Abstract
Tissue engineering and cell therapy for regenerative medicine have great potential to treat chronic disorders. In musculoskeletal disorders, mesenchymal stromal cells (MSCs) have been identified as a relevant cell type in cell and regenerative strategies due to their multi-lineage potential, although this is likely to be a result of their trophic and immunomodulatory effects on other cells. This PRISMA systematic review aims to assess whether the age of the patient influences the chondrogenic potential of MSCs in regenerative therapy. We identified a total of 3027 studies after performing a search of four databases, including Cochrane, Web of Science, Medline, and PubMed. After applying inclusion and exclusion criteria, a total of 14 papers were identified that were reviewed, assessed, and reported. Cell surface characterization and proliferation, as well as the osteogenic, adipogenic, and chondrogenic differentiation, were investigated as part of the analysis of these studies. Most included studies suggest a clear link between aged donor MSCs and diminished clonogenic and proliferative potential. Our study reveals a heterogeneous and conflicting range of outcomes concerning the chondrogenic, osteogenic, and adipogenic potential of MSCs in relation to age. Further investigations on the in vitro effects of chronological age on the chondrogenic potential of MSCs should follow the outcomes of this systematic review, shedding more light on this complex relationship.
1. Introduction
Mesenchymal stromal cells (MSCs) have been the topic of much interest with regard to their potential and much of current research has intertwined varying definitions for what these specifically represent. The current established definition for these cells suggests that MSCs are whole-tissue-specific cells that can switch morphology into other cell types. Some of this has been clarified by seminal work from Caplan et al. [1], which has shown that the majority of MSCs are not stem cells. MSCs are now believed to secrete immunomodulatory and trophic cells that alter the biology of the region in which it is active [1].
MSCs were initially derived from the bone marrow [2], but the complexities and inconvenience associated with obtaining MSCs from this source [3] led to the development of protocols for the isolation of MSCs from pericytes and adventitial progenitor cells from other tissues in the body [4], including adipose tissue, peripheral blood, lungs, dental tissues (dental pulp stem cells (DPSCs), stem cells from human exfoliated deciduous teeth (SHED)) [5,6,7] and neonatal-birth-associated tissues (e.g., placenta, umbilical cord and cord blood) [8].
MSCs can differentiate into other mesoderm-derived specialised cells such as osteoblasts, chondrocytes, adipocytes, tenocytes, and myocytes [9,10,11], and are also capable of differentiating into non-mesoderm-derived cells such as glial cells and neural cells [12,13,14,15,16,17,18]. Morphologically, MSCs can be characterised by a spindle-like shape in the undifferentiated state [19]. These cells can also be characterised by the cell surface markers [20] they express or lack: MSCs typically express CD73, CD90, CD105 [21], cell adhesion molecules (e.g., CD54/ICAM–1 or CD106/VCAM–1) [22,23] and some cytokine receptors (e.g., IL–1R and TNF–aR). MSCs are also typically negative for the cell surface markers CD11b, CD14, CD19, CD34, CD45, CD79a HLA–DR and vWF [24,25]. A precise MSC characterization profile is difficult to establish due to the heterogeneity of MSCs. The International Society for Cellular Therapy (ISCT) proposed the minimum criteria that characterize cells as MSCs [26].
MSCs have an important potential therapeutic use in human musculoskeletal disease. A key driver for research in MSCs is biological therapies for osteoarthritis (OA). OA is the most common articular cartilage degenerative disorder affecting different joints of the body, albeit mainly affecting the joints in the knee, hip, hands, and spine [27]. Characterised by articular pain, OA leads to impaired joint function and progressive disability [28]. Due to the lack of disease-modifying treatment for OA, current strategies are limited to analgesic control and lifestyle modification [29]. Past studies have shown that MSCs can differentiate into the main components of articular cartilage: (1) proteoglycans, providing compressive stiffness to the cartilage; and (2) type II collagen, contributing to tensile strength and resilience of the cartilage [30]. As with many pathologies, advancing age is associated with OA development; age in itself is not a pathological process but represents the accumulation of changes that contribute to disease. With over 75% of OA sufferers being those over 65 [31], the quality and integrity of MSCs in treating older patients have been questioned and studied.
Several properties of MSCs make them putative treatment options in diseases characterised by degeneration. These include (1) selective migration and homing to the inflammatory microenvironment [11,32]; (2) secretion of trophic factors, such as growth factors, cytokines, morphogens, anti-apoptotic factors, and exosomes [33,34,35]; and (3) immunoregulatory responses to antibody production, T cell activation and cytokine secretion by NK cells [9,36]. These properties make MSCs behave in a non-stem-cell-like fashion. Due to the poor regenerative potential of articular cartilage [37], the aforementioned properties of MSCs alongside their chondrogenic potential could make them useful in orthopaedic applications through their immunomodulatory effects on other cells as well as immune cells.
Mesenchymal progenitors give rise to chondrocytes in vitro that have the potential to lead to cartilage development. Chondrocytes are metabolically active cells that differentiate and proliferate during development and are the predominant cells in healthy cartilage [38]. However, there is no detection of cell proliferation in adult cartilage. Only 1–5% of cartilage is occupied by the chondrocytes in adults; the rest is the extracellular matrix [39]. A large volume of proteoglycans, collagen, glycoproteins and hyaluronan, also known as extracellular matrix compounds, are turned over and synthesized by chondrocytes [40]. The mechanical and chemical environment of chondrocytes are factors that affect their metabolic activity [41]. Although skeletal development can result from the proliferation of chondrocytes in vivo, it can also occur in vitro [42]. The formation of chondrocytes, resulting in cartilage development, is called chondrogenic differentiation and takes 28 days to form in vitro [43].
The osteogenic and adipogenic differentiation process of MSC in vitro lasts almost three weeks. Lineage commitment and maturation are the two main stages of these processes [44]. The first differentiation from bone marrow stem cells, described by Friedenstein et al. in the late 1960s, was the formation of osteoblasts. This differentiation was identified as osteogenic. The formation of fat cells—adipocytes derived from stem cells—is called adipogenesis, where preadipocytes differentiate into mature adipocytes. There are two cell populations adipocytes can arise from: either from bone marrow progenitor cells (migration to adipose tissue) or from preadipocytes (adipose-tissue-resident) [45].
MSCs, when cultured on fibronectin and bFGF-coated wells, undergo neurogenesis, differentiating into neuronal cells. After 14 days, they display characteristics of both neurons and glial cells, confirmed by positive markers NF200 (68.9%), GFAP (15.4%), and Gal-C (12.3%). This successful differentiation process holds potential for tissue regeneration in nervous system disorders and has been a key focus in clinical trials [5,46,47].
