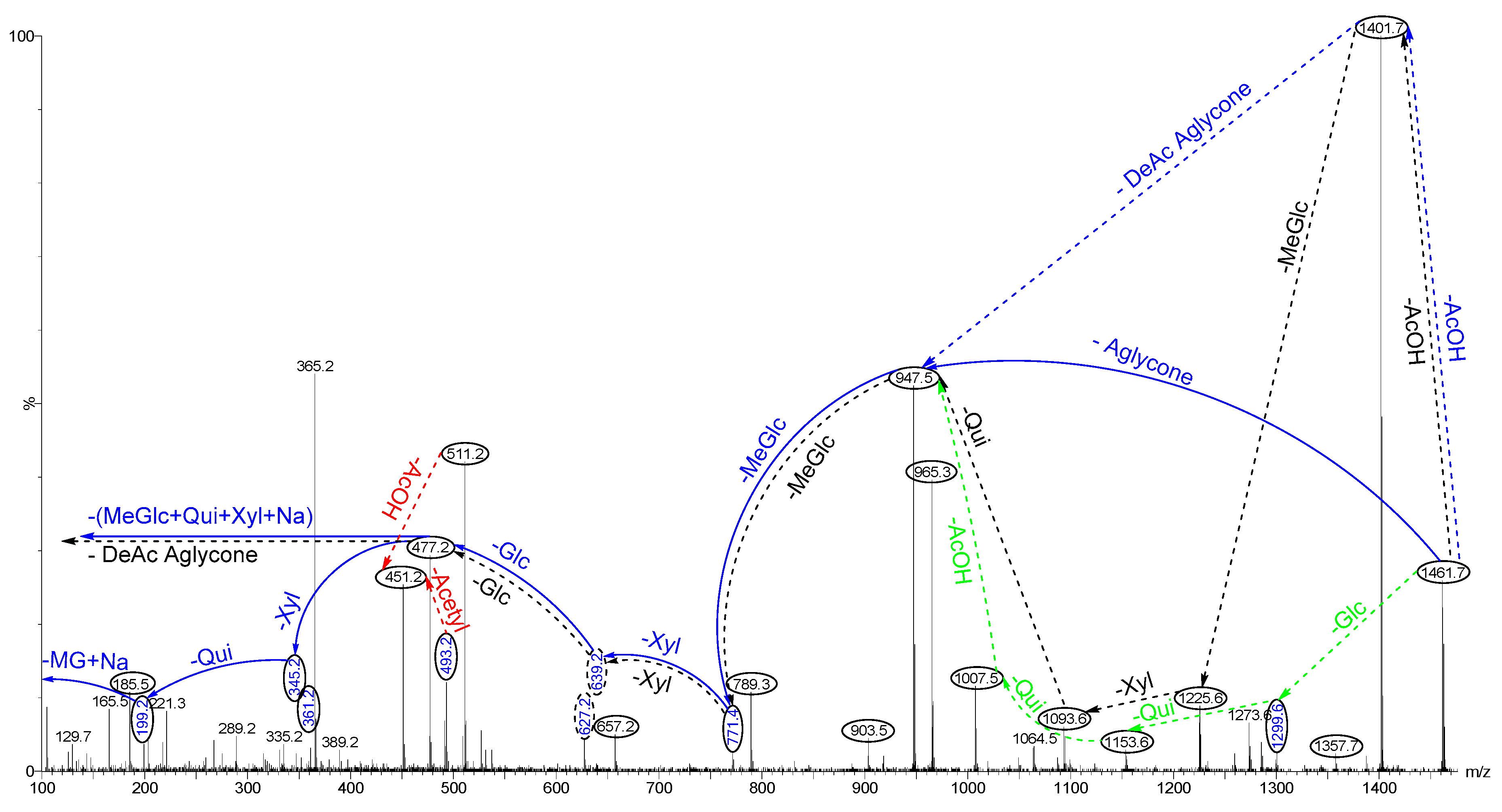
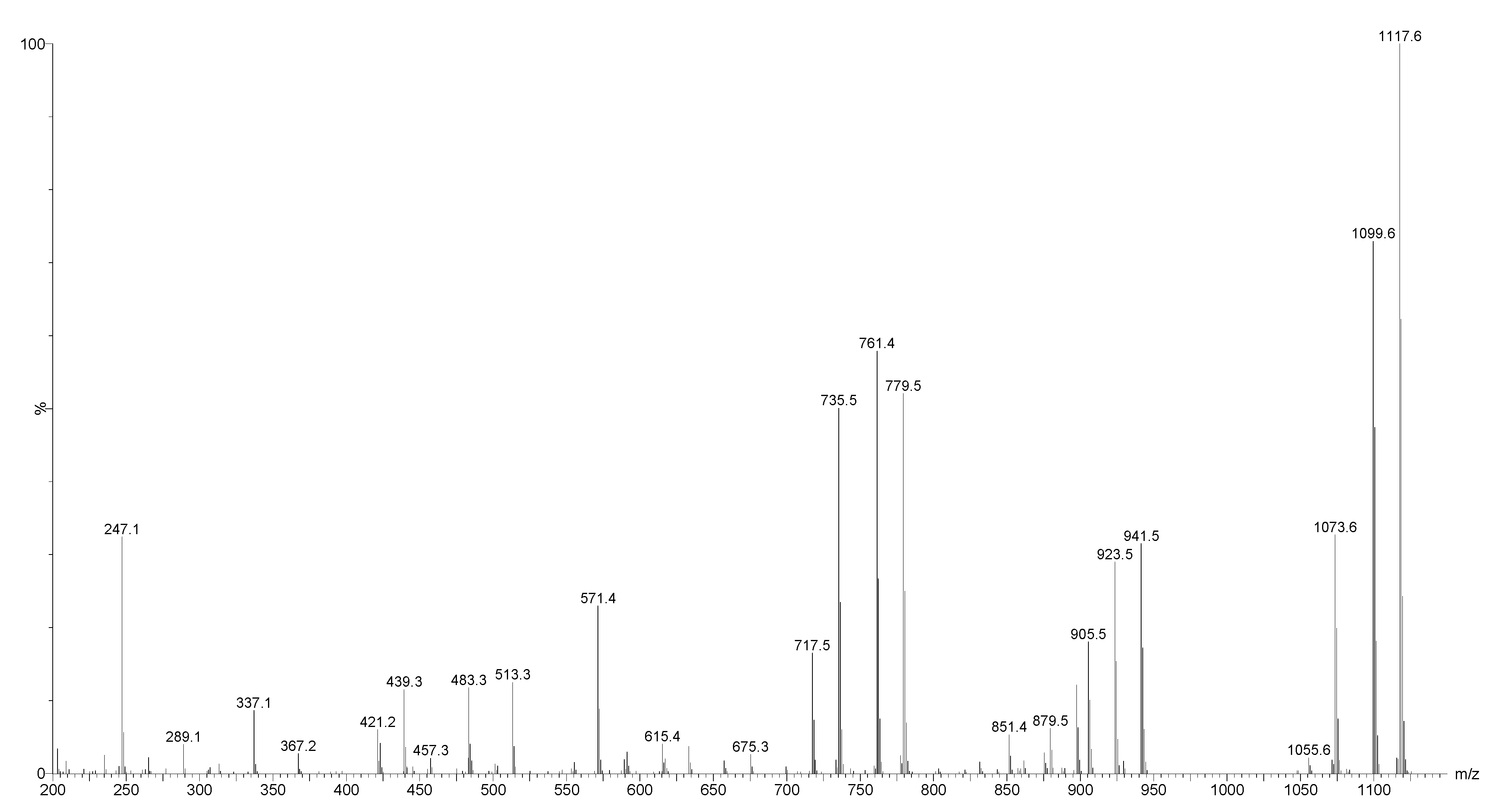
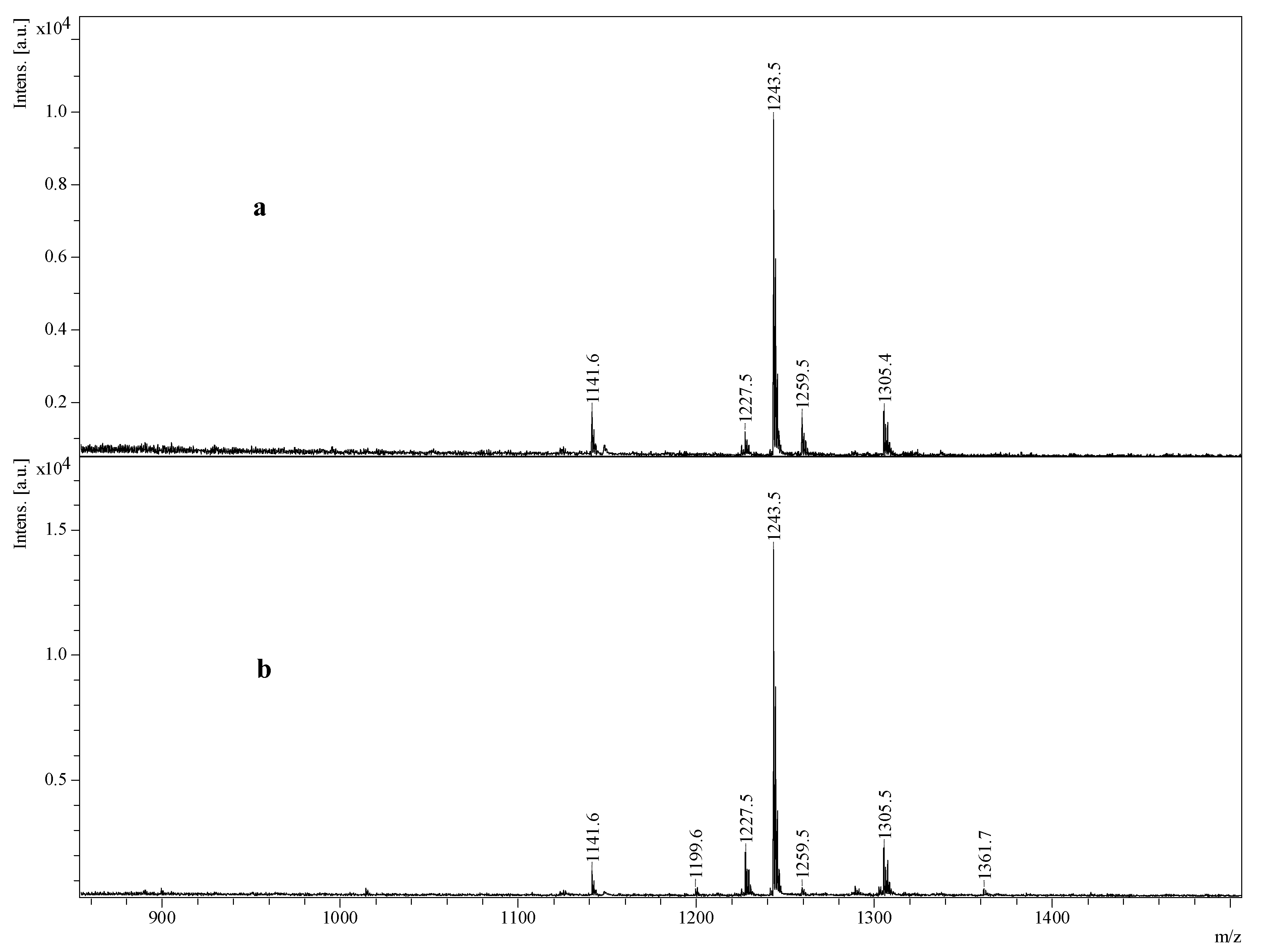
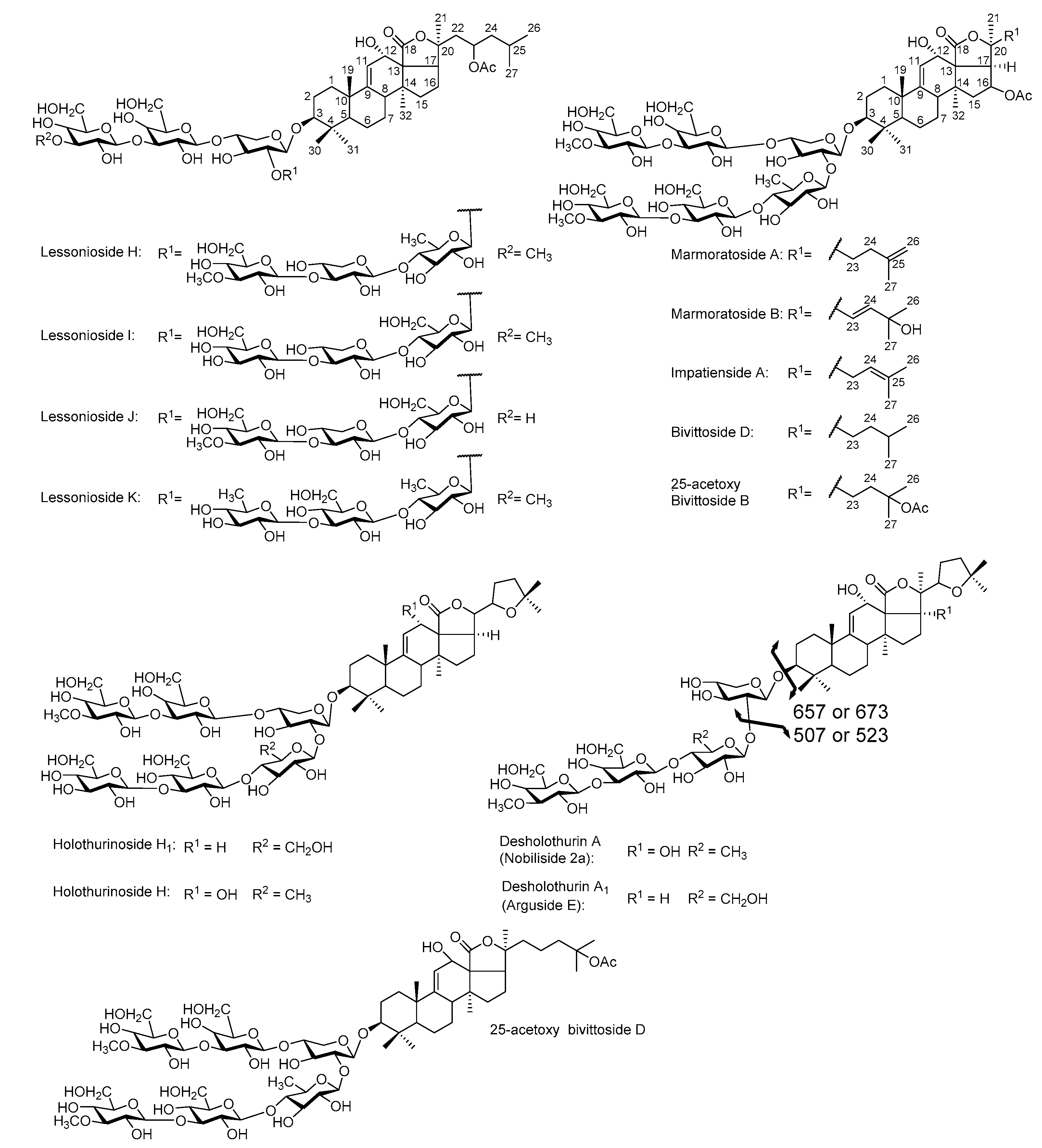
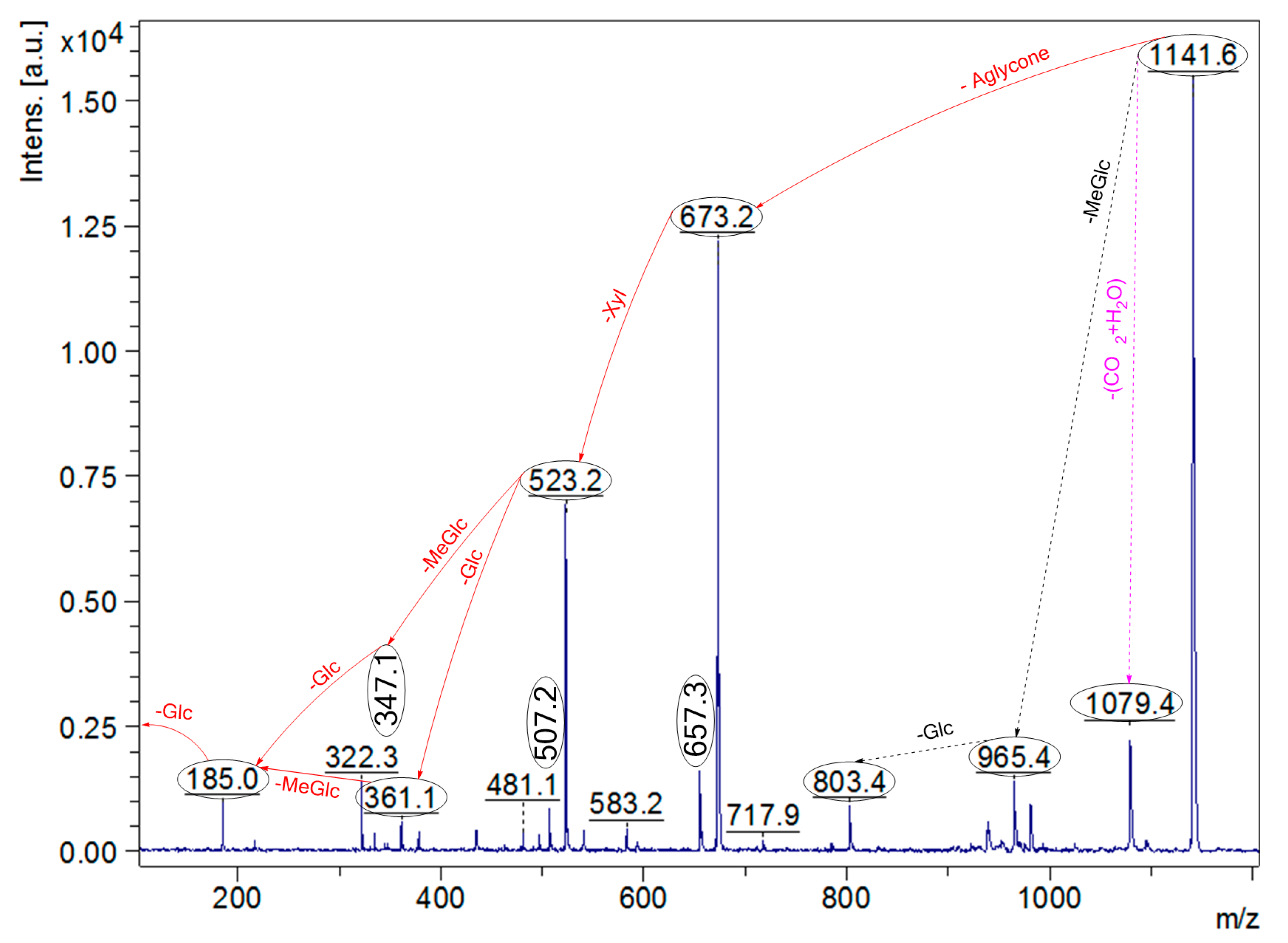
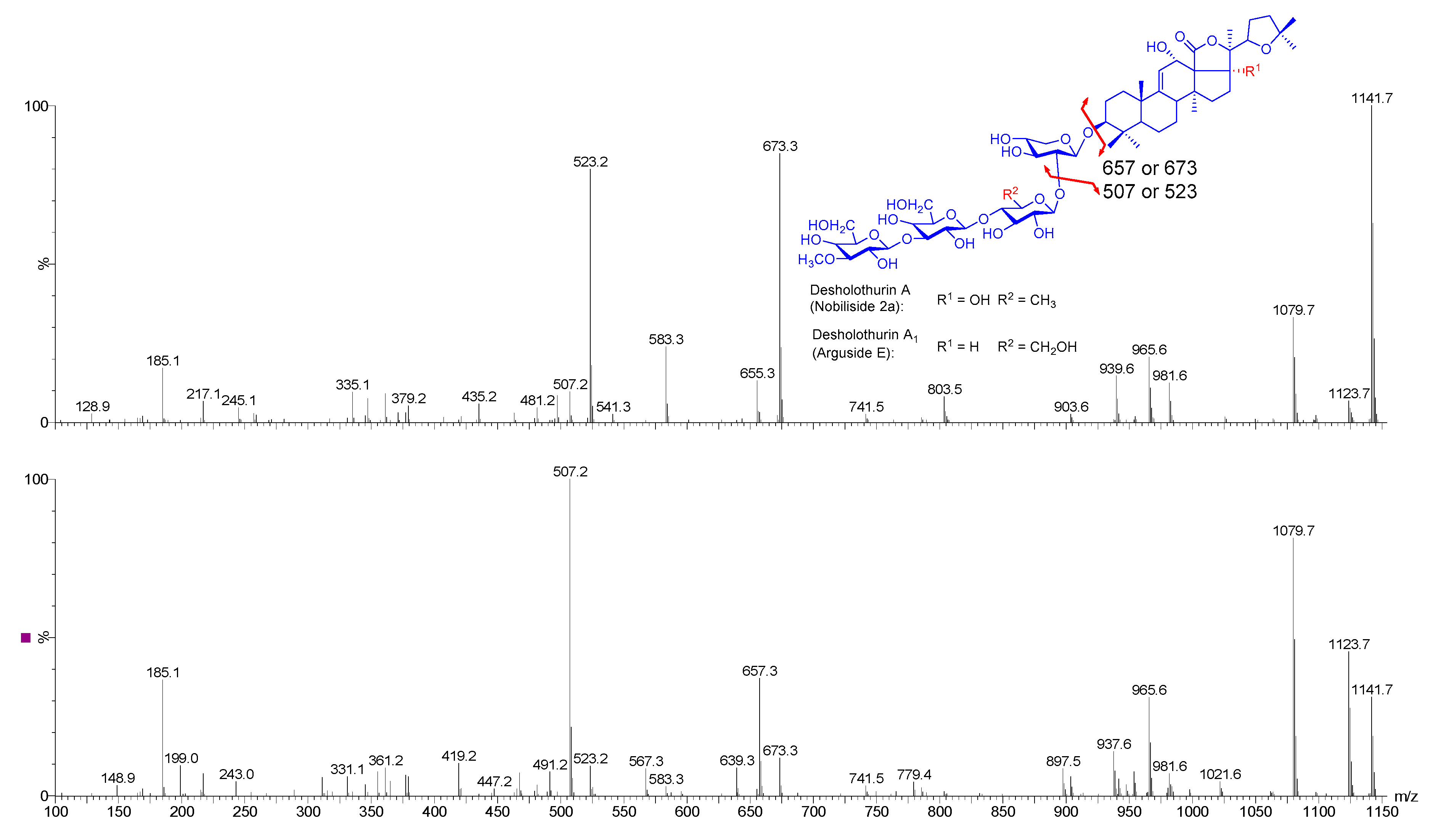
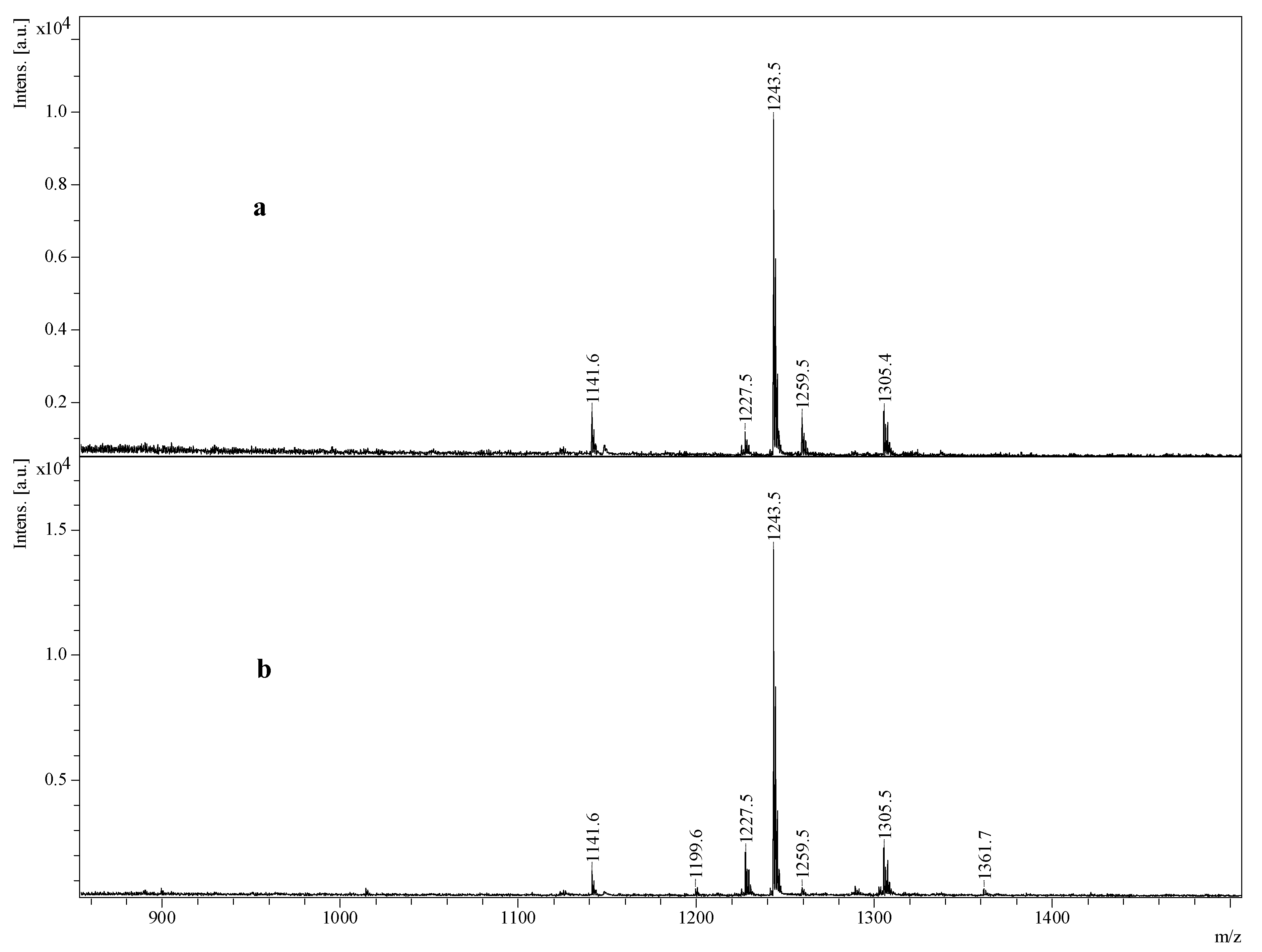
2.4. Structure Elucidation of Saponins by Tandem Mass Spectrometry Analysis
2.5. Structural Determination of Saponins by MALDI MS/MS
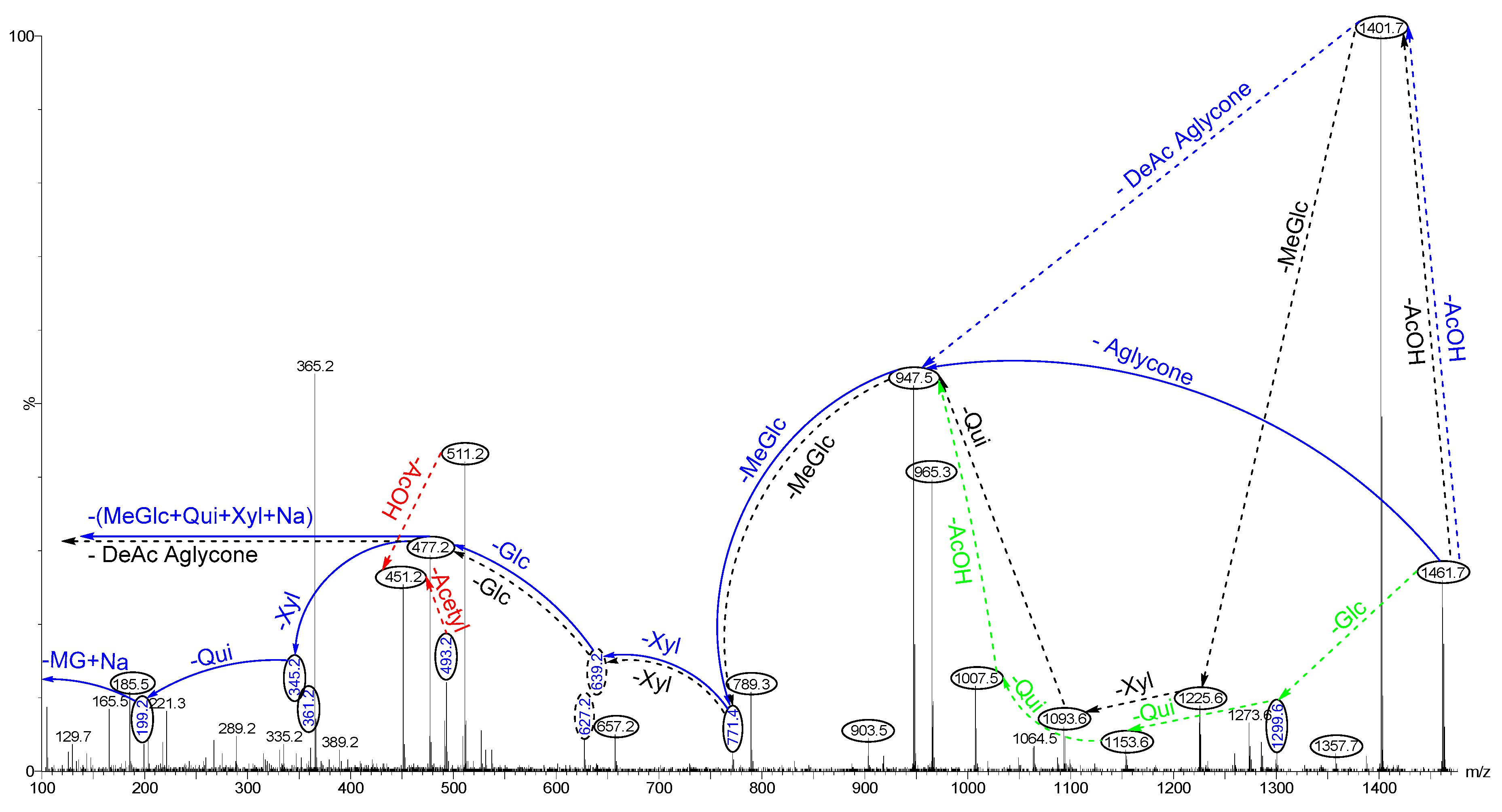
2.5.1. Chemical Analysis of Saponins by ESI-MS/MS
2.5.2. Negative Ion Mode ESI-MS/MS
2.6. Common Saponins between the Viscera and Body Wall
2.6.1. Unique Saponins in the Body Wall
2.6.2. Distribution of Saponin (Body Wall vs. Viscera)
2.7. Bioactivity of Sea Cucumber Fractions and Saponins
2.7.1. Antifungal and Antibacterial Activities of Purified Saponins
2.7.2. Anti-Oxidant Activity of Sea Cucumber Extracts
3. Materials and Methods
3.1. Sea Cucumber Sample
3.2. Chemicals
3.3. Extraction and Purification Protocols
3.4. Purification of the Extract
3.5. High-Performance Centrifugal Partition Chromatography (HPCPC or CPC)
3.6. Thin Layer Chromatography (TLC)
3.7. Mass Spectrometry
3.7.1. MALDI
3.7.2. ESI MS
3.8. Antifungal Activity Assay (Plug Type Diffusion Assay)
3.9. Antibacterial Activity Assay
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Purcell, S.W.; Samyn, Y.; Conand, C. Commercially Important Sea Cucumbers of the World; FAO Species Catalogue for Fishery Purposes No. 6; FAO: Rome, Italy, 2012; p. 150. [Google Scholar]
- Bahrami, Y.; Zhang, W.; Franco, C. Discovery of novel saponins from the viscera of the sea cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 2633–2667. [Google Scholar] [CrossRef] [PubMed]
- Purcell, S.W. Value, market preferences and trade of beche-de-mer from Pacific Island sea cucumbers. PLoS ONE 2014, 9, e95075. [Google Scholar] [CrossRef] [PubMed]
- Pangestuti, R.; Arifin, Z. Medicinal and health benefit effects of functional sea cucumbers. J. Tradit. Complement. Med. 2018, 8, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y. Discovery of Novel Saponins as Potential Future Drugs from Sea Cucumber Viscera; Flinders University: Adelaide, Australia, 2015. [Google Scholar]
- Bahrami, Y.; Franco, M.M.C. Structure elucidation of new acetylated saponins, Lessoniosides A, B, C, D, and E, and non-acetylated saponins, Lessoniosides F and G, from the viscera of the sea cucumber Holothuria lessoni. Mar. Drugs 2015, 13, 597–617. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Franco, C.M.M. Acetylated triterpene glycosides and their biological activity from holothuroidea reported in the past six decades. Mar. Drugs 2016, 14, 147. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, M.; Wisztorski, M.; Decroo, C.; De Winter, J.; Caulier, G.; Hennebert, E.; Eeckhaut, I.; Fournier, I.; Flammang, P.; Gerbaux, P. Inter- and intra-organ spatial distributions of sea star saponins by MALDI imaging. Anal. Bioanal. Chem. 2015. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.-C.; Xue, C.-H.; Zhang, T.-T.; Wang, Y.-M. Saponins from sea cucumber and their biological activities. J. Agric. Food Chem. 2018, 66, 7222–7237. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Kalinin, V.I.; Andrijaschenko, P.V.; Dmitrenok, P.S.; Chingizova, E.A.; Ermakova, S.P.; Malyarenko, O.S.; Dautova, T.N. Nine new triterpene glycosides, magnumosides A1–A4, B1, B2, C1, C2 and C4, from the vietnamese sea cucumber Neothyonidium (=Massinium) magnum: Structures and activities against tumor cells independently and in synergy with radioactive irradiation. Mar. Drugs 2017, 15, 256. [Google Scholar] [CrossRef] [PubMed]
- Bahrami, Y.; Zhang, W.; Chataway, T.; Franco, C. Structural elucidation of novel saponins in the sea cucumber Holothuria lessoni. Mar. Drugs 2014, 12, 4439–4473. [Google Scholar] [CrossRef] [PubMed]
