1. Introduction
Ceratonia siliqua L. is an evergreen tree belonging to the
Leguminosae (
Fabaceae) family (subfamily
Caesalpinioideae) and it is widely known as the carob tree. Originating in the Middle East, it is widely encountered in the Mediterranean area, but it can grow in warm- or temperate-climate areas all over the world, even in arid and semiarid regions [
1]. The carob tree has been domesticated and cultivated since ancient times; hence, a great variety of local cultivars exist in every country [
2]. For instance, only in Southern Italy, fifty-four
C. siliqua cultivars have been described [
3]. In our days, the countries with the largest production of carob are Portugal, Italy, Spain, Morocco, Turkey, Greece, and Cyprus [
4], with approximately 80% of the Greek production concentrated in the island of Crete [
2].
C. siliqua cultivation throughout the years has aimed mainly at the collection of the fruits (carobs), but also at ornamental and other uses; e.g., nowadays, it is used against deforestation [
5,
6]. Carobs have the form of elongated pods (also known as locust beans) containing the pulp and the seeds (the latter usually account for 8–12% of the pod weight), while the pulp consists of a rough outer layer, the pericarp, and a soft inner layer, the mesocarp. The pulp comprises mainly sugars (mostly sucrose, and to a lesser extent glucose and fructose) and insoluble fiber (such as cellulose and hemicellulose), tannins, and other phenolic compounds (gallic acid and gallotannins), being poor in fat and protein [
1,
7]. The seeds contain much more protein and fewer sugars, as well as a low concentration of fat and many phenolic compounds [
1,
8,
9].
Since ancient times, carob has been used as food for humans and animals, and during hard times like World War II in Crete or the Spanish Civil War, it has saved many lives [
4,
10]. Beyond its nutritional uses, carob has always had multiple pharmaceutical applications, as indicated through traditional medicinal and ethnomedicinal reports [
11]. It has been proposed for glycemic control and antidiabetic activities, for combating hyperlipidemia, for anti-inflammatory effects especially in the digestive tract, for cytotoxic and anticancer effects, as well as for anxiolytic and antidepressant activities [
7,
12,
13,
14]. A less studied aspect regards the use of carob in the cosmetic industry, although the polysaccharide galactomannan (locust bean gum or E410) from the seed endosperm is already being used in the cosmetic sector, mainly due to its physical properties [
15]. A recent study suggested that carob extracts, beyond their well-known antioxidant properties, also possess photoprotective activity [
16].
The present study is part of an effort to support carob tree cultivation in Crete given its ecological advantages over other species, such as its very high fixation of carbon dioxide and its low requirements for water irrigation [
17]. As mentioned above, many different
C. siliqua cultivars exist in Crete, some of them imported from other countries. Based on the practical knowledge of the local producers, a typical cultivar existing in Crete for many years was selected as a primary target of the study, the so-called Imera (or Hemere) cultivar [
2]. This cultivar was recently subjected to the sequencing, assembly, and annotation of its genome, which resulted in the first, nearly chromosome-level assembly for
C. siliqua [
18]. The material used for this study was collected from trees in Pines in the area of Elounda Lasithiou. Another cultivar imported from Cyprus but acclimatized in Crete, the so-called Tylliria, was also studied for reasons of comparison [
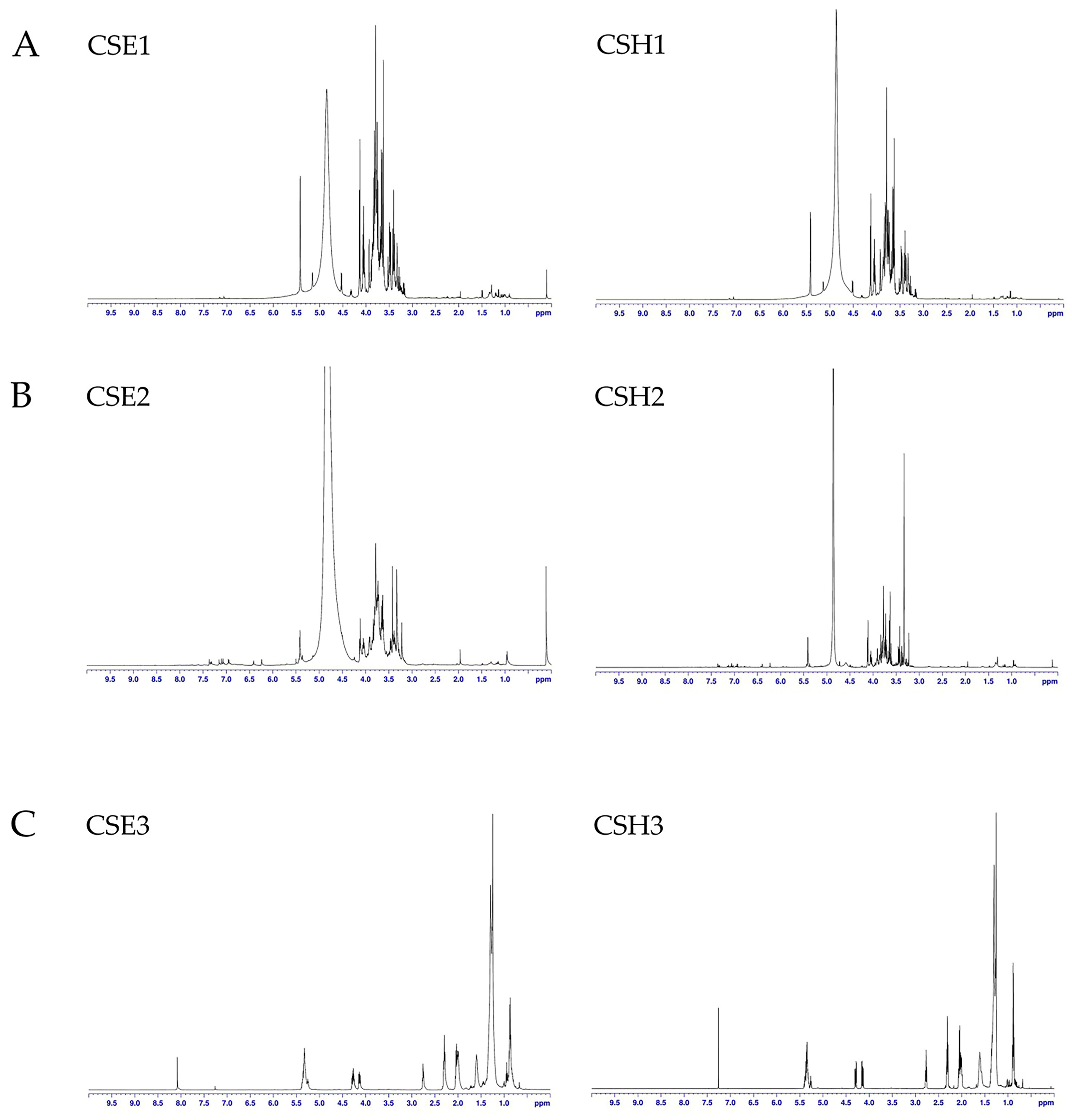
2]. The material used in this study was harvested from trees in Melidochori, near Heraklion. The local producers use Imera carobs mainly for flour production and for livestock food, while Tylliria is ideal for the production of carob syrup. Extracts from the two cultivars designated CSE (
C. siliqua Elounda) and CSH (
