Identification of 5-Hydroxymethylfurfural (5-HMF) as an Active Component Citrus Jabara That Suppresses FcεRI-Mediated Mast Cell Activation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
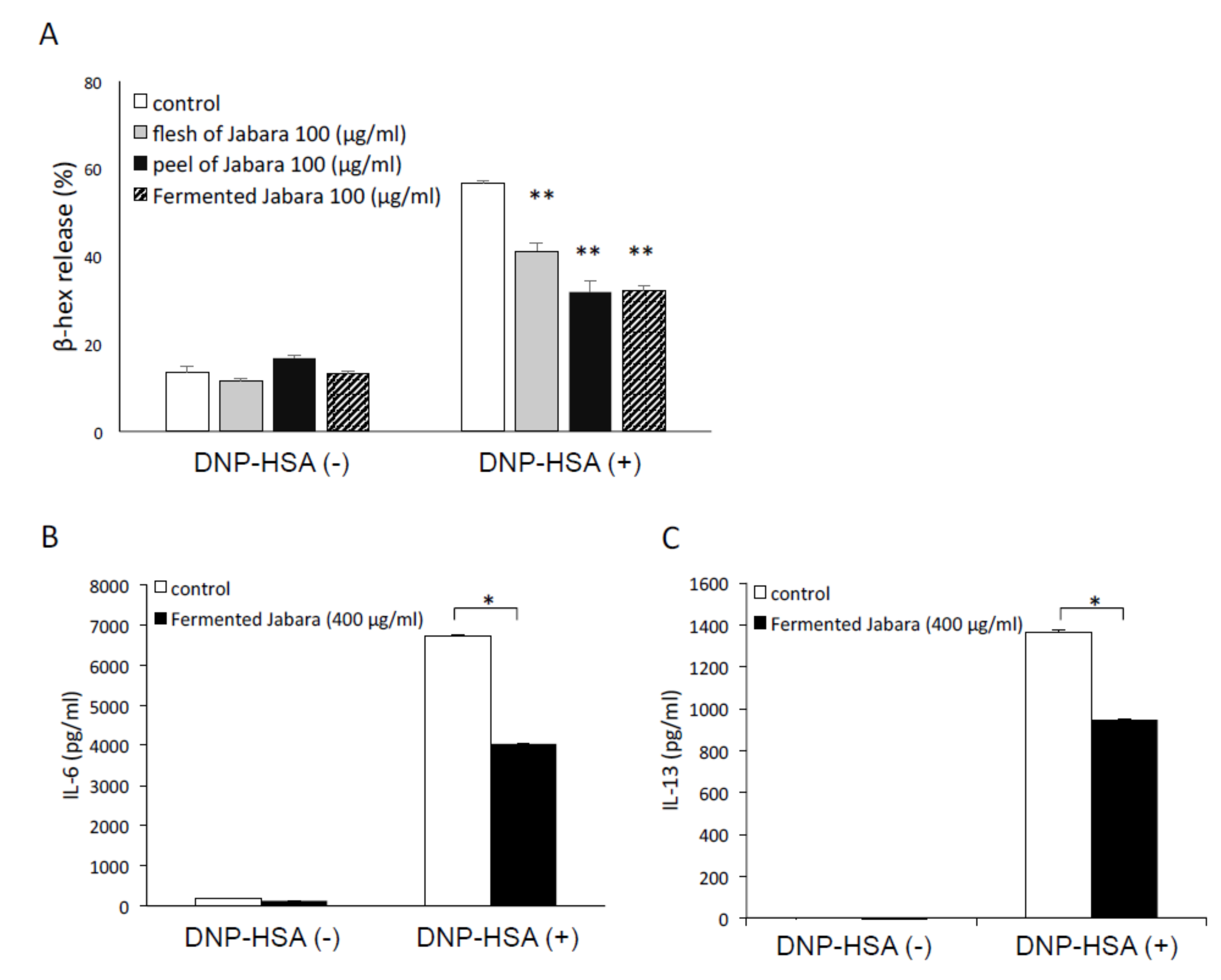
2.1. Fermented Jabara Suppressed Mast Cell Activation
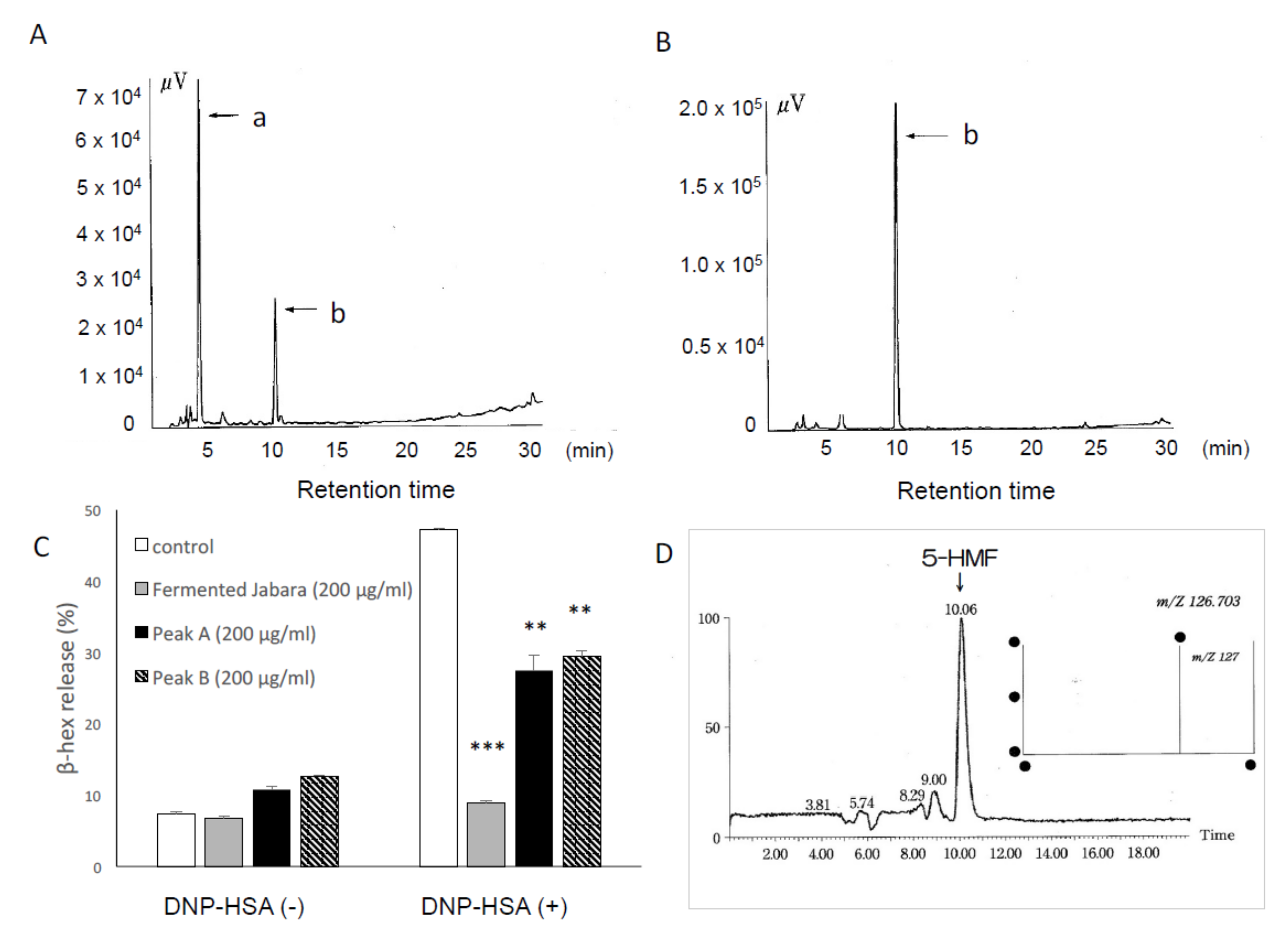
2.2. 5-HMF is the Active Component in Fermented Jabara that Inhibits Mast Cell Activation
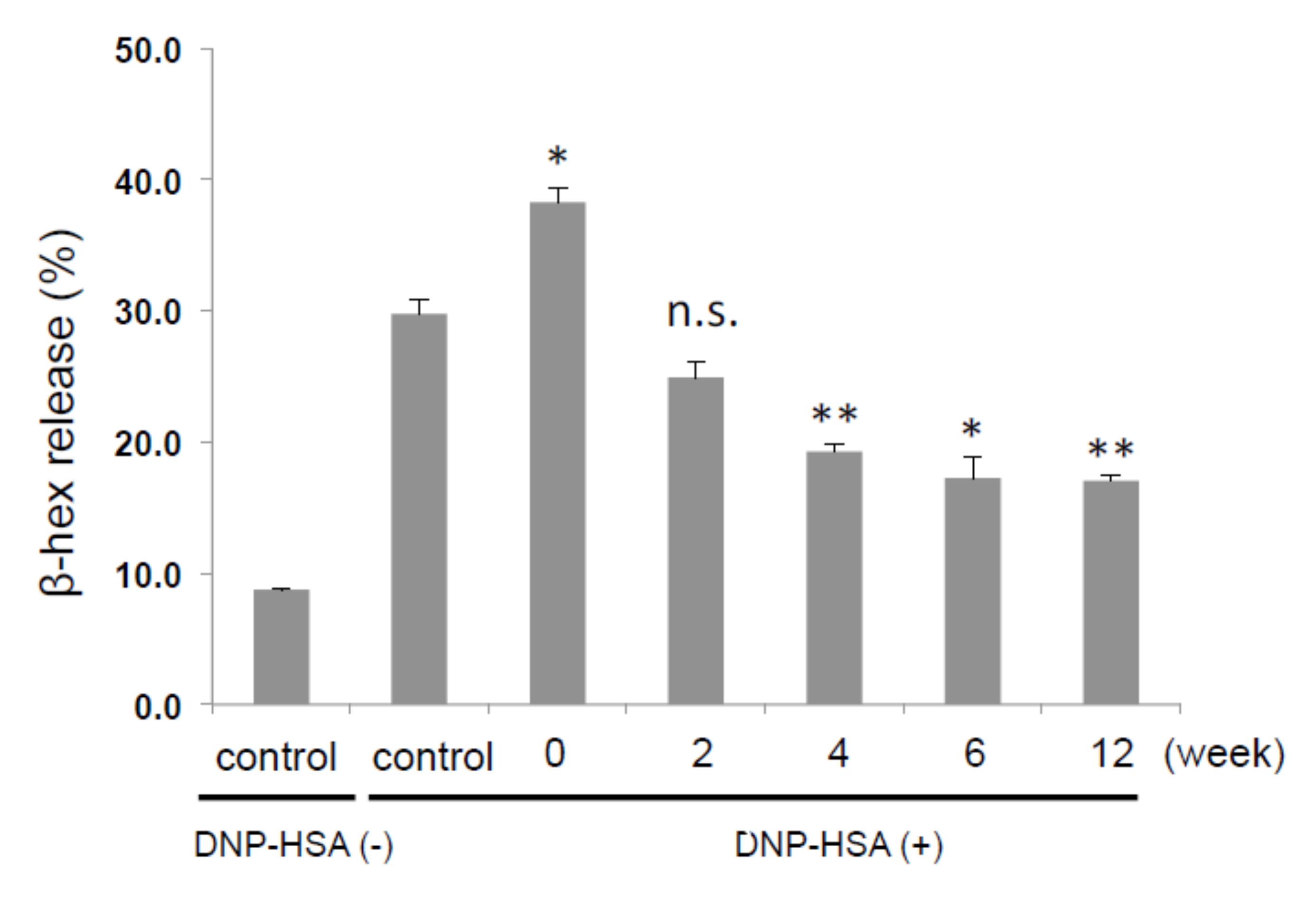
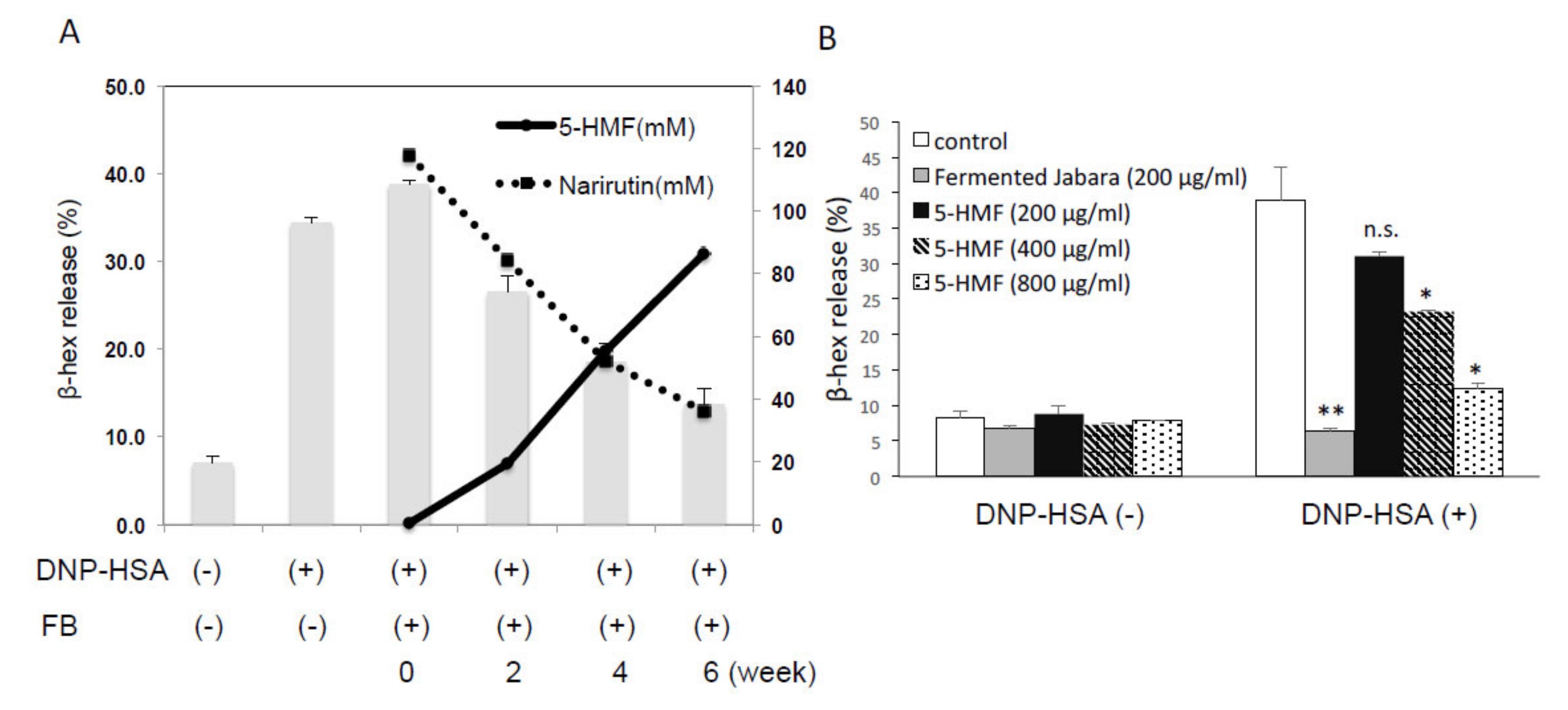
2.3. 5-HMF Increased as Fermentation Progressed
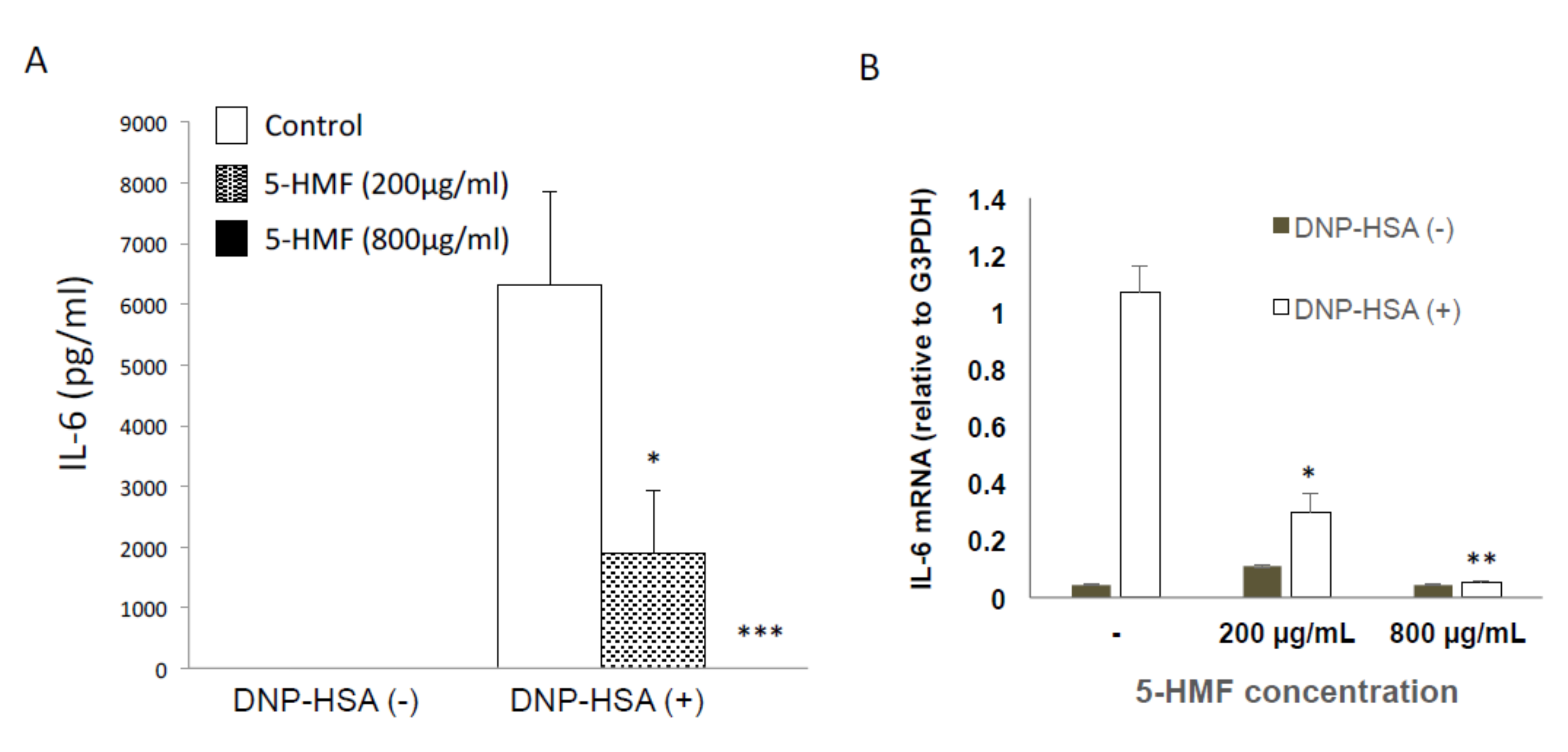
2.4. Effect of 5-HMF on the Expression of Proinflammatory Cytokine
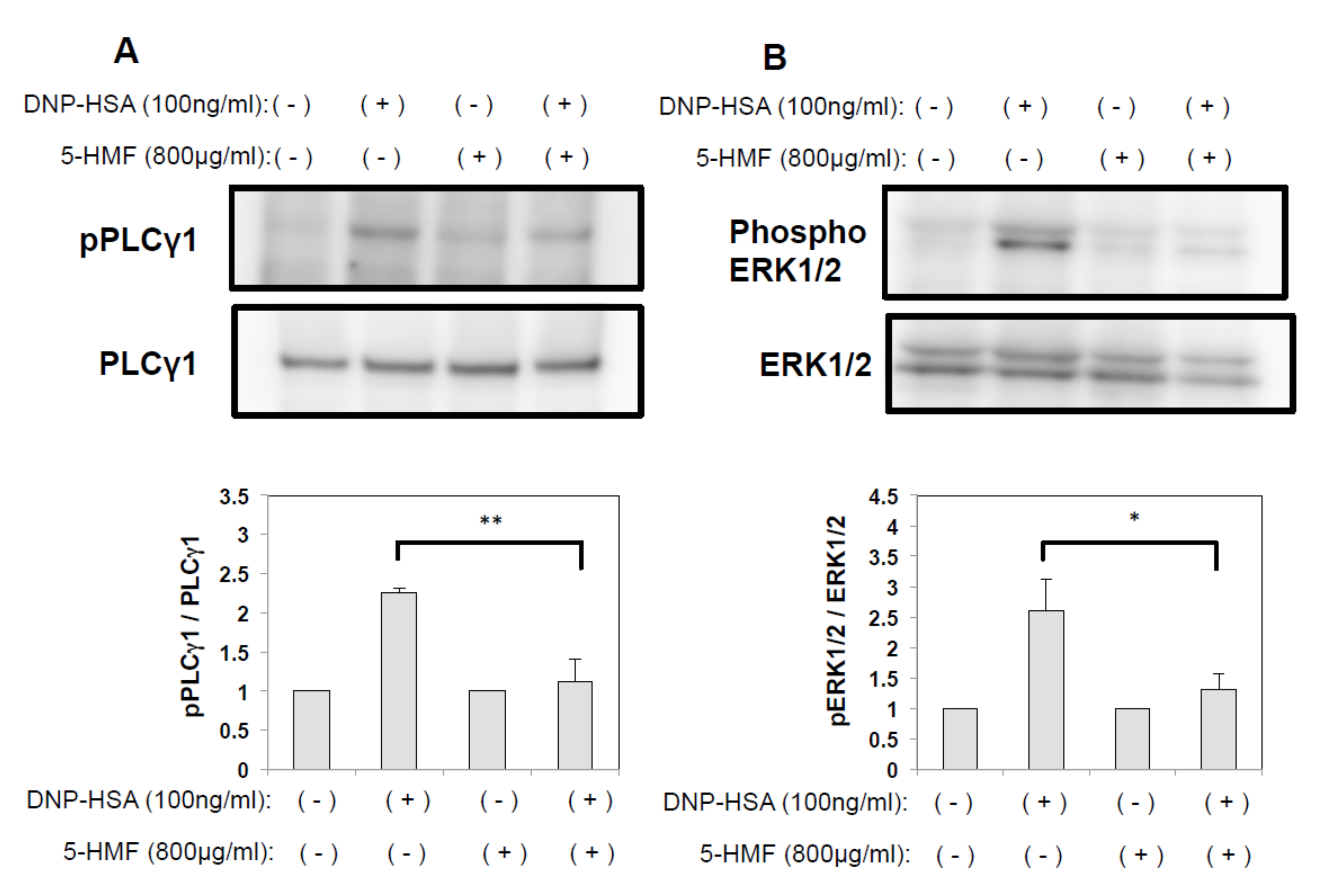
2.5. The effect of 5-HMF on the PLCγ and MAPK Signaling Pathways
3. Discussion
4. Materials and Methods
4.1. Fermentation of Jabara
4.2. Reagents and Antibodies
4.3. Mice
4.4. Cell Culture
4.5. BMMC Degranulation Assay
4.6. Measurement of Cytokines
4.7. Extraction from Fermented Jabara
4.8. Identification of the Active Component from Fermented Jabara
4.9. Cell lysates and Immunoblotting
4.10. Real-time PCR Analysis
4.11. Cell Viability
