
Molecular and Epigenetic Control of Aldosterone Synthase, CYP11B2 and 11-Hydroxylase, CYP11B1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Epigenetic Control of Gene Expression
3. DNA Methylation
4. Histone Modifications
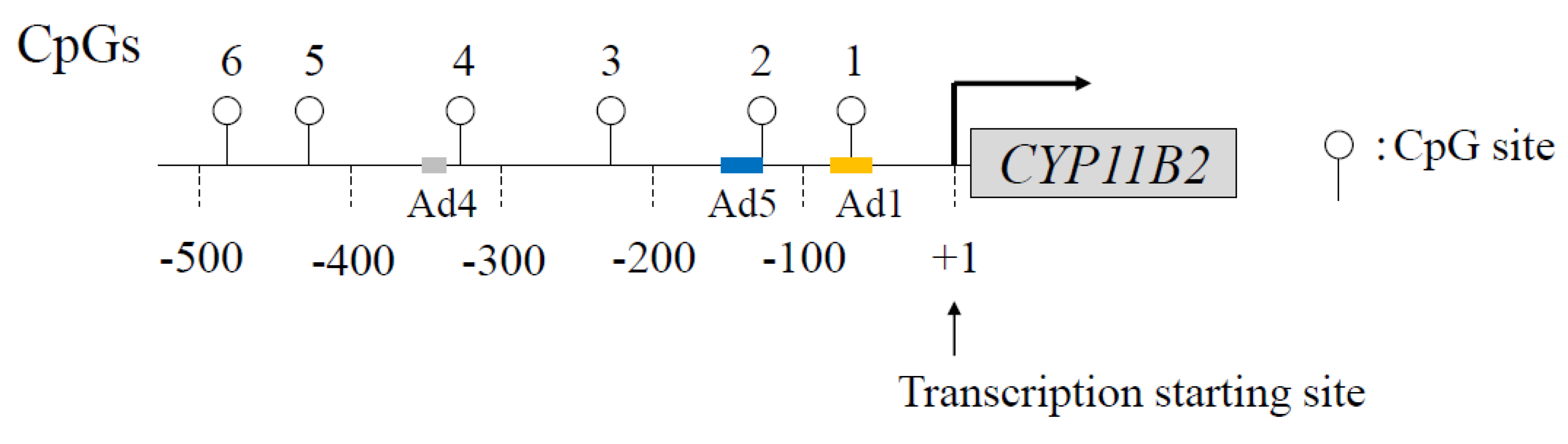
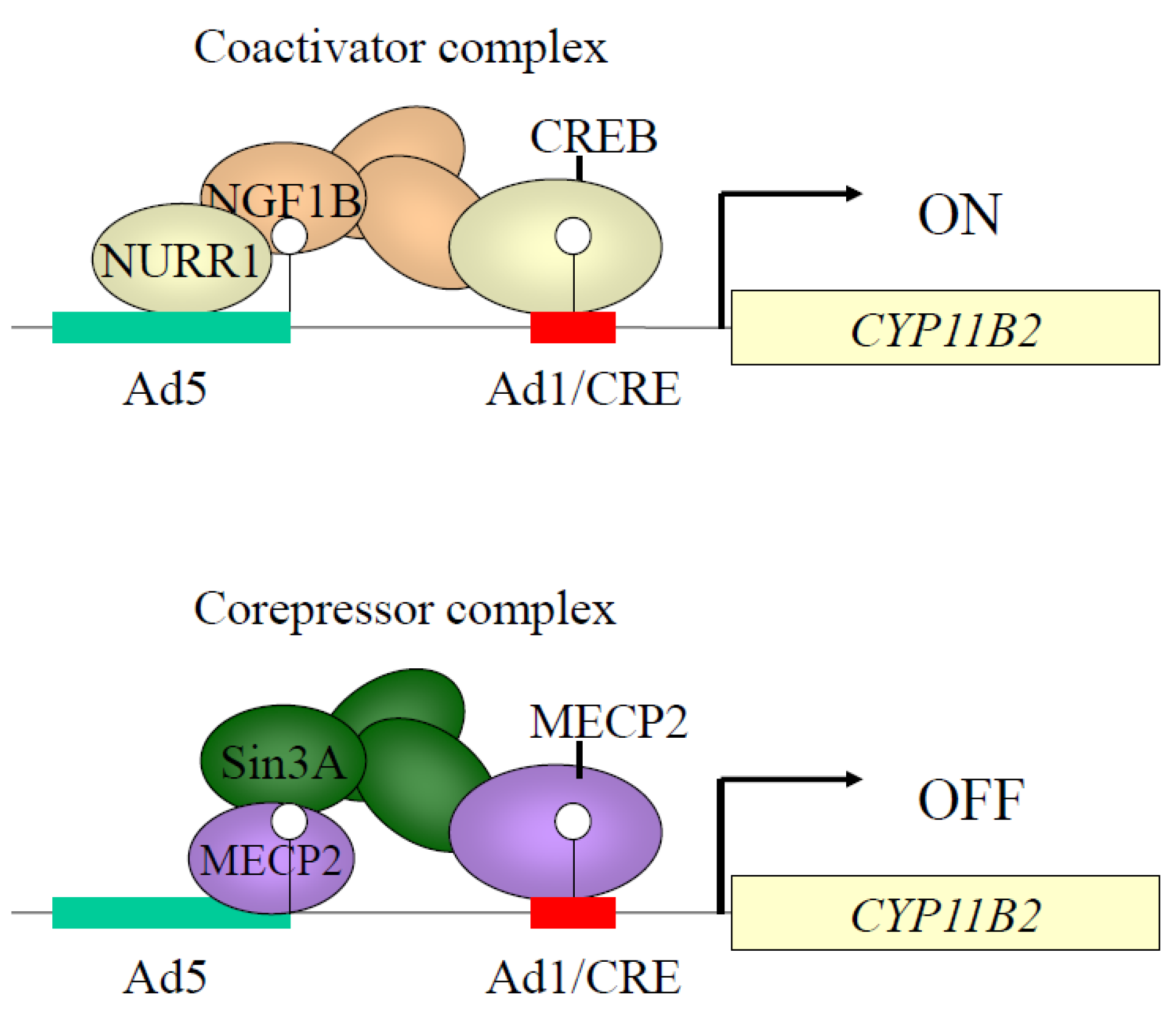
5. Epigenetic Control of CYP11B2
6. Epigenetic Modification of CYP11B2 in Aldosterone-Producing Adenoma
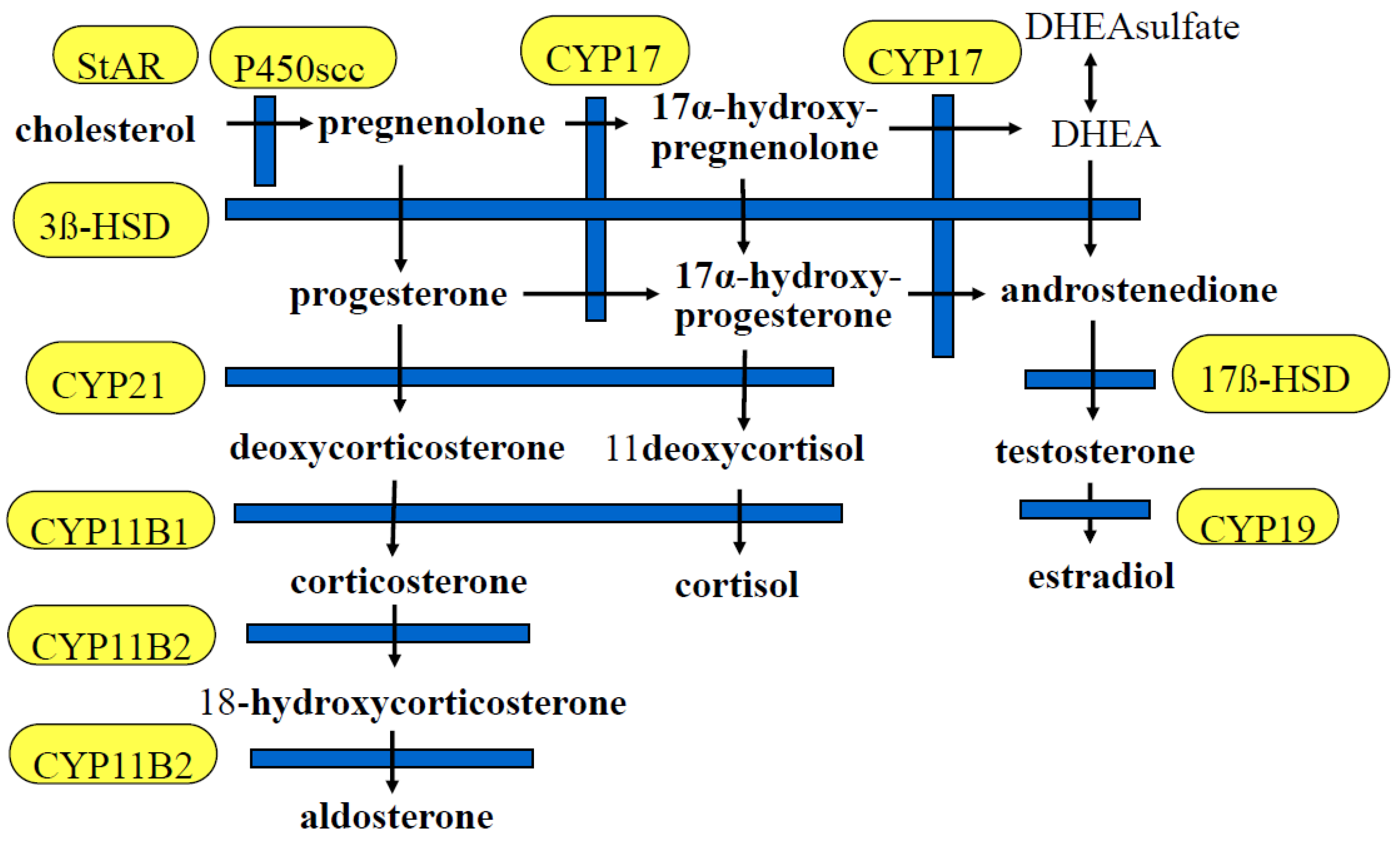
7. Extra-Adrenal Mineralo- and Glucocorticoid Synthesis
8. Epigenetic Control of CYP11B1
9. Epigenetic Modification of CYP11B1 in Cortisol-Producing Adenoma
10. Epigenetic Modification of CYP11B1 in Aldosterone-Producing Adenoma with Autonomous Cortisol Secretion
11. MicroRNAs (miRNAs) in Post-Transcriptional Regulation
12. Epigenesis of the Other Steroid Hormone Synthase Genes
12.1. Steroidogenic Acute Regulatory Protein (StAR)
12.2. Cytochrome P450 Family 11, Subfamily A, Polypeptide 1 (CYP11A1)
12.3. Aromatase (CYP19A1)
12.4. 17α-Hydroxylase (CYP17A1)
13. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ACE | angiotensin converting enzyme; |
| Ad | cis-acting element; |
| AMP | adenocine monophosphate; |
| ARB | AT1R blocker; |
| APA | aldosterone-producing adenoma; |
| APCC | aldosterone-producing cell cluster; |
| ATF | activating transcription factor; |
| AT1R | type2 angiotensin II receptor; |
| CACNA1D | Calcium Voltage-Gated Channel Subunit Alpha1 D |
| CEBP | CCAAT enhancer binding protein; |
| CPA | coerisol-producing adenoma |
| CREB | cyclic AMP responsive element binding protein; |
| CYP11B1 | 11β-hydroxylase |
| CYP11B2 | aldosterone synthetase; |
| GR | glucocorticoid receptor; |
| GRE | GR element; |
| HNF1A | hepatocyte nuclear factor1 homeobox A; |
| HPA | hypothalamic-pituitary-adrenal; |
| KCNJ | potassium inwardly rectifying channel subfamily J; |
| MBD | methyl-CpG-binding domain; |
| MECP | methyl-CpG-binding protein; |
| miRNA | microRNA; |
| Mi2 | chromodomain-helicase-DNA-binding protein Mi-2 homolog; |
| MR | mineralocorticoid receptor; |
| MRA | mineralocorticoid receptor antagonist; |
| NGFI-B | nerve growth factor-induced clone B; |
| NBRE-1 | NGFI-B response element; |
| NFA | non-functioning adenoma; |
| NR4A | nuclear receptor 4 group A; |
| NURR1 | nuclear receptor-related factor 1; |
| PA | primary aldosteronism; |
| RAS | renin-angiotensin system; |
| SETDB | histone-lysine N-methyltransferase; |
| Sin3A | SIN3 transcription regulator family member A; |
| STAT | signal transducer and activation transcription factor; |
| TSS | transcription start site. |
References
- Chong, C.; Hamid, A.; Yao, T.; Garza, A.E.; Pojoga, L.H.; Adler, G.K.; Romero, J.R.; Williams, G.H. Regulation of aldosterone secretion by mineralocorticoid receptor-mediated signaling. J. Endocrinol. 2017, 232, 525–534. [Google Scholar] [CrossRef] [PubMed]
- Miller, B.S.; Auchus, R.J. Evaluation and treatment of patients with hypercortisolism: A review. JAMA Surg. 2020, 155, 1152–1159. [Google Scholar] [CrossRef] [PubMed]
- Karaca, Z.; Grossman, A.; Kelestimur, F. Investigation of the hypothalamo-pituitary-adrenal (HPA) axis: A contemporary synthesis. Rev. Endocr. Metab. Disord. 2021, 22, 179–204. [Google Scholar] [CrossRef] [PubMed]
- Buffolo, F.; Tetti, M.; Mulatero, P.; Monticone, S. Aldosterone as a mediator of cardiovascular damage. Hypertension 2022, 79, 1899–1911. [Google Scholar] [CrossRef]
- Takeda, Y. Vascular synthesis of aldosterone: Role in hypertension. Mol. Cell Endocrinol. 2004, 217, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Sanchez, E.P.; Gomez-Sanchez, C.M.; Plonczynski, M.; Gomez-Sanchez, C.E. Aldosterone synthesis in the brain contributes to Dahl salt-sensitive rat hypertension. Exp. Physiol. 2010, 95, 120–130. [Google Scholar] [CrossRef]