Advancing age is associated with declining MSC function, including reduced proliferation and differentiation potential, enhanced cellular apoptosis, and reduced wound-healing properties [48,49,50]. Epigenetic and genetic mechanisms have been shown to underlie these senescence-related changes [51]. Despite these findings, studies on the effect of ageing on the chondrogenic potential of MSCs have yielded inconsistent results. It is worth noting that the papers included in these studies are in their early stages, making it unlikely for them to undergo senescence. Additionally, the culture conditions remained consistent within each paper, indicating that any observed changes in age-related characteristics are not attributed to senescence [52,53].
This systematic review, therefore, aims to summarise the evidence and evaluate the methodological quality of studies that have examined the effect of ageing on the MSC chondrogenic potential. In addition, this systematic review assesses whether the chronological age of patients affected the proliferative and chondrogenic capacity for differentiation with mesenchymal stromal cells. Finally, this systematic review reports the available literature and studies that are present and amalgamates these together to formulate a review of these here.
2. Methods
2.1. Search Strategy
A systematic review of the literature was performed according to the Preferred Reporting Items for Systematic Reviews (PRISMA) guidelines [54]. A search of the literature was carried out exploring four databases: Cochrane, PubMed, Medline, and Web of Science. These were performed in the last week of July and the first week of August. For our search strategy, we used the following search terms: “age” or “aging” and “mesenchymal stem cells” or “mesenchymal stem cell” or “mesenchymal stromal cells” or “mesenchymal stromal cell” and “cell surface characterisation” or “cell surface” or “differentiation potential” or “differentiation” and “in vitro”.
In total, 23 studies were extracted from Cochrane (1946—last week of July 2023), 992 from PubMed (1996—last week of July 2023), 703 from Medline (1946—first week of August 2023), and 1309 from Web of Science (1900—last week of July 2023).
Overall, 966 papers were identified as duplicates and removed. Once the inclusion and exclusion criteria were applied and papers were screened based on abstract and title, 1927 papers were excluded. A total number of 144 studies were assessed based on the full text and excluded based on the exclusion criteria listed below. Following this process, which is explained in detail in Figure 1, we identified a final number of 14 papers, which we used for data extraction.
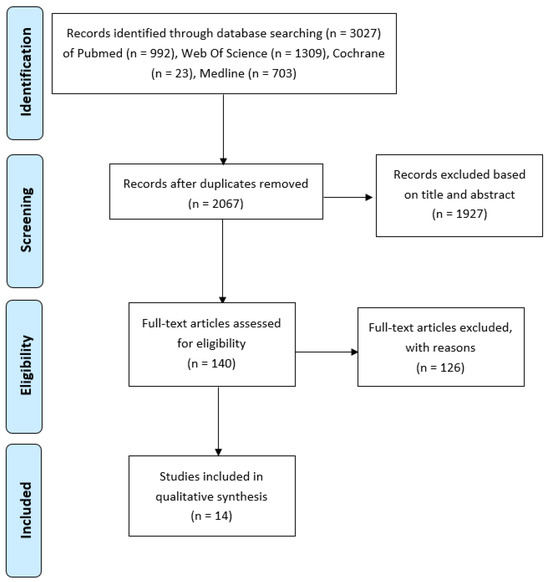
Figure 1.
PRISMA Flow Diagram (Figure 1 in [54]).
The search was conducted by A.V., D.A., K.K., and N.C., and two authors (W.K. and A.V.) independently screened titled abstracts. In cases of disagreement, papers were included for full review.
The study was registered on the PROSPERO database with the registration number (459279).
2.2. Inclusion Criteria
- In vitro studies involving adult human subjects;
- Studies with a reference to subjects’ age;
- Studies looking at MSCs and the source of extraction of cells specified;
- Studies that refer to chondrogenic differentiation;
- English language.
2.3. Exclusion Criteria
- Duplicate studies;
- Those not in the English language;
- Non-human studies;
- Studies using samples from patients with systemic diseases;
- Any paper other than research papers was excluded;
- Studies looking at non-mesenchymal cells, e.g., embryonic, umbilical cord, and periodontal MSCs.
2.4. Data Extraction
An Excel spreadsheet was used to present the data that were extracted from each study. The data extracted are presented in four tables listing the reference of the papers, a brief description of the study, subjects’ number and chronological age, source of the MSCs, culture conditions, proliferation analysis, MSC cell surface characterisation, and chondrogenic, adipogenic and osteogenic differentiation.
2.5. Quality Assessment
A quality check for each paper was carried out using a modified version of the “OHAT risk of bias rating tool for Human and Animal studies” from the Office of Health Assessment and Translation (OHAT tool) [55]. Any differences in the results were solved by discussion.
3. Results
3.1. General Characteristics of the Papers
In this systematic review, 14 studies were included, as presented in Figure 1. The earliest study that is included that met the inclusion and exclusion criteria is from 2007 [56] and the latest 2019. For most papers, MSCs were isolated from different tissues of healthy or osteoarthritic patients, and the proliferation and chondrogenic differentiation of MSCs were investigated. Moreover, some papers presented data for adipogenic and osteogenic differentiation. The samples used for each paper varied between 6 and 260. Seven papers compared the effects of age between three different age groups, three papers presented data for two age groups, three for one group (younger and older) and only one had four different age groups. Where possible, we excluded whole groups or data from people under 18 years old. Bone MSCs were isolated for seven papers. More specifically, bone marrow (BM) MSCs were isolated mainly from the iliac crest and tibia. Furthermore, MSCs of adipose tissue were isolated for five papers and ligaments MSCs (ACL) for two papers. Finally, all the studies used similar culture conditions, which allowed us to compare the data they extracted with each other. They isolated MSCs by processing different tissues, washing them, digesting them in collagenase, filtering them, washing the pellet, and then resuspending them in a fresh medium and plating the MSCs in flasks under tissue culture conditions. The non-adherent cells were removed after 48 h. The medium was changed on different days for each study passage when confluent enough (Table 1).
3.2. Proliferation of MSCs
Most studies assessed (11 out of 14) found an association between increased age of donor source of MSCs and diminished initial proliferative rate, as well as the capacity to maintain proliferation and clonogenic potential. Three studies did not identify proliferative or clonogenic differences between aged and young donor MSCs, [57,58,59] possibly due to the protocols for isolation and/or maintenance of MSCs having rate-limiting effects (Table 2).

Table 2.
Proliferation and MSC characterization.
Table 1.
General characteristics of the studies.
Table 1.