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kawakami, T.; Ando, T.; Kimura, M.; Wilson, B.S.; Kawakami, Y. Mast cells in atopic dermatitis. Curr. Opin. Immunol. 2009, 21, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Tsai, M. IgE and mast cells in allergic disease. Nat. Med. 2012, 18, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Dema, B.; Suzuki, R.; Rivera, J. Rethinking the role of immunoglobulin E and its high-affinity receptor: New insights into allergy and beyond. Int. Arch. Allergy Immunol. 2014, 164, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Galli, S.J.; Nakae, S.; Tsai, M. Mast cells in the development of adaptive immune responses. Nat. Immunol. 2005, 6, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Mukai, K.; Tsai, M.; Saito, H.; Galli, S.J. Mast cells as sources of cytokines, chemokines, and growth factors. Immunol. Rev. 2018, 282, 121–150. [Google Scholar] [CrossRef]
- Minatoguchi, S.; Ohno, Y.; Funaguchi, N.; Baila, B.; Nagashima, K.; Fujiwara, H. Effect of “Jabara” juice on symptoms and QOL in patients cedar pollionsis. Clin. Immunol. Allergol. 2008, 50, 360–364. [Google Scholar]
- Kubo, M.; Matsuda, H.; Tomohiro, N.; Harima, S. History and therapeutic evaluation of Citrus hassaku HORT. Tanaka. Yakushigaku Zasshi 2004, 39, 363–364. [Google Scholar]