- Kalinin, V.I.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Stonik, V.A. Triterpene glycosides from sea cucucmbers (Holothuroidea, Echinodermata). Biological activities and functions. Stud. Nat. Prod. Chem. 2008, 35, 135–196. [Google Scholar]
- Yun, S.-H.; Sim, E.-H.; Han, S.-H.; Han, J.-Y.; Kim, S.-H.; Silchenko, A.S.; Stonik, V.A.; Park, J.-I. Holotoxin A1 induces apoptosis by activating acid sphingomyelinase and neutral sphingomyelinase in K562 and human primary leukemia cells. Mar. Drugs 2018, 16, 123. [Google Scholar] [CrossRef] [PubMed]
- Aminin, D.L.; Menchinskaya, E.S.; Pisliagin, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I. Anticancer activity of sea cucumber triterpene glycosides. Mar. Drugs 2015, 13, 1202–1223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-J.; Zhu, K.-Q. A novel antitumor compound nobiliside D isolated from sea cucumber (Holothuria nobilis Selenka). Exp. Ther. Med. 2017, 14, 1653–1658. [Google Scholar] [CrossRef] [PubMed]
- Mitu, S.A.; Bose, U.; Suwansa-ard, S.; Turner, L.H.; Zhao, M.; Elizur, A.; Ogbourne, S.M.; Shaw, P.N.; Cummins, S.F. Evidence for a saponin biosynthesis pathway in the body wall of the commercially significant sea cucumber Holothuria scabra. Mar. Drugs 2017, 15, 349. [Google Scholar] [CrossRef] [PubMed]
- Decroo, C.; Colson, E.; Lemaur, V.; Caulier, G.; De Winter, J.; Cabrera-Barjas, G.; Cornil, J.; Flammang, P.; Gerbaux, P. Ion mobility mass spectrometry of saponin ions. Rapid Commun. Mass Spectrom. 2018. [Google Scholar] [CrossRef] [PubMed]
- Decroo, C.; Colson, E.; Demeyer, M.; Lemaur, V.; Caulier, G.; Eeckhaut, I.; Cornil, J.; Flammang, P.; Gerbaux, P. Tackling saponin diversity in marine animals by mass spectrometry: Data acquisition and integration. Anal. Bioanal. Chem. 2017, 409, 3115–3126. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Stonik, V.A.; Avilov, S.A.; Kalinin, V.I.; Kalinovsky, A.I.; Zaharenko, A.M.; Smirnov, A.V.; Mollo, E.; Cimino, G. Holothurins B2, B3, and B4, new triterpene glycosides from mediterranean sea cucumbers of the genus holothuria. J. Nat. Prod. 2005, 68, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Hori, M.; Kan, K.; Yasuzawa, T.; Matsui, M.; Suzuki, S.; Kitagawa, I. Marine natural products. XXVII: Distribution of lanostane-type triterpene oligoglycosides in ten kinds of Okinawan Sea cucumbers. Chem. Pharm. Bull. 1991, 39, 2282–2287. [Google Scholar] [CrossRef]
- Kitagawa, I.; Nishino, T.; Matsuno, T.; Akutsu, H.; Kyogoku, Y. Structure of holothurin B a pharmacologically active triterpene-oligoglycoside from the sea cucumber Holothuria leucospilota Brandt. Tetrahedron Lett. 1978, 19, 985–988. [Google Scholar] [CrossRef]
- Wu, J.; Yi, Y.H.; Tang, H.F.; Wu, H.M.; Zou, Z.R.; Lin, H.W. Nobilisides A–C, three new triterpene glycosides from the sea cucumber Holothuria nobilis. Planta Med. 2006, 72, 932–935. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Zhang, W.; Yi, Y.H.; Liu, B.S.; Pan, M.X.; Wang, X.H. A novel sulfated holostane glycoside from sea cucumber Holothuria leucospilota. Chem. Biodivers. 2010, 7, 1764–1769. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yi, Y.H.; Li, L.; Wang, X.H.; Liu, B.S.; Sun, P.; Pan, M.X. A new triterpene glycoside from sea cucumber Holothuria leucospilota. Chin. Chem. Lett. 2007, 18, 161–164. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M.; Son, B.W.; Suzuki, S.; Kyogoku, Y. Marine natural products. XIX: Pervicosides A, B, and C, lanostane-type triterpene-oligoglycoside sulfates from the sea cucumber Holothuria pervicax. Chem. Pharm. Bull. 1989, 37, 1230–1234. [Google Scholar] [CrossRef]