General characteristics of the studies.
| References | Brief Description of Study | Source of MSCs | Number of Subjects | Age | Culture Conditions |
|---|---|---|---|---|---|
| Scharstuhl et al., 2007 [56] | BM-MSCs were isolated from the femoral shaft at total hip replacement. | BM
| 98 | 24–92 years | In total, 25 mL of BM was collected, and the mononuclear cell fraction isolated through density gradient centrifugation. After 48 h, non-adherent cells were removed. |
| Stolzing et al., 2008 [60] | Investigated MSC from donors of various ages and determined their “fitness” by measuring various age and senescence markers in relation to their differentiation capacity and functionality. | BM
| 57 | Group I: 19–40 years old—“adult” group, Group II: >40 years old—“aged” group used in our analyses. Group III: 7–18 years old—“young” group not included in our review. | Lympho-prep was used for BM MNC separation and cryopreservation in liquid nitrogen. In the CFU-f test, 5 × 106 BM MNCs were initially plated. Subsequent passages of MSCs were cultured at 1 × 106 cells in T75 culture flasks. |
| Alm, J. J. et al., 2010 [61] | The study examined MSCs in fracture patients looking at cell surface markers, proliferation through several passages as well as osteogenic, chondrogenic and adipogenic differentiation. | BM
| 41 | Group I: (76–95) Group II: (75–85) (19–60) | MNCs isolated, plated at 2 × 106 (BM) or 5 × 106 (PB) cells in 25-cm2 flasks. Non-adherent cells discarded after 48 h. Cells trypsinized after 14–21 days and re-plated at 1000 cells/cm2 in flasks. |
| Fickert et al., 2011 [62] | The study investigated the influence of donor age on proliferation and osteogenic differentiation in long-term ex vivo cultures of primary human MSCs from patients in different age groups. | BM
| 15 | Group I: <50 yearsGroup II: 50–65 yearsGroup III: >65 years | Density gradient used and MNCs isolated. |
| Alt, E. U. et al., 2012 [63] | Adipose-tissue-derived MSCs (ASCs) were isolated from young, middle age, and aged healthy volunteers to investigate the effect of ageing on the self-renewal and differentiation potential of ASCs | ASCs
| 40 | 15–71 Group I: <20 yearsGroup II: 30–40 yearsGroup III: >50 years | In total, 50 g tissue digested with collagenase I, RBC lysis buffer used. ASCs From three groups plated at densities of 1000 to 25 cells/cm2 in 12-well dishes. Analysis at day 10. |
| Siegel et al., 2013 [64] | BM-MSCs were assessed for phenotype, in vitro growth, colony formation, telomerase activity, differentiation capacity, T cell proliferation suppression, cytokine and trophic factor secretion, and receptor expression. Expression of Oct4, Nanog, Prdm14, and SOX2 mRNA was compared to pluripotent stem cells. | BM | 53 | 13–80 years | Isolated mononuclear cells seeded at 1 × 105 cells/cm2 in standard culture medium with 10% pooled human AB serum. |
| Ding, D.-C. et al., 2013 [57] | ASC isolated from abdominal subcutaneous fat of women undergoing gynaecological surgery. | Adipose Tissue Abdominal subcutaneous fat | 27 | Group I: (30–39 y) Group II: (40–49 y) Group III: (50–60 y) | ASCs dissociated with collagenase, passaged at 80% confluence, 1:3 ratio. |
| Choudhery, M. S. et al., 2014 [65] | Assessed effects of age on ASC expansion and differentiation. Measured expression of p16 and p21, population doublings (PD), superoxide dismutase (SOD) activity, cellular senescence, and differentiation potential. | Adipose Tissue | 29 | Group I: (<30) Group II: (35–55 y) Group III: (>60) | ASCs isolated via enzymatic digestion and then plated. |
| Ruzzini, L et al., 2014 [66] | Tendon stem cells (TSCs) were isolated through magnetic sorting from the hamstring tendons of six patients. TSC percentage, morphology and clonogenic potential were evaluated, as well as the expression of specific surface markers. | Hamstring tendons | 6 | Group I: (20–22) Group II: (28–31) Group III: (49–50) | Tendon biopsies yielded MSCs from fat and muscle via digestion and centrifugation, then cultured. |
| Lee, D.-H. et al., 2015 [67] | This study assessed the phenotypic and functional differences in ACL-MSCs isolated from younger and older donors and evaluated the correlation between ACL-MSC proportion and donor age. | ACL remnants from ACL reconstruction or TKA | 69 | Group I: Young—ACL reconstruction—29.67 ± 10.92 years Group II: Old—TKA—67.96 ± 5.22 years | Isolated ACL fascicles, washed, minced, digested, and plated after centrifugation and filtration. |
| Marędziak, M. et al., 2016 [68] | The study evaluated fibroblast colony forming unit (CFUF) count, proliferation rate, population doubling time (PDT), and lineage-specific differentiation parameters (osteogenic, adipogenic, chondrogenic). | Subcutaneous adipose tissue. | 32 | 22 to 77 years old Group I: >20 years (mean age 24 ± 1.4 years; n = 8), Group II: >50 years (mean age 57.5 ± 0.7 years; n = 8), Group III: >60 years (mean age 67 ± 1.4 years; n = 8), and Group IV: >70 years (mean age 75 ± 2.8 years; n = 8). | Tissue samples digested in collagenase, centrifuged, and cells resuspended in culture medium for culturing. |
| Kawagishi-Hotta, M et al., 2017 [69] | ASCs were assessed for proliferation, as well as adipogenic, osteogenic, and chondrogenic differentiation potentials in vitro. Individual donor characteristics were analyzed via principal component analysis (PCA) based on these parameters. | Adipose tissue | 260 | 5–97 years old | Subcutaneous adipose tissue digested to obtain SVF cells and ASCs, cultured in FGF-supplemented medium. |
| Prall, W. C. et al., 2018 [59] | This study found similar properties in hMSCs from iliac crest and proximal tibia, including proliferation and differentiation capabilities. | BM
| 46 | Group I: young (18–49 years) Group II: Aged (≥50 years) | MSCs were isolated by washing the bone graft material and digesting it. |
| Andrzejewska, A. et al., 2019 [58] | Compared adult and elder BM-MSCs from a biobank, evaluating growth kinetics, gene expression, and differentiation potential. | Metaphyseal Bone Marrow
| 23 | Mean age of adults = 38 years, Mean age of elderly = 72 years | Isolatedvia centrifugation and density grdients, cultured under standard conditions in an expansion medium. |
3.3. Characterization of MSCs
All papers characterized the cells they processed. In the majority of the papers, characterization was performed using flow cytometry. Thirteen studies stained for CD90, a well-accepted MSC cell surface marker. One study did not stain for CD90 but stained for CD44, CD146, and STRO1 [66]. This study investigated the absence of CD34, as it proved a non-hematopoietic correlation (Table 2). Eleven papers used CD105, and eight papers used CD73. Based on the ISCT criteria, nine studies stained for negative markers CD34 or CD45. Some papers mentioned additional positive markers and some other negative markers. Some studies found no correlation between MSC phenotype and donor age, [56,57,58,59,62,63,65], whereas some studies indicated a correlation between MSC cell surface markers and donor age. In one study, young patients observed high levels of increased expression of MCAM, VCAM-1, ALCAM, PDGFRβ, PDL-1, Thy1, and CD71 [64]. A study that investigated CD44, CD90, CD105, and Stro-1, presented significant age-related changes such as increased expression levels of CD44 and a decrease in other cell surface markers [60]. Another study presented BM-MSCs and PB-MSCs from three younger fracture patients being less than 1% positive for CD45, CD14, CD19, and HLA-DR, and 98% positive for CD73, CD105, and CD90 [61]. A higher expression of CD73 was observed in the youngest group of patients in comparison to all the other groups, which indicated a variation connected with donor age and CD73. However, no differences were found with regard to the percentage of MSC surface antigen expression [68]. On the other hand, another study presented a higher expression of CD73 on ASCs from elderly donors compared to young donors. A significantly lower expression of CD105 was observed in ASCs of the high proliferation of young group donors compared to those in the low-potential group [69] (Table 2).
