
Insights on Phytohormonal Crosstalk in Plant Response to Nitrogen Stress: A Focus on Plant Root Growth and Development



Abstract
:1. Introduction
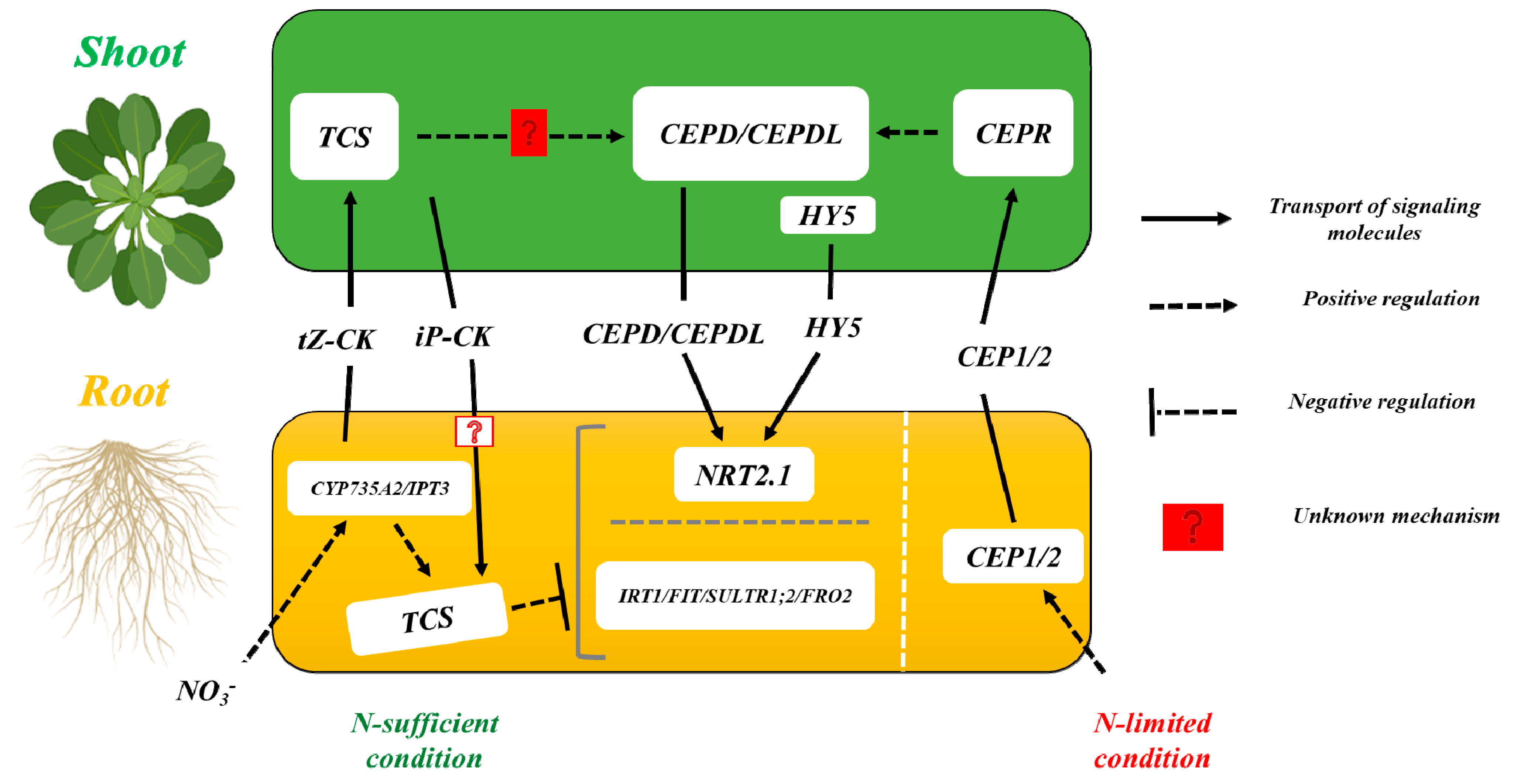
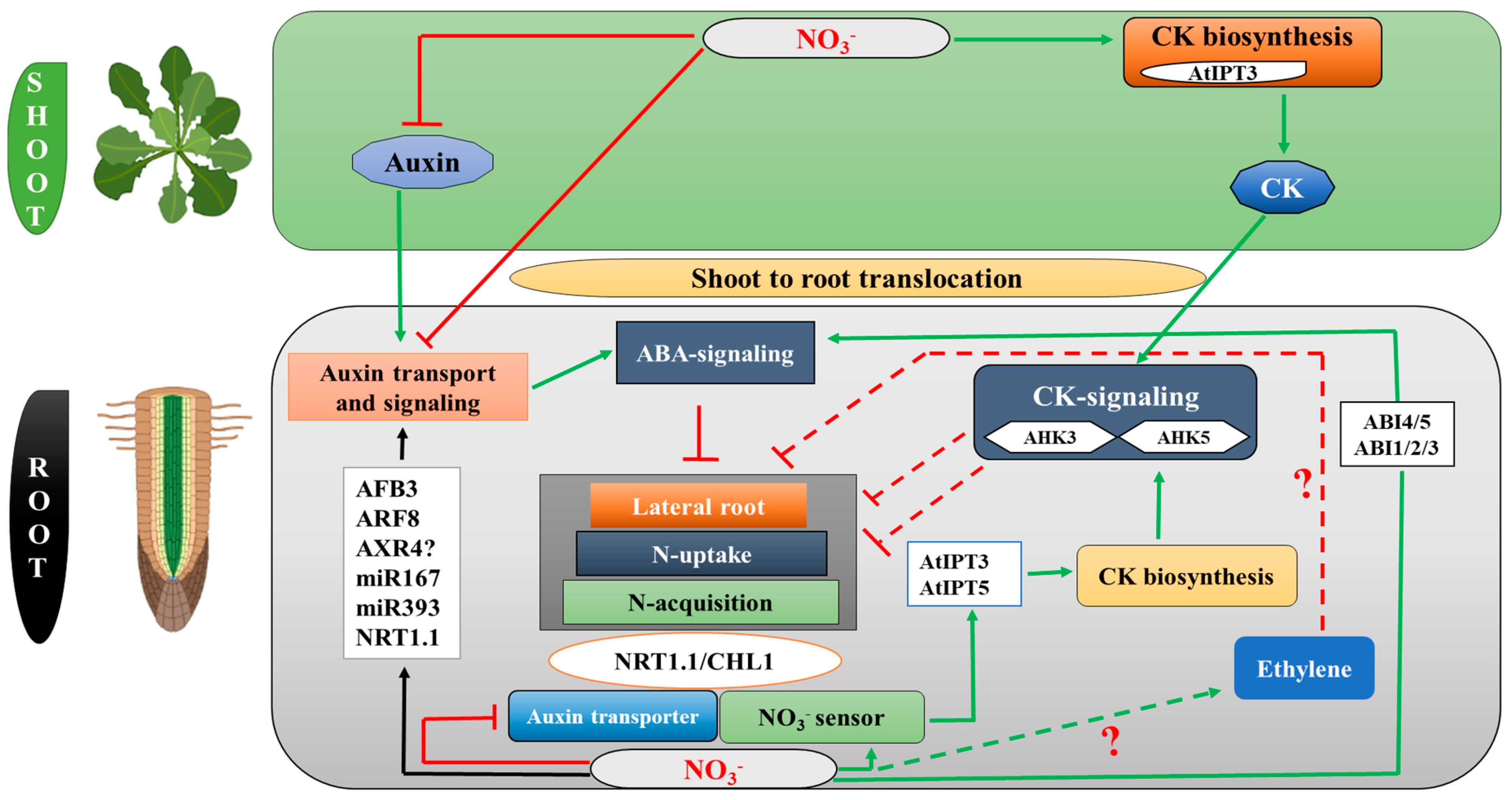
2. Cytokinin (CK) Modulates RSA in Response to Nitrogen Stress
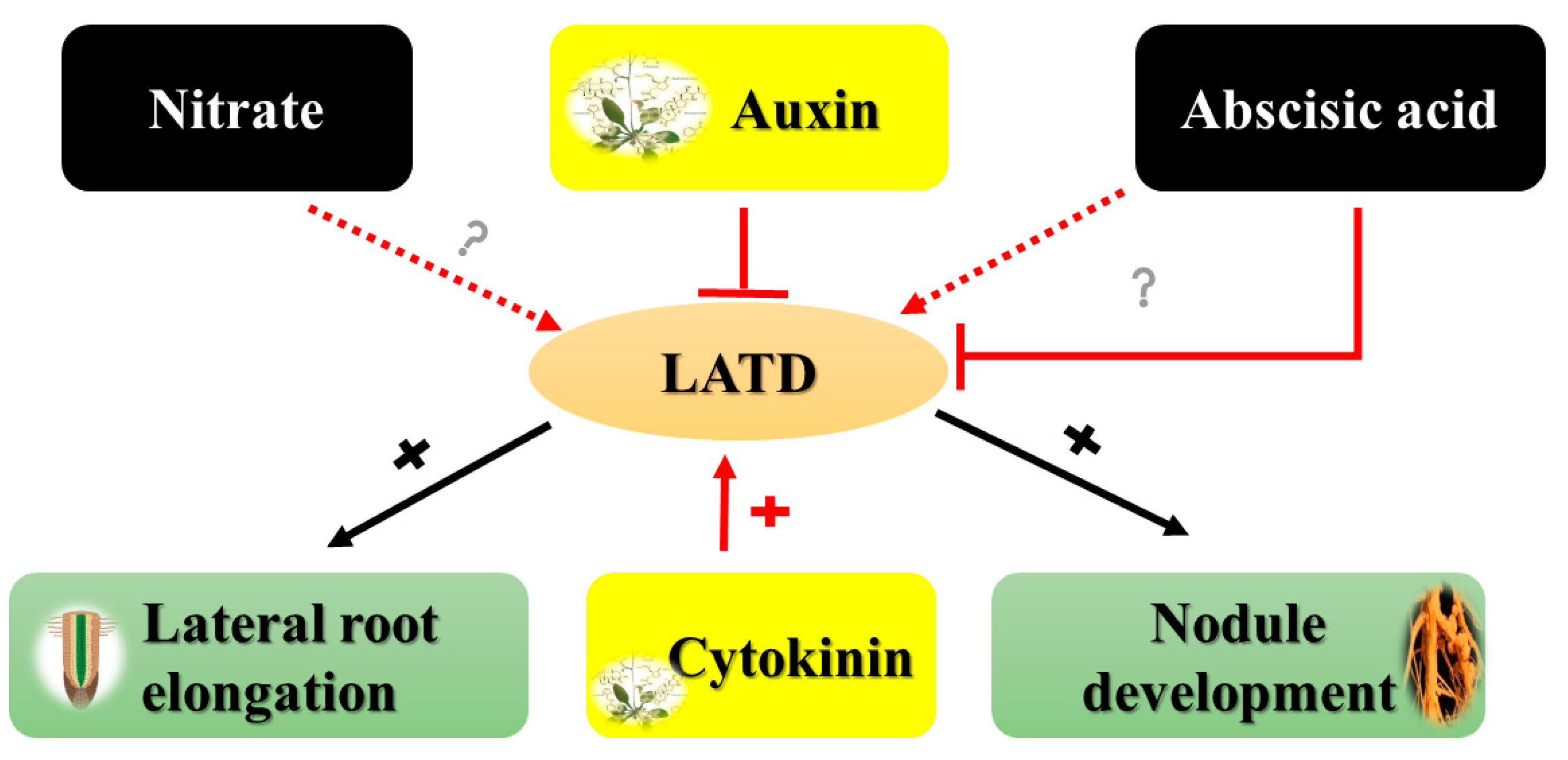
3. Abscisic Acid (ABA) Modulates RSA in Response to Nitrogen Stress
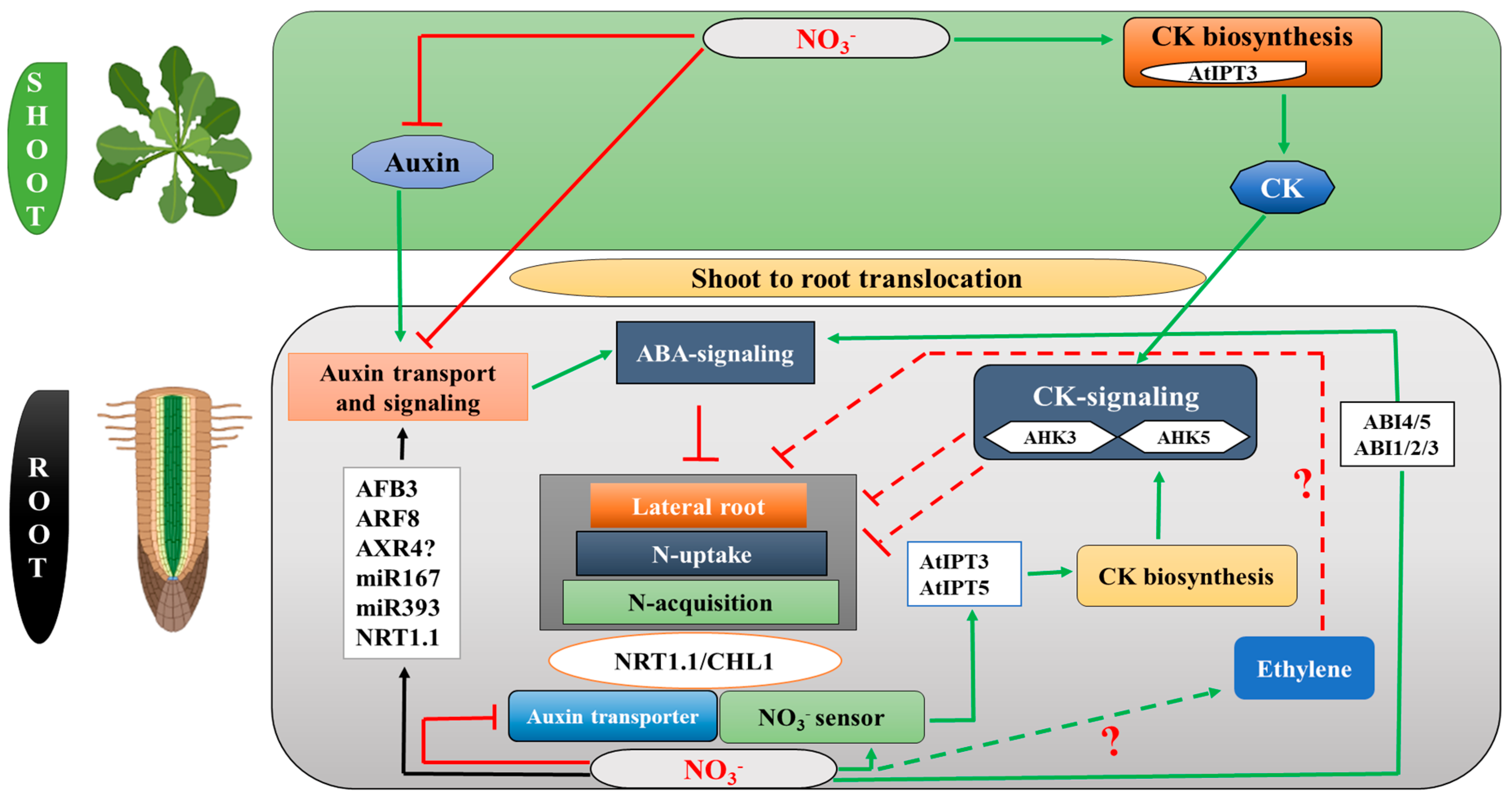
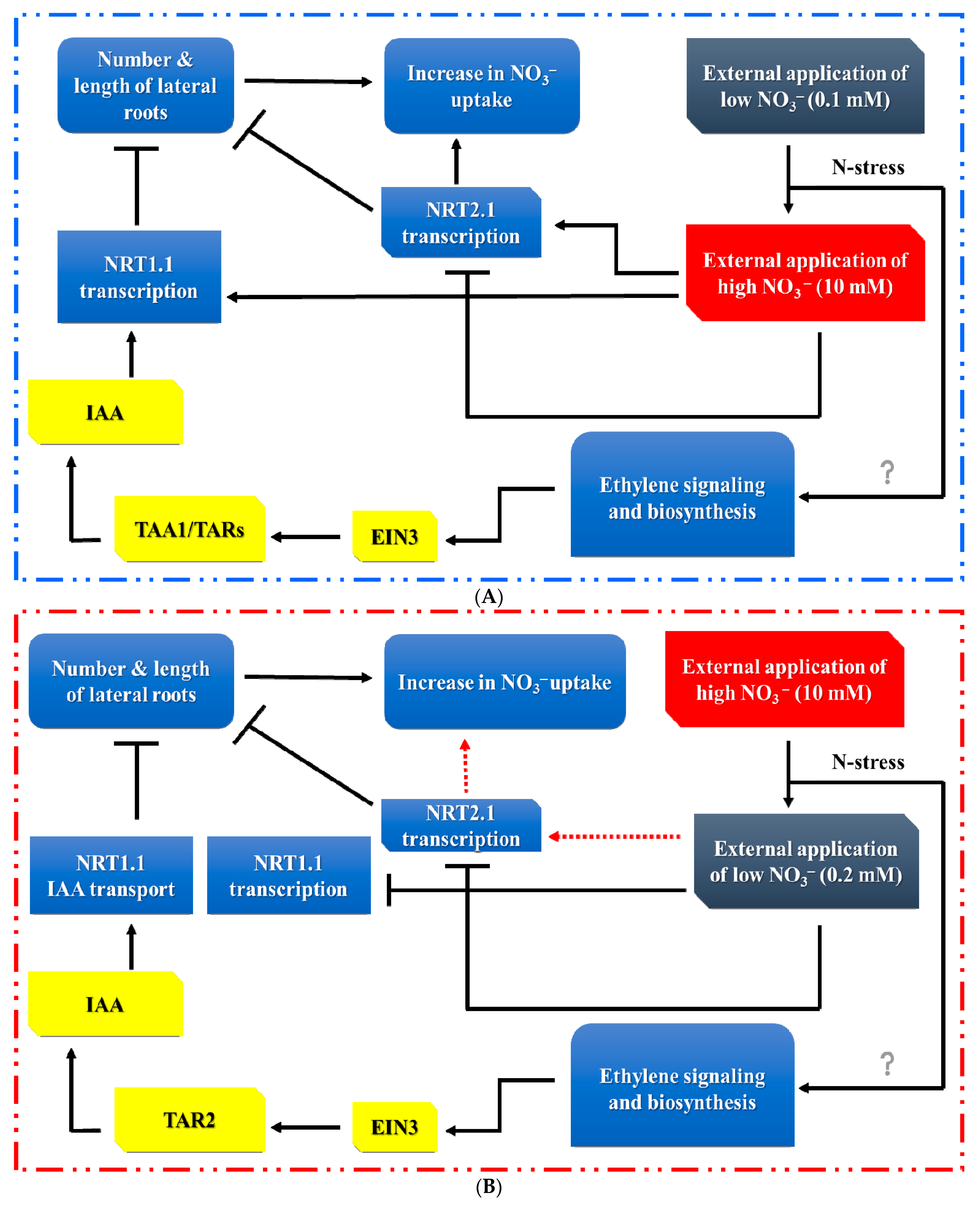
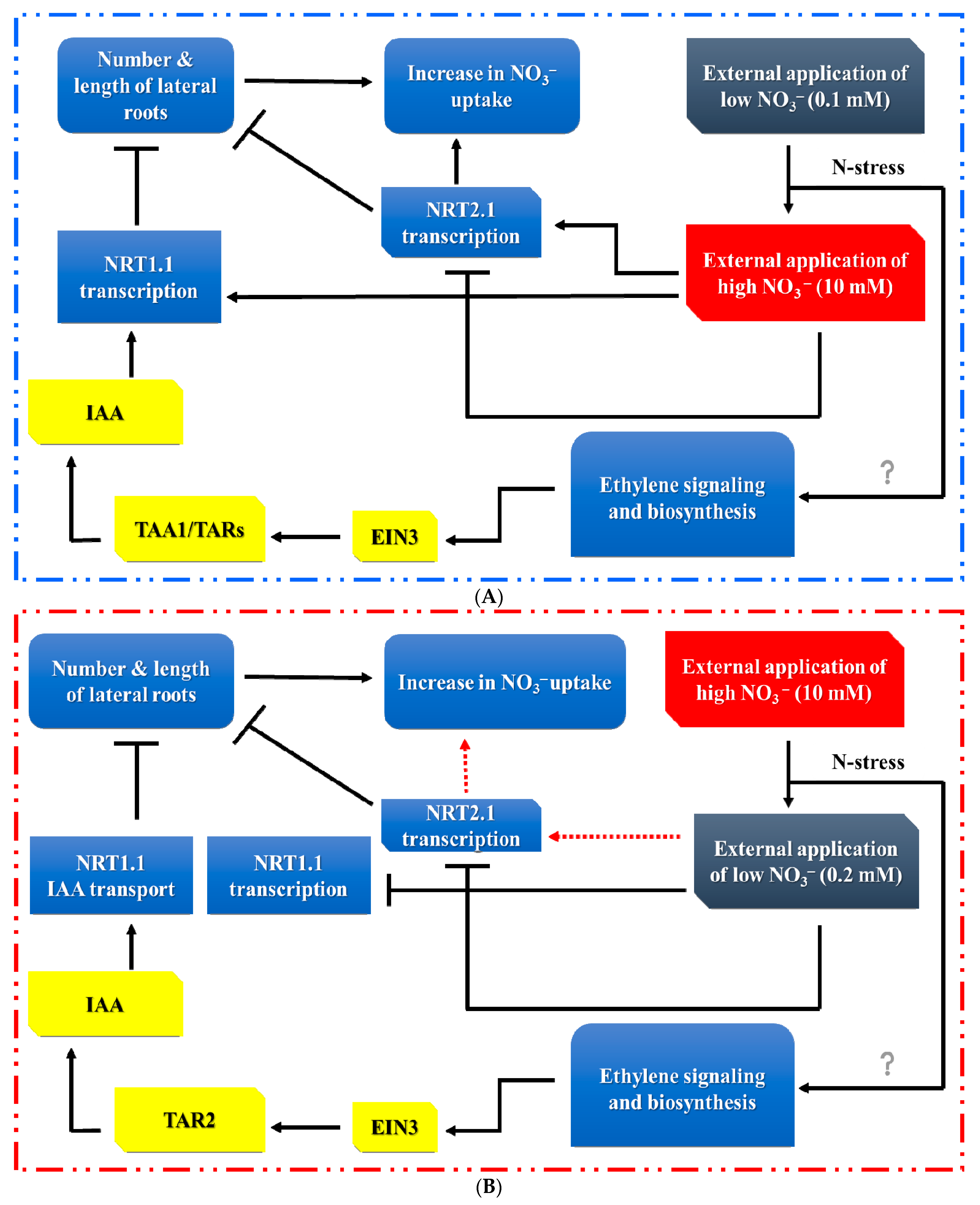
4. Auxin (AUX) Modulates RSA in Response to Nitrogen Supply
5. Ethylene (ETH) Modulates RSA in Response to Nitrogen Stress
6. Brassinosteroid (BR), Strigolactones (SLs), Jasmonic Acid (JA), and Salicylic Acid (SA) Modulate RSA in Response to Nitrogen Stress
7. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ahmad, N.; Su, B.; Ibrahim, S.; Kuang, L.; Tian, Z.; Wang, X.; Wang, H.; Dun, X. Deciphering the Genetic Basis of Root and Biomass Traits in Rapeseed (Brassica napus L.) through the Integration of GWAS and RNA-Seq under Nitrogen Stress. Int. J. Mol. Sci. 2022, 23, 7958. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Ibrahim, S.; Tian, Z.; Kuang, L.; Wang, X.; Wang, H.; Dun, X. Quantitative Trait Loci Mapping Reveals Important Genomic Regions Controlling Root Architecture and Shoot Biomass under Nitrogen, Phosphorus, and Potassium Stress in Rapeseed (Brassica napus L.). Front. Plant Sci. 2022, 13, 4666. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, A.; Qiang, D.; Zhun, W.; Xiangru, W.; Huiping, G.; Hengheng, Z.; Nianchang, P.; Xiling, Z.; Meizhen, S. Growth and Nitrogen Metabolism Are Associated with Nitrogen-Use Efficiency in Cotton Genotypes. Plant Physiol. Biochem. 2020, 149, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Anas, M.; Liao, F.; Verma, K.K.; Sarwar, M.A.; Mahmood, A.; Chen, Z.L.; Li, Q.; Zeng, X.P.; Liu, Y.; Li, Y.R. Fate of Nitrogen in Agriculture and Environment: Agronomic, Eco-Physiological and Molecular Approaches to Improve Nitrogen Use Efficiency. Biol. Res. 2020, 53, 47. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Hu, B.; Chu, C. Nitrogen Use Efficiency in Crops: Lessons from Arabidopsis and Rice. J. Exp. Bot. 2017, 68, 2477–2488. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.K.; Bali, S.K. A Review of Methods to Improve Nitrogen Use Efficiency in Agriculture. Sustainability 2017, 10, 51. [Google Scholar] [CrossRef]
- Javed, T.; Indu, I.; Singhal, R.K.; Shabbir, R.; Shah, A.N.; Kumar, P.; Jinger, D.; Dharmappa, P.M.; Shad, M.A.; Saha, D.; et al. Recent Advances in Agronomic and Physio-Molecular Approaches for Improving Nitrogen Use Efficiency in Crop Plants. Front. Plant Sci. 2022, 13, 877544. [Google Scholar] [CrossRef]
- Kant, S.; Bi, Y.M.; Rothstein, S.J. Understanding Plant Response to Nitrogen Limitation for the Improvement of Crop Nitrogen Use Efficiency. J. Exp. Bot. 2011, 62, 1499–1509. [Google Scholar] [CrossRef]
- Muratore, C.; Espen, L.; Prinsi, B. Nitrogen Uptake in Plants: The Plasma Membrane Root Transport Systems from a Physiological and Proteomic Perspective. Plants 2021, 10, 681. [Google Scholar] [CrossRef]
- Ueda, Y.; Konishi, M.; Yanagisawa, S. Molecular Basis of the Nitrogen Response in Plants. Soil Sci. Plant Nutr. 2017, 63, 329–341. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, Y.; Hang, H. Differential Contributions of to Nitrogen Use in Response to a Variable Inorganic Nitrogen Supply in Plantlets of Two Brassicaceae Species in Vitro. Plant Methods 2019, 15, 86. [Google Scholar] [CrossRef]
- Agusti, J.; Ramireddy, E.; Brumos, J. Editorial: Integration of Hormonal Signals Shaping Root Growth, Development, and Architecture. Front. Plant Sci. 2021, 12, 10–12. [Google Scholar] [CrossRef] [PubMed]
- Vissenberg, K.; Claeijs, N.; Balcerowicz, D.; Schoenaers, S. Hormonal Regulation of Root Hair Growth and Responses to the Environment in Arabidopsis. J. Exp. Bot. 2020, 71, 2412–2427. [Google Scholar] [PubMed]
- Notaguchi, M.; Okamoto, S. Dynamics of Long-Distance Signaling via Plant Vascular Tissues. Front. Plant Sci. 2015, 6, 161. [Google Scholar] [CrossRef] [PubMed]
- Sami, F.; Siddiqui, H.; Hayat, S. Interaction of Glucose and Phytohormone Signaling in Plants. Plant Physiol. Biochem. 2019, 135, 119–126. [Google Scholar] [CrossRef]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Atanassova, R. The Sugar-Signaling Hub: Overview of Regulators and Interaction with the Hormonal and Metabolic Network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef]
- Li, L.; Liu, K.H.; Sheen, J. Dynamic Nutrient Signaling Networks in Plants. Annu. Rev. Cell Dev. Biol. 2021, 37, 341–367. [Google Scholar] [CrossRef]
- Barneix, A.J. Physiology and Biochemistry of Source-Regulated Protein Accumulation in the Wheat Grain. J. Plant Physiol. 2007, 164, 581–590. [Google Scholar] [CrossRef]
- EL Sabagh, A.; Islam, M.S.; Hossain, A.; Iqbal, M.A.; Mubeen, M.; Waleed, M.; Reginato, M.; Battaglia, M.; Ahmed, S.; Rehman, A.; et al. Phytohormones as Growth Regulators During Abiotic Stress Tolerance in Plants. Front. Agron. 2022, 4, 765068. [Google Scholar] [CrossRef]
- Jia, Z.; Giehl, R.F.H.; von Wirén, N. Nutrient Hormone Relations: Driving Root Plasticity in Plants. Mol. Plant 2022, 15, 86–103. [Google Scholar] [CrossRef]
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under Stress: Involvement of Auxin and Cytokinin. Int. J. Mol. Sci. 2017, 18, 1427. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and Their Metabolic Engineering for Abiotic Stress Tolerance in Crop Plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Li, N.; Euring, D.; Cha, J.Y.; Lin, Z.; Lu, M.; Huang, L.J.; Kim, W.Y. Plant Hormone-Mediated Regulation of Heat Tolerance in Response to Global Climate Change. Front. Plant Sci. 2021, 11, 627969. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Hisano, H.; Hojo, Y.; Matsuura, T.; Ikeda, Y.; Mori, I.C.; Senthil-Kumar, M. Global Profiling of Phytohormone Dynamics during Combined Drought and Pathogen Stress in Arabidopsis thaliana Reveals ABA and JA as Major Regulators. Sci. Rep. 2017, 7, 4017. [Google Scholar] [CrossRef] [PubMed]
- Kazan, K. Auxin and the Integration of Environmental Signals into Plant Root Development. Ann. Bot. 2013, 112, 1655–1665. [Google Scholar] [CrossRef]
- Hirayama, T.; Mochida, K. Plant Hormonomics: A Key Tool for Deep Physiological Phenotyping to Improve Crop Productivity. Plant Cell Physiol. 2022, 63, 1826–1839. [Google Scholar] [CrossRef]