- Funaguchi, N.; Ohno, Y.; La, B.L.; Asai, T.; Yuhgetsu, H.; Sawada, M.; Takemura, G.; Minatoguchi, S.; Fujiwara, T.; Fujiwara, H. Narirutin inhibits airway inflammation in an allergic mouse model. Clin. Exp. Pharmacol. Physiol. 2007, 34, 766–770. [Google Scholar] [CrossRef]
- Murata, K.; Takano, S.; Masuda, M.; Iinuma, M.; Matsuda, H. Anti-degranulating activity in rat basophil leukemia RBL-2H3 cells of flavanone glycosides and their aglycones in citrus fruits. J. Nat. Med. 2013, 67, 643–646. [Google Scholar] [CrossRef]
- Gonzalez-Espinosa, C.; Odom, S.; Olivera, A.; Hobson, J.P.; Martinez, M.E.; Oliveira-Dos-Santos, A.; Barra, L.; Spiegel, S.; Penninger, J.M.; Rivera, J. Preferential signaling and induction of allergy-promoting lymphokines upon weak stimulation of the high affinity IgE receptor on mast cells. J. Exp. Med. 2003, 197, 1453–1465. [Google Scholar] [CrossRef]
- Jeuring, H.J.; Kuppers, F.J. High performance liquid chromatography of furfural and hydroxymethylfurfural in spirits and honey. J. Assoc. Off. Anal. Chem. 1980, 63, 1215–1218. [Google Scholar] [CrossRef] [PubMed]