- Briones, A.M.; Nguyen Dinh Cat, A.; Callera, G.E.; Yogi, A.; Burger, D.; He, Y.; Corrêa, J.W.; Gagnon, A.M.; Gomez-Sanchez, C.E.; Gomez-Sanchez, E.P.; et al. Adipocytes produce aldosterone through calcineurin-dependent signaling pathways: Implications in diabetes mellitus-associated obesity and vascular dysfunction. Hypertension 2012, 59, 1069–1078. [Google Scholar] [CrossRef]
- Mohamed, D.M.; Shaqura, M.; Li, X.; Shakibaei, M.; Beyer, A.; Treskatsch, S.; Schäfer, M.; Mousa, S.A. Aldosterone synthase in peripheral sensory neurons contributes to mechanical hypersensitivity during local inflammation in rats. Anesthesiology 2020, 132, 867–880. [Google Scholar] [CrossRef]
- Phan, T.S.; Schink, L.; Mann, J.; Merk, V.M.; Zwicky, P.; Mundt, S.; Simon, D.; Kulms, D.; Abraham, S.; Legler, D.F.; et al. Keratinocytes control skin immune homeostasis through de novo-synthesized glucocorticoids. Sci. Adv. 2021, 7, eabe0337. [Google Scholar] [CrossRef]
- Mueller, M.; Cima, I.; Noti, M.; Fuhrer, A.; Jakob, S.; Dubuquoy, L.; Schoonjans, K.; Brunner, T. The nuclear receptor LRH-1 critically regulates extra-adrenal glucocorticoid synthesis in the intestine. J. Exp. Med. 2006, 203, 2057–2062. [Google Scholar] [CrossRef]
- Miller, W.L. Steroidogenesis: Unanswered questions. Trends Endocrinol. Metab. 2017, 28, 771–793. [Google Scholar] [CrossRef] [PubMed]
- Schiffer, L.; Anderko, S.; Hannemann, F.; Eiden-Plach, A.; Bernhardt, R.J. The CYP11B subfamily. J. Steroid. Biochem. Mol. Biol. 2015, 151, 38–51. [Google Scholar] [CrossRef] [PubMed]
- Van de Wiel, E.; Chaman Baz, A.H.; Küsters, B.; Mukai, K.; van Bonzel, L.; van Erp, M.; Deinum, J.; Langenhuijsen, J. Changes of the CYP11B2 expressing zona glomerulosa in human adrenals from birth to 40 years of age. Hypertension 2022, 79, 2565–2572. [Google Scholar] [CrossRef] [PubMed]
- Rosenfield, R.L. Normal and premature adrenarche. Endocr. Rev. 2021, 42, 783–814. [Google Scholar] [CrossRef]
- Rainey, W. E Adrenal zonation: Clues from 11beta-hydroxylase and aldosterone synthase. Mol. Cell. Endocrinol. 1999, 151, 151–160. [Google Scholar] [CrossRef]
- Bassett, M.H.; Zhang, Y.; White, P.C.; Rainey, W.E. Regulation of human CYP11B2 and CYP11B1: Comparing the role of the common CRE/Ad1 element. Endocr. Res. 2000, 26, 941–951. [Google Scholar] [CrossRef]
- Sabari, B.R.; Zhang, D.; Allis, C.D.; Zhao, Y. Metabolic regulation of gene expression through histone acylations. Nat. Rev. Mol. Cell Biol. 2017, 18, 90–101. [Google Scholar] [CrossRef]
- Köhler, F.; Rodríguez-Paredes, M. DNA methylation in epidermal differentiation, aging, and cancer. J. Investig. Dermatol. 2020, 140, 38–47. [Google Scholar] [CrossRef]
- Ghanbarian, H.; Yıldız, M.T.; Tutar, Y. MicroRNA targeting. Methods Mol. Biol. 2022, 2257, 105–130. [Google Scholar]
- Salameh, Y.; Bejaoui, Y.; El Hajj, N. DNA methylation biomarkers in aging and age-related diseases. Front Genet. 2020, 11, 171. [Google Scholar] [CrossRef]
- Liang, M. Epigenetic mechanisms and hypertension. Hypertension 2018, 72, 1244–1254. [Google Scholar] [CrossRef]
- Demura, M.; Demura, Y.; Takeda, Y.; Saijoh, K. Dynamic regulation of the angiotensinogen gene by DNA methylation, which is influenced by various stimuli experienced in daily life. Hypertens. Res. 2015, 38, 519–527. [Google Scholar] [CrossRef]
- Parry, A.; Rulands, S.; Reik, W. Active turnover of DNA methylation during cell fate decisions. Nat. Rev. Genet. 2021, 22, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Stillman, B. Histone modifications: Insights into their influence on gene expression. Cell 2018, 175, 6–9. [Google Scholar] [CrossRef]
- McClure, J.J.; Li, X.; Chou, C.J. Advances and challenges of HDAC inhibitors in cancer therapeutics. Adv. Cancer Res. 2018, 138, 183–211. [Google Scholar] [PubMed]