- Oracz, K.; Karpiński, S. Phytohormones Signaling Pathways and ROS Involvement in Seed Germination. Front. Plant Sci. 2016, 7, 864. [Google Scholar] [CrossRef]
- Pozo, M.J.; López-Ráez, J.A.; Azcón-Aguilar, C.; García-Garrido, J.M. Phytohormones as Integrators of Environmental Signals in the Regulation of Mycorrhizal Symbioses. New Phytol. 2015, 205, 1431–1436. [Google Scholar] [CrossRef]
- Wang, M.; Le Gourrierec, J.; Jiao, F.; Demotes-Mainard, S.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Crespel, L.; Bertheloot, J.; Chen, J.; et al. Convergence and Divergence of Sugar and Cytokinin Signaling in Plant Development. Int. J. Mol. Sci. 2021, 22, 1282. [Google Scholar] [CrossRef]
- Nacry, P.; Bouguyon, E.; Gojon, A. Nitrogen Acquisition by Roots: Physiological and Developmental Mechanisms Ensuring Plant Adaptation to a Fluctuating Resource. Plant Soil 2013, 370, 1–29. [Google Scholar] [CrossRef]
- Guan, P. Dancing with Hormones: A Current Perspective of Nitrate Signaling and Regulation in Arabidopsis. Front. Plant Sci. 2017, 8, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Nowicka, B. Modifications of Phytohormone Metabolism Aimed at Stimulation of Plant Growth, Improving Their Productivity and Tolerance to Abiotic and Biotic Stress Factors. Plants 2022, 11, 3430. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Hu, X.; Wang, T.; Xu, G. Modulation of Plant Root Traits by Nitrogen and Phosphate: Transporters, Long-Distance Signaling Proteins and Peptides, and Potential Artificial Traps. Breed. Sci. 2021, 71, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Guan, P.; Wang, R.; Nacry, P.; Breton, G.; Kay, S.A.; Pruneda-Paz, J.L.; Davani, A.; Crawford, N.M. Nitrate Foraging by Arabidopsis Roots Is Mediated by the Transcription Factor TCP20 through the Systemic Signaling Pathway. Proc. Natl. Acad. Sci. USA 2014, 111, 15267–15272. [Google Scholar] [CrossRef]
- Ötvös, K.; Marconi, M.; Vega, A.; O’Brien, J.; Johnson, A.; Abualia, R.; Antonielli, L.; Montesinos, J.C.; Zhang, Y.; Tan, S.; et al. Modulation of Plant Root Growth by Nitrogen Source-defined Regulation of Polar Auxin Transport. EMBO J. 2021, 40, e106862. [Google Scholar] [CrossRef]
- Liu, B.; Wu, J.; Yang, S.; Schiefelbein, J.; Gan, Y. Nitrate Regulation of Lateral Root and Root Hair Development in Plants. J. Exp. Bot. 2020, 71, 4405–4414. [Google Scholar] [CrossRef]
- Li, M.; Wang, T.; Zhang, H.; Liu, S.; Li, W.; Abou Elwafa, S.F.; Tian, H. TaNRT2.1-6B Is a Dual-Affinity Nitrate Transporter Contributing to Nitrogen Uptake in Bread Wheat under Both Nitrogen Deficiency and Sufficiency. Crop J. 2022, 10, 993–1005. [Google Scholar] [CrossRef]
- Sun, J.; Zheng, N. Molecular Mechanism Underlying the Plant NRT1.1 Dual-Affinity Nitrate Transporter. Front. Physiol. 2015, 6, 386. [Google Scholar] [CrossRef] [PubMed]
- Guan, M.Y.; Chen, M.X.; Cao, Z.Z. NRT2.1, a Major Contributor to Cadmium Uptake Controlled by High-Affinity Nitrate Transporters. Ecotoxicol. Environ. Saf. 2021, 218, 112269. [Google Scholar] [CrossRef] [PubMed]
- Orsel, M.; Krapp, A.; Daniel-Vedele, F. Analysis of the NRT2 Nitrate Transporter Family in Arabidopsis. Structure and Gene Expression. Plant Physiol. 2002, 129, 886–896. [Google Scholar] [CrossRef]
- Krouk, G.; Tillard, P.; Gojon, A. Regulation of the High-Affinity NO3- Uptake System by NRT1.1-Mediated NO3- Demand Signaling in Arabidopsis. Plant Physiol. 2006, 142, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Ruffel, S.; Krouk, G.; Ristova, D.; Shasha, D.; Birnbaum, K.D.; Coruzzi, G.M. Nitrogen Economics of Root Foraging: Transitive Closure of the Nitrate-Cytokinin Relay and Distinct Systemic Signaling for N Supply vs. Demand. Proc. Natl. Acad. Sci. USA 2011, 108, 18524–18529. [Google Scholar] [CrossRef]
- Ohkubo, Y.; Kuwata, K.; Matsubayashi, Y. A Type 2C Protein Phosphatase Activates High-Affinity Nitrate Uptake by Dephosphorylating NRT2.1. Nat. Plants 2021, 7, 310–316. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, F.; Crawford, N.M.; Wang, Y. Molecular Regulation of Nitrate Responses in Plants. Int. J. Mol. Sci. 2018, 19, 2039. [Google Scholar] [CrossRef]
- Jung, J.K.H.; McCouch, S. Getting to the Roots of It: Genetic and Hormonal Control of Root Architecture. Front. Plant Sci. 2013, 4, 186. [Google Scholar] [CrossRef]
- Sun, C.H.; Yu, J.Q.; Hu, D.G. Nitrate: A Crucial Signal during Lateral Roots Development. Front. Plant Sci. 2017, 8, 485. [Google Scholar] [CrossRef]
- Raspor, M.; Motyka, V.; Kaleri, A.R.; Ninković, S.; Tubić, L.; Cingel, A.; Ćosić, T. Integrating the Roles for Cytokinin and Auxin in de Novo Shoot Organogenesis: From Hormone Uptake to Signaling Outputs. Int. J. Mol. Sci. 2021, 22, 8554. [Google Scholar] [CrossRef]
- Sakakibara, H. Cytokinin Biosynthesis and Transport for Systemic Nitrogen Signaling. Plant J. 2021, 105, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Zhang, Y.; Du, W.; Xia, H.; Fan, S.; Zhang, B. Signaling Responses to N Starvation: Focusing on Wheat and Filling the Putative Gaps With Findings Obtained in Other Plants. A Review. Front. Plant Sci. 2021, 12, 656696. [Google Scholar] [CrossRef] [PubMed]
- Decouard, B.; Bailly, M.; Rigault, M.; Marmagne, A.; Arkoun, M.; Soulay, F.; Caïus, J.; Paysant-Le Roux, C.; Louahlia, S.; Jacquard, C.; et al. Genotypic Variation of Nitrogen Use Efficiency and Amino Acid Metabolism in Barley. Front. Plant Sci. 2022, 12, 807798. [Google Scholar] [CrossRef]
- Sakakibara, H.; Takei, K.; Hirose, N. Interactions between Nitrogen and Cytokinin in the Regulation of Metabolism and Development. Trends Plant Sci. 2006, 11, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Zaman, M.; Kurepin, L.V.; Catto, W.; Pharis, R.P. Enhancing Crop Yield with the Use of N-Based Fertilizers Co-Applied with Plant Hormones or Growth Regulators. J. Sci. Food Agric. 2015, 95, 1777–1785. [Google Scholar] [CrossRef]
- Takei, K.; Ueda, N.; Aoki, K.; Kuromori, T.; Hirayama, T.; Shinozaki, K.; Yamaya, T.; Sakakibara, H. AtIPT3 Is a Key Determinant of Nitrate-Dependent Cytokinin Biosynthesis in Arabidopsis. Plant Cell Physiol. 2004, 45, 1053–1062. [Google Scholar] [CrossRef]
- Vangheluwe, N.; Beeckman, T. Lateral Root Initiation and the Analysis of Gene Function Using Genome Editing with Crispr in Arabidopsis. Genes 2021, 12, 884. [Google Scholar] [CrossRef]
- Torres-Martínez, H.H.; Rodríguez-Alonso, G.; Shishkova, S.; Dubrovsky, J.G. Lateral Root Primordium Morphogenesis in Angiosperms. Front. Plant Sci. 2019, 10, 206. [Google Scholar] [CrossRef] [PubMed]