3.4. Other Outcomes
The ability of BM-MSCs to self-replicate was positively related to the expression of the Prdm14 mRNA (n = 18). The presence of Oct4 (n = 24) or Nanog mRNA (n = 24) was related to the lack of self-replicability of BM-MSCs [64]. In these cells, markers suggestive of senescence of the cells were noted with increased levels of ROS levels and increasing oxidative damage. These resulted in p21 and p53 levels increasing resulting in reduced MSC numbers and subsequently loss of differentiative potential. Increasing age resulted in progressively increased levels of NO, whilst superoxide dismutase (SOD) activity was reduced with increasing age. Biomarkers of ageing in MSCs included evidence of carbonylation of protein, and levels of lipofuscin and AGEs, which were markedly increased with increasing age and utilised to assess levels of oxidative stress of MSCs. Cellular homeostasis controlled by heat shock proteins (HSPs) was disrupted with increasing age as a result of reduced HSP levels. When passaged at the beginning, MSCs, irrespective of age, exhibited morphology that was similar in nature with spindles prominently present during replication. These are lost with further passaging as cells and the properties of the cells change with increased cell size [60]. Typically, samples from the cells in older tissue had less viable mononuclear cells [65] (Table 2).
3.5. Chondrogenic Differentiation of MSCs
In contrast to the findings in the majority of studies that suggest a clear link between aged donor MSCs and diminished clonogenic and proliferative potential, no such consensus exists with regard to the capacity for chondrogenic differentiation (Table 3). This is consistent with current thinking that while MSC donor age plays a role in chondrogenic differentiative capacity, other factors are also important. This explains why many of the studies that have been explored do not show homogeneity in their results (Table 3). Of the 14 studies identified, only 4 demonstrated reduced chondrogenicity with age, 5 showed no difference or no statistical difference, and 5 did not report any results after investigating the chondrogenesis.
Table 3.
Chondrogenic differentiation.
3.6. Adipogenic and Osteogenic Differentiation of MSCs
Similar to the observation for chondrogenic differentiation, analyses of the impact of donor age on adipogenic (12 studies) and osteogenic (13 studies) differentiation of MSCs, did not yield a consensus (Table 4). Again, while it is likely that donor age and isolation/maintenance procedures play a role in differentiative capacity, other factors may be important. Finally, Scharstuhl et al. [56] did not investigate adipogenic or osteogenic differentiation. One study did not investigate the adipogenic differentiation [62]. From the 14 studies within our literature search, 4 papers demonstrated adipogenic differentiation reduction with age, 8 showed no difference or no statistical difference and 2 failed to report adipogenicity. With regards to osteogenic potential, six studies demonstrated a significant reduction in osteogenesis with age, seven studies did not demonstrate any meaningful difference in potential, and one failed to report osteogenecity.
Table 4.
Osteogenic and adipogenic differentiation.
3.7. Quality of Studies
A modified version of the OHAT tool was used to grade each study using some of the 11 questions listed in the Table below. Overall, eight studies were differently low risk. However, all 14 studies had probably a high risk of the “Blinding of research personnel” (Table 5). There was some concern towards the “Accounting for important confounding/modifying variables“ for four studies [67], and in five studies, there was concern towards other potential threats regarding “internal validity (bias)“. Overall, none of the studies were definitely high risk; all 14 studies included in this review were of low risk and high quality.
Table 5.
Risk of bias analysis/assessment.
4. Discussion
4.1. Risk Factors of Osteoarthritis
Although the aetiology of osteoarthritis is not fully understood yet, which is a limitation for fully treating OA, there are risk factors that lead to osteoarthritis. The risk factors of OA could be categorized into modifiable, known as the secondary form of osteoarthritis, and non-modifiable, which represents the primary form of OA [70]. Unfortunately, the mechanism of the primary form of OA is not well understood. Heredity and age are both related to the primary form of osteoarthritis. Genetic predisposition plays an important role in the development of OA in its primary form [71]. On the other hand, secondary OA is mainly due to insults to the native joint such as articular injury, and predisposes the individual to developing secondary osteoarthritis through a culmination of variation in joint loading (biomechanical changes) as well as the biochemical milieu during acute injury which can be deleterious to cartilage [72].
Age is a well-accepted risk factor for OA. With the increase in the age of a patient, there is a higher risk of developing OA. Many studies consider weight and in particular, obesity as one of the highest risk factors for osteoarthritis as a result of the pressure and forces on the synovium and cartilage. Sex appears to play an important role in the risks of osteoarthritis. Females are at higher risk of OA, especially those who have undergone menopause. Although genetic risk factors are important, these have not been extensively explored [73]. Physical activity or intense exercise increases OA and can be compounded by intra-articular injuries (be they meniscal or ligamentous in nature). Although heavy physical workload did not demonstrate any significant proof for causing osteoarthritis, occupations that require kneeling and lifting appear to be a risk factor for OA [73]. Some occupations, like construction work and farming, have a high risk of causing future abnormalities to the knees of a patient [71]. Lifestyle could be considered a risk factor as well. For example, smoking varies as a risk factor with equivocal results with some studies presenting it as a protective factor and some as a risk factor for future OA [71].