- Gu, F.; Lin, Y.; Wang, Z.; Wu, X.; Ye, Z.; Wang, Y.; Lan, H. Biological roles of LSD1 beyond its demethylase activity. Cell Mol. Life Sci. 2020, 77, 3341–3350. [Google Scholar] [CrossRef]
- Huang, Y.; Ting, P.Y.; Yao, T.M.; Homma, T.; Brooks, D.; Katayama Rangel, I.; Adler, G.K.; Romero, J.R.; Williams, J.S.; Pojoga, L.H.; et al. Histone demethylase LSD1 deficiency and biological sex: Impact on blood pressure and aldosterone production. J Endocrinol. 2019, 240, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Miyamori, I.; Yoneda, T.; Hatakeyama, H.; Inaba, S.; Furukawa, K.; Mabuchi, H.; Takeda, R. Regulation of aldosterone synthase in human vascular endothelial cells by angiotensin II and adrenocorticotropin. J. Clin. Endocrinol. Metab. 1996, 81, 2797–2800. [Google Scholar]
- Guagliardo, N.A.; Klein, P.M.; Gancayco, C.A.; Lu, A.; Leng, S.; Makarem, R.R.; Cho, C.; Rusin, C.G.; Breault, D.T.; Barrett, P.Q.; et al. Angiotensin II induces coordinated calcium bursts in aldosterone-producing adrenal rosettes. Nat. Commun. 2020, 11, 1679. [Google Scholar] [CrossRef]
- Ali, Y.; Dohi, K.; Okamoto, R.; Katayama, K.; Ito, M. Novel molecular mechanisms in the inhibition of adrenal aldosterone synthesis: Action of tolvaptan via vasopressin V2 receptor-independent pathway. Br. J. Pharmacol. 2019, 176, 1315–1327. [Google Scholar] [CrossRef]
- Vanderriele, P.E.; Caroccia, B.; Seccia, T.M.; Piazza, M.; Lenzini, L.; Torresan, F.; Iacobone, M.; Unger, T.; Rossi, G.P. The angiotensin type 2 receptor in the human adrenocortical zona glomerulosa and in aldosterone-producing adenoma: Low expression and no functional role. Clin. Sci. 2018, 132, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Lauber, M.; Boni-Schnetzler, M.; Miiller, J. Potassium raises cytochrome P-450 mRNA level in zona glomerulosa of rat adrenals. Mol. Cell Endocrinol. 1990, 72, 159–166. [Google Scholar] [PubMed]
- Holland, O.B.; Carr, B. Modulation of aldosterone synthase messenger ribonucleic acid levels by dietary sodium and potassium and by adrenocorticotropin. Endocrinology 1993, 132, 2666–2673. [Google Scholar] [CrossRef] [PubMed]
- Peters, B.; Teubner, P.; Clausmeyer, S.; Puschner, T.; Maser-Gluth, C.; Wrede, H.J.; Kränzlin, B.; Peters, J. StAR expression and the long-term aldosterone response to high-potassium diet in Wistar-Kyoto and spontaneously hypertensive rats. Am. J. Physiol. Endocrinol. Metab. 2007, 292, E16–E23. [Google Scholar] [CrossRef]
- Demura, M.; Wang, F.; Yoneda, T.; Karashima, S.; Mori, S.; Oe, M.; Kometani, M.; Sawamura, T.; Cheng, Y.; Maeda, Y.; et al. Multiple noncoding exons 1 of nuclear receptors NR4A family (nerve growth factor-induced clone B, Nur-related factor 1 and neuron-derived orphan receptor 1) and NR5A1 (steroidogenic factor 1) in human cardiovascular and adrenal tissues. J. Hypertens. 2011, 29, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Dierks, A.D.; Urs, D.; Lichtenauer, U.D.; Sackmann, S.; Spyroglou, A.; Shapiro, I.; Geyer, M.; Manonopoulou, J.; Reincke, M.; Hantel, C.; et al. Identification of adrenal genes regulated in a potassium-dependent manner. J. Mol. Endocrinol. 2010, 45, 193–206. [Google Scholar] [CrossRef]
- Takeda, Y.; Demura, M.; Wang, F.; Karashima, S.; Yoneda, T.; Kometani, M.; Hashimoto, A.; Aono, D.; Horike, S.; Meguro-Horike, M.; et al. Epigenetic regulation of aldosterone synthase gene by sodium and angiotensin II. J. Am. Heart Assoc. 2018, 7, e008281. [Google Scholar] [CrossRef]
- Wang, F.; Demura, M.; Cheng, Y.; Zhu, A.; Karashima, S.; Yoneda, T.; Demura, Y.; Maeda, Y.; Namiki, M.; Ono, K.; et al. Dynamic CCAAT/enhancer binding protein-associated changes of DNA methylation in the angiotensinogen gene. Hypertension 2014, 63, 281–288. [Google Scholar] [CrossRef]