- Janzowski, C.; Glaab, V.; Samimi, E.; Schlatter, J.; Eisenbrand, G. 5-Hydroxymethylfurfural: Assessment of mutagenicity, DNA-damaging potential and reactivity towards cellular glutathione. Food Chem. Toxicol. 2000, 38, 801–809. [Google Scholar] [CrossRef]
- Antal, M.J., Jr.; Mok, W.S.; Richards, G.N. Mechanism of formation of 5-(hydroxymethyl)-2-furaldehyde from D-fructose an sucrose. Carbohydr. Res. 1990, 199, 91–109. [Google Scholar] [CrossRef]
- Lo Coco, F.; Novelli, V.; Valentini, C.; Ceccon, L. High-performance liquid chromatographic determination of 2-furaldehyde and 5-hydroxymethyl-2-furaldehyde in fruit juices. J. Chromatogr. Sci. 1997, 35, 578–583. [Google Scholar] [CrossRef][Green Version]
- Shapla, U.M.; Solayman, M.; Alam, N.; Khalil, M.I.; Gan, S.H. 5-Hydroxymethylfurfural (HMF) levels in honey and other food products: Effects on bees and human health. Chem. Cent. J. 2018, 12, 35. [Google Scholar] [CrossRef]
- Murkovic, M.; Pichler, N. Analysis of 5-hydroxymethylfurfual in coffee, dried fruits and urine. Mol. Nutr. Food Res. 2006, 50, 842–846. [Google Scholar] [CrossRef]
- Yamada, P.; Nemoto, M.; Shigemori, H.; Yokota, S.; Isoda, H. Isolation of 5-(hydroxymethyl)furfural from Lycium chinense and its inhibitory effect on the chemical mediator release by basophilic cells. Planta Med. 2011, 77, 434–440. [Google Scholar] [CrossRef]