4.2. Chronological Age and MSCs Behavior In Vitro and In Vivo
In vivo, ageing varies in comparison to in vitro ageing in its definition. In vivo, ageing is represented by the chronological age of a donor, whereas in vitro, this is typically represented by the loss of stem cell characteristics as cell lineage differentiates into a particular phenotype during expansion. The earlier the passage of the cells, the closer the environment is to the human body it is, and therefore, cells behave under similar conditions. Colony formation ability, osteogenic potential, and proliferative capacity are reduced in older MSC donors in comparison to younger ones. Most studies suggest a direct correlation between prefiltration and MSC donor age. The proliferation potential decreases with the increase in age. The rate of proliferation in vivo is low and there is no de-differentiation of chondrocyte reported. In vitro, they are cultured monolayer, and they are lost in the twenty-day phenotype [74]. Some investigations lead to a correlation between senescence and donor age of MSCs; one of the reasons might be the elevated expression of the BAX gene and p21 and P53 (its pathway genes) [75]. Cartilage, like all other tissues, presents changes with the increase in chronological age. Decreases in mitosis and metabolic activity are observed due to the extinction of the blood vessels. Aged cartilage is thinner because of declining proteoglycan levels and increased collagen crosslinking [76].
4.3. Effects—MSCs Characterization
The studies we reviewed here characterized the cells as MSCs using some of the ISCT positive and negative cell surface markers. However, a few studies did not include all the minimum criteria that ISCT proposed. An MSC population that included some, but not all the minimal criteria, was described and presented by several studies included in this review. Finding an optimum, unified definition of what characterizes MSCs through identifying specific surface markers and functional assays will provide homogeneity between studies and would be favourable for future studies. The heterogeneity seen in these studies means that there are varying opinions of what defines an MSC and can add extra variables to these studies.
4.4. Effects—Chondrogenic Potential
Chondrogenic potential is compromised with age, although this finding is not uniform between studies. Only 4 of 14 studies state that with an increase in age, there is a decrease in the chondrogenic potential of MSCs [60,63,65,68]. What causes this loss of potential is speculated to involve the difficulty in stimulating genes, allowing for chondrogenic growth factors and an environment conducive to these cells [77]. The interplay with other factors may represent why there is a lack of equipoise with regard to the influence of age in chondrogenesis. Then, different factors may influence the chondrocyte differentiation in addition to just age. Identifying these factors remains an area of current research. The role of immune cells within these regions is believed to influence the ability of an MSC to show chondrogenic potential and may represent the damaged tissue within cartilage. Identifying these particular biomarkers will become important in helping us better understand which biochemical pathways influence MSCs’ aid with cartilage repair and will provide a useful tool in patient stratification in the future. This may also explain the heterogeneity in the results found. Further analysis in understanding the differentiating genes between different aged donors can help reviewers understand the differential gene profile through transcriptomics and metabolomics of MSCs.
4.5. Adipogenic and Osteogenic Potential
Four studies we reviewed reported a correlation between adipogenic potential and age; six reported a correlation between osteogenic potential and age. The potential of osteogenic and adipogenic differentiation is decreasing with the increase in the age of MSC donors. This may point to similar mechanisms influencing both of these and may represent a biochemical pathway that works in tandem with each other, depending on the state of the neighbouring cells. Some of the studies identified within our study appear to conclude that this regenerative potential is diminished with age. This is likely a result of cumulative mutations within these cells that drive the cell into an apoptotic pathway and resultant cell death [78]. It is important to note that not all studies observed this relationship. The majority of the reviewed papers did not find any significant effect of age on the differentiation capabilities of MSCs into adipocytes and osteoblasts. This divergence in findings could be attributed to variations in study design, sample size, cell culture conditions, or other factors that may influence the observed outcomes. Reversing these to prevent this from activating and potentially upregulating these mechanisms will be further identified with the use of novel genomics, metabolomics, and transcriptomic technology and provide a potential avenue for researchers to explore in the future.
5. Conclusions
This systematic review demonstrates a heterogeneous conflicting range of outcomes concerning the chondrogenic, osteogenic, and adipogenic potential of MSCs in relation to age. While several studies present a correlation when comparing chronological age with the osteogenic and adipogenic potential of MSCs within the literature, others present varied findings on this matter. In addition, most studies that we reviewed suggest a clear link between aged donor MSCs and diminished clonogenic and proliferative potential. This does not appear to be reflected in chondrogenesis in these cells, as only four reported a decline in the chondrogenic potential with increased age. Chronological age-related changes in MSC function have important implications for the use of these cells in clinical applications for an ageing population. Further investigations on the in vitro effects of chronological age on the chondrogenic potential of MSC should follow the outcomes of this systematic review, shedding more light on this complex relationship. The results from this study should be used to plan further investigations looking at the effects of chronological age on cellular senescence and identify pathways that could be targeted to potentially reverse any age-related changes. Understanding the pathomechanisms involved in the process can highlight potential reversal targets in promoting and maintaining cartilage repair.
Osteoarthritis continues to be an ever-increasing, debilitating ailment for patients and health professionals must be prepared to treat and provide them with differing ways for symptomatic and functional relief. A wave of strategies including MSC therapy could provide an avenue for the prevention of the development of primary and secondary OA and using new-age technology such as genomics, transcriptomics, and functional assays can help scientists target this disease once and for all.
Author Contributions
A.V. and W.K. were responsible for the conceptualization of the study, methodology, manuscript preparation, and review. A.V. performed the initial formal analysis. Data curation was performed by D.A., K.K., N.C. and A.V. Project supervision and funding acquisition were performed by W.K., M.A.B. and A.W.M. All authors have read and agreed to the published version of the manuscript.