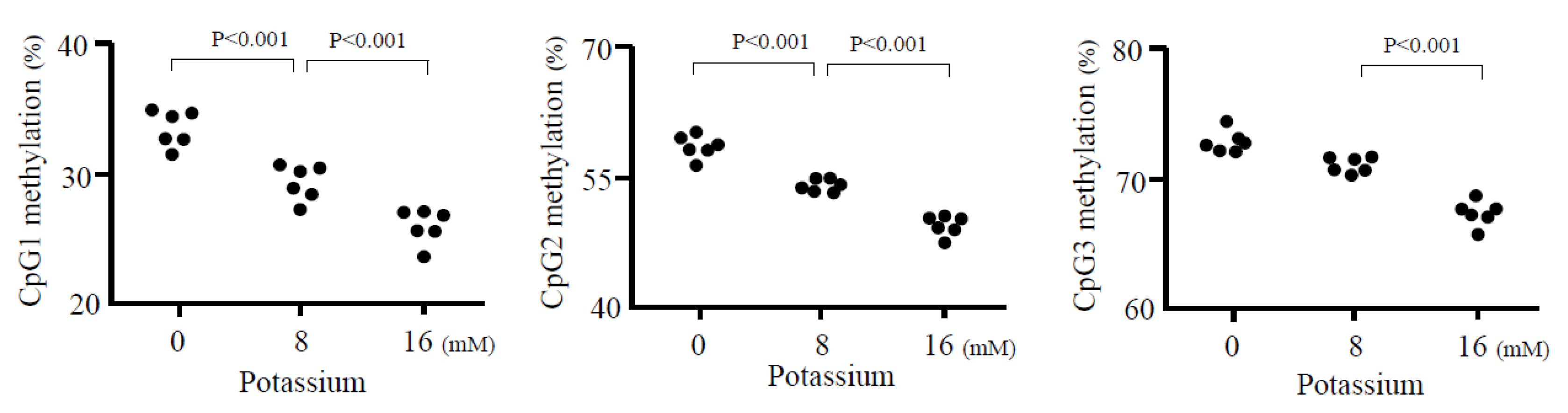
- Takeda, Y.; Demura, M.; Wang, F.; Karashima, S.; Yoneda, T.; Kometani, M.; Aomo, D.; Hashimoto, A.; Horike, S.; Meguro-Horike, M.; et al. Effect of potassium on DNA methylation of aldosterone synthase gene. J. Hypertens. 2021, 39, 1018–1024. [Google Scholar] [CrossRef]
- Edwards, J.R.; Yarychkivska, O.; Boulard, M.; Bestor, T.H. DNA methylation and DNA methyltransferases. Epigenetics Chromatin. 2017, 8, 10–23. [Google Scholar] [CrossRef]
- Nogueira, E.F.; Vargas, C.A.; Otis, M.; Gallo-Payet, N.G.; Bollag, W.B.; Rainey, W.E. Angiotensin-II acute regulation of rapid response genes in human, bovine, and rat adrenocortical cells. J. Molec. Endocrinol. 2007, 39, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, K.; Harris, R.B.S.; Rainey, W.E.; Seki, T. Sodium deficiency regulates rat adrenal zona glomerulosa gene expression. Endocrinology 2014, 155, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Carey, R.M. Aldosterone and cardiovascular disease. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 194–198. [Google Scholar] [CrossRef]
- Reincke, M.; Bancos, I.; Mulatero, P.; Scholl, U.I.; Stowasser, M.; Williams, T.A. Diagnosis and treatment of primary aldosteronism. Lancet Diabetes Endocrinol. 2021, 9, 876–892. [Google Scholar] [CrossRef]
- Lyubarova, R.; Gosmanova, E.O. Mineralocorticoid receptor blockade in end-stage renal disease. Curr. Hypertens. Rep. 2017, 19, 40. [Google Scholar] [CrossRef]
- Gao, X.; Yamazaki, Y.; Tezuka, Y.; Omata, K.; Ono, Y.; Morimoto, R.; Nakamura, Y.; Suzuki, T.; Satoh, F.; Sasano, H. Pathology of Aldosterone Biosynthesis and its Action. Tohoku J. Exp. Med. 2021, 254, 1–15. [Google Scholar] [CrossRef]
- Mogi, M. Aldosterone breakthrough from a pharmacological perspective. Hypertens Res. 2022, 45, 967–997. [Google Scholar] [CrossRef]
- Vergaro, G.; Passino, C.; Emdin, M.J. No aldosterone breakthrough with the neprilysin inhibitor sacubitril. Am. Coll. Cardiol. 2019, 73, 3037–3038. [Google Scholar] [CrossRef]
- Takeda, Y. Effects of eplerenone, a selective mineralocorticoid receptor antagonist, on clinical and experimental salt-sensitive hypertension. Hypertens. Res. 2009, 32, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, A.; Takeda, Y.; Karashima, S.; Kometani, M.; Aono, D.; Higashikata, T.; Konishi, S.; Yoneda, T.; Takeda, Y. Impact of mineralocorticoid receptor blockade with direct renin inhibition in angiotensin II-dependent hypertensive mice. Hypertens. Res. 2020, 43, 1099–1104. [Google Scholar] [CrossRef]
- Hughes-Austin, J.M.; Rifkin, D.E.; Beben, T.; Katz, R.; Sarnak, M.J.; Deo, R.; Hoofnagle, A.N.; Homma, S.; Siscovick, D.S.; Sotoodehnia, N.; et al. The relation of serum potassium concentration with cardiovascular events and mortality in community-living individuals. Clin. J. Am. Soc. Nephrol. 2017, 12, 245–252. [Google Scholar] [CrossRef]
- Weir, M.R.; Bushinsky, D.A.; Benton, W.W.; Woods, S.D.; Mayo, M.R.; Arthur, S.P.; Pitt, B.; Bakris, G.L. Effect of patiromer on hyperkalemia recurrence in older chronic kidney disease patients taking RAAS inhibitors. Am. J. Med. 2018, 131, 555–564. [Google Scholar] [CrossRef]
- Sakthiswary, R.; Wong, M.; Nor Azmi, K. Spot urine potassium as a potential screening test for aldosterone breakthrough. Clin. Ter. 2012, 163, 195–198. [Google Scholar]
- Vaidya, A.; Mulatero, P.; Baudrand, R.; Adler, G.K. The expanding spectrum of primary aldosteronism: Implications for diagnosis, pathogenesis, and treatment. Endocr Rev. 2018, 39, 1057–1088. [Google Scholar] [CrossRef]
- Funder, J.W.; Carey, R.M. Primary aldosteronism: Where are we now? Where to from here? Hypertension 2022, 79, 726–735. [Google Scholar] [CrossRef]