- Alizadeh, M.; Khodaei, H.; Mesgari Abbasi, M.; Saleh-Ghadimi, S. Assessing the effect of 5-hydroxymethylfurfural on selected components of immune responses in mice immunised with ovalbumin. J. Sci. Food Agric. 2017, 97, 3979–3984. [Google Scholar] [CrossRef]
- Ziadlou, R.; Barbero, A.; Stoddart, M.J.; Wirth, M.; Li, Z.; Martin, I.; Wang, X.L.; Qin, L.; Alini, M.; Grad, S. Regulation of Inflammatory Response in Human Osteoarthritic Chondrocytes by Novel Herbal Small Molecules. Int. J. Mol. Sci. 2019, 20, 5745. [Google Scholar] [CrossRef]
- Kong, F.; Fan, C.; Yang, Y.; Lee, B.H.; Wei, K. 5-hydroxymethylfurfural-embedded poly (vinyl alcohol)/sodium alginate hybrid hydrogels accelerate wound healing. Int. J. Biol. Macromol. 2019, 138, 933–949. [Google Scholar] [CrossRef]
- Kong, F.; Lee, B.H.; Wei, K. 5-Hydroxymethylfurfural Mitigates Lipopolysaccharide-Stimulated Inflammation via Suppression of MAPK, NF-kappaB and mTOR Activation in RAW 264.7 Cells. Molecules 2019, 24, 275. [Google Scholar] [CrossRef] [PubMed]