- Moreira, S.; Bishopp, A.; Carvalho, H.; Campilho, A. AHP6 Inhibits Cytokinin Signaling to Regulate the Orientation of Pericycle Cell Division during Lateral Root Initiation. PLoS ONE 2013, 8, e56370. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A.; Zheng, B. Molecular Responses during Plant Grafting and Its Regulation by Auxins, Cytokinins, and Gibberellins. Biomolecules 2019, 9, 397. [Google Scholar] [CrossRef]
- Li, H.; Testerink, C.; Zhang, Y. How Roots and Shoots Communicate through Stressful Times. Trends Plant Sci. 2021, 26, 940–952. [Google Scholar] [CrossRef]
- Ramireddy, E.; Chang, L.; Schmülling, T. Cytokinin as a Mediator for Regulating Root System Architecture in Response to Environmental Cues. Plant Signal. Behav. 2014, 9, e27771. [Google Scholar] [CrossRef]
- Bellegarde, F.; Gojon, A.; Martin, A. Signals and Players in the Transcriptional Regulation of Root Responses by Local and Systemic N Signaling in Arabidopsis thaliana. J. Exp. Bot. 2017, 68, 2553–2565. [Google Scholar] [CrossRef] [PubMed]
- Vidal, E.A.; Alvarez, J.M.; Araus, V.; Riveras, E.; Brooks, M.D.; Krouk, G.; Ruffel, S.; Lejay, L.; Crawford, N.M.; Coruzzi, G.M.; et al. Nitrate in 2020: Thirty Years from Transport to Signaling Networks. Plant Cell 2020, 32, 2094–2119. [Google Scholar] [CrossRef]
- Asim, M.; Ullah, Z.; Xu, F.; An, L.; Aluko, O.O.; Wang, Q.; Liu, H. Nitrate Signaling, Functions, and Regulation of Root System Architecture: Insights from Arabidopsis thaliana. Genes 2020, 11, 633. [Google Scholar] [CrossRef]
- Hirose, N.; Takei, K.; Kuroha, T.; Kamada-Nobusada, T.; Hayashi, H.; Sakakibara, H. Regulation of Cytokinin Biosynthesis, Compartmentalization and Translocation. J. Exp. Bot. 2008, 59, 75–83. [Google Scholar] [CrossRef]
- Brenner, W.G.; Romanov, G.A.; Köllmer, I.; Bürkle, L.; Schmülling, T. Immediate-Early and Delayed Cytokinin Response Genes of Arabidopsis thaliana Identified by Genome-Wide Expression Profiling Reveal Novel Cytokinin-Sensitive Processes and Suggest Cytokinin Action through Transcriptional Cascades. Plant J. 2005, 44, 314–333. [Google Scholar] [CrossRef]
- Kiba, T.; Naitou, T.; Koizumi, N.; Yamashino, T.; Sakakibara, H.; Mizuno, T. Combinatorial Microarray Analysis Revealing Arabidopsis Genes Implicated in Cytokinin Responses through the His→Asp Phosphorelay Circuitry. Plant Cell Physiol. 2005, 46, 339–355. [Google Scholar] [CrossRef]
- To, J.P.C.; Kieber, J.J. Cytokinin Signaling: Two-Components and More. Trends Plant Sci. 2008, 13, 85–92. [Google Scholar] [CrossRef]
- Kawai, M.; Tabata, R.; Ohashi, M.; Honda, H.; Kamiya, T.; Kojima, M.; Takebayashi, Y.; Oishi, S.; Okamoto, S.; Hachiya, T.; et al. Regulation of Ammonium Acquisition and Use in Oryza Longistaminata Ramets under Nitrogen Source Heterogeneity. Plant Physiol. 2022, 188, 2364–2376. [Google Scholar] [CrossRef]
- Lebedeva, M.A.; Yashenkova, Y.S.; Dodueva, I.E.; Lutova, L.A. Molecular Dialog between Root and Shoot via Regulatory Peptides and Its Role in Systemic Control of Plant Development. Russ. J. Plant Physiol. 2020, 67, 985–1002. [Google Scholar] [CrossRef]
- Fan, X.; Naz, M.; Fan, X.; Xuan, W.; Miller, A.J.; Xu, G. Plant Nitrate Transporters: From Gene Function to Application. J. Exp. Bot. 2017, 68, 2463–2475. [Google Scholar] [CrossRef]
- Hou, M.; Yu, M.; Li, Z.; Ai, Z.; Chen, J. Molecular Regulatory Networks for Improving Nitrogen Use Efficiency in Rice. Int. J. Mol. Sci. 2021, 22, 40. [Google Scholar] [CrossRef]
- Sakakibara, H. CYTOKININS: Activity, Biosynthesis, and Translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Higuchi, M.; Pischke, M.S.; Mähönen, A.P.; Miyawaki, K.; Hashimoto, Y.; Seki, M.; Kobayashi, M.; Shinozaki, K.; Kato, T.; Tabata, S.; et al. In Planta Functions of the Arabidopsis Cytokinin Receptor Family. Proc. Natl. Acad. Sci. USA 2004, 101, 8821–8826. [Google Scholar] [CrossRef]
- Iwama, A.; Yamashino, T.; Tanaka, Y.; Sakakibara, H.; Kakimoto, T.; Sato, S.; Kato, T.; Tabata, S.; Nagatani, A.; Mizuno, T. AHK5 Histidine Kinase Regulates Root Elongation through an ETR1-Dependent Abscisic Acid and Ethylene Signaling Pathway in Arabidopsis thaliana. Plant Cell Physiol. 2007, 48, 375–380. [Google Scholar] [CrossRef]
- Dello Ioio, R.; Linhares, F.S.; Scacchi, E.; Casamitjana-Martinez, E.; Heidstra, R.; Costantino, P.; Sabatini, S. Cytokinins Determine Arabidopsis Root-Meristem Size by Controlling Cell Differentiation. Curr. Biol. 2007, 17, 678–682. [Google Scholar] [CrossRef]
- Ehrary, A.; Rosas, M.; Carpinelli, S.; Davalos, O.; Cowling, C.; Fernandez, F.; Escobar, M. Glutaredoxin AtGRXS8 Represses Transcriptional and Developmental Responses to Nitrate in Arabidopsis thaliana Roots. Plant Direct 2020, 4, e00227. [Google Scholar] [CrossRef]
- Patterson, K.; Walters, L.A.; Cooper, A.M.; Olvera, J.G.; Rosas, M.A.; Rasmusson, A.G.; Escobar, M.A. Nitrate-Regulated Glutaredoxins Control Arabidopsis Primary Root Growth. Plant Physiol. 2016, 170, 989–999. [Google Scholar] [CrossRef]
- Poitout, A.; Crabos, A.; Petřík, I.; Novák, O.; Krouk, G.; Lacombe, B.; Ruffel, S. Responses to Systemic Nitrogen Signaling in Arabidopsis Roots Involve Trans-Zeatin in Shoots. Plant Cell 2018, 30, 1243–1257. [Google Scholar] [CrossRef]
- Yang, Q.; Zhang, J.; Kojima, M.; Takebayashi, Y.; Uragami, T.; Kiba, T.; Sakakibara, H.; Lee, Y. ABCG11 Modulates Cytokinin Responses in Arabidopsis thaliana. Front. Plant Sci. 2022, 13, 976267. [Google Scholar] [CrossRef]
- Tang, X.; Peng, Y.; Li, Z.; Guo, H.; Xia, X.; Li, B.; Yin, W. The Regulation of Nitrate Reductases in Response to Abiotic Stress in Arabidopsis. Int. J. Mol. Sci. 2022, 23, 1202. [Google Scholar] [CrossRef]
- Subudhi, P.K.; Garcia, R.S.; Coronejo, S.; Tapia, R. Comparative Transcriptomics of Rice Genotypes with Contrasting Responses to Nitrogen Stress Reveals Genes Influencing Nitrogen Uptake through the Regulation of Root Architecture. Int. J. Mol. Sci. 2020, 21, 5759. [Google Scholar] [CrossRef]
- Harris, J.M. Abscisic Acid: Hidden Architect of Root System Structure. Plants 2015, 4, 548–572. [Google Scholar] [CrossRef]
- Zheng, Z.L. Carbon and Nitrogen Nutrient Balance Signaling in Plants. Plant Signal. Behav. 2009, 4, 584–591. [Google Scholar] [CrossRef]
- Araus, V.; Swift, J.; Alvarez, J.M.; Henry, A.; Coruzzi, G.M. A Balancing Act: How Plants Integrate Nitrogen and Water Signals. J. Exp. Bot. 2020, 71, 4442–4451. [Google Scholar] [CrossRef]