- Howard, B.; Wang, Y.; Xekouki, P.; Faucz, F.R.; Jain, M.; Zhang, L.; Meltzer, P.G.; Stratakis, C.A.; Kebebew, E. Integrated analysis of genome-wide methylation and gene expression shows epigenetic regulation of CYP11B2 in aldosteronomas. J. Clin. Endocrinol. Metab. 2014, 99, E536–E543. [Google Scholar] [CrossRef] [PubMed]
- Takeda, Y.; Demura, N.; Yoneda, T.; Takeda, Y. DNA Methylation of the Angiotensinogen Gene, AGT, and the Aldosterone Synthase Gene, CYP11B2 in Cardiovascular Diseases. Int. J. Mol. Sci. 2021, 22, 4587. [Google Scholar] [CrossRef]
- Di Dalmazi, G.; Morandi, L.; Rubin, B.; Pilon, C.; Asioli, S.; Vicennati, V.; De Leo, A.; Ambrosi, F.; Santini, D.; Pagotto, U.; et al. DNA methylation of steroidogenic enzymes in benign adrenocortical tumors: New insights in aldosterone-producing adenomas. J. Clin. Endocrinol. Metab. 2020, 105, dgaa585. [Google Scholar] [CrossRef] [PubMed]
- Yoshii, Y.; Oki, K.; Gomez-Sanchez, C.E.; Ohno, H.; Itcho, K.; Kobuke, K.; Yoneda, M. Hypomethylation of CYP11B2 in aldosterone-producing adenoma. Hypertension 2016, 68, 1432–1437. [Google Scholar] [CrossRef] [PubMed]
- Nishimoto, K.; Seki, T.; Kurihara, I.; Yokota, K.; Omura, M.; Nishikawa, T.; Shibata, H.; Kosaka, T.; Oya, M.; Suematsu, M.; et al. Case report: Nodule development from subcapsular aldosterone producing cell clusters causes hyperaldosteronism. J. Clin. Endocrinol. Metab. 2016, 101, 6–9. [Google Scholar] [CrossRef]
- Murakami, M.; Yoshimoto, T.; Nakabayashi, K.; Nakano, Y.; Fukaishi, T.; Tsuchiya, K.; Minami, I.; Bouchi, R.; Okamura, K.; Fujii, Y.; et al. Molecular characteristics of the KCNJ5 mutated aldosterone-producing adenomas. Endocr. Relat. Cancer. 2017, 24, 531–541. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, K.; Boulkroun, S.; Baron, S.; Nanba, K.; Wack, M.; Rainey, W.E.; Rocha, A.; Giscos-Douriez, I.; Meatchi, T.; Amar, L.; et al. Genetic, cellular, and molecular heterogeneity in adrenals with aldosterone-p ne-producing adenoma. Hypertension 2020, 75, 1034–1044. [Google Scholar] [CrossRef] [PubMed]
- Omata, K.; Anand, S.K.; Hovelson, D.H.; Liu, C.J.; Yamazaki, Y.; Nakamura, Y.; Ito, S.; Satoh, F.; Sasano, H.; Rainey, W.E.; et al. Aldosterone-producing cell clusters frequently harbor somatic mutations and accumulate with age in mormal adrenals. J. Endocr. Soc. 2017, 1, 787–799. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Meyer, L.S.; Feuchtinger, A.; Kunzke, T.; Knösel, T.; Reincke, M.; Walch, A.; Williams, T.A. Mass spectrometry imaging establishes 2 distinct metabolic phenotypes of aldosterone-producing cell clusters in primary aldosteronism. Hypertension 2020, 75, 634–644. [Google Scholar] [CrossRef]
- Rossi, G.P. Primary aldosteronism: JACC state-of-the-art review. J. Am. Coll. Cardiol. 2019, 74, 2799–2811. [Google Scholar] [CrossRef]
- Vaidya, A.; Hundemer, G.L.; Nanba, K.; Parksook, W.W.; Brown, J.M. Primary aldosteronism: State-of-the-art review. Am. J. Hypertens. 2022, 35, 967–988. [Google Scholar] [CrossRef]
- Fischer, E.; Beuschlein, F.; Degenhart, C.; Jung, P.; Bidlingmaier, M.; Reincke, M. Spontaneous remission of idiopathic aldosteronism after long-term treatment with spironolactone: Results from the German Conn’s Registry. Clin. Endocrinol. 2012, 76, 473–477. [Google Scholar] [CrossRef]
- Yoneda, T.; Demura, M.; Takata, H.; Kometani, M.; Karashima, S.; Yamagishi, M.; Takeda, Y. Unilateral primary aldosteronism with spontaneous remission after long-term spironolactone therapy. J. Clin. Endocrinol. Metab. 2012, 97, 1109–1113. [Google Scholar] [CrossRef]
- Ye, P.; Yamashita, T.; Pollock, D.M.; Sasano, H.; Rainey, W.E. Contrasting effects of eplerenone and spironolactone on adrenal cell steroidogenesis. Horm. Metab. Res. 2009, 41, 35–39. [Google Scholar] [CrossRef]