- Baba, Y.; Nishida, K.; Fujii, Y.; Hirano, T.; Hikida, M.; Kurosaki, T. Essential function for the calcium sensor STIM1 in mast cell activation and anaphylactic responses. Nat. Immunol. 2008, 9, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.; Jou, S.T.; Chen, Y.; Hoffmeyer, A.; Wang, D. Phospholipase C gamma 2 is essential for specific functions of Fc epsilon R and Fc gamma R. J. Immunol. 2002, 169, 6743–6752. [Google Scholar] [CrossRef] [PubMed]
- Turner, H.; Kinet, J.P. Signalling through the high-affinity IgE receptor Fc epsilonRI. Nature 1999, 402 (Suppl. 6760), B24–B30. [Google Scholar] [CrossRef] [PubMed]
- Vig, M.; Kinet, J.P. Calcium signaling in immune cells. Nat. Immunol. 2009, 10, 21–27. [Google Scholar] [CrossRef]
- Wolkart, G.; Schrammel, A.; Koyani, C.N.; Scherubel, S.; Zorn-Pauly, K.; Malle, E.; Pelzmann, B.; Andra, M.; Ortner, A.; Mayer, B. Cardioprotective effects of 5-hydroxymethylfurfural mediated by inhibition of L-type Ca(2+) currents. Br. J. Pharmacol. 2017, 174, 3640–3653. [Google Scholar] [CrossRef]
- Gibbs, B.F.; Grabbe, J. Inhibitors of PI 3-kinase and MEK kinase differentially affect mediator secretion from immunologically activated human basophils. J. Leukoc. Biol. 1999, 65, 883–890. [Google Scholar] [CrossRef]