C. siliqua Heraklion) were subjected to a comparative study of their activities (including antioxidant, photoprotective, cytotoxic, and enzyme-modifying ones) on skin cell cultures and in cell-free assays, towards their putative use in cosmetic and other applications. Seed extracts were studied separately from deseeded pod extracts, since the former are considered by-products of flour production that may prove useful as additives in food, pharmaceutical, and cosmetic products [
19].
3. Discussion
The present study focuses on the comparative analysis of carob extracts obtained from two distinct
C. siliqua cultivars encountered in different geographical locations on the island of Crete. It has been shown previously that gender and cultivar significantly influence the chemical content and the biological activities of carob extracts with Portuguese origin [
33]. This also seems to be the case with the two Cretan cultivars of the present study, at least concerning the chemical content; only 11 out of 27 essential oil compounds (41%) were found to be common in the pods of the two cultivars, while also the fatty acid and the carbohydrate composition varied considerably (
Table 2,
Table 3 and
Table 4). The higher sucrose content of CSH compared to CSE reflects the knowledge of the local producers, who prefer the first cultivar for the production of carob syrup. As far as we know, there is only one study concerning the fatty acid and carbohydrate content of Cretan carob fruits—without, however, more information on the selected cultivars—assessing their composition in three different growth stages, the second one being the most appropriate for comparison with our results, based on the collection period of the plant material [
34]. Our results are in line with the above study concerning palmitic acid, but there are certain differences regarding ω3 and ω6 fatty acids since we detected lower amounts of linolenic acid and higher percentages of the linoleic one (
Table 3). Moreover, our results agree regarding the content of the main carob carbohydrate (sucrose), with minor differences in the glucose and fructose contents. Still, the fatty acid and carbohydrate contents determined in both studies are within the limits that have been generally described in the literature [
35,
36].
The extracts from the two Cretan cultivars were further subjected to a comparative study of a series of biological activities related to their potential for use in cosmetics. Their effect on human skin fibroblast viability was evaluated and none of the carob extracts exhibited cytotoxicity up to the concentration of 200 μg/mL (
Table S1). On the other hand, some of the fractions, i.e., CSE1.2 and CSE2.1 (under serum-free and low-serum conditions), did result in reduced viability; hence, lower noncytotoxic concentrations were chosen for the respective experiments (
Table S2). We did not locate any other studies in the literature regarding carob extracts’ effects on human skin fibroblasts; however, liposome encapsulated
C. siliqua pod extracts tested on 3T3 embryonic mouse fibroblasts, and immortalized human keratinocytes (HaCaT) have been reported to be noncytotoxic [
37], in accordance with our findings. There are many studies in the literature regarding carob extracts’ cytotoxicity towards a variety of different cancer cell lines [
33,
38,
39,
40,
41,
42,
43,
44,
45,
46,
47,
48,
49,
50,
51]. Cytotoxic activity has been mainly found with polar extracts and is believed to be a result of synergy between many compounds contained in carob [
7,
44]. Although not directly related to the scope of the present study, we tested the extracts from the two Cretan cultivars on human epidermoid carcinoma (A431) and human fibrosarcoma (HT-1080) cells. CSE3 at the highest concentration tested was marginally cytotoxic for A431 cells, while fraction CSE1.2, and to a lesser extent fractions CSE1.1 and CSE2.1, were cytotoxic for both cell lines (
Tables S1 and S2). A 30% aqueous methanolic leaf extract from the Imera (Elounda) cultivar was also cytotoxic for HT-1080 fibrosarcoma cells (
Table S3). Regarding the different efficacies observed among the various fractions, these may be related to the amount of carbohydrates, such as sucrose, which may exert protective effects, counteracting the cytotoxic activities of other constituents [
40,
52]. The cytotoxic activities of Cretan carob extracts and their fractions against cancer cells are being further investigated in a separate study.