- Caulier, G.; Flammang, P.; Gerbaux, P.; Eeckhaut, I. When a repellent becomes an attractant: Harmful saponins are kairomones attracting the symbiotic Harlequin crab. Sci. Rep. 2013, 3, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, I.; Kobayashi, M.; Hori, M.; Kyogoku, Y. Marine natural products. XVIII. Four lanostane-type triterpene oligoglycosides, bivittosides A, B, C and D, from the Okinawan sea cucumber Bohadschia bivittata mitsukuri. Chem. Pharm. Bull. 1989, 37, 61–67. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M.; Kyogoku, Y. Marine natural products. IX. Structural elucidation of triterpenoidal oligoglycosides from the Bahamean sea cucumber Actinopyga agassizi Selenka. Chem. Pharm. Bull. 1982, 30, 2045–2050. [Google Scholar] [CrossRef]
- Rodriguez, J.; Castro, R.; Riguera, R. Holothurinosides: New antitumour non sulphated triterpenoid glycosides from the sea cucumber Holothuria forskalii. Tetrahedron 1991, 47, 4753–4762. [Google Scholar] [CrossRef]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Elucidation of molecular diversity and body distribution of saponins in the sea cucumber Holothuria forskali (Echinodermata) by mass spectrometry. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2009, 152, 124–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.S.; Yi, Y.H.; Li, L.; Sun, P.; Han, H.; Sun, G.Q.; Wang, X.H.; Wang, Z.L. Argusides D and E, two new cytotoxic triterpene glycosides from the sea cucumber Bohadschia argus Jaeger. Chem. Biodivers. 2008, 5, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Elbandy, M.; Rho, J.; Afifi, R. Analysis of saponins as bioactive zoochemicals from the marine functional food sea cucumber Bohadschia cousteaui. Eur. Food Res. Technol. 2014, 238, 937–955. [Google Scholar] [CrossRef]
- Wu, J.; Yi, Y.; Zou, Z. Two new triterpene glycosides from sea cucumber Holothuria nobilis. Chin. Tradit. Herb. Drugs 2006, 37, 497. [Google Scholar]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Qualitative and quantitative saponin contents in five sea cucumbers from the Indian ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.S.; Yi, Y.H.; Li, L.; Zhang, S.L.; Han, H.; Weng, Y.Y.; Pan, M.X. Arguside A: A new cytotoxic triterpene glycoside from the sea cucumber Bohadschia argus Jaeger. Chem. Biodivers. 2007, 4, 2845–2851. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Yi, Y.; Wu, H.; Wu, J.; Liaw, C.-C.; Lee, K.-H. Intercedensides A−C, three new cytotoxic triterpene glycosides from the sea cucumber Mensamaria intercedens Lampert. J. Nat. Prod. 2003, 66, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-Y.; Yi, Y.-H.; Tang, H.-F. Bioactive triterpene glycosides from the sea cucumber Holothuria fuscocinerea. J. Nat. Prod. 2006, 69, 1492–1495. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Yi, Y.; Xu, Q.; La, M.; Zhang, H. Two new cytotoxic triterpene glycosides from the sea cucumber Holothuria scabra. Planta Med. 2009, 75, 1608–1612. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, I.; Nishino, T.; Kyogoku, Y. Structure of holothurin A a biologically active triterpene-oligoglycoside from the sea cucumber Holothuria leucospilota Brandt. Tetrahedron Lett. 1979, 20, 1419–1422. [Google Scholar] [CrossRef]
- Han, H.; Yi, Y.H.; Li, L.; Liu, B.S.; La, M.P.; Zhang, H.W. Antifungal active triterpene glycosides from sea cucumber Holothuria scabra. Acta Pharm. Sin. 2009, 44, 620–624. [Google Scholar]
- Han, H.; Li, L.; Yi, Y.; Wang, X.; Pan, M. Triterpene glycosides from sea cucumber Holothuria scabra with cytotoxic activity. Chin. Herb. Med. 2012, 4, 183–188. [Google Scholar]
- Caulier, G.; Van Dyck, S.; Gerbaux, P.; Eeckhaut, I.; Flammang, P. Review of saponin diversity in sea cucumbers belonging to the family Holothuriidae. SPC Beche-de-Mer. Inf. Bull. 2011, 31, 48–54. [Google Scholar]
- Kalinin, V.I.; Stonik, V.A. Glycosides of marine invertebrates. Structure of Holothurin A2 from the holothurian Holothuria edulis. Chem. Nat. Compd. 1982, 18, 196–200. [Google Scholar] [CrossRef]