- Chen, J.; Liu, S.; Zhang, S.; Ge, C.; Shen, Q.; Ma, H.; Zhang, X.; Dong, H.; Zhao, X.; Pang, C. Nitrogen Modulates Cotton Root Morphology by Affecting Abscisic Acid (ABA) and Salicylic Acid (SA) Content. Arch. Agron. Soil Sci. 2021, 67, 1722–1738. [Google Scholar] [CrossRef]
- Harris, J.M.; Ondzighi-Assoume, C.A. Environmental Nitrate Signals through Abscisic Acid in the Root Tip. Plant Signal. Behav. 2017, 12, e1273303. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Upadhyay, N.; Kumar, N.; Yadav, G.; Singh, J.; Mishra, R.K.; Kumar, V.; Verma, R.; Upadhyay, R.G.; Pandey, M.; et al. Abscisic Acid Signaling and Abiotic Stress Tolerance in Plants: A Review on Current Knowledge and Future Prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef]
- Lu, Y.; Sasaki, Y.; Li, X.; Mori, I.C.; Matsuura, T.; Hirayama, T.; Sato, T.; Yamaguchij, J. ABI1 Regulates Carbon/Nitrogen-Nutrient Signal Transduction Independent of ABA Biosynthesis and Canonical ABA Signalling Pathways in Arabidopsis. J. Exp. Bot. 2015, 66, 2763–2771. [Google Scholar] [CrossRef]
- Zhang, H.; Rong, H.; Pilbeam, D. Signalling Mechanisms Underlying the Morphological Responses of the Root System to Nitrogen in Arabidopsis thaliana. J. Exp. Bot. 2007, 58, 2329–2338. [Google Scholar] [CrossRef]
- Su, H.; Wang, T.; Ju, C.; Deng, J.; Zhang, T.; Li, M.; Tian, H.; Wang, C. Abscisic Acid Signaling Negatively Regulates Nitrate Uptake via Phosphorylation of NRT1.1 by SnRK2s in Arabidopsis. J. Integr. Plant Biol. 2021, 63, 597–610. [Google Scholar] [CrossRef]
- Signora, L.; De Smet, I.; Foyer, C.H.; Zhang, H. ABA Plays a Central Role in Mediating the Regulatory Effects of Nitrate on Root Branching in Arabidopsis. Plant J. 2001, 28, 655–662. [Google Scholar] [CrossRef]
- Zhao, Y.; Xing, L.; Wang, X.; Hou, Y.-J.; Gao, J.; Wang, P.; Duan, C.-G.; Zhu, X.; Zhu, J.-K. The ABA Receptor PYL8 Promotes Lateral Root Growth by Enhancing MYB77-Dependent Transcription of Auxin-Responsive Genes. Sci. Signal. 2014, 7, ra53. [Google Scholar] [CrossRef] [PubMed]
- Ondzighi-Assoume, C.A.; Chakraborty, S.; Harris, J.M. Environmental Nitrate Stimulates Abscisic Acid Accumulation in Arabidopsis Root Tips by Releasing It from Inactive Stores. Plant Cell 2016, 28, 729–745. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.A.A.; Vega, A.; Bouguyon, E.; Krouk, G.; Gojon, A.; Coruzzi, G.; Gutiérrez, R.A.A. Nitrate Transport, Sensing, and Responses in Plants. Mol. Plant 2016, 9, 837–856. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Bousquet, A.; Harris, J.M. Abscisic Acid and Lateral Root Organ Defective/Numerous Infections and Polyphenolics Modulate Root Elongation via Reactive Oxygen Species in Medicago truncatula. Plant Physiol. 2014, 166, 644–658. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Mitchell, D.M.; Harris, J.M. Abscisic Acid Rescues the Root Meristem Defects of the Medicago truncatula Latd Mutant. Dev. Biol. 2007, 304, 297–307. [Google Scholar] [CrossRef] [PubMed]
- Yendrek, C.R.; Lee, Y.C.; Morris, V.; Liang, Y.; Pislariu, C.I.; Burkart, G.; Meckfessel, M.H.; Salehin, M.; Kessler, H.; Wessler, H.; et al. A Putative Transporter Is Essential for Integrating Nutrient and Hormone Signaling with Lateral Root Growth and Nodule Development in Medicago truncatula. Plant J. 2010, 62, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Morre-Le Paven, M.C.; Viau, L.; Hamon, A.; Vandecasteele, C.; Pellizzaro, A.; Bourdin, C.; Laffont, C.; Lapied, B.; Lepetit, M.; Frugier, F.; et al. Characterization of a Dual-Affinity Nitrate Transporter MtNRT1.3 in the Model Legume Medicago truncatula. J. Exp. Bot. 2011, 62, 5595–5605. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, R.; Salehin, M.; Sarah Adeyemo, O.; Salazar, C.; Shulaev, V.; Janine Sherrier, D.; Dickstein, R. Functional Assessment of the Medicago truncatula NIP/LATD Protein Demonstrates That It Is a High-Affinity Nitrate Transporter. Plant Physiol. 2012, 160, 906–916. [Google Scholar] [CrossRef] [PubMed]
- Maymon, T.; Eisner, N.; Bar-Zvi, D. The Abcisic Acid Insensitive (ABI) 4 Transcription Factor Is Stabilized by Stress, ABA and Phosphorylation. Plants 2022, 11, 2179. [Google Scholar] [CrossRef] [PubMed]
- Lin, Z.; Li, Y.; Wang, Y.; Liu, X.; Ma, L.; Zhang, Z.; Mu, C.; Zhang, Y.; Peng, L.; Xie, S.; et al. Initiation and Amplification of SnRK2 Activation in Abscisic Acid Signaling. Nat. Commun. 2021, 12, 2456. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Zhao, X.-Q.; Zhu, Y.-G.; Li, B.; Tong, Y.-P.; Li, Z.-S. Regulation of the High-Affinity Nitrate Transport System in Wheat Roots by Exogenous Abscisic Acid and Glutamine. J. Integr. Plant Biol. 2007, 49, 1719–1725. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, P.; Liu, Q.; Li, G.; Di, D.; Xia, G.; Kronzucker, H.J.; Fang, S.; Chu, J.; Shi, W. TAANR1-TABG1 and TawAbi5-TANRT2S/NARS Link ABA Metabolism and Nitrate Acquisition in Wheat Roots. Plant Physiol. 2020, 182, 1440–1453. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Klopotek, Y.; Hajirezaei, M.R.; Zerche, S.; Franken, P.; Druege, U. Role of Auxin Homeostasis and Response in Nitrogen Limitation and Dark Stimulation of Adventitious Root Formation in Petunia Cuttings. Ann. Bot. 2019, 124, 1053–1066. [Google Scholar] [CrossRef]
- Zhao, B.; Liu, Q.; Wang, B.; Yuan, F. Roles of Phytohormones and Their Signaling Pathways in Leaf Development and Stress Responses. J. Agric. Food Chem. 2021, 69, 3566–3584. [Google Scholar] [CrossRef]
- Vanneste, S.; Friml, J. Auxin: A Trigger for Change in Plant Development. Cell 2009, 136, 1005–1016. [Google Scholar] [CrossRef]
- De Jong, M.; Ongaro, V.; Ljung, K. Auxin and Strigolactone Signaling Are Required for Modulation of Arabidopsis Shoot Branching by Nitrogen Supply. Plant Physiol. 2014, 166, 384–395. [Google Scholar] [CrossRef]
- Gho, Y.S.; Song, M.Y.; Bae, D.Y.; Choi, H.; Jung, K.H. Rice Pin Auxin Efflux Carriers Modulate the Nitrogen Response in a Changing Nitrogen Growth Environment. Int. J. Mol. Sci. 2021, 22, 3243. [Google Scholar] [CrossRef]
- Xu, P.; Ma, W.; Hu, J.; Cai, W. The Nitrate-Inducible NAC Transcription Factor NAC056 Controls Nitrate Assimilation and Promotes Lateral Root Growth in Arabidopsis thaliana. PLoS Genet. 2022, 18, e1010090. [Google Scholar] [CrossRef]
- Lay-Pruitt, K.S.; Takahashi, H. Integrating N Signals and Root Growth: The Role of Nitrate Transceptor NRT1.1 in Auxin-Mediated Lateral Root Development. J. Exp. Bot. 2020, 71, 4365–4368. [Google Scholar] [CrossRef]
- Hu, Q.Q.; Shu, J.Q.; Li, W.M.; Wang, G.Z. Role of Auxin and Nitrate Signaling in the Development of Root System Architecture. Front. Plant Sci. 2021, 12, 690363. [Google Scholar] [CrossRef]