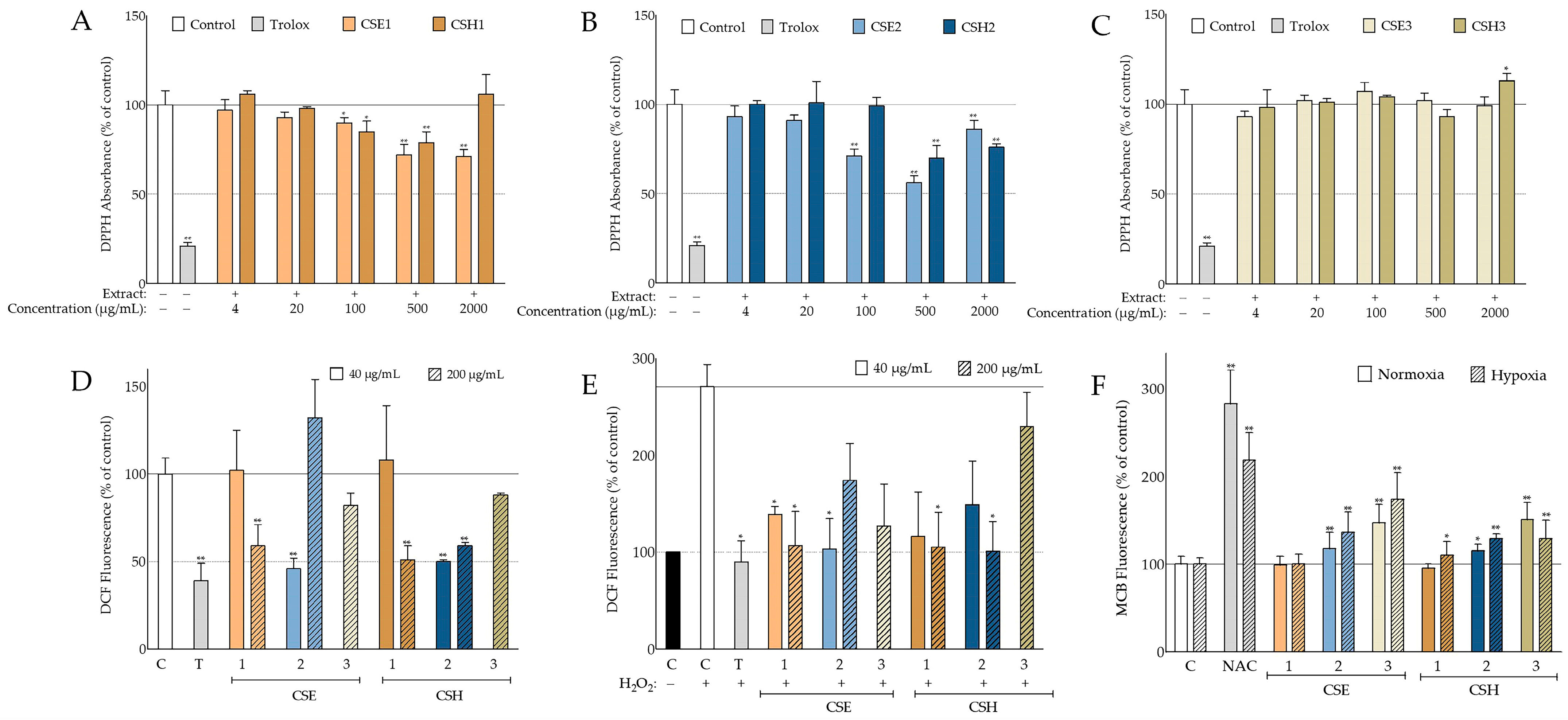
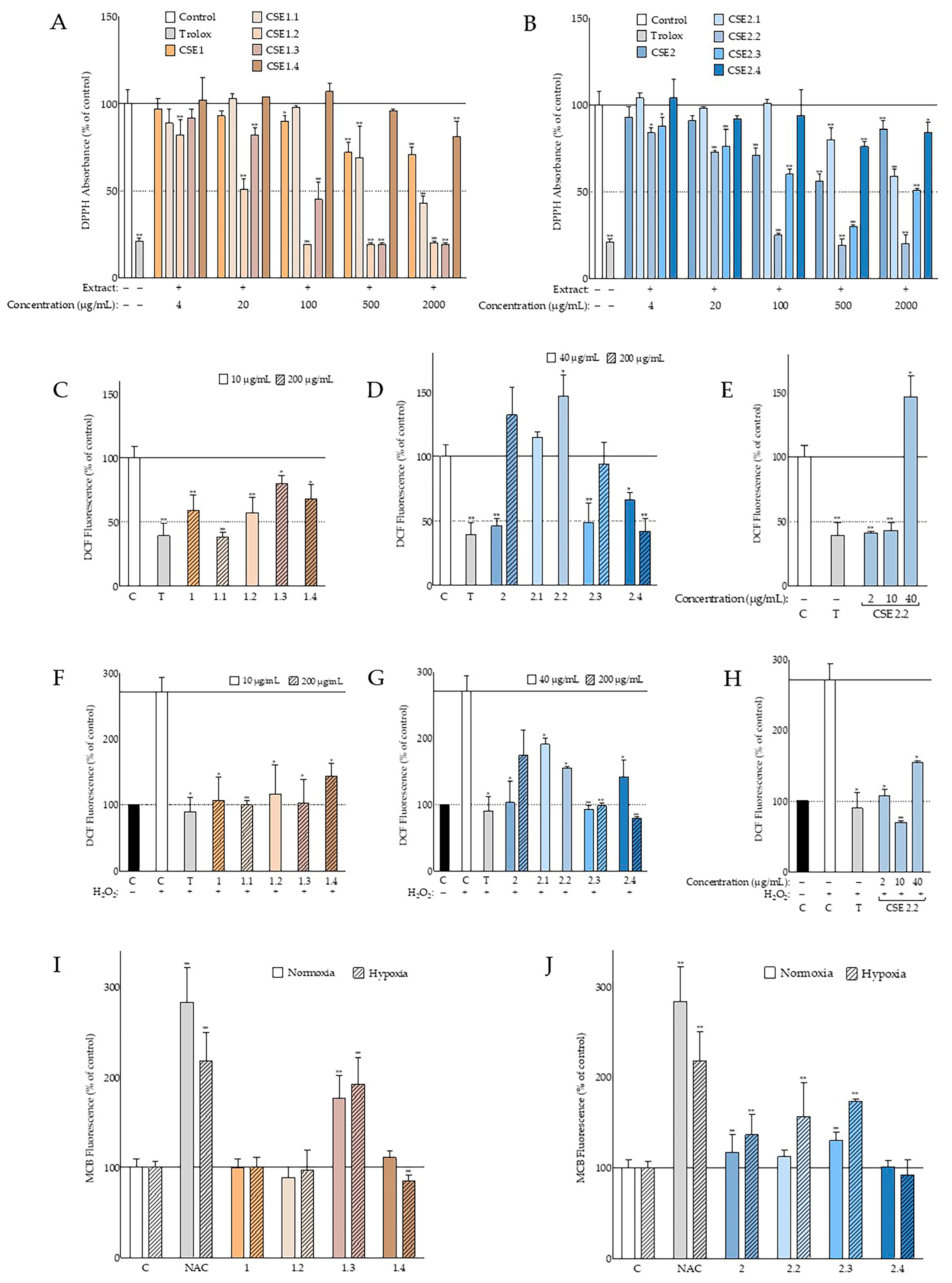
A main target of our study was the assessment of the antioxidant activity of the extracts from the two Cretan carob tree cultivars. The antioxidant activity of C.