Funding
The authors gratefully acknowledge the financial support of Evelyn Medical Research Grant (Project Ref: 20-15) as well as of the Versus Arthritis (Formerly Arthritis Research UK) through Versus Arthritis Tissue Engineering and Regenerative Therapies Centre (Grant 21156).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
This scientific paper was supported by the Onassis Foundation - Scholarship ID: F ZS 069-1/2022-2023.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caplan, A.I. Mesenchymal stem cells: Time to change the name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. The development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen cells. Cell Tissue Kinet. 1970, 3, 393–403. [Google Scholar] [CrossRef]
- Ding, D.C.; Chang, Y.H.; Shyu, W.C.; Lin, S.Z. Human umbilical cord mesenchymal stem cells: A new era for stem cell therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef]
- Santilli, F.; Fabrizi, J.; Pulcini, F.; Santacroce, C.; Sorice, M.; Monache, S.D.; Mattei, V. Gangliosides and Their Role in Multilineage Differentiation of Mesenchymal Stem Cells. Biomedicines 2022, 10, 3112. [Google Scholar] [CrossRef]
- Martellucci, S.; Santacroce, C.; Manganelli, V.; Santilli, F.; Piccoli, L.; Cassetta, M.; Misasi, R.; Sorice, M.; Mattei, V. Isolation, propagation, and prion protein expression during neuronal differentiation of human dental pulp stem cells. J. Vis. Exp. 2019, 2019, e59282. [Google Scholar] [CrossRef]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef]
- Mortada, I.; Mortada, R. Epigenetic changes in mesenchymal stem cells differentiation. Eur. J. Med. Genet. 2018, 61, 114–118. [Google Scholar] [CrossRef]
- Yao, P.; Zhou, L.; Zhu, L.; Zhou, B.; Yu, Q. Mesenchymal Stem Cells: A Potential Therapeutic Strategy for Neurodegenerative Diseases. Eur. Neurol. 2020, 83, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Kassem, M.; Kristiansen, M.; Abdallah, B.M. Mesenchymal stem cells: Cell biology and potential use in therapy. Basic Clin. Pharmacol. Toxicol. 2004, 95, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Vallone, V.B.F.; Romaniuk, M.A.; Choi, H.; Labovsky, V.; Otaegui, J.; Chasseing, N.A. Mesenchymal stem cells and their use in therapy: What has been achieved? Differentiation 2013, 85, 1–10. [Google Scholar] [CrossRef]
- Khan, A.A.; Huat, T.J.; Al Mutery, A.; El-Serafi, A.T.; Kacem, H.H.; Abdallah, S.H.; Reza, M.F.; Abdullah, J.M.; Jaafar, H. Significant transcriptomic changes are associated with differentiation of bone marrow-derived mesenchymal stem cells into neural progenitor-like cells in the presence of bFGF and EGF. Cell Biosci. 2020, 10, 126. [Google Scholar] [CrossRef]
- George, S.; Hamblin, M.R.; Abrahamse, H. Differentiation of Mesenchymal Stem Cells to Neuroglia: In the Context of Cell Signalling. Stem Cell Rev. Rep. 2019, 15, 814–826. [Google Scholar] [CrossRef]
- Urrutia, D.N.; Caviedes, P.; Mardones, R.; Minguell, J.J.; Vega-Letter, A.M.; Jofre, C.M. Comparative study of the neural differentiation capacity of mesenchymal stromal cells from different tissue sources: An approach for their use in neural regeneration therapies. PLoS ONE 2019, 14, e0213032. [Google Scholar] [CrossRef]
- Hernández, R.; Jiménez-Luna, C.; Perales-Adán, J.; Perazzoli, G.; Melguizo, C.; Prados, J. Differentiation of human mesenchymal stem cells towards neuronal lineage: Clinical trials in nervous system disorders. Biomol. Ther. 2020, 28, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.T.J.; Gronthos, S.; Shi, S. Critical reviews in oral biology & medicine: Mesenchymal stem cells derived from dental tissues vs. those from other sources: Their biology and role in Regenerative Medicine. J. Dent. Res. 2009, 88, 792–806. [Google Scholar] [CrossRef] [PubMed]
- Mareschi, K.; Novara, M.; Rustichelli, D.; Ferrero, I.; Guido, D.; Carbone, E.; Medico, E.; Madon, E.; Vercelli, A.; Fagioli, F. Neural differentiation of human mesenchymal stem cells: Evidence for expression of neural markers and eag K+ channel types. Exp. Hematol. 2006, 34, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Black, I.B.; Woodbury, D. Adult rat and human bone marrow stromal stem cells differentiate into neurons. Blood Cells Mol. Dis. 2001, 27, 632–636. [Google Scholar] [CrossRef]
- Roberts, I. Mesenchymal stem cells Definition and characteristics of MSC Sources of MSC. Vox. Sang. 2004, 87, 38–41. [Google Scholar] [CrossRef]
- Yelick, P.C.; Zhang, W. Mesenchymal stem cells. In Tissue Engineering Principles and Practices; CRC Press: Boca Raton, FL, USA, 2012; pp. 10-1–10-14. [Google Scholar] [CrossRef]
- Bühring, H.J.; Battula, V.L.; Treml, S.; Schewe, B.; Kanz, L.; Vogel, W. Novel markers for the prospective isolation of human MSC. Ann. N. Y. Acad. Sci. 2007, 1106, 262–271. [Google Scholar] [CrossRef]
- Brooke, G.; Tong, H.; Levesque, J.P.; Atkinson, K. Molecular trafficking mechanisms of multipotent mesenchymal stem cells derived from human bone marrow and placenta. Stem Cells Dev. 2008, 17, 929–940. [Google Scholar] [CrossRef]
- Yang, Z.X.; Han, Z.-B.; Ji, Y.R.; Wang, Y.W.; Liang, L.; Chi, Y.; Yang, S.G.; Na Li, L.; Luo, W.F.; Li, J.P.; et al. CD106 Identifies a Subpopulation of Mesenchymal Stem Cells with Unique Immunomodulatory Properties. PLoS ONE 2013, 8, e59354. [Google Scholar] [CrossRef]
- Metcalf, D. Concise Review: Hematopoietic Stem Cells and Tissue Stem Cells: Current Concepts and Unanswered Questions. Stem Cells 2007, 25, 2390–2395. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.; Lee, H.-S.; Naidansaren, P.; Kim, H.-K.; O, E.; Cha, J.-H.; Ahn, H.-Y.; Yang, P.I.; Shin, J.-C.; Joe, Y.A. Proangiogenic features of Wharton’s jelly-derived mesenchymal stromal/stem cells and their ability to form functional vessels. Int. J. Biochem. Cell Biol. 2013, 45, 560–570. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, T.W.; McCabe, P.S.; McBeth, J. Update on the epidemiology, risk factors and disease outcomes of osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2018, 32, 312–326. [Google Scholar] [CrossRef]