- Jing, H.; Strader, L.C. Interplay of Auxin and Cytokinin in Lateral Root Development. Int. J. Mol. Sci. 2019, 20, 486. [Google Scholar] [CrossRef]
- Casimiro, I.; Marchant, A.; Bhalerao, R.P.; Beeckman, T.; Dhooge, S.; Swarup, R.; Graham, N.; Inzé, D.; Sandberg, G.; Casero, P.J.; et al. Auxin Transport Promotes Arabidopsis Lateral Root Initiation. Plant Cell 2001, 13, 843–852. [Google Scholar] [CrossRef]
- Tian, Q.; Chen, F.; Liu, J.; Zhang, F.; Mi, G. Inhibition of Maize Root Growth by High Nitrate Supply Is Correlated with Reduced IAA Levels in Roots. J. Plant Physiol. 2008, 165, 942–951. [Google Scholar] [CrossRef] [PubMed]
- Kudoyarova, G.R.; Dodd, I.C.; Veselov, D.S.; Rothwell, S.A.; Yu. Veselov, S. Common and Specific Responses to Availability of Mineral Nutrients and Water. J. Exp. Bot. 2015, 66, 2133–2144. [Google Scholar] [CrossRef] [PubMed]
- Ravazzolo, L.; Trevisan, S.; Forestan, C.; Varotto, S.; Sut, S.; Dall’acqua, S.; Malagoli, M.; Quaggiotti, S. Nitrate and Ammonium Affect the Overall Maize Response to Nitrogen Availability by Triggering Specific and Common Transcriptional Signatures in Roots. Int. J. Mol. Sci. 2020, 21, 686. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Pu, X.; Jia, H.; Zhou, Y.; Ye, G.; Yang, Y.; Na, T.; Wang, J. Transcriptome Analysis Reveals Multiple Effects of Nitrogen Accumulation and Metabolism in the Roots, Shoots, and Leaves of Potato (Solanum tuberosum L.). BMC Plant Biol. 2022, 22, 282. [Google Scholar] [CrossRef]
- Canales, J.; Moyano, T.C.; Villarroel, E.; Gutiérrez, R.A. Systems Analysis of Transcriptome Data Provides New Hypotheses about Arabidopsis Root Response to Nitrate Treatments. Front. Plant Sci. 2014, 5, 22. [Google Scholar] [CrossRef]
- Mounier, E.; Pervent, M.; Ljung, K.; Gojon, A.; Nacry, P. Auxin-Mediated Nitrate Signalling by NRT1.1 Participates in the Adaptive Response of Arabidopsis Root Architecture to the Spatial Heterogeneity of Nitrate Availability. Plant Cell Environ. 2014, 37, 162–174. [Google Scholar] [CrossRef]
- Abualia, R.; Ötvös, K.; Novák, O.; Bouguyon, E.; Domanegg, K.; Krapp, A.; Nacry, P.; Gojon, A.; Lacombe, B.; Benková, E. Molecular Framework Integrating Nitrate Sensing in Root and Auxin-Guided Shoot Adaptive Responses. Proc. Natl. Acad. Sci. USA 2022, 119, e2122460119. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Z.S.; Xia, J.Q.; Alfatih, A.; Song, Y.; Huang, Y.J.; Wan, G.Y.; Sun, L.Q.; Tang, H.; Liu, Y.; et al. Rice NIN-LIKE PROTEIN 4 Plays a Pivotal Role in Nitrogen Use Efficiency. Plant Biotechnol. J. 2021, 19, 448–461. [Google Scholar] [CrossRef]
- Hayashi, K.I. The Interaction and Integration of Auxin Signaling Components. Plant Cell Physiol. 2012, 53, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Schrader, J.; Baba, K.; May, S.T.; Palme, K.; Bennett, M.; Bhalerao, R.P.; Sandberg, G. Polar Auxin Transport in the Wood-Forming Tissues of Hybrid Aspen Is under Simultaneous Control of Developmental and Environmental Signals. Proc. Natl. Acad. Sci. USA 2003, 100, 10096–10101. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and Signaling Aspects Underpinning the Regulation of Plant Carbon Nitrogen Interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Xin, W.; Zhang, L.; Gao, J.; Zhang, W.; Yi, J.; Zhen, X.; Bi, C.; He, D.; Liu, S.; Zhao, X. Adaptation Mechanism of Roots to Low and High Nitrogen Revealed by Proteomic Analysis. Rice 2021, 14, 5. [Google Scholar] [CrossRef] [PubMed]
- Caba, J.M.; Centeno, M.L.; Fernández, B.; Gresshoff, P.M.; Ligero, F. Inoculation and Nitrate Alter Phytohormone Levels in Soybean Roots: Differences between a Supernodulating Mutant and the Wild Type. Planta 2000, 211, 98–104. [Google Scholar] [CrossRef]
- Ambreetha, S.; Chinnadurai, C.; Marimuthu, P.; Balachandar, D. Plant-Associated Bacillus Modulates the Expression of Auxin-Responsive Genes of Rice and Modifies the Root Architecture. Rhizosphere 2018, 5, 57–66. [Google Scholar] [CrossRef]
- Song, X.; Xiong, Y.; Kong, X.; Huang, G. Roles of Auxin Response Factors in Rice Development and Stress Responses. Plant. Cell Environ. 2022. [Google Scholar] [CrossRef]
- Wu, L.; Wang, J.L.; Li, X.F.; Guo, G.Q. Cytokinin-controlled Gradient Distribution of Auxin in Arabidopsis Root Tip. Int. J. Mol. Sci. 2021, 22, 3874. [Google Scholar] [CrossRef]
- Kirolinko, C.; Hobecker, K.; Wen, J.; Mysore, K.S.; Niebel, A.; Blanco, F.A.; Zanetti, M.E. Auxin Response Factor 2 (ARF2), ARF3, and ARF4 Mediate Both Lateral Root and Nitrogen Fixing Nodule Development in Medicago truncatula. Front. Plant Sci. 2021, 12, 659061. [Google Scholar] [CrossRef]
- Luo, P.; Di, D.W.; Wu, L.; Yang, J.; Lu, Y.; Shi, W. MicroRNAs Are Involved in Regulating Plant Development and Stress Response through Fine-Tuning of TIR1/AFB-Dependent Auxin Signaling. Int. J. Mol. Sci. 2022, 23, 510. [Google Scholar] [CrossRef]
- Fang, X.Z.; Fang, S.Q.; Ye, Z.Q.; Liu, D.; Zhao, K.L.; Jin, C.W. NRT1.1 Dual-Affinity Nitrate Transport/Signalling and Its Roles in Plant Abiotic Stress Resistance. Front. Plant Sci. 2021, 12, 715694. [Google Scholar] [CrossRef]
- Zhang, Y.; Ma, Y.; Zhao, D.; Tang, Z.; Zhang, T.; Zhang, K. Genetic Regulation of Lateral Root Development. Plant Signal. Behav. 2022. [Google Scholar] [CrossRef]
- Swarup, R.; Bhosale, R. Developmental Roles of AUX1/LAX Auxin Influx Carriers in Plants. Front. Plant Sci. 2019, 10, 1306. [Google Scholar] [CrossRef]
- Jiang, J.; Zhu, H.; Li, N.; Batley, J.; Wang, Y. The MiR393-Target Module Regulates Plant Development and Responses to Biotic and Abiotic Stresses. Int. J. Mol. Sci. 2022, 23, 9477. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Xing, X.; Wang, Y.; Tran, A.; Crawford, N.M. A Genetic Screen for Nitrate Regulatory Mutants Captures the Nitrate Transporter Gene NRT1.1. Plant Physiol. 2009, 151, 472–478. [Google Scholar] [CrossRef] [PubMed]
- Smolko, A.; Bauer, N.; Pavlovi, I.; Pěnčík, A. Altered Root Growth, Auxin Metabolism and Distribution in Arabidopsis thaliana Exposed to Salt and Osmotic Stress. Int. J. Mol. Sci. 2021, 22, 7993. [Google Scholar] [CrossRef]
- Ding, Z.J.; Yan, J.Y.; Li, C.X.; Li, G.X.; Wu, Y.R.; Zheng, S.J. Transcription Factor WRKY46 Modulates the Development of Arabidopsis Lateral Roots in Osmotic/Salt Stress Conditions via Regulation of ABA Signaling and Auxin Homeostasis. Plant J. 2015, 84, 56–69. [Google Scholar] [CrossRef]