- Dong, P.; Xue, C.; Du, Q. Separation of two main triterpene glycosides from sea cucumber Pearsonothuria graeffei by high-speed countercurrent chromatography. Acta Chromatogr. 2008, 20, 269–276. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M.; Inamoto, T.; Fuchida, M.; Kyogoku, Y. Marine natural products. XIV. Structures of echinosides A and B, antifungal lanostane-oligosides from the sea cucumber Actinopyga echinites (Jaeger). Chem. Pharm. Bull. 1985, 33, 5214–5224. [Google Scholar] [CrossRef] [PubMed]
- Thanh, N.V.; Dang, N.H.; Kiem, P.V.; Cuong, N.X.; Huong, H.T.; Minh, C.V. A new triterpene glycoside from the sea cucumber Holothuria scabra collected in Vietnam. Asean J. Sci. Technol. Dev. 2006, 23, 253–259. [Google Scholar] [CrossRef]
- Yuan, W.H.; Yi, Y.H.; Tang, H.F.; Xue, M.; Wang, Z.L.; Sun, G.Q.; Zhang, W.; Liu, B.S.; Li, L.; Sun, P. Two new holostan-type triterpene glycosides from the sea cucumber Bohadschia marmorata Jaeger. Chem. Pharm. Bull. 2008, 56, 1207–1211. [Google Scholar] [CrossRef] [PubMed]
- Chanley, J.D.; Ledeen, R.; Wax, J.; Nigrelli, R.F.; Sobotka, H.; Holothurin, I. The isolation, properties and sugar components of holothurin A1. J. Am. Chem. Soc. 1959, 81, 5180–5183. [Google Scholar] [CrossRef]
- Elyakov, G.B.; Stonik, V.A.; Levina, E.V.; Slanke, V.P.; Kuznetsova, T.A.; Levin, V.S. Glycosides of marine invertebrates—I. A comparative study of the glycoside fractions of pacific sea cucumbers. Comp. Biochem. Physiol. B 1973, 44, 325–336. [Google Scholar] [CrossRef]
- Elyakov, G.B.; Kuznetsova, T.A.; Stonik, V.A.; Levin, V.S.; Albores, R. Glycosides of marine invertebrates. IV. A comparative study of the glycosides from Cuban sublittoral holothurians. Comp. Biochem. Physiol. B 1975, 52, 413–417. [Google Scholar] [CrossRef]
- Yasumoto, T.; Nakamura, K.; Hashimoto, Y. A new saponin, holothurin B, isolated from sea-cucumber, Holothuria vagabunda and Holothuria lubrica. Agric. Biol. Chem. 1967, 31, 7–10. [Google Scholar] [CrossRef]
- Matsuno, T.; Iba, J. Studies on the saponins of the sea cucumber. Yakugaku Zasshi 1966, 86, 637–638. [Google Scholar] [PubMed]
- Dang, N.H.; Thanh, N.V.; Kiem, P.V.; Huong le, M.; Minh, C.V.; Kim, Y.H. Two new triterpene glycosides from the Vietnamese sea cucumber Holothuria scabra. Arch. Pharm. Res. 2007, 30, 1387–1391. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.H.; Yi, Y.H.; Tan, R.X.; Wang, Z.L.; Sun, G.Q.; Xue, M.; Zhang, H.W.; Tang, H.F. Antifungal triterpene glycosides from the sea cucumber Holothuria (Microthele) axiloga. Planta Med. 2009, 75, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Van Dyck, S.; Flammang, P.; Meriaux, C.; Bonnel, D.; Salzet, M.; Fournier, I.; Wisztorski, M. Localization of secondary metabolites in marine invertebrates: Contribution of MALDI MSI for the study of saponins in Cuvierian tubules of H. forskali. PLoS ONE 2010, 5, e13923. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.Q.; Li, L.; Yi, Y.H.; Yuan, W.H.; Liu, B.S.; Weng, Y.Y.; Zhang, S.L.; Sun, P.; Wang, Z.L. Two new cytotoxic nonsulfated pentasaccharide holostane (=20-hydroxylanostan-18-oic acid γ-lactone) glycosides from the sea cucumber Holothuria grisea. Helv. Chim. Acta 2008, 91, 1453–1460. [Google Scholar] [CrossRef]
- Bhatnagar, S.; Dudouet, B.; Ahond, A.; Poupat, C.; Thoison, O.; Clastres, A.; Laurent, D.; Potier, P. Marine-invertebrates. 4. Saponins and sapogenins from a seacucumber, Actinopyga flammea. Bull. Soc. Chim. Fr. 1985, 1, 124–129. [Google Scholar]
- Van Dyck, S.; Caulier, G.; Todesco, M.; Gerbaux, P.; Fournier, I.; Wisztorski, M.; Flammang, P. The triterpene glycosides of Holothuria forskali: Usefulness and efficiency as a chemical defense mechanism against predatory fish. J. Exp. Biol. 2011, 214 Pt 8, 1347–1356. [Google Scholar] [CrossRef]