- MacKenzie, S.M.; Connell, J.M.; Davies, E. Non-adrenal synthesis of aldosterone: A reality check. Mol. Cell. Endocrinol. 2012, 350, 163–167. [Google Scholar] [CrossRef]
- Takeda, Y. Role of cardiovascular aldosterone in hypertension. Curr. Med. Chem. Cardiovasc. Hematol. Agents. 2005, 3, 261–266. [Google Scholar] [CrossRef]
- Fujisaki, M.; Nagoshi, T.; Nishikawa, T.; Date, T.; Yoshimura, M. Rapid induction of aldosterone synthesis in cultured neonatal rat cardiomyocytes under high glucose conditions. Biomed. Res. Int. 2013, 2013, 161396. [Google Scholar] [CrossRef] [PubMed]
- Takata, H.; Takeda, Y.; Zhu, A.; Cheng, Y.; Yoneda, T.; Demura, M.; Yagi, K.; Karashima, S.; Yamagishi, M. Protective effects of mineralocorticoid receptor blockade against neuropathy in experimental diabetic rats. Diabetes Obes. Metab. 2012, 14, 155–162. [Google Scholar] [CrossRef]
- Tsybouleva, N.; Zhang, L.; Chen, S.; Patel, R.; Lutucuta, S.; Nemoto, S.; De Freitas, G.; Entman, M.; Carabello, B.A.; Roberts, R.; et al. Aldosterone, through novel signaling proteins, is a fundamental molecular bridge between the genetic defect and the cardiac phenotype of hypertrophic cardiomyopathy. Circulation 2009, 109, 1284–1291. [Google Scholar] [CrossRef]
- Takeda, Y.; Yoneda, T.; Demura, M.; Usukura, M.; Mabuchi, H. Calcineurin inhibition attenuates mineralocorticoid-induced cardiac hypertrophy. Circulation 2002, 105, 677–679. [Google Scholar] [CrossRef] [PubMed]
- Garnier, A.; Bendall, J.K.; Fuchs, S.; Escoubet, B.; Rochais, F.; Hoerter, J.; Delcayre, C. Cardiac specific increase in aldosterone production induces coronary dysfunction in aldosterone synthase-transgenic mice. Circulation 2004, 110, 1819–1825. [Google Scholar] [CrossRef] [PubMed]
- Alesutan, I.; Voelkl, J.; Feger, M.; Kratschmar, D.V.; Castor, T.; Mia, S.; Sacherer, M.; Viereck, R.; Borst, O.; Leibrock, C.; et al. Involvement of vascular aldosterone synthase in phosphate-induced osteogenic transformation of vascular smooth muscle cells. Sci. Rep. 2017, 7, 2059. [Google Scholar] [CrossRef]
- Takeda, Y.; Yoneda, T.; Demura, M.; Miyamori, I.; Mabuchi, H. Cardiac aldosterone production in genetically hypertensive rats. Hypertension 2000, 36, 495–500. [Google Scholar] [CrossRef]
- Yoshimura, M.; Nakamura, S.; Ito, T.; Nakayama, M.; Harada, E.; Mizuno, Y.; Sakamoto, T.; Yamamuro, M.; Saito, Y.; Nakao, K.; et al. Expression of aldosterone synthase gene in failing human heart: Quantitative analysis using modified real-time polymerase chain reaction. J. Clin. Endocrinol. Metab. 2002, 87, 3936–3940. [Google Scholar] [CrossRef]
- Kater, C.E.; Giorgi, R.B.; Costa-Barbosa, F.A. Classic and current concepts in adrenal steroidogenesis: A reappraisal. Arch. Endocrinol. Metab. 2022, 66, 77–87. [Google Scholar] [CrossRef]
- Slominski, R.M.; Tuckey, R.C.; Manna, P.R.; Jetten, A.M.; Postlethwaite, A.; Raman, C.; Slominski, A.T. Extra-adrenal glucocorticoid biosynthesis: Implications for autoimmune and inflammatory disorders. Genes. Immun. 2020, 21, 150–168. [Google Scholar] [CrossRef]
- Takeda, Y.; Miyamori, I.; Yoneda, T.; Iki, K.; Hatakeyama, H.; Blair, I.A.; Hsieh, F.Y.; Takeda, R. Synthesis of corticosterone in the vascular wall. Endocrinology 1994, 135, 2283–2286. [Google Scholar] [CrossRef] [PubMed]
- Salehzadeh, M.; Soma, K. K Glucocorticoid production in the thymus and brain: Immunosteroids and neurosteroids. Brain Behav Immun. Health. 2021, 18, 100352. [Google Scholar] [CrossRef]
- Bassett, M.H.; Mayhew, B.; Rehman, K.; White, P.C.; Mantero, F.; Arnaldi, G.; Stewart, P.M.; Bujalska, I.; Rainey, W.E. Expression profiles for steroidogenic enzymes in adrenocortical disease. J. Clin. Endocrinol. Metab. 2005, 90, 5446–5455. [Google Scholar] [CrossRef] [PubMed]