- Undurraga, S.F.; Ibarra-henríquez, C.; Fredes, I.; Álvarez, J.M.; Gutiérrez, R.A. Nitrate Signaling and Early Responses in Arabidopsis Roots. J. Exp. Bot. 2017, 68, 2541–2551. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.-H.; Miao, Z.-Q.; Qi, G.-F.; Wu, J.; Cai, X.-T.; Mao, J.-L.; Xiang, C.-B. MADS-Box Transcription Factor AGL21 Regulates Lateral Root Development and Responds to Multiple External and Physiological Signals. Mol. Plant 2014, 7, 1653–1669. [Google Scholar] [CrossRef]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. Int. J. Mol. Sci. 2019, 6343. [Google Scholar] [CrossRef]
- Forestan, C.; Farinati, S.; Varotto, S. The Maize PIN Gene Family of Auxin Transporters. Front. Plant Sci. 2012, 3, 16. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Ball, G.; Hodgman, C.; Lu, C. Analysis of Gene Regulatory Networks of Maize in Response to Nitrogen. Genes 2018, 9, 151. [Google Scholar] [CrossRef]
- Qin, H.; He, L.; Huang, R. The Coordination of Ethylene and Other Hormones in Primary Root Development. Front. Plant Sci. 2019, 10, 874. [Google Scholar] [CrossRef]
- Qin, H.; Huang, R. Auxin Controlled by Ethylene Steers Root Development. Int. J. Mol. Sci. 2018, 19, 3656. [Google Scholar] [CrossRef] [PubMed]
- Tian, Q.Y.; Sun, P.; Zhang, W.H. Ethylene Is Involved in Nitrate-Dependent Root Growth and Branching in Arabidopsis thaliana. New Phytol. 2009, 184, 918–931. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, P.; Liu, P.; Song, Y.; Zhang, D. Genetic Effects and Expression Patterns of the Nitrate Transporter (NRT) Gene Family in Populus tomentosa. Front. Plant Sci. 2021, 12, 661635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.B.; Meng, S.; Gong, J.M. The Expected and Unexpected Roles of Nitrate Transporters in Plant Abiotic Stress Resistance and Their Regulation. Int. J. Mol. Sci. 2018, 19, 3535. [Google Scholar] [CrossRef]
- Zheng, D.; Han, X.; An, Y.; Guo, H.; Xia, X.; Yin, W. The Nitrate Transporter NRT2.1 Functions in the Ethylene Response to Nitrate Deficiency in Arabidopsis. Plant Cell Environ. 2013, 36, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Brumos, J.; Li, H.; Ji, Y.; Ke, M.; Gong, X.; Zeng, Q.; Li, W.; Zhang, X.; An, F.; et al. A Small-Molecule Screen Identifies L-Kynurenine as a Competitive Inhibitor of TAA1/TAR Activity in Ethylene-Directed Auxin Biosynthesis and Root Growth in Arabidopsis. Plant Cell 2011, 23, 3944–3960. [Google Scholar] [CrossRef]
- Leblanc, A.; Renault, H.; Lecourt, J.; Etienne, P.; Deleu, C.; Deunff, E. Le Elongation Changes of Exploratory and Root Hair Systems Induced by Aminocyclopropane Carboxylic Acid and Aminoethoxyvinylglycine Affect Nitrate Uptake and BnNrt2.1 and BnNrt1.1 Transporter Gene Expression in Oilseed Rape. Plant Physiol. 2008, 146, 1928–1940. [Google Scholar] [CrossRef]
- Fatma, M.; Asgher, M.; Iqbal, N.; Rasheed, F.; Sehar, Z.; Sofo, A.; Khan, N.A. Ethylene Signaling under Stressful Environments: Analyzing Collaborative Knowledge. Plants 2022, 11, 2211. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Nazar, R.; Syeed, S.; Masood, A.; Khan, N.A. Exogenously-Sourced Ethylene Increases Stomatal Conductance, Photosynthesis, and Growth under Optimal and Deficient Nitrogen Fertilization in Mustard. J. Exp. Bot. 2011, 62, 4955–4963. [Google Scholar] [CrossRef] [PubMed]
- Jahan, B.; Iqbal, N.; Fatma, M.; Sehar, Z.; Masood, A.; Sofo, A. Ethylene Supplementation Combined with Split Application of Nitrogen and Sulfur Protects Salt-Inhibited Photosynthesis through Optimization of Proline Metabolism and Antioxidant. Plants 2021, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, F.; Ji, Z.; Belfield, E.J.; Ding, Z.J.; Zheng, S.J.; Harberd, N.P. Ethylene Signaling Modulates Arabidopsis thaliana Nitrate Metabolism. Planta 2022, 255, 94. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Ma, M.; Feng, Y.; Li, H.; Wang, Y.; Ma, Y.; Li, M.; An, F. EIN2-Directed Translational Regulation of Ethylene Signaling in Arabidopsis. Cell 2015, 163, 670–683. [Google Scholar] [CrossRef]
- Gu, S.; Wang, L.; Cheuh, C.; Lo, W.; Lindsey, K.; Wang, L. CHITINASE LIKE1 Regulates Root Development of Dark-Grown Seedlings by Modulating Ethylene Biosynthesis in Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 600. [Google Scholar] [CrossRef]
- Li, G.; Li, B.; Dong, G.; Feng, X.; Kronzucker, H.J.; Shi, W. Ammonium-Induced Shoot Ethylene Production Is Associated with the Inhibition of Lateral Root Formation in Arabidopsis. J. Exp. Bot. 2013, 64, 1413–1425. [Google Scholar] [CrossRef]
- Jia, Z.; Giehl, R.F.H.; Meyer, R.C.; Altmann, T.; von Wirén, N. Natural Variation of BSK3 Tunes Brassinosteroid Signaling to Regulate Root Foraging under Low Nitrogen. Nat. Commun. 2019, 10, 2378. [Google Scholar] [CrossRef]
- Chai, S.; Chen, J.; Yue, X.; Li, C.; Zhang, Q.; de Dios, V.R.; Yao, Y.; Tan, W. Interaction of BES1 and LBD37 Transcription Factors Modulates Brassinosteroid-Regulated Root Forging Response under Low Nitrogen in Arabidopsis. Front. Plant Sci. 2022, 13, 998961. [Google Scholar] [CrossRef]
- Wei, Z.; Li, J. Regulation of Brassinosteroid Homeostasis in Higher Plants. Front. Plant Sci. 2020, 11, 583622. [Google Scholar] [CrossRef]
- Jia, Z.; Giehl, R.F.H.; Von Wirén, N. The Root Foraging Response under Low Nitrogen Depends on Dwarf1-Mediated Brassinosteroid Biosynthesis. Plant Physiol. 2020, 183, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Rozhon, W.; Akter, S.; Fernandez, A.; Poppenberger, B. Inhibitors of Brassinosteroid Biosynthesis and Signal Transduction. Molecules 2019, 24, 4372. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.; Walk, T.C.; Han, P.; Chen, L.; Zhang, S.; Li, Y.; Hu, X.; Xie, L.; Yang, Y.; Liu, J.; et al. Adaption of Roots to Nitrogen Deficiency Revealed by 3D Quantification and Proteomic Analysis. Plant Physiol. 2019, 179, 329–347. [Google Scholar] [CrossRef] [PubMed]
- Kaniganti, S.; Bhattacharya, J.; Petla, B.P.; Reddy, P.S. Strigolactone, a Neglected Plant Hormone, with a Great Potential for Crop Improvement: Crosstalk with Other Plant Hormones. Environ. Exp. Bot. 2022, 204, 105072. [Google Scholar] [CrossRef]
- Santoro, V.; Schiavon, M.; Gresta, F.; Ertani, A.; Cardinale, F.; Sturrock, C.J.; Celi, L.; Schubert, A. Strigolactones Control Root System Architecture and Tip Anatomy in Solanum lycopersicum L. Plants under P Starvation. Plants 2020, 9, 612. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Tao, J.; Liu, S.; Huang, S.; Chen, S.; Xie, X.; Yoneyama, K.; Zhang, Y.; Xu, G. Strigolactones Are Involved in Phosphate- and Nitrate-Deficiency-Induced Root Development and Auxin Transport in Rice. J. Exp. Bot. 2014, 65, 6735–6746. [Google Scholar] [CrossRef]