- Karin, M. Signal transduction from the cell surface to the nucleus through the phosphorylation of transcription factors. Curr. Opin. Cell Biol. 1994, 6, 415–424. [Google Scholar] [CrossRef]
- Davis, R.J. Transcriptional regulation by MAP kinases. Mol. Reprod. Dev. 1995, 42, 459–467. [Google Scholar] [CrossRef]
- Nishida, K.; Hasegawa, A.; Yamasaki, S.; Uchida, R.; Ohashi, W.; Kurashima, Y.; Kunisawa, J.; Kimura, S.; Iwanaga, T.; Watarai, H.; et al. Mast cells play role in wound healing through the ZnT2/GPR39/IL-6 axis. Sci. Rep. 2019, 9, 10842. [Google Scholar] [CrossRef]
- Uchida, R.; Egawa, T.; Fujita, Y.; Furuta, K.; Taguchi, H.; Tanaka, S.; Nishida, K. Identification of the minimal region of peptide derived from ADP-ribosylation factor1 (ARF1) that inhibits IgE-mediated mast cell activation. Mol. Immunol. 2019, 105, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Uchida, R.; Xiang, H.; Arai, H.; Kitamura, H.; Nishida, K. L-Type Calcium Channel-Mediated Zinc Wave Is Involved in the Regulation of IL-6 by Stimulating Non-IgE with LPS and IL-33 in Mast Cells and Dendritic Cells. Biol. Pharm. Bull. 2019, 42, 87–93. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uchida, R.; Kato, M.; Hattori, Y.; Kikuchi, H.; Watanabe, E.; Kobayashi, K.; Nishida, K. Identification of 5-Hydroxymethylfurfural (5-HMF) as an Active Component Citrus Jabara That Suppresses FcεRI-Mediated Mast Cell Activation. Int. J. Mol. Sci. 2020, 21, 2472. https://doi.org/10.3390/ijms21072472