- Kubota-Nakayama, F.; Nakamura, Y.; Konosu-Fukaya, S.; Azmahani, A.; Ise, K.; Yamazaki, Y.; Kitawaki, Y.; Felizola, S.J.; Ono, Y.; Omata, K.; et al. Expression of steroidogenic enzymes and their transcription factors in cortisol-producing adrenocortical adenomas: Immunohistochemical analysis and quantitative real-time polymerase chain reaction studies. Hum. Pathol. 2016, 54, 165–173. [Google Scholar] [CrossRef]
- Ahn, C.H.; Na, H.Y.; Park, S.Y.; Yu, H.W.; Kim, S.J.; Choi, J.Y.; Lee, K.E.; Kim, S.W.; Jung, K.C.; Kim, J.H. Expression of CYP11B1 and CYP11B2 in adrenal adenoma correlates with clinical characteristics of primary aldosteronism. Clin. Endocrinol. 2022, 96, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, X.-D.; Vaheri, A.; Voutilainen, R. DNA methylation affects cell proliferation, cortisol secretion and steroidogenic gene expression in human adrenocortical NCI-H295R cells. J. Mol. Endocrinol. 2004, 33, 651–662. [Google Scholar] [CrossRef]
- Kometani, M.; Yoneda, T.; Demura, M.; Koide, H.; Nishimoto, K.; Mukai, K.; Gomez-Sanchez, C.E.; Akagi, T.; Yokota, T.; Horike, S.I.; et al. Cortisol overproduction results from DNA methylation of CYP11B1 in hypercortisolemia. Sci. Rep. 2017, 7, 11205. [Google Scholar] [CrossRef]
- Ronchi, C.L.; Di Dalmazi, G.; Faillot, S.; Sbiera, S.; Assié, G.; Weigand, I.; Calebiro, D.; Schwarzmayr, T.; Appenzeller, S.; Rubin, B.; et al. Genetic landscape of sporadic unilateral adrenocortical adenomas without PRKACA p.leu 206arg mutation. J. Clin. Endocrinol. Metab. 2016, 101, 3526–3538. [Google Scholar] [CrossRef]
- Yanase, T.; Oki, Y.; Katabami, T.; Otsuki, M.; Kageyama, K.; Tanaka, T.; Kawate, H.; Tanabe, M.; Doi, M.; Akehi, Y.; et al. New diagnostic criteria of adrenal subclinical Cushing’s syndrome: Opinion from the Japan Endocrine Society. Endocr. J. 2018, 65, 83–393. [Google Scholar] [CrossRef]
- Abe, I.; Sugimoto, K.; Miyajima, T.; Ide, T.; Minezaki, M.; Takeshita, K.; Takahara, S.; Nakagawa, M.; Fujimura, Y.; Kudo, T.; et al. Clinical investigation of adrenal incidentalomas in Japanese patients of the fukuoka Rregion with updated diagnostic criteria for sub-clinical Cushing’s syndrome. Intern. Med. 2018, 57, 2467–2472. [Google Scholar] [CrossRef] [PubMed]
- Ichijo, T.; Ueshiba, H.; Nawata, H.; Yanase, T. A nationwide survey of adrenal incidentalomas in Japan: The first report of clinical and epidemiological features. Endocr. J. 2020, 67, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Katabami, T.; Matsuba, R.; Kobayashi, H.; Nakagawa, T.; Kurihara, I.; Ichijo, T.; Tsuiki, M.; Wada, N.; Ogawa, Y.; Sone, M.; et al. Primary aldosteronism with mild autonomous cortisol secretion increases renal complication risk. Eur. J. Endocrinol. 2022, 186, 645–655. [Google Scholar] [CrossRef]
- Arlt, W.; Lang, K.; Sitch, A.J.; Dietz, A.S.; Rhayem, Y.; Bancos, I.; Feuchtinger, A.; Chortis, V.; Gilligan, L.C.; Ludwig, P.; et al. Steroid metabolome analysis reveals prevalent glucocorticoid excess in primary aldosteronism. JCI Insight 2017, 2, e93136. [Google Scholar] [CrossRef] [PubMed]
- Akehi, Y.; Yanase, T.; Motonaga, R.; Umakoshi, H.; Tsuiki, M.; Takeda, Y.; Yoneda, T.; Kurihara, I.; Itoh, H.; Katabami, T.; et al. Japan Primary Aldosteronism Study Group. High prevalence of diabetes in patients with primary aldosteronism (PA) associated with subclinical hypercortisolism and prediabetes more prevalent in bilateral than unilateral PA: A large, multicenter cohort study in Japan. Diabetes Care 2019, 42, 938–945. [Google Scholar]
- Inoue, K.; Horikoshi, H.; Omura, M.; Tsurutani, Y.; Saito, J.; Nishikawa, T. Association between aldosterone and hypertension among patients with overt and subclinical hypercortisolism. J. Endoc. Soc. 2023, 7, bvac167. [Google Scholar] [CrossRef]