- Yuan, W.H.; Yi, Y.H.; Tang, H.F.; Liu, B.S.; Wang, Z.L.; Sun, G.Q.; Zhang, W.; Li, L.; Sun, P. Antifungal triterpene glycosides from the sea cucumber Bohadschia marmorata. Planta Med. 2009, 75, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.S.; Yi, Y.H.; Li, L.; Sun, P.; Yuan, W.H.; Sun, G.Q.; Han, H.; Xue, M. Argusides B and C, two new cytotoxic triterpene glycosides from the sea cucumber Bohadschia argus Jaeger. Chem. Biodivers. 2008, 5, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Stonik, V.A.; Chumak, A.D.; Isakov, V.V.; Belogortseva, N.I.; Chirva, V.Y.; Elyakov, G.B. Glycosides of marine invertebrates. VII. Structure of holothurin B from Holothuria atra. Chem. Nat. Compd. 1979, 15, 453–457. [Google Scholar] [CrossRef]
- Liu, J.; Yang, X.; He, J.; Xia, M.; Xu, L.; Yang, S. Structure analysis of triterpene saponins in Polygala tenuifolia by electrospray ionization ion trap multiple-stage mass spectrometry. J. Mass Spectrom. 2007, 42, 861–873. [Google Scholar] [CrossRef] [PubMed]
- Demeyer, M.; De Winter, J.; Caulier, G.; Eeckhaut, I.; Flammang, P.; Gerbaux, P. Molecular diversity and body distribution of saponins in the sea star Asterias rubens by mass spectrometry. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 168, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Liu, B.S.; Yi, Y.H.; Li, L.; Gui, M.; Tang, H.F.; Zhang, D.Z.; Zhang, S.L. A new cytotoxic lanostane-type triterpene glycoside from the sea cucumber Holothuria impatiens. Chem. Biodivers. 2007, 4, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-H.; Zou, Z.-R.; Yi, Y.-H.; Han, H.; Li, L.; Pan, M.-X. Variegatusides: New non-sulphated triterpene glycosides from the sea cucumber Stichopus variegates Semper. Mar. Drugs 2014, 12, 2004–2018. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Kalinovsky, A.I.; Avilov, S.A.; Andryjaschenko, P.V.; Dmitrenok, P.S.; Martyyas, E.A.; Kalinin, V.I.; Jayasandhya, P.; Rajan, G.C.; Padmakumar, K.P. Structures and biological activities of Typicosides A1, A2, B1, C1 and C2, triterpene glycosides from the sea cucumber Actinocucumis typica. Nat. Prod. Commun. 2013, 8, 301–310. [Google Scholar] [PubMed]
- Bondoc, K.G.V.; Lee, H.; Cruz, L.J.; Lebrilla, C.B.; Juinio-Meñez, M.A. Chemical fingerprinting and phylogenetic mapping of saponin congeners from three tropical holothurian sea cucumbers. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2013, 166, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Bakus, G.J. Defensive mechanisms and ecology of some tropical holothurians. Mar. Biol. 1968, 2, 23–32. [Google Scholar] [CrossRef]
- Mercier, A.; Sims, D.W.; Hamel, J.F. Advances in Marine Biology: Endogenous and Exogenous Control of Gametogenesis and Spawning in Echinoderms; Academic Press: New York, NY, USA, 2009; Volume 55. [Google Scholar]
- Kalinin, V.I. System-theoretical (Holistic) approach to the modelling of structural-functional relationships of biomolecules and their evolution: An example of triterpene glycosides from sea cucumbers (Echinodermata, Holothurioidea). J. Theor. Biol. 2000, 206, 151–168. [Google Scholar] [CrossRef] [PubMed]
- Popov, R.S.; Ivanchina, N.V.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Dolmatov, I.Y.; Stonik, V.A.; Dmitrenok, P.S. Metabolite profiling of triterpene glycosides of the far eastern sea cucumber eupentacta fraudatrix and their distribution in various body components using LC-ESI QTOF-MS. Mar. Drugs 2017, 15, 302. [Google Scholar] [CrossRef] [PubMed]
- Kitagawa, I.; Kobayashi, M.; Imamoto, T.; Yasuzawa, T.; Kyogoku, Y. The structures of six antifungal oligoglycosides, stichlorosides A1, A2, B1, B2, C1 and C2, from the sea cucumber Stichopus chloronotus Brandt. Chem. Pharm. Bull. 1981, 29, 2387–2391. [Google Scholar] [CrossRef]