- Alvi, A.F.; Sehar, Z.; Fatma, M.; Masood, A.; Khan, N.A. Strigolactone: An Emerging Growth Regulator for Developing Resilience in Plants. Plants 2022, 11, 2604. [Google Scholar] [CrossRef]
- Ito, S.; Ito, K.; Abeta, N.; Takahashi, R.; Sasaki, Y.; Yajima, S. Effects of Strigolactone Signaling on Arabidopsis Growth under Nitrogen Deficient Stress Condition. Plant Signal. Behav. 2016, 11, e1126031. [Google Scholar] [CrossRef]
- Wang, Y.; Mostafa, S.; Zeng, W.; Jin, B. Function and Mechanism of Jasmonic Acid in Plant Responses to Abiotic and Biotic Stresses. Int. J. Mol. Sci. 2021, 22, 8568. [Google Scholar] [CrossRef]
- Ruan, J.; Zhou, Y.; Zhou, M.; Yan, J.; Khurshid, M.; Weng, W.; Cheng, J.; Zhang, K. Jasmonic Acid Signaling Pathway in Plants. Int. J. Mol. Sci. 2019, 20, 2479. [Google Scholar] [CrossRef]
- Kamali, S.; Singh, A. Jasmonates as Emerging Regulators of Plants Response to Variable Nutrient Environment. CRC. Crit. Rev. Plant Sci. 2022, 41, 271–285. [Google Scholar] [CrossRef]
- Conesa, C.M.; Saez, A.; Navarro-Neila, S.; de Lorenzo, L.; Hunt, A.G.; Sepúlveda, E.B.; Baigorri, R.; Garcia-Mina, J.M.; Zamarreño, A.M.; Sacristán, S.; et al. Alternative Polyadenylation and Salicylic Acid Modulate Root Responses to Low Nitrogen Availability. Plants 2020, 9, 251. [Google Scholar] [CrossRef] [PubMed]
- Silva-Navas, J.; Conesa, C.M.; Saez, A.; Navarro-Neila, S.; Garcia-Mina, J.M.; Zamarreño, A.M.; Baigorri, R.; Swarup, R.; del Pozo, J.C. Role of Cis-Zeatin in Root Responses to Phosphate Starvation. New Phytol. 2019, 224, 242–257. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, Y.; Kim, J.W.; Lee, H.; Lee, W.S.; Kim, S.; Wang, Z.; Kim, S. Identification of Arabidopsis BAK1-Associating Receptor-Like Kinase 1 (BARK1) and Characterization of Its Gene Expression and Brassinosteroid-Regulated Root Phenotypes. Plant Cell Physiol. 2013, 1, 1620–1634. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Giehl, R.F.H.; von Wirén, N. Local Auxin Biosynthesis Acts Downstream of Brassinosteroids to Trigger Root Foraging for Nitrogen. Nat. Commun. 2021, 12, 5437. [Google Scholar] [CrossRef]
- Kim, J.Y.; Song, J.T.; Seo, H.S. Ammonium-Mediated Reduction in Salicylic Acid Content and Recovery of Plant Growth in Arabidopsis Siz1 Mutants Is Modulated by NDR1 and NPR1. Plant Signal. Behav. 2021, 16, 1928819. [Google Scholar] [CrossRef]
- Xing, J.; Wang, Y.; Yao, Q.; Zhang, Y.; Zhang, M.; Li, Z. Brassinosteroids Modulate Nitrogen Physiological Response and Promote Nitrogen Uptake in Maize (Zea mays L.). Crop J. 2022, 10, 166–176. [Google Scholar] [CrossRef]
- Manoli, A.; Trevisan, S.; Quaggiotti, S.; Varotto, S. Identification and Characterization of the BZR Transcription Factor Family and Its Expression in Response to Abiotic Stresses in Zea mays L. Plant Growth Regul. 2018, 84, 423–436. [Google Scholar] [CrossRef]
- Yang, S.; Yuan, D.; Zhang, Y.; Sun, Q.; Xuan, Y.H. BZR1 Regulates Brassinosteroid-Mediated Activation of AMT1;2 in Rice. Front. Plant Sci. 2021, 12, 665883. [Google Scholar] [CrossRef]
- Jiao, X.; Wang, H.; Yan, J.; Kong, X.; Liu, Y.; Chu, J.; Chen, X.; Fang, R.; Yan, Y. Promotion of BR Biosynthesis by MiR444 Is Required for Ammonium-Triggered Inhibition of Root Growth. Plant Physiol. 2020, 182, 1454–1466. [Google Scholar] [CrossRef]
- Sun, H.; Li, W.; Burritt, D.J.; Tian, H.; Zhang, H.; Liang, X.; Miao, Y.; Mostofa, M.G.; Tran, L.S.P. Strigolactones Interact with Other Phytohormones to Modulate Plant Root Growth and Development. Crop J. 2022, 10, 1517–1527. [Google Scholar] [CrossRef]
- Ogura, T.; Goeschl, C.; Filiault, D.; Mirea, M.; Slovak, R.; Wolhrab, B.; Satbhai, S.B.; Busch, W. Root System Depth in Arabidopsis Is Shaped by EXOCYST70A3 via the Dynamic Modulation of Auxin Transport. Cell 2019, 178, 400–412.e16. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Singh, D.; Saksena, H.B.; Sharma, M.; Tiwari, A.; Awasthi, P.; Botta, H.K.; Shukla, B.N.; Laxmi, A. Understanding the Intricate Web of Phytohormone Signalling in Modulating Root System Architecture. Int. J. Mol. Sci. 2021, 22, 5508. [Google Scholar] [CrossRef]
- Li, S.; Tian, Y.; Wu, K.; Ye, Y.; Yu, J.; Zhang, J.; Liu, Q.; Hu, M.; Li, H.; Tong, Y.; et al. Modulating Plant Growth–Metabolism Coordination for Sustainable Agriculture. Nature 2018, 560, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Reddy, A.R. Simplifying the Root Dynamics: From Complex Hormone–Environment Interactions to Specific Root Architectural Modulation. Plant Growth Regul. 2018, 85, 337–349. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Arabidopsis | |||
|---|---|---|---|
| Gene Name | Gene ID | Functions | References |
| AtGRXS1 | At1g03020 | NO3− upregulated GRXs in shoots to modulate primary root (PR) elongation through CK signaling | [75,76] |
| AtGRXS3 | At4g15700 | ||
| AtGRXS4 | At4g15680 | ||
| AtGRXS5 | At4g15690 | ||
| AtGRXS6 | At3g62930 | ||
| AtGRXS8 | At4g15660 | ||
| AtGRXS11 | At3g62950 | ||
| AtIPT3 | At4g17870 | Involved in CK biosynthesis, modulating lateral root (LR) elongation in response to partial NO3− deficiency | [77] |
| AtIPT5 | At3g50500 | ||
| AtIPT7 | At5g66880 | ||
| AtABCG14 | At1g31770 | Involved in CK transport in response to N | [78] |
| CYP735A2 | At1g67110 | Involved in CK biosynthesis and modulating RSA in response to N-stress | [79] |
| Rice | |||
| OsIPT4 | Os03g0810100 | Essential for CK biosynthesis dependent on the glumine-related signal | [80] |
| OsIPT5 | Os07g0211700 | ||
| OsIPT7 | Os05g0551700 | ||
| OsIPT8 | Os01g0688300 | ||
| Arabidopsis | |||
|---|---|---|---|
| Gene Name | Gene ID | Functions | References |
| AtPYL2 | At2g26040 | Involved in ABA signaling and PR and LR growth in response to NO3− | [87] |
| AtPYL4 | At2g38310 | ||
| AtABI1 | At4g26080 | ||
| AtPYL1 | At5g46790 | ||
| AtABI2 | At5g57050 | ||
| AtHAB1 | At1g72770 | ||
| AtPP2Ca | At3g11410 | ||
| AtPYR1 | At4g17870 | ||
| AtSnRK2.2 | At3g50500 | Involved in ABA signaling and PR growth in response to NO3− | [89,100] |
| AtSnRK2.3 | At5g66880 | ||
| AtSnRK2.6 | At4g33950 | ||
| Wheat | |||
| TaGS2-2Ab | TraesCS2A02g500400 | Nitrogen use efficiency and ABA signaling | [49] |
| TaNAR2.1 | TraesCS6D02G193100 | Nitrogen uptake and assimilation, ABA biosynthesis | [101,102] |
| TaNAR2.2 | TraesCS5D02G506100 | ||
| TaNRT2.1 | TraesCS6A02G030900 | ||
| TaNRT2.2 | TraesCS6D02G035800 | ||
| TaWabi5 | TraesCS5B02G235600 | Involved in ABA signaling in response to low N | [103] |