siliqua extracts was extensively investigated in cell-free systems. Previously, higher DPPH radical scavenging has been reported with methanolic carob leaf extracts compared to pulp ones, as well as a correlation between antioxidant activity and gender [
53]. Leaf decoctions were also found to be more active than the pulp and seed ones [
54], as well as more potent in total flavonoids, tannins, and phenols in comparison to the pulp [
55]. Their results are in accordance with our findings since carob leaf 30% aqueous methanolic extract, CSE4, from the Imera cultivar (Elounda) was the only extract capable of reducing DPPH absorbance more than 50%, hence allowing the calculation of its IC
50 at 294 ± 35 μg/mL (
Table S3). On the other hand, the scavenging abilities of aqueous and methanolic carob leaf extracts from Turkey have been reported to be higher than the above calculated for the Imera cultivar [
56]. Regarding the role of the solvent selected for extraction, our results from the seed extracts of both studied cultivars indicate that polar solvents are more efficient for the recovery of the DPPH-scavenging activities (
Figure 2B vs. 2C), in agreement with previous studies using DPPH or a similar method, i.e., 2,2′-azino-bis[3-ethylbenzthiazoline-6-sulphonic acid] (ABTS) [
57,
58]. In these studies, as well as in the present one, inferior activity for hexane carob pulp and/or seed extracts compared to aqueous and methanolic ones was observed. Other solvent systems, such as 80% aqueous acetone [
59,
60], have also been reported to be very efficient for deseeded carob pod extraction.
In general, the IC
50 values reported in the literature for methanolic carob pod extracts vary from around 100–200 μg/mL [
61] to around 10 mg/mL [
33], and this may be due to the different geographical origins, the cultivar, the gender, or the ripening stage [
62]. Regarding the two Cretan cultivars presented here, the reduction in DPPH absorbance for CSE1 and CSH1 (
Figure 2A) was not sufficient to calculate an IC
50 (the highest concentration tested being 2 mg/mL); however, IC
50 values of 1177 ± 240, 17 ± 1, and 79 ± 22 μg/mL were obtained from fractions CSE1.1, CSE1.2, and CSE1.3, respectively (
Figure 6A). It is important to note that the carobs used in this study had reached maturity, while more potent antioxidant activities have been reported for unripe carob pulp extracts than ripe ones [
49,
60,
63,
64]. On the other hand, ethyl acetate ripe seed extracts had lower IC
50 values when compared to the unripe ones [
49]. Regarding the carob seed extracts, the best antioxidant activity was exhibited by the 30% aqueous methanolic extract CSE2 that managed to decrease DPPH radical absorbance to 56% of the negative control at 500 μg/mL and at 71% at 100 μg/mL (
Figure 2B). This efficacy was comparable to other studies based on Moroccan carob seeds [
65] but inferior to another using carob seeds from Algeria [
66]. However, the fractions CSE2.2 and CSE2.3 of the seed extract yielded much better IC
50 values (35 ± 5 and 140 ± 68 μg/mL, respectively;
Figure 6B).
The promising results obtained from the cell-free assay led us to evaluate the antioxidant capacity of the carob extracts under study in the cellular context. Indeed, the extracts from both Cretan cultivars proved to be efficient in suppressing not only the basal ROS levels in human skin fibroblast cultures, but also the increase in ROS levels provoked by hydrogen peroxide (
Figure 2D,E). Certain extracts and fractions were found to be as potent as the positive control (Trolox) (
Figure 2D,E and
Figure 6C–H). The inferior antioxidant activity following extraction with hexane compared to the 30% aqueous-methanol system, that was observed in the DPPH experiments, was also supported by the intracellular ROS levels. Our results agree with findings in the literature, where pretreatment with
C. siliqua pod extract was shown to ameliorate the viability of 3T3 embryonic mouse fibroblasts, immortalized human keratinocytes, and A431 squamous carcinoma cells exposed to H
2O
2 [
37], while an aqueous carob pod extract was found to protect against H
2O
2-induced DNA damage [
42]. Interestingly, reducing effects of carob extract on intracellular ROS levels have been also reported for human sperm cells [
67]. Extracts from other parts of the carob tree, such as sapwood and leaf, have been shown to revert H
2O
2-stimulated intracellular ROS levels of HeLa cells [
51,
51], though germ flour extract was observed to increase ROS levels in the same cell type [
47]. To our knowledge, this is the first study reporting the effects of
C. siliqua extracts and fractions on basal and H
2O
2-challenged ROS levels of human skin fibroblasts.