- Mokhlesi, A.; Saeidnia, S.; Gohari, A.R.; Shahverdi, A.R.; Nasrolahi, A.; Farahani, F.; Khoshnood, R.; Es’ haghi, N. Biological activities of the sea cucumber Holothuria leucospilota. Asian J. Anim. Vet. Adv. 2012, 7, 243–249. [Google Scholar]
- Kuznetsova, T.A.; Anisimov, M.M.; Popov, A.M.; Baranova, S.I.; Afiyatullov, S.; Kapustina, I.I.; Antonov, A.S.; Elyakov, G.B. A comparative study in vitro of physiological activity of triterpene glycosides of marine invertebrates of echinoderm type. Comp. Biochem. Physiol. C 1982, 73, 41–43. [Google Scholar] [CrossRef]
- Abraham, T.J.; Nagarajan, J.; Shanmugam, S.A. Antimicrobial substances of potential biomedical importance from holothurian species. Indian J. Mar. Sci. 2002, 31, 161–164. [Google Scholar]
- Park, S.Y.; Lim, H.K.; Lee, S.; Cho, S.K.; Park, S.; Cho, M. Biological effects of various solvent fractions derived from Jeju Island red sea cucumber (Stichopus japonicus). J. Korean Soc. Appl. Biol. Chem. 2011, 54, 718–724. [Google Scholar] [CrossRef]
- Husni, A.; Shin, I.-S.; You, S.; Chung, D. Antioxidant properties of water and aqueous ethanol extracts and their crude saponin fractions from a far-eastern sea cucumber, Stichopus japonicus. Food Sci. Biotechnol. 2009, 18, 419–424. [Google Scholar]
- Raza, W.; Yang, X.; Wu, H.; Wang, Y.; Xu, Y.; Shen, Q. Isolation and characterisation of fusaricidin-type compound-producing strain of Paenibacillus polymyxa SQR-21 active against Fusarium oxysporum f.sp. nevium. Eur. J. Plant Pathol. 2009, 125, 471–483. [Google Scholar] [CrossRef]
- Almuzara, M.; Limansky, A.; Ballerini, V.; Galanternik, L.; Famiglietti, A.; Vay, C. In vitro susceptibility of Achromobacter spp. isolates: Comparison of disk diffusion, Etest and agar dilution methods. Int. J. Antimicrob. Agents 2010, 35, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Wikler, M.A. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically, 9th ed.; CLSI Document M07-A9; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 32. [Google Scholar]


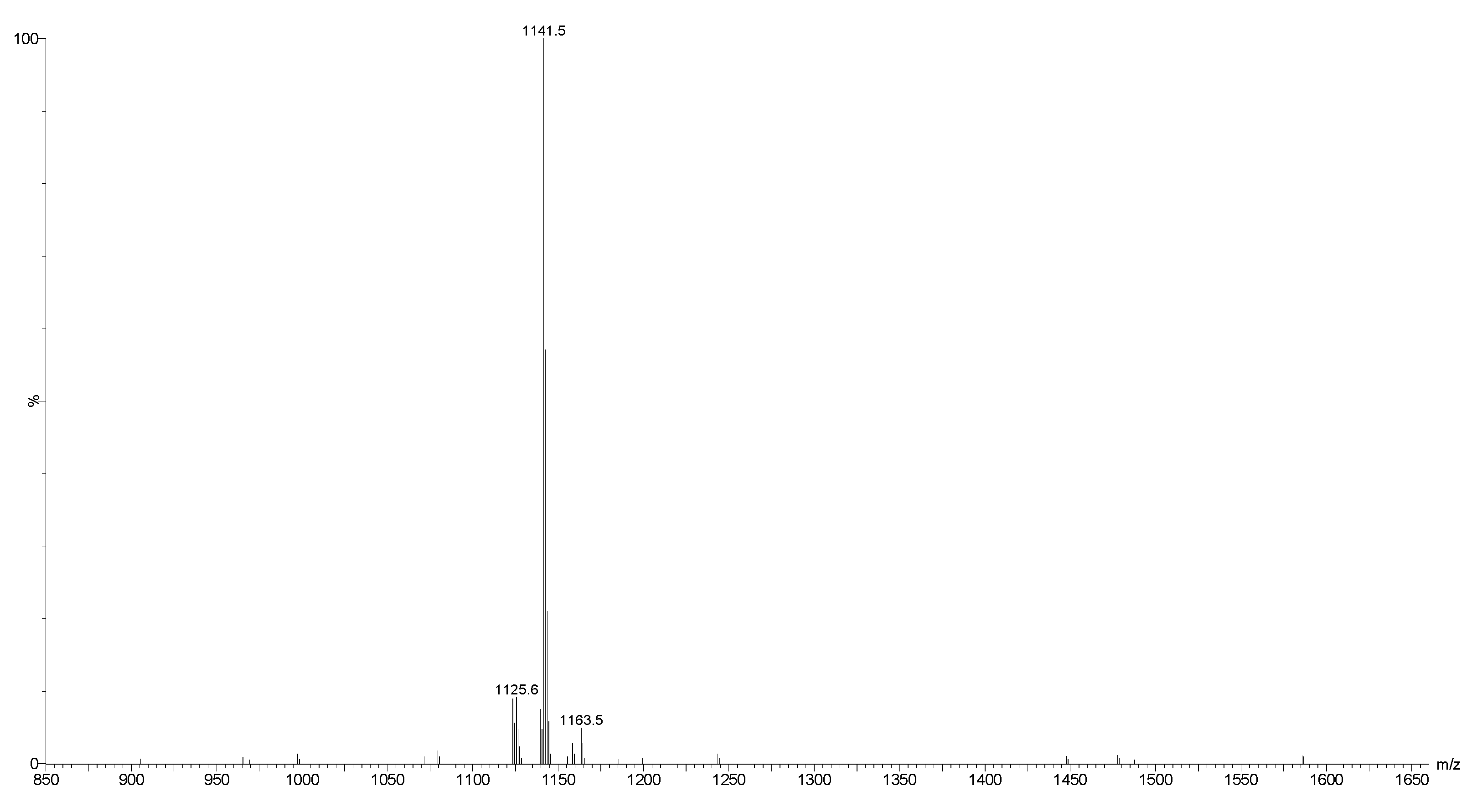


{kind=link}
{kind=link}
{kind=link}
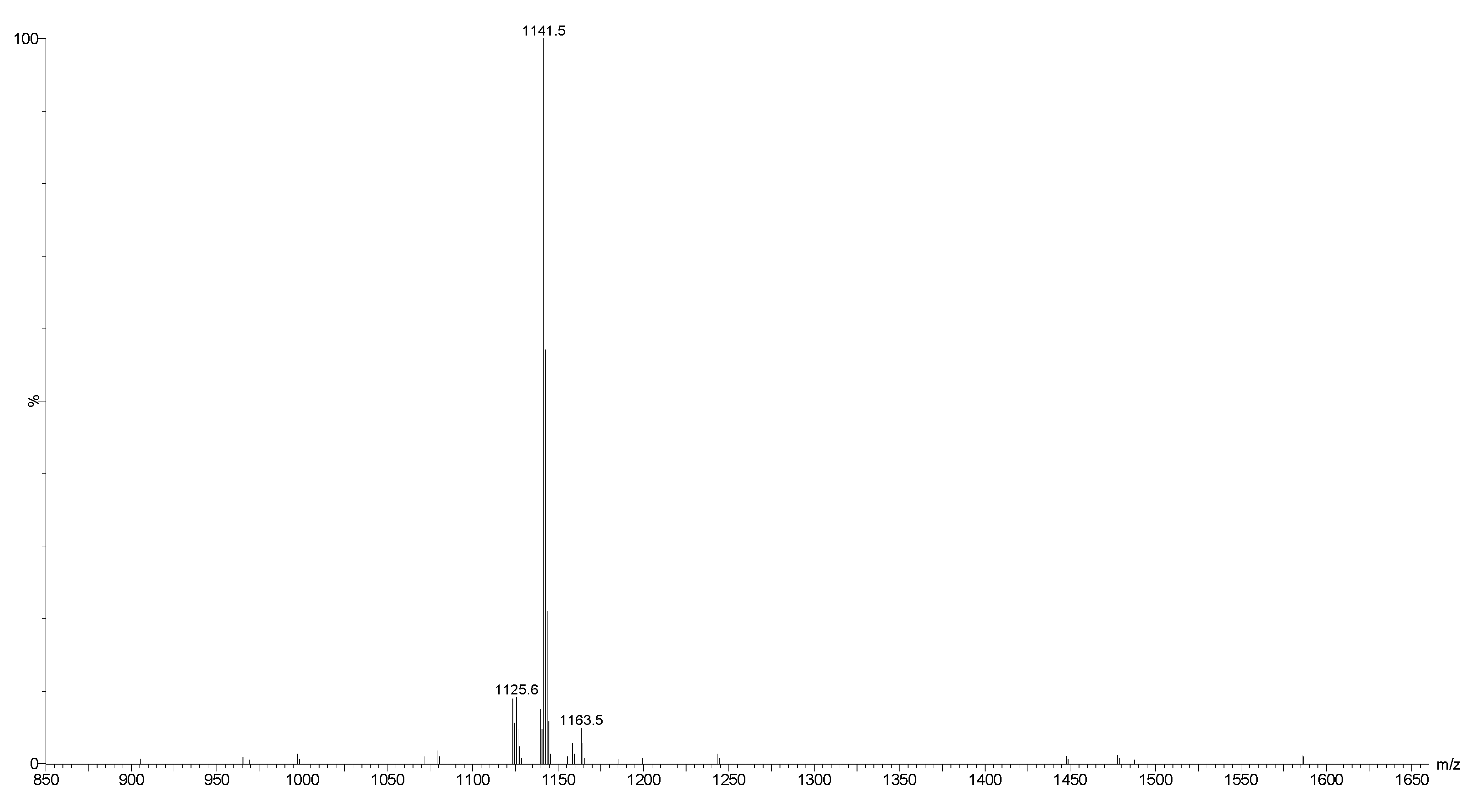{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| [M + Na]+ m/z | MW | Formula | Compound Name | Body Wall | Viscera | Novel (N)/Published (P) | References |
|---|---|---|---|---|---|---|---|
| 889.4 | 866 | C41H63NaO16S | Holothurin B3 | Yes | Yes | P | [19] |
| C42H67NaO15S | Unidentified | Yes | Yes | N | − | ||
| 905.4 | 882 | C41H63NaO17S | Holothurin B4 | Yes | Yes | P | [2,19] |
| Holothurin B | Yes | Yes | P | [20,21] | |||
| Nobiliside B | Yes | Yes | P | [22] | |||
| 907.4 | 884 | C41H65NaO17S | Holothurin B2 | No | Yes | P | [19] |
| Leucospilotaside B | No | Yes | P | [23] | |||
| 911.6 | 888 | C45H92O16 | Unidentified | Yes | Yes | N | − |
| 917.4 | 994 | C44H71NaO15S | Unidentified | No | Yes | N | − |
| 921.4 | 898 | C41H63NaO18S | Leucospilotaside A | No | Yes | P | [24] |
| 1034.1 | 1011 | a * | Unidentified | Yes | Yes | N | − |
| 1065.5 | 1042 | C48H82O24 | Unidentified | No | Yes | N | − |
| 1069.5 | 1046 | C52H86O21 | Unidentified | Yes | No | N | - |
| 1071.5 | 1048 | C47H93NaO21S | Unidentified | Yes | Yes | N | [2,11] |
| 1079.5 | 1056 | C53H84O21 | Unidentified | Yes | Yes | N | - |
| 1083.3 | 1060 | C58H64O25 | Unidentified | No | Yes | N | [2,11] |
| 1085.5 | 1062 | C53H90O21 | Unidentified | No | Yes | N | - |
| 1087.5 | 1064 | C52H88O22 C47H93NaO22S | Unidentified | Yes | Yes | N | [2,11] |
| 1101.6 | 1078 | C52H86O23 | Unidentified | Yes | Yes | N | - |
| 1103.5 | 1080 | C52H88O23 | Unidentified | Yes | No | N | - |
| 1107.7 | 1084 | C54H84O22 | Unidentified | Yes | Yes | N | - |
| 1109.5 | 1086 | C54H86O22 | DS-pervicoside B | Yes | Yes | P | [25] |
| 1111.5 | 1088 | C54H88O22 | Bivitoside B | Yes | Yes | P | [26,27] |