| TaBG1 | TraesCS6A02G048200 | Involved in ABA biosynthesis in response to low N | |
| Arabidopsis | |||
|---|---|---|---|
| Gene Name | Gene ID | Functions | References |
| AtGOXL3 | At1g75620 | N-regulated putative auxin efflux carries | [136] |
| AtPILS2 | At1g71090 | ||
| AtPILS6 | At5g01990 | ||
| AtPLT1 | At3g20840 | Regulating cell elongation in the process of severe NO3− deficiency and inhibiting PR elongation | [110] |
| AtPLT2 | At5g51190 | ||
| AtWOX5 | At3g11260 | ||
| AtWRKY46 | At2g46400 | Regulated by high NH4+ and activates auxin conjugating genes GH3.1, 3.6 to modulate PR elongation | [137] |
| miR167a | At3g22886 | Regulated by glutamine and glutamate to modulate LR development | [62] |
| AtARF8 | At5g37020 | ||
| miR393 | At2g39885 | NO3−-responsive miR393/AFB3 regulatory module controlling RSA | [110] |
| AtAFB3 | At1g12820 | ||
| AtOBP4 | At5g60850 | Modulating LR initiation by auxin signaling in response to NO3− supply | [138] |
| AtNAC4 | At5g07680 | ||
| AtSNX1 | At5g57090 | N-regulated auxin-transport-related gene | [35] |
| AtAXR4 | At1g54990 | Mediating LR elongation in response to local NO3− supply | [110] |
| AtPIN1 | At1g73590 | Modulating PR elongation through mediating auxin flow in response to NO3− supply | [132] |
| AtPIN2 | At5g06140 | ||
| AtPIN4 | At2g01420 | ||
| AtPIN7 | At5g01990 | ||
| AtAUX1 | At2g38120 | ||
| AtLAX2 | At2g21050 | ||
| AtLAX3 | At1g77690 | ||
| AtAGL21 | At4g37940 | Involved in auxin homeostasis and modulating LR elongation under NO3− deficiency | [139] |
| AtGH3.1 | At2g14960 | Regulating auxin levels and NH4+ sensitivity in roots | [88] |
| AtGH3.2 | At4g37390 | ||
| AtGH3.3 | At2g23170 | ||
| AtGH3.4 | At1g59500 | ||
| AtGH3.6 | At5g54510 | ||
| AtYUC3 | At1g04610 | Promoting auxin levels in LR tips and modulating LR elongation in response to N deficiency | [140] |
| AtYUC5 | At5g43890 | ||
| AtYUC7 | At2g33230 | ||
| AtYUC8 | At4g28720 | ||
| AtTAR1 | At1g23320 | Involved in auxin biosynthesis and LR growth in response to low N | [49] |
| AtTAR2 | At4g24670 | ||
| AtTAA1 | At1g70560 | ||
| Maize | |||
| ZmPIN1 | Zm00001d044812 | Upregulated by local, high NO3− in roots | [141] |
| ZmPIN9 | Zm00001d043179 | ||
| Rice | |||
| OsDNR1 | Os01g0178000 | Regulating auxin homeostasis, N uptake, and assimilation | [142] |
| OsARF1 | Os01g0236300 | Auxin response factors, positiveregulates N uptake | [129] |
| OsARF5 | Os02g0141100 | ||
| OsARF6 | Os02g0164900 | ||
| OsARF17 | Os06g0677800 | ||
| OsARF19 | Os06g0702600 | ||
| OsARF24 | Os12g0479400 | ||
| OsARF25 | Os12g0613700 | ||
| OsPIN1 | Os11g0137000 | Auxin efflux carriers, regulated by local NO3− supply and modulate LR growth and seminal root (SR) elongation | [107] |
| OsPIN2 | Os06g0660200 | ||
| OsPIN5 | Os01g0919800 | ||
| OsPIN9 | Os01g0802700 | ||
| OsPIN10 | Os05g0576900 | ||
| OsAUX1 | Os01g0856500 | Auxin influx carriers, regulated by local NO3− supply and modulate LRP establishment | [127] |
| OsAXR4 | Os11g0544100 | ||
| Wheat | |||
| TaTAR2.1-3A | TraesCS3A02G093000 | Involved in auxin biosynthesis in response to low N | [49] |
| TaTAR2.5 | TraesCS1B02G133900 | ||
| Arabidopsis | |||
|---|---|---|---|
| Gene Name | Gene ID | Functions | References |
| AtCTR1 | At5g03730 | Involved in ETH signaling in response to low NO3− | [154] |
| AtEIL1 | At2g27050 | ||
| AtEIN3 | At3g20770 | ||
| AtEIN2 | At5g03280 | Involved in NO3−-dependent root growth and branching | [155] |
| AtETR1 | At1g66340 | ||
| AtACS2 | At1g01480 | Involved in ETH biosynthesis in response to high NO3− | [156,157] |
| AtACS4 | At2g22810 | ||
| AtACS5 | At5g65800 | ||
| AtACS6 | At4g11280 | ||
| AtACS7 | At4g26200 | ||
| AtACS8 | At4g37770 | ||
| AtACS11 | At4g08040 | ||
| AtACO1 | At2g19590 | ||
| AtACO2 | At1g62380 | ||
| Arabidopsis | ||||
|---|---|---|---|---|
| Hormones | Gene Name | Gene ID | Functions | References |
| BR | AtBAK1 | At4g33430 | Involved in BR signaling and low NO3− promoting PR elongation | [158,174] |
| AtBSK3 | At4g00710 | |||
| AtBR6ox2 | At3g30180 | N deficiency regulating BR biosynthesis genes | [175] | |
| AtDWF4 | At3g50660 | |||
| AtCPD | At5g05690 | |||
| AtDWF1 | At3g19820 | Involved in BR biosynthesis and root foraging response | [161] | |
| SA | AtSIZ1 | At5g60410 | Regulating SA levels and activating NIA1 and NIA2 | [176] |
| Maize | ||||
| BR | ZmBRI1 | Zm00001d011721 | Involved in BR signaling and PR elongation in response to low NO3− stress | [177] |
| ZmDET2 | Zm00001d007910 | N-regulated BR-biosynthesis-related genes | [178] | |
| ZmBZR1 | Zm00001d046305 | |||
| ZmCPD | Zm00001d052475 | |||
| ZmDWF4 | Zm00001d028325 | |||
| Rice | ||||
| BR | OsRAVL1 | Os04g0581400 | Involved in BR signaling and involved in BR signaling and regulating OsAMT1;2 and NH4+ uptake | [179] |
| OsBZR1 | Os07g0580500 | |||
| OsMADS23 | Os08g0431900 | BR biosynthesis gene OsBRD1 involved in NH4+-inhibited SR elongation | [180] | |
| OsMADS25 | Os04g0304400 | |||
| OsMADS27 | Os02g0579600 | |||
| OsMADS57 | Os02g0731200 | |||
| OsMADS61 | Os04g0461300 | |||
| miR444 | Os02g0731300 | |||
| OsBRD1 | Os03g0602300 | |||
| SL | OsSPL14 | Os08g0509600 | SL signaling target proteins and modulating SR elongation | [80] |
| OsSPL17 | Os09g0491532 | |||
| OsD10 | Os01g0746400 | Involved in SL biosynthesis in response to NO3− | [181] | |
| OsD17 | Os04g0550600 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, N.; Jiang, Z.; Zhang, L.; Hussain, I.; Yang, X. Insights on Phytohormonal Crosstalk in Plant Response to Nitrogen Stress: A Focus on Plant Root Growth and Development. Int. J. Mol. Sci. 2023, 24, 3631. https://doi.org/10.3390/ijms24043631
Ahmad N, Jiang Z, Zhang L, Hussain I, Yang X. Insights on Phytohormonal Crosstalk in Plant Response to Nitrogen Stress: A Focus on Plant Root Growth and Development. International Journal of Molecular Sciences. 2023; 24(4):3631. https://doi.org/10.3390/ijms24043631
Chicago/Turabian StyleAhmad, Nazir, Zhengjie Jiang, Lijun Zhang, Iqbal Hussain, and Xiping Yang. 2023. "Insights on Phytohormonal Crosstalk in Plant Response to Nitrogen Stress: A Focus on Plant Root Growth and Development" International Journal of Molecular Sciences 24, no. 4: 3631. https://doi.org/10.3390/ijms24043631
APA StyleAhmad, N., Jiang, Z., Zhang, L., Hussain, I., & Yang, X. (2023). Insights on Phytohormonal Crosstalk in Plant Response to Nitrogen Stress: A Focus on Plant Root Growth and Development. International Journal of Molecular Sciences, 24(4), 3631. https://doi.org/10.3390/ijms24043631