Uchida R, Kato M, Hattori Y, Kikuchi H, Watanabe E, Kobayashi K, Nishida K. Identification of 5-Hydroxymethylfurfural (5-HMF) as an Active Component Citrus Jabara That Suppresses FcεRI-Mediated Mast Cell Activation. International Journal of Molecular Sciences. 2020; 21(7):2472. https://doi.org/10.3390/ijms21072472
Chicago/Turabian StyleUchida, Ryota, Michiko Kato, Yuka Hattori, Hiroko Kikuchi, Emi Watanabe, Katsuumi Kobayashi, and Keigo Nishida. 2020. "Identification of 5-Hydroxymethylfurfural (5-HMF) as an Active Component Citrus Jabara That Suppresses FcεRI-Mediated Mast Cell Activation" International Journal of Molecular Sciences 21, no. 7: 2472. https://doi.org/10.3390/ijms21072472
APA StyleUchida, R., Kato, M., Hattori, Y., Kikuchi, H., Watanabe, E., Kobayashi, K., & Nishida, K. (2020). Identification of 5-Hydroxymethylfurfural (5-HMF) as an Active Component Citrus Jabara That Suppresses FcεRI-Mediated Mast Cell Activation. International Journal of Molecular Sciences, 21(7), 2472. https://doi.org/10.3390/ijms21072472