A further approach for assessing the antioxidant properties of the carob extracts was the evaluation of the intracellular GSH levels (
Figure 2D and
Figure 6I,J). Both seed extracts were found to increase the GSH levels of human skin fibroblasts, as well as some of the fractions tested. The fact that hexane seed extracts CSE3 and CSH3—that are expected to contain fatty acids, among others—exhibited high activities in this assay may be related to previous observations that the exposure of endothelial cells to fatty acids for over 12 h leads to an increase in GSH levels [
68]. In general, under hypoxic conditions, the percentage of GSH stimulation was higher compared to normoxic ones, most probably due to the fact that hypoxia minimized basal GSH levels, as previously shown in various cell types [
69,
70]. Although there are several reports on the beneficial effects of various carob extracts on the GSH levels in whole organisms, mainly as part of their antioxidant defenses and during detoxification processes [
71,
72,
73,
74], this is the first study showing that
C. siliqua extracts stimulate GSH at the cellular level.
The above protective effects of the Cretan carob extracts against ROS and oxidative stress were also evident when the activation of two signaling molecules, i.e., PKB/Akt and AMPKα was examined. Akt is considered a key molecule promoting cell survival [
24], while AMPK is characterized as a guardian of metabolism and mitochondrial homeostasis [
75]. AMPK is a downstream target of Akt activation; however, it has been recently shown that AMPK itself can activate Akt under cellular stress conditions [
76]. We observed that the treatment of human skin fibroblasts with carob seed extract induced Akt activation (
Figure 3A), suggesting an augmentation of the survival cell mechanisms. Pretreatment of the cells with seed extracts from both Cretan
C. siliqua cultivars enhanced the subsequent H
2O
2-induced Akt activation (
Figure 3B), further supporting cell survival, while on the other hand, it suppressed AMPKα phosphorylation. Other natural products with antioxidant and survival activities have been also shown to suppress H
2O
2-induced AMPKα phosphorylation [