| 1121.5 | 1098 | C54H82O23 | Unidentified | No | Yes | N | - |
| 1123.5 | 1100 | C54H84O23 | Unidentified | Yes | Yes | N | [2,11] |
| 1125.5 | 1102 | C54H86O23 | Holothurinosides C/C1 | Yes | Yes | P | [28,29] |
| 1127.6 | 1104 | C53H84O24C54H88O23 | Holothurinosides X/Y/Z | Yes | Yes | P | [2,11] |
| 1139.5 | 1116 | C54H84O24 | Unidentified | No | Yes | N | - |
| 1141.5 | 1118 | C54H86O24 | Desholothurin A (Nobiliside 2a), Desholothurin A1 (Arguside E) | Yes | Yes | P | [2,28,29,30,31,32,33] |
| 1149.2 | 1126 | a * | Holothurinoside T | No | Yes | P | − |
| 1157.5 | 1134 | C54H86O25 | Holothurinoside J1 Unidentified | Yes | Yes | P N | [2,11,34] |
| 1163.5 | 1140 | C54H92O25 | Unidentified | Yes | Yes | N | - |
| 1167.8 | 1144 | C56H88O24 | Arguside A | No | Yes | P | [35] |
| 1173.5 | 1150 | C57 H82O24 | Unidentified | Yes | Yes | N | - |
| 1179.5 | 1156 | C57H88O24C54H85NaO23S | Unidentified | Yes | Yes | N | - |
| 1181.4 | 1158 | C57H90O24 | Unidentified | No | Yes | N | - |
| 1189.5 | 1166 | C59H97O24 | Unidentified | Yes | No | N | - |
| 1193.5 | 1170 | C54H83NaO24S | Unidentified | Yes | Yes | N | [2,11] |
| 1197.5 | 1174 | C54H87NaO24S | Unidentified | Yes | Yes | N | - |
| 1199.5 | 1176 | C54H64O29C56H88O26 | Unidentified Arguside D | Yes | Yes | N P | [2,31] |
| 1205.5 | 1182 | C57H82O26 C55H83NaO24S | Unidentified | Yes | Yes | N | - |
| 1207.5 | 1184 | C55H83NaO24S | Unidentified | Yes | Yes | N | - |
| 1211.5 | 1188 | C54H85NaO25S | Unidentified | Yes | Yes | N | - |
| 1221.5 | 1198 | C56H78O28C55H83NaO25S | Unidentified Intercedenside A | Yes | Yes | N P | [2,36] |
| 1223.5 | 1200 | C55H85NaO25S | Unidentified | No | Yes | N | - |
| 1225.5 | 1202 | C54H83NaO26S | Unidentified | No | Yes | N | − |
| 1227.5 | 1204 | C54H85NaO26S | Fuscocinerosides B/C, Scabraside A or 24–dehydroechinoside A, Unidentified | Yes | Yes | P | [11,28,37,38,39,40,41,42] |
| 1229.5 | 1206 | C54H87NaO26S | Holothurin A2, Echinoside A Pervicoside B | Yes | Yes | P | [20,26,40,43,44,45,46] |
| 1237.5 | 1214 | C56H78O29C55H83NaO26S | Unidentified | Yes | Yes | N | - |
| 1243.5 | 1220 | C54H85NaO27S | Holothurin A Scabraside B 17-Hydroxy fuscocineroside B, 25-Hydroxy fuscocinerosiden B | Yes | Yes | P | [19,20,33,38,39,46,47,48,49,50,51,52] |
| 1245.5 | 1222 | C54H87NaO27S | Holothurin A1 Holothurin A4 Scabraside D | No | Yes | P | [40,41,53] |
| 1259.5 | 1236 | C54H85NaO28S | Holothurin A3 Holothurin D | Yes | Yes | P P | [2,11,53] |
| 1261.5 | 1238 | C54H87NaO28S | Unidentified | No | Yes | N | − |
| 1265.5 | 1242 | C56H83NaO27S | Unidentified | Yes | Yes | N | [2] |
| 1269.5 | 1246 | C60H94O27 | Cousteside G | No | Yes | P | [32] |
| 1271.6 | 1248 | C60H96O27 | Impatienside B Cousteside H | Yes | Yes | P | [32,54] |
| 1273.6 | 1250 | C60H98O27 | Cousteside J | Yes | Yes | P | [2,32] |
| 1281.4 | 1258 | C54H87NaO29S | Unidentified | No | Yes | N | - |
| 1283.4 | 1260 | C54H89NaO29S C61H96O27 | Unidentified | No | Yes | N | - |
| 1285.6 | 1262 | C56H87NaO28S | Fuscocineroside A | Yes | Yes | P | [37] |
| 1287.6 | 1264 | C60H96O28 | Holothurinoside E, | Yes | Yes | P | [30,55] |
| Holothurinoside E1 | Yes | Yes | P | [30,55] | |||
| Holothurinoside O | Yes | Yes | P | [2,11] | |||
| Holothurinoside P | Yes | Yes | P | [2,11] | |||
| 17-dehydroxy holothurinoside A | Yes | Yes | P | [32,56] | |||
| Cousteside E | Yes | Yes | P | [32] | |||
| Cousteside F | Yes | Yes | P | [32] | |||
| C56H89NaO28S | 22-acetoxy-echinoside A | Yes | Yes | P | [57] | ||
| 1289.6 | 1266 | C60H98O28 | Griseaside A Cousteside I | Yes Yes | Yes Yes | P P | [56] [32] |
| 1301.6 | 1278 | C61H98O28C60H94O29 | Holothurinoside M Unidentified | Yes | Yes | P N | [11,58] |
| 1303.6 | 1280 | C60H96O29 | Holothurinoside A | Yes | Yes | P | [29,30] |
| Holothurinoside A1 | Yes | Yes | P | [29,30] | |||