- Glyn-Jones, S.; Palmer, A.J.R.; Agricola, R.; Price, A.J.; Vincent, T.L.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, P.; El-Jawhari, J.J.; Giannoudis, P.V.; Burska, A.N.; Ponchel, F.; Jones, E.A. Age-related Changes in Bone Marrow Mesenchymal Stromal Cells: A Potential Impact on Osteoporosis and Osteoarthritis Development. Cell Transplant. 2017, 26, 1520–1529. [Google Scholar] [CrossRef]
- Kempson, G.E.; Muir, H.; Swanson, S.A.V.; Freeman, M.A.R. Correlations between stiffness and the chemical constituents of cartilage on the human femoral head. BBA Gen. Subj. 1970, 215, 70–77. [Google Scholar] [CrossRef]
- Xia, B.; Chen, D.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Osteoarthritis Pathogenesis: A Review of Molecular Mechanisms. Calcif. Tissue Int. 2014, 95, 495–505. [Google Scholar] [CrossRef]
- Zhao, Q.; Ren, H.; Han, Z. Mesenchymal stem cells: Immunomodulatory capability and clinical potential in immune diseases. J. Cell. Immunother. 2016, 2, 3–20. [Google Scholar] [CrossRef]
- Samsonraj, R.M.; Raghunath, M.; Nurcombe, V.; Hui, J.H.; van Wijnen, A.J.; Cool, S.M. Concise Review: Multifaceted Characterization of Human Mesenchymal Stem Cells for Use in Regenerative Medicine. Stem Cells Transl. Med. 2017, 6, 2173–2185. [Google Scholar] [CrossRef] [PubMed]
- Biancone, L.; Bruno, S.; Deregibus, M.C.; Tetta, C.; Camussi, G. Therapeutic potential of mesenchymal stem cell-derived microvesicles. Nephrol. Dial. Transplant. 2012, 27, 3037–3042. [Google Scholar] [CrossRef] [PubMed]
- Kumaresan, S.; Vaiyapuri, S.; Kang, J.-H.; Dubey, N.; Manivasagam, G.; Yun, K.-D.; Park, S.-W. Perspective Chapter: Additive Manufactured Zirconia-Based Bio-Ceramics for Biomedical Applications; Intech: London, UK, 2022. [Google Scholar] [CrossRef]
- Cook, K.D.; Waggoner, S.N.; Whitmire, J.K. NK cells and their ability to modulate T cells during virus infections. Crit. Rev. Immunol. 2014, 34, 359–388. [Google Scholar] [CrossRef] [PubMed]
- Suchorska, W.M.; Augustyniak, E.; Richter, M.; Trzeciak, T. Gene expression profile in human induced pluripotent stem cells: Chondrogenic differentiation in vitro, part A. Mol. Med. Rep. 2017, 15, 2387–2401. [Google Scholar] [CrossRef]
- Marconi, A.; Hancock-Ronemus, A.; Gillis, J.A. Adult chondrogenesis and spontaneous cartilage repair in the skate, Leucoraja erinacea. bioRxiv 2019. [Google Scholar] [CrossRef]
- Kuettner, K.E. Biochemistry of articular cartilage in health and disease. Clin. Biochem. 1992, 25, 155–163. [Google Scholar] [CrossRef]
- Gale, A.L.; Linardi, R.L.; McClung, G.; Mammone, R.M.; Ortved, K.F. Comparison of the chondrogenic differentiation potential of equine synovial membrane-derived and bone marrow-derived mesenchymal stem cells. Front. Vet. Sci. 2019, 6, 178. [Google Scholar] [CrossRef]
- Goldring, M.B. Chondrogenesis, chondrocyte differentiation, and articular cartilage metabolism in health and osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 269–285. [Google Scholar] [CrossRef]
- Boeuf, S.; Richter, W. Chondrogenesis of mesenchymal stem cells: Role of tissue source and inducing factors. Stem Cell Res. Ther. 2010, 1, 31. [Google Scholar] [CrossRef]
- Mueller, M.B.; Fischer, M.; Zellner, J.; Berner, A.; Dienstknecht, T.; Prantl, L.; Kujat, R.; Nerlich, M.; Tuan, R.S.; Angele, P. Hypertrophy in mesenchymal stem cell chondrogenesis: Effect of TGF-β isoforms and chondrogenic conditioning. Cells Tissues Organs 2010, 192, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Robert, A.W.; Marcon, B.H.; Dallagiovanna, B.; Shigunov, P. Adipogenesis, Osteogenesis, and Chondrogenesis of Human Mesenchymal Stem/Stromal Cells: A Comparative Transcriptome Approach. Front. Cell Dev. Biol. 2020, 8, 561. [Google Scholar] [CrossRef] [PubMed]
- Karagianni, M.; Brinkmann, I.; Kinzebach, S.; Grassl, M.; Weiss, C.; Bugert, P.; Bieback, K. A comparative analysis of the adipogenic potential in human mesenchymal stromal cells from cord blood and other sources. Cytotherapy 2013, 15, 76–88.e2. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Jahagirdar, B.N.; Reinhardt, R.L.; Schwartz, R.E.; Keene, C.D.; Ortiz-Gonzalez, X.R.; Reyes, M.; Lenvik, T.; Lund, T.; Blackstad, M.; et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418, 41–49, Erratum in Nature 2007, 447, 879–880. [Google Scholar] [CrossRef]
- Maltman, D.J. Role of mesenchymal stem cells in neurogenesis and nervous system repair. Neurochem. Int. 2011, 59, 347–356. [Google Scholar] [CrossRef]
- Qu, C.; Puttonen, K.A.; Lindeberg, H.; Ruponen, M.; Hovatta, O.; Koistinaho, J.; Lammi, M.J. Chondrogenic differentiation of human pluripotent stem cells in chondrocyte co-culture. Int. J. Biochem. Cell Biol. 2013, 45, 1802–1812. [Google Scholar] [CrossRef]
- Zhang, Y.; Herman, B. Ageing and apoptosis. Mech. Ageing Dev. 2002, 123, 245–260. [Google Scholar] [CrossRef]
- McCulloch, K.; Litherland, G.J.; Rai, T.S. Cellular senescence in osteoarthritis pathology. Aging Cell 2017, 16, 210–218. [Google Scholar] [CrossRef]
- Marasa, B.S.; Srikantan, S.; Martindale, J.L.; Kim, M.M.; Lee, E.K.; Gorospe, M.; Abdelmohsen, K. MicroRNA profiling in human diploid fibroblasts uncovers miR-519 role in replicative senescence. Aging 2010, 2, 333–343. [Google Scholar] [CrossRef]
- Kapetanos, K.; Asimakopoulos, D.; Christodoulou, N.; Vogt, A.; Khan, W. Chronological age affects msc senescence in vitro—a systematic review. Int. J. Mol. Sci. 2021, 22, 7945. [Google Scholar] [CrossRef]
- Alzer, H.; Kalbouneh, H.; Alsoleihat, F.; Abu Shahin, N.; Ryalat, S.; Alsalem, M.; Alahmad, H.; Tahtamouni, L. Age of the donor affects the nature of in vitro cultured human dental pulp stem cells. Saudi Dent. J. 2021, 33, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Watthew, J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- OHAT Risk of Bias Rating Tool for Human and Animal Studies. Psychology. 2015. Available online: https://ntp.niehs.nih.gov/ntp/ohat/pubs/riskofbiastool_508.pdf (accessed on 30 August 2023).