77]. Interestingly, the extracts obtained with hexane as a solvent (CSE3 and CSH3), which had shown inferior antioxidant capacity in the DPPH and DCFH-DA assays compared to the 30% aqueous methanolic ones (CSE2 and CSH2;
Figure 2), were also unable to suppress AMPKα phosphorylation (
Figure 3B). Interestingly, a recent study reported that in a different cellular context, i.e., in breast cancer cells, carob extracts showed the ability to suppress Akt phosphorylation, hence suppressing the detrimental cancer cell survival [
49].
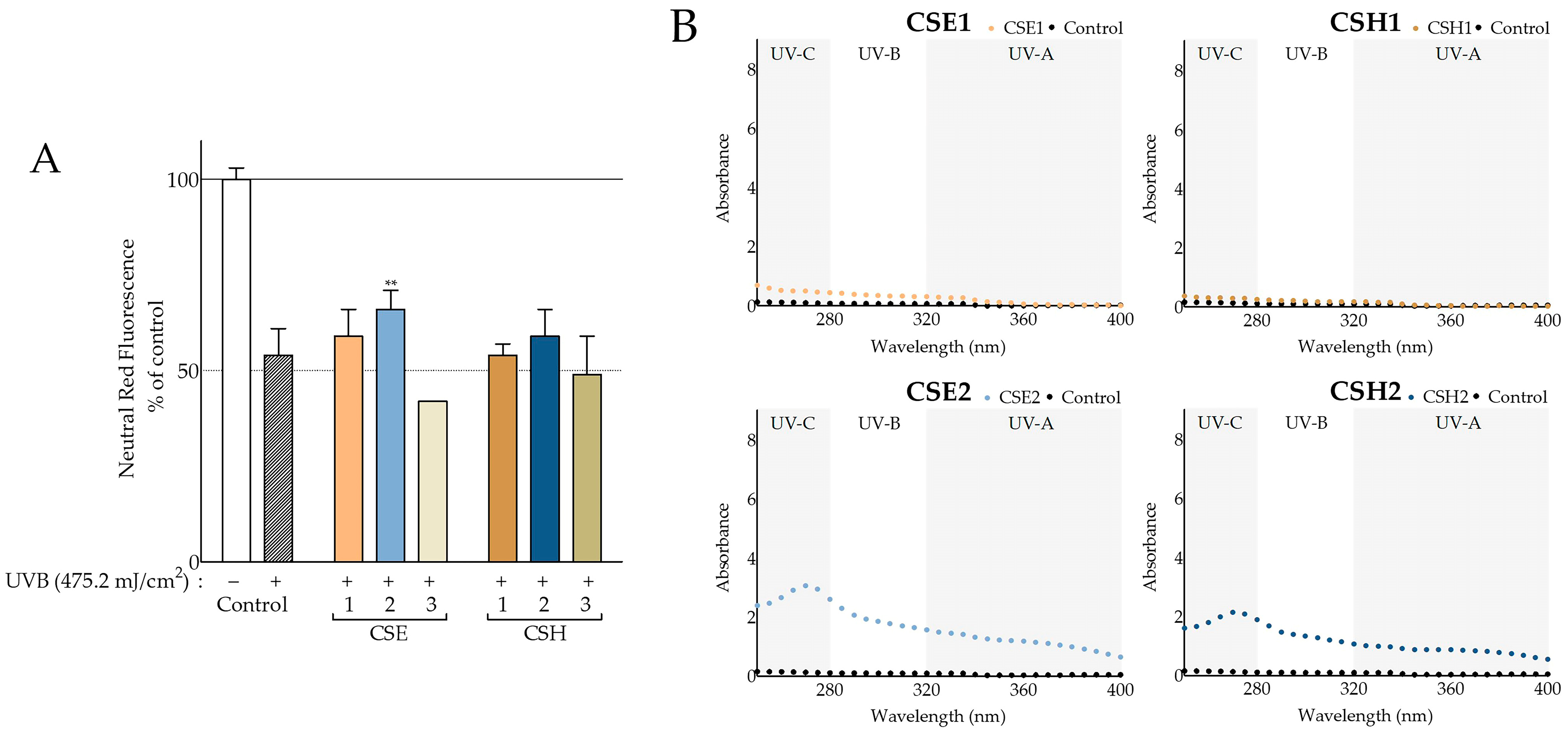
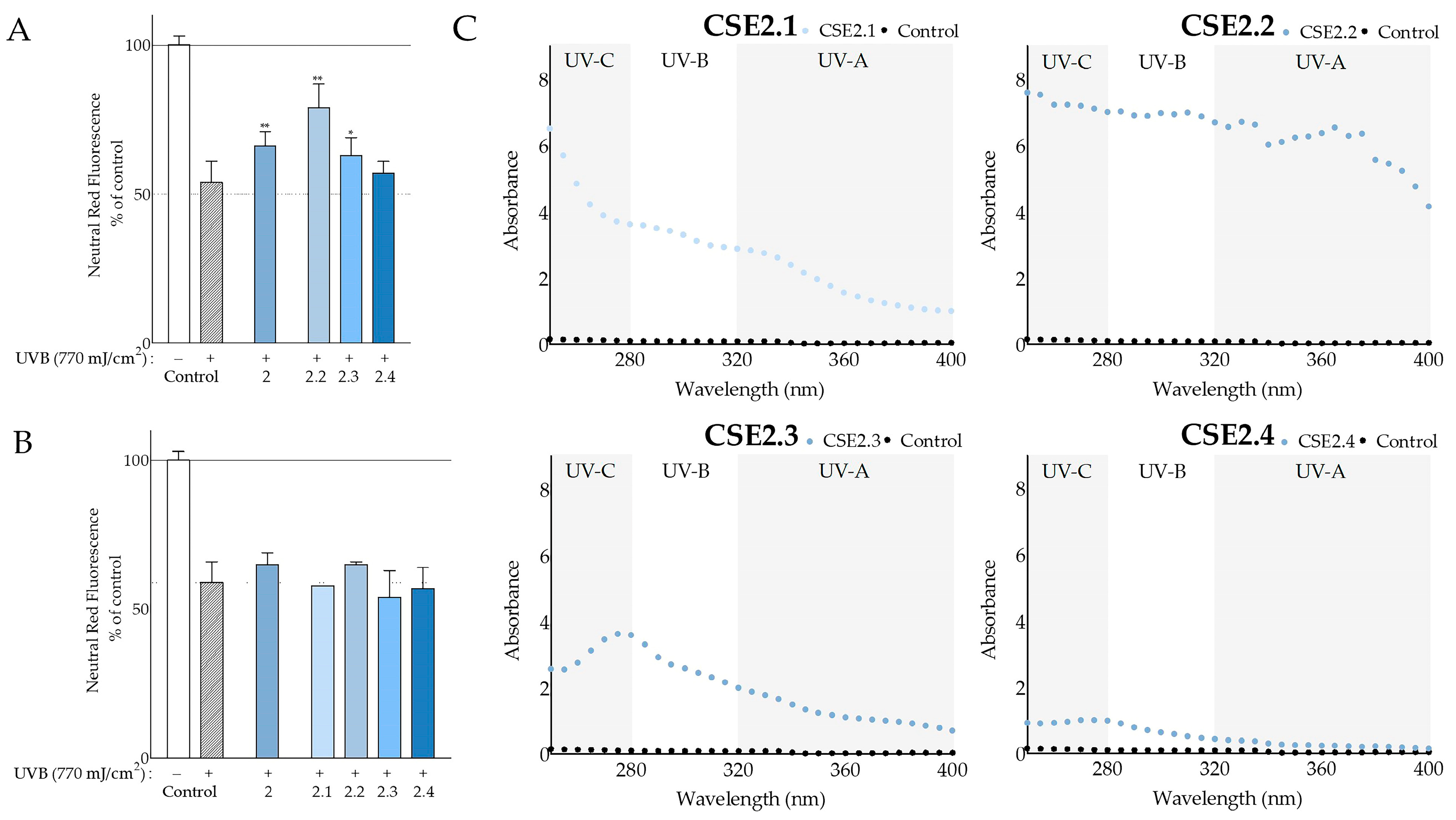
Beyond the antioxidant properties, the ability of the carob extracts to protect against UV radiation was evaluated, being a feature of great importance to cosmetics. This was assessed at the level of simple UV-screening, as well as at the more complicated level of conferring protection against UV-induced cytotoxicity. The screening capacity was found to be enhanced in aqueous methanolic seed extracts of both cultivars (
Figure 4B) compared to deseeded pod extracts, in agreement—at least qualitatively—with observations based on plant material from other countries [
78]. Overall, the aqueous methanolic seed extracts from both Cretan cultivars absorbed more than the hexane seed extracts. A recent study on the effects of processing in ethanolic
C. siliqua pod extracts regarding their antioxidant and photoprotective potential [
16] included a calculation of in vitro SPF, which ranged from 8.62 to 22.37. In our study, we found lower SPF values, ranging from 2.14 to 3.95, regarding the deseeded pod extracts. Even so, similar SPF values of 18.19 and 13.07 were obtained from CSE2 and CSH2, respectively. A high SPF value (32.44) was also observed with the leaf extract, CSE4 (
Table S3). Furthermore, we determined even higher SPF values with carob seed extract fractions CSE2.1, CSE2.2, and CSE2.4 of 31.97, 69.87, and 25.06, respectively. To the best of our knowledge, this is the first study to examine the photoprotective effect of carob extracts on UVB-treated fibroblasts. We showed an increase in the viability of fibroblasts exposed to UVB irradiation that had been pretreated with CSE2 (
Figure 4A), which might be related to its high SPF value. In addition, its fraction CSE2.2 showed even greater photoprotective qualities in both assays (
Figure 7).