| Holothurinoside Q | Yes | Yes | P | [2,11] | |||
| Holothurinoside S | Yes | Yes | P | [2,11] | |||
| Holothurinoside R | Yes | Yes | P | [2,11] | |||
| Holothurinoside R1 | Yes | Yes | P | [2,11] | |||
| Cousteside C | Yes | Yes | P | [32] | |||
| 1305.6 | 1282 | C60H98O29 | Unidentified | Yes | Yes | N | [2] |
| 1307.6 | 1284 | C60H100O29 | Unidentified | Yes | Yes | N | [2] |
| 1317.6 | 1294 | C61H98O29 | Unidentified Holothurinoside L | Yes | Yes | N P | [2,11,26] |
| 1319.5 | 1296 | C60H96O30 | Unidentified | Yes | Yes | N | - |
| 1329.7 | 1306 | C62H98O29 | Arguside F | No | Yes | P | [54] |
| 1335.3 | 1312 | C60H96O31 | Unidentified | Yes | Yes | N | [2] |
| 1349.8 | 1326 | C61H98O31 | Unidentified | No | Yes | N | - |
| 1356.4 | 1333 | a * | Unidentified | No | Yes | N | − |
| 1361.7 | 1338 | C63H102O30 | Unidentified | Yes | Yes | N | - |
| 1377.3 | 1354 | C63H102O31 | Unidentified | No | Yes | N | - |
| 1409.4 | 1386 | C61H78O36 | Unidentified | Yes | Yes | N | [2] |
| 1411.7 | 1388 | C62H116O33 | Unidentified | No | Yes | N | − |
| 1415.7 | 1392 | C66H104O31 | Unidentified | No | Yes | N | - |
| 1417.7 | 1394 | C66H106O31 | Unidentified | Yes | Yes | N | - |
| 1419.7 | 1396 | C66H108O31 | Unidentified | Yes | Yes | N | [2] |
| 1431.4 | 1408 | C66H104O32 | Unidentified | No | Yes | N | - |
| 1435.7 | 1412 | C66H108O32 | Unidentified | Yes | Yes | N | [2] |
| 1447.7 | 1424 | C67H108O32 | Unidentified Impatienside A Marmoratoside A | Yes | Yes | N P | - [59] |
| 1449.7 | 1426 | C67H110O32 | Bivittoside D | No | Yes | P | [27] |
| 1453.6 | 1430 | C66H94O34 | Unidentified | Yes | Yes | N | - |
| 1459.7 | 1436 | C68H108O32 | Unidentified | Yes | No | N | - |
| 1461.7 | 1438 | C68H110O32 | Unidentified | Yes | No | N | - |
| 1463.7 | 1440 | C67H108O33 | Holothurinosides H/H1 Holothurin C Cousteside A 17α-hydroxy impatienside A Marmoratoside B | Yes | No | P | [26,55] [32] [59] |
| 1465.7 | 1442 | C67H110O33 | Argusides B/C | No | Yes | P | [60] |
| 1475.7 | 1452 | C68H108O33C65H112O35 | Unidentified | Yes | Yes | N | [11] |
| 1477.7 | 1454 | C68H110O33C65H114O35 | Lessoniosides A/B/C/D/E Unidentified | Yes | Yes | P | [6] |
| 1479.7 | 1456 | C67H108O34 | Holothurinosides I/I1 | No | Yes | P | [55] |
| 1481.7 | 1458 | C66H106O35C67H110O34 | Unidentified | Yes | Yes | N | [2] |
| 1489.7 | 1466 | C68H106O34 | Unidentified | Yes | No | N | - |
| 1491.5 | 1468 | C68H108O34 | Unidentified | No | Yes | N | - |
| 1493.7 | 1470 | C68H110O34C65H114O36 | Unidentified | No | Yes | N | − |
| 1495.7 | 1472 | C67H108O35 | Holothurinoside K1 Unidentified | No | Yes | P N | [34] − |
| 1507.7 | 1484 | C69H112O34 | 25-acetoxy bivittoside D | Yes | Yes | P | [59] |
| 1521.7 | 1498 | C69H110O35 | Unidentified | Yes | Yes | N | - |
| 1535.7 | 1412 | C69H108O36 | Unidentified | Yes | No | N | - |
| 1539.7 | 1416 | C69H112O36 | Unidentified | Yes | No | N | - |
| 1591.7 | 1568 | C66H120O41 | Unidentified | No | Yes | N | − |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bahrami, Y.; Zhang, W.; M. M. Franco, C. Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities. Mar. Drugs 2018, 16, 423. https://doi.org/10.3390/md16110423
Bahrami Y, Zhang W, M. M. Franco C. Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities. Marine Drugs. 2018; 16(11):423. https://doi.org/10.3390/md16110423
Chicago/Turabian StyleBahrami, Yadollah, Wei Zhang, and Christopher M. M. Franco. 2018. "Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities" Marine Drugs 16, no. 11: 423. https://doi.org/10.3390/md16110423
APA StyleBahrami, Y., Zhang, W., & M. M. Franco, C. (2018). Distribution of Saponins in the Sea Cucumber Holothuria lessoni; the Body Wall Versus the Viscera, and Their Biological Activities. Marine Drugs, 16(11), 423. https://doi.org/10.3390/md16110423