- Nusrin, S.; Tong, S.K.; Chaturvedi, G.; Wu, R.S.; Giesy, J.P.; Kong, R.Y. Regulation of CYP11B1 and CYP11B2 steroidogenic genes by hypoxia-inducible miR-10b in H295R cells. Mar. Pollut. Bull. 2014, 85, 344–351. [Google Scholar] [CrossRef]
- Robertson, S.; MacKenzie, S.M.; Alvarez-Madrazo, S.; Diver, L.A.; Lin, J.; Stewart, P.M.; Fraser, R.; Connell, J.M.; Davies, E. MicroRNA-24 is a novel regulator of aldosterone and cortisol production in the human adrenal cortex. Hypertension 2013, 62, 572–578. [Google Scholar] [CrossRef]
- Lenzini, L.; Caroccia, B.; Campos, A.G.; Fassina, A.; Belloni, A.S.; Seccia, T.M.; Kuppusamy, M.; Ferraro, S.; Skander, G.; Bader, M.; et al. Lower expression of the TWIK-related acid-sensitive K+ channel 2 (TASK-2) gene is a hallmark of aldosterone-producing adenoma causing human primary aldosteronism. J. Clin. Endocrinol. Metab. 2014, 99, E674–E682. [Google Scholar] [CrossRef]
- Vetrivel, S.; Zhang, R.; Engel, M.; Oßwald, A.; Watts, D.; Chen, A.; Wielockx, B.; Sbiera, S.; Reincke, M.; Riester, A. Characterization of adrenal miRNA-based dysregulations in Cushing’s syndrome. Int. J. Mol. Sci. 2022, 23, 7676. [Google Scholar] [CrossRef]
- Sugino, N. Molecular mechanisms of luteinization. Obstet. Gynecol. Sci. 2014, 57, 93–101. [Google Scholar] [CrossRef]
- Okada, M.; Lee, L.; Maekawa, R.; Sato, S.; Kajimura, T.; Shinagawa, M.; Tamura, T.; Taketani, T.; Asada, H.; Tamura, H.; et al. Epigenetic changes of the cyp11a1 promoter region in granulosa cells undergoing luteinization during ovulation in female rats. Endocrinology 2016, 157, 3344–3354. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Yan, L.; Qiao, J. Increased expression of P450scc and CYP17 in development of endogenous hyperandrogenism in a rat model of PCOS. Endocrine 2013, 43, 184–190. [Google Scholar] [CrossRef]
- Krasic, J.; Fucic, A.; Sincic, N.; Sindicic Dessardo, N.; Starcevic, M.; Guszak, V.; Ceppi, M.; Bruzzone, M.; Kralik, S. Comparison of estradiol, testostosterone, and CYP19 methylation levels between full-term and preterm newborns. Horm. Res. Paediatr. 2021, 94, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Spitschak, M.; Vanselow, J. Bovine large luteal cells show increasing de novo DNA methylation of the main ovarian CYP19A1 promoter P2. Gen. Comp. Endocrinol. 2012, 178, 7–45. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; He, F.; Wen, H.; Li, J.; Ni, M.; Chi, M.; Qian, K.; Bu, Y.; Zhang, D.; Si, Y.; et al. DNA methylation status of cyp17-II gene correlated with its expression pattern and reproductive endocrinology during ovarian development stages of Japanese flounder (Paralichthys olivaceus). Gene 2013, 527, 82–88. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takeda, Y.; Demura, M.; Kometani, M.; Karashima, S.; Yoneda, T.; Takeda, Y. Molecular and Epigenetic Control of Aldosterone Synthase, CYP11B2 and 11-Hydroxylase, CYP11B1. Int. J. Mol. Sci. 2023, 24, 5782. https://doi.org/10.3390/ijms24065782
Takeda Y, Demura M, Kometani M, Karashima S, Yoneda T, Takeda Y. Molecular and Epigenetic Control of Aldosterone Synthase, CYP11B2 and 11-Hydroxylase, CYP11B1. International Journal of Molecular Sciences. 2023; 24(6):5782. https://doi.org/10.3390/ijms24065782
Chicago/Turabian StyleTakeda, Yoshimichi, Masashi Demura, Mitsuhiro Kometani, Shigehiro Karashima, Takashi Yoneda, and Yoshiyu Takeda. 2023. "Molecular and Epigenetic Control of Aldosterone Synthase, CYP11B2 and 11-Hydroxylase, CYP11B1" International Journal of Molecular Sciences 24, no. 6: 5782. https://doi.org/10.3390/ijms24065782
APA StyleTakeda, Y., Demura, M., Kometani, M., Karashima, S., Yoneda, T., & Takeda, Y. (2023). Molecular and Epigenetic Control of Aldosterone Synthase, CYP11B2 and 11-Hydroxylase, CYP11B1. International Journal of Molecular Sciences, 24(6), 5782. https://doi.org/10.3390/ijms24065782