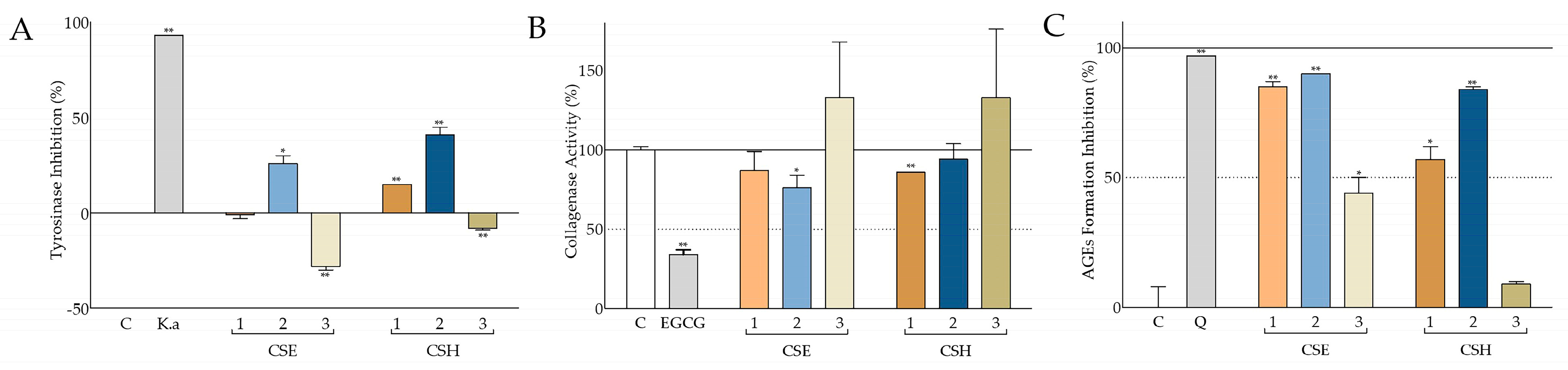
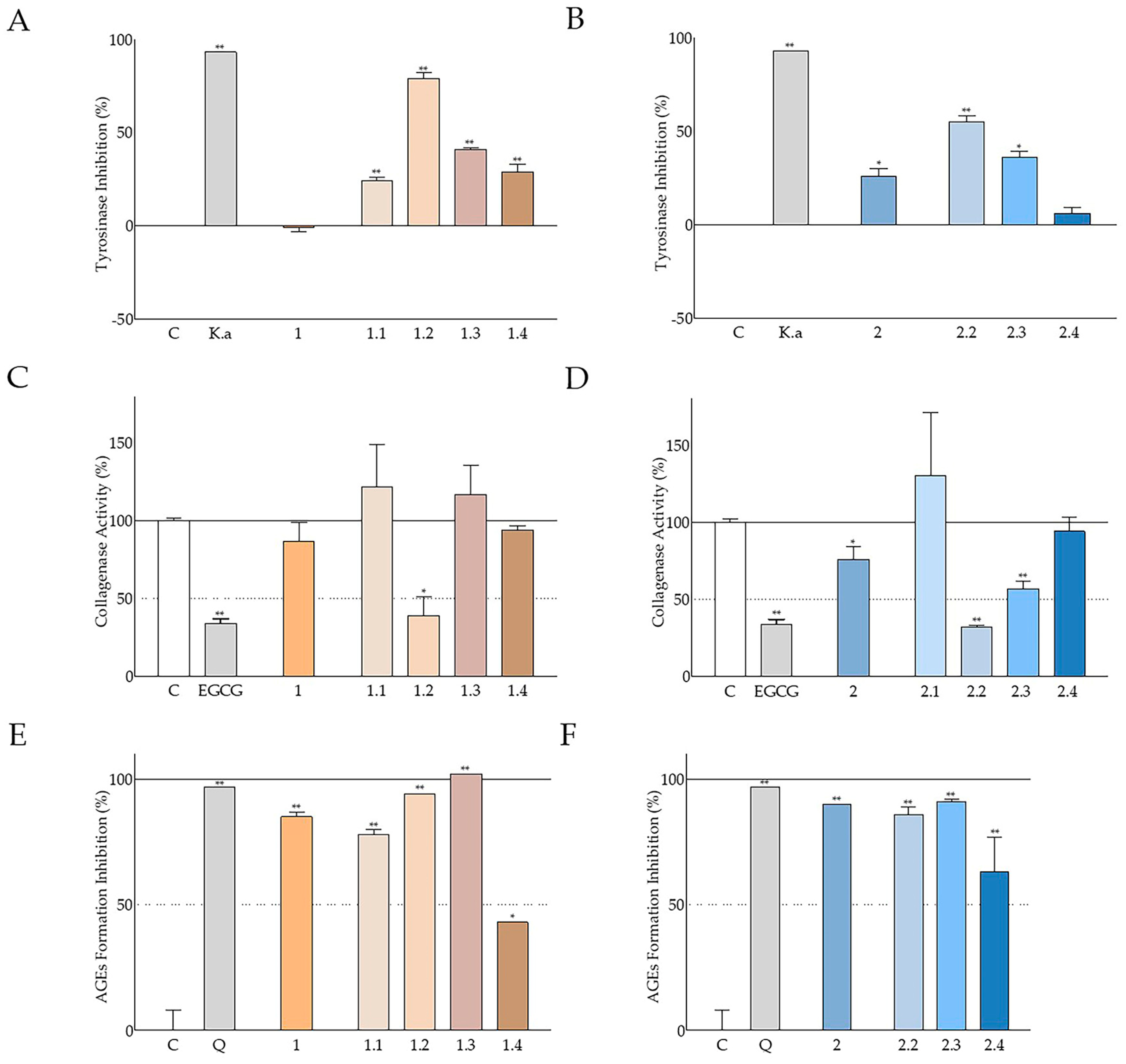
In the present study, a significant inhibition of tyrosinase activity was observed, mainly by the aqueous methanolic seed extracts from both cultivars, and to a lesser extent by the deseeded pod extract from Heraklion (
Figure 5A). The hexane seed extracts were inactive in this assay. Following fractionation, the best antityrosinase activity was observed with the fractions partitioned with ethyl acetate and butanol (
Figure 8A,B). A similar though less intense inhibitory activity against tyrosinase has been reported for a Turkish carob ethanolic seed extract [
79]. In a previous study of the antimelanogenic properties of
C. siliqua ethanolic leaf, bark, and fruit extracts, monophenolase activity was found to be potently inhibited by all extracts, while diphenolase activity was inhibited mainly by the leaf extract [
80], in agreement with our observations regarding the aqueous methanolic leaf extract from the Imera cultivar (Elounda;
Table S3). Comparable inhibitory effects have been also reported with carob leaf methanolic and aqueous extracts from Turkish plants [
56].