- Scharstuhl, A.; Schewe, B.; Benz, K.; Gaissmaier, C.; Buhring, H.-J.; Stoop, R. Chondrogenic potential of human adult mesenchymal stem cells is independent of age or osteoarthritis etiology. Stem Cells 2007, 25, 3244–3251. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.-C.; Chou, H.-L.; Hung, W.-T.; Liu, H.-W.; Chu, T.-Y. Human adipose-derived stem cells cultured in keratinocyte serum free medium: Donor’s age does not affect the proliferation and differentiation capacities. J. Biomed. Sci. 2013, 20, 59. [Google Scholar] [CrossRef]
- Andrzejewska, A.; Catar, R.; Schoon, J.; Qazi, T.H.; Sass, F.A.; Jacobi, D.; Blankenstein, A.; Reinke, S.; Krüger, D.; Streitz, M.; et al. Multi-Parameter Analysis of Biobanked Human Bone Marrow Stromal Cells Shows Little Influence for Donor Age and Mild Comorbidities on Phenotypic and Functional Properties. Front. Immunol. 2019, 10, 2474. [Google Scholar] [CrossRef]
- Prall, W.C.; Saller, M.M.; Scheumaier, A.; Tucholski, T.; Taha, S.; Böcker, W.; Polzer, H. Proliferative and osteogenic differentiation capacity of mesenchymal stromal cells: Influence of harvesting site and donor age. Injury 2018, 49, 1504–1512. [Google Scholar] [CrossRef]
- Stolzing, A.; Jones, E.; McGonagle, D.; Scutt, A. Age-related changes in human bone marrow-derived mesenchymal stem cells: Consequences for cell therapies. Mech. Ageing Dev. 2008, 129, 163–173. [Google Scholar] [CrossRef]
- Alm, J.J.; Koivu, H.M.A.; Heino, T.J.; Hentunen, T.A.; Laitinen, S.; Aro, H.T. Circulating plastic adherent mesenchymal stem cells in aged hip fracture patients. J. Orthop. Res. 2010, 28, 1634–1642. [Google Scholar] [CrossRef]
- Fickert, S.; Schröter-Bobsin, U.; Groß, A.-F.; Hempel, U.; Wojciechowski, C.; Rentsch, C.; Corbeil, D.; Günther, K.P. Human mesenchymal stem cell proliferation and osteogenic differentiation during long-term ex vivo cultivation is not age dependent. J. Bone Miner. Metab. 2010, 29, 224–235. [Google Scholar] [CrossRef]
- Alt, E.U.; Senst, C.; Murthy, S.N.; Slakey, D.P.; Dupin, C.L.; Chaffin, A.E.; Kadowitz, P.J.; Izadpanah, R. Aging alters tissue resident mesenchymal stem cell properties. Stem Cell Res. 2012, 8, 215–225. [Google Scholar] [CrossRef]
- Siegel, G.; Kluba, T.; Hermanutz-Klein, U.; Bieback, K.; Northoff, H.; Schafer, R. Phenotype, donor age and gender affect function of human bone marrow-derived mesenchymal stromal cells. BMC Med. 2013, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Ruzzini, L.; Abbruzzese, F.; Rainer, A.; Longo, U.G.; Trombetta, M.; Maffulli, N.; Denaro, V. Characterization of age-related changes of tendon stem cells from adult human tendons. Knee Surg. Sports Traumatol. Arthrosc. 2014, 22, 2856–2866. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-H.; Ng, J.; Kim, S.-B.; Sonn, C.H.; Lee, K.-M.; Han, S.-B. Effect of Donor Age on the Proportion of Mesenchymal Stem Cells Derived from Anterior Cruciate Ligaments. PLoS ONE 2015, 10, e0117224. [Google Scholar] [CrossRef][Green Version]
- Maredziak, M.; Marycz, K.; Tomaszewski, K.A.; Kornicka, K.; Henry, B.M. The Influence of Aging on the Regenerative Potential of Human Adipose Derived Mesenchymal Stem Cells. Stem Cells Int. 2016, 2016, 2152435. [Google Scholar] [CrossRef]
- Kawagishi-Hotta, M.; Hasegawa, S.; Igarashi, T.; Yamada, T.; Takahashi, M.; Numata, S.; Kobayashi, T.; Iwata, Y.; Arima, M.; Yamamoto, N.; et al. Enhancement of individual differences in proliferation and differentiation potentials of aged human adipose-derived stem cells. Regen. Ther. 2017, 6, 29–40. [Google Scholar] [CrossRef]
- Palazzo, C.; Nguyen, C.; Lefevre-Colau, M.M.; Rannou, F.; Poiraudeau, S. Risk factors and burden of osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 134–138. [Google Scholar] [CrossRef]
- Blagojevic, M.; Jinks, C.; Jeffery, A.; Jordan, K.P. Risk factors for onset of osteoarthritis of the knee in older adults: A systematic review and meta-analysis. Osteoarthr. Cartil. 2010, 18, 24–33. [Google Scholar] [CrossRef]
- Silverwood, V.; Blagojevic-Bucknall, M.; Jinks, C.; Jordan, J.L.; Protheroe, J.; Jordan, K.P. Current evidence on risk factors for knee osteoarthritis in older adults: A systematic review and meta-analysis. Osteoarthr. Cartil. 2015, 23, 507–515. [Google Scholar] [CrossRef]
- Berran, Y.; Charles, L.E.; Baker, B.; Burchfiel, C.M. Occupational and genetic risk factors for osteoarthritis: A review. HHS Public Access. 2015, 50, 261–273. [Google Scholar] [CrossRef]
- Dahlin, R.L.; Meretoja, V.V.; Ni, M.; Kasper, F.K.; Mikos, A.G. Chondrogenic phenotype of articular chondrocytes in monoculture and co-culture with mesenchymal stem cells in flow perfusion. Tissue Eng. Part A 2014, 20, 2883–2891. [Google Scholar] [CrossRef]
- Yang, Y.H.K. Aging of mesenchymal stem cells: Implication in regenerative medicine. Regen. Ther. 2018, 9, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Lotza, M.; Loeserb, R.F. Effects of aging on articular cartilage homeostasis. Bone 2012, 51, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Payne, K.A.; Didiano, D.M.; Chu, C.R. Donor sex and age influence the chondrogenic potential of human femoral bone marrow stem cells. Osteoarthr. Cartil. 2010, 18, 705–713. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Sui, B.; Xiang, L.; Yan, X.; Wu, D.; Shi, S.; Hu, X. Emerging understanding of apoptosis in mediating mesenchymal stem cell therapy. Cell Death Dis. 2021, 12, 596. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).