Though the inhibitory effects against collagenase activity were not so intense, statistically significant reduction in the activity of this enzyme was mainly observed with the leaf extract (CSE4;
Table S3) and to a lesser extent with seed extract CSE2 and deseeded pod extract CSH1 (
Figure 5B). However, upon fractionation,
C. siliqua pod and seed extract fractions CSE1.2, CSE2.2, and CSE2.3 were found to be very active, with CSE1.2 and CSE2.2 even attenuating collagenase activity to a similar extent to the reference compound (EGCG) (
Figure 8C,D). Even though locust bean gum, a polysaccharide from
C. siliqua seed, has been found to inhibit collagenase [
81], as far as we know, this is the first study assessing the effect of carob extracts on collagenase activity.
Natural products’ interference with AGE formation is usually studied as a means to counteract diabetic complications; however, it is also important for cosmetics [
82]. Our findings regarding the ability of all aqueous-methanol carob extracts, from both Cretan cultivars, as well as their fractions to inhibit glucose-induced AGE formation were very promising, since CSE1 and CSE2 had activities comparable to the positive control (quercetin;
Figure 5C), whilst fraction CSE1.3 completely eliminated AGE formation (
Figure 8E). Again, hexane extracts were found to be less active (
Figure 5C), while the leaf extract CSE4 (
Table S3) was highly active, in agreement with another study in the literature [
83].
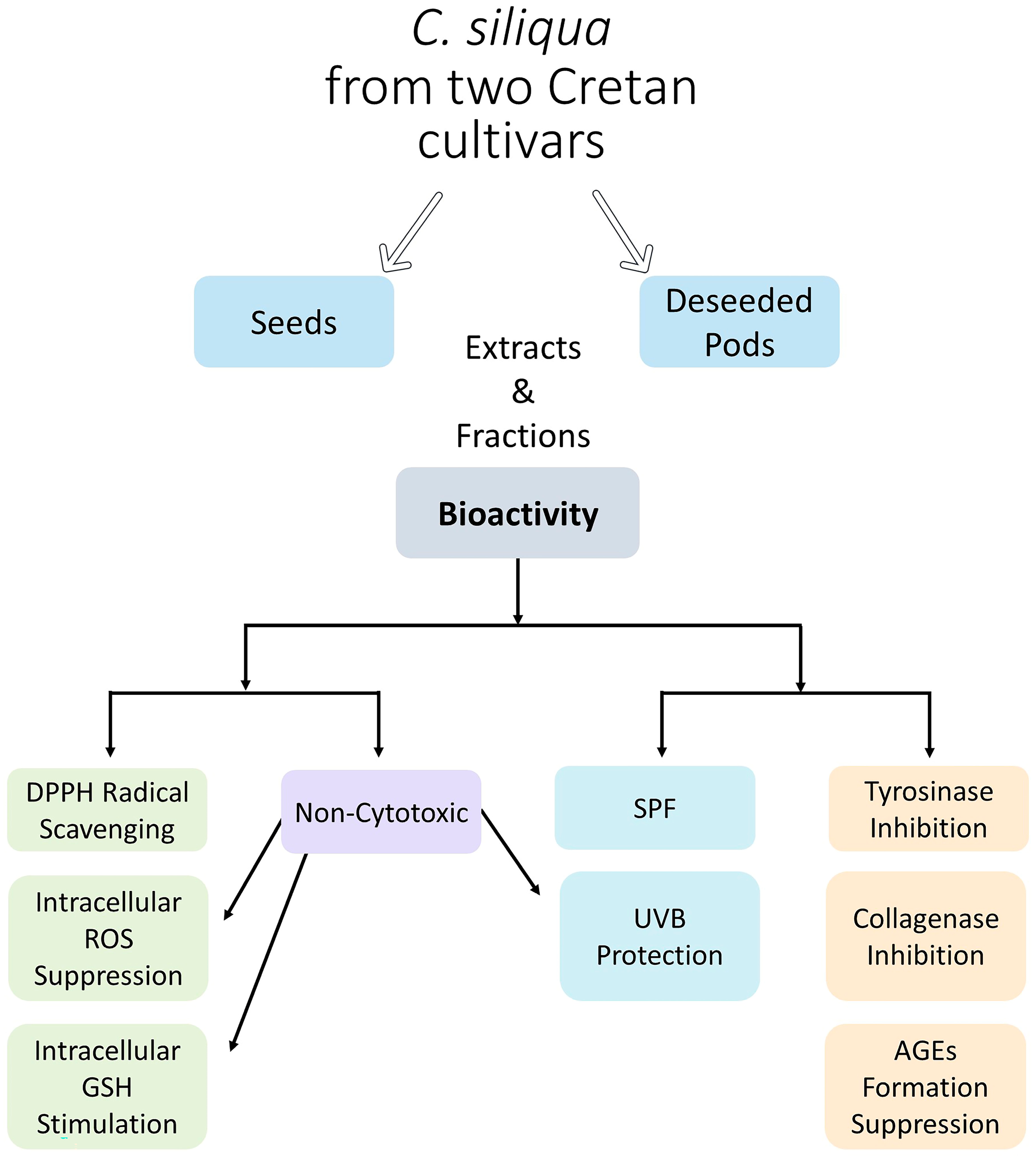
All the above activities characterizing carob extracts and studied either on human skin fibroblasts in vitro or in cell-free assay systems were, for the first time, investigated from a perspective aiming at the putative use of these extracts in cosmetics. Indeed, the cumulative capacity of these extracts for scavenging free radicals, suppressing ROS, stimulating GSH, inhibiting tyrosinase and collagenase activities, blocking glucose-induced AGEs, and in certain cases counteracting UVB effects (
Scheme 1) makes them ideal candidates for skin care products.


 ,
, 












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}