The WRKY Transcription Factor GmWRKY12 Confers Drought and Salt Tolerance in Soybean



 and
and
Abstract
1. Introduction
2. Results
2.1. Identification of GmWRKYs Up-Regulated under Drought/Salt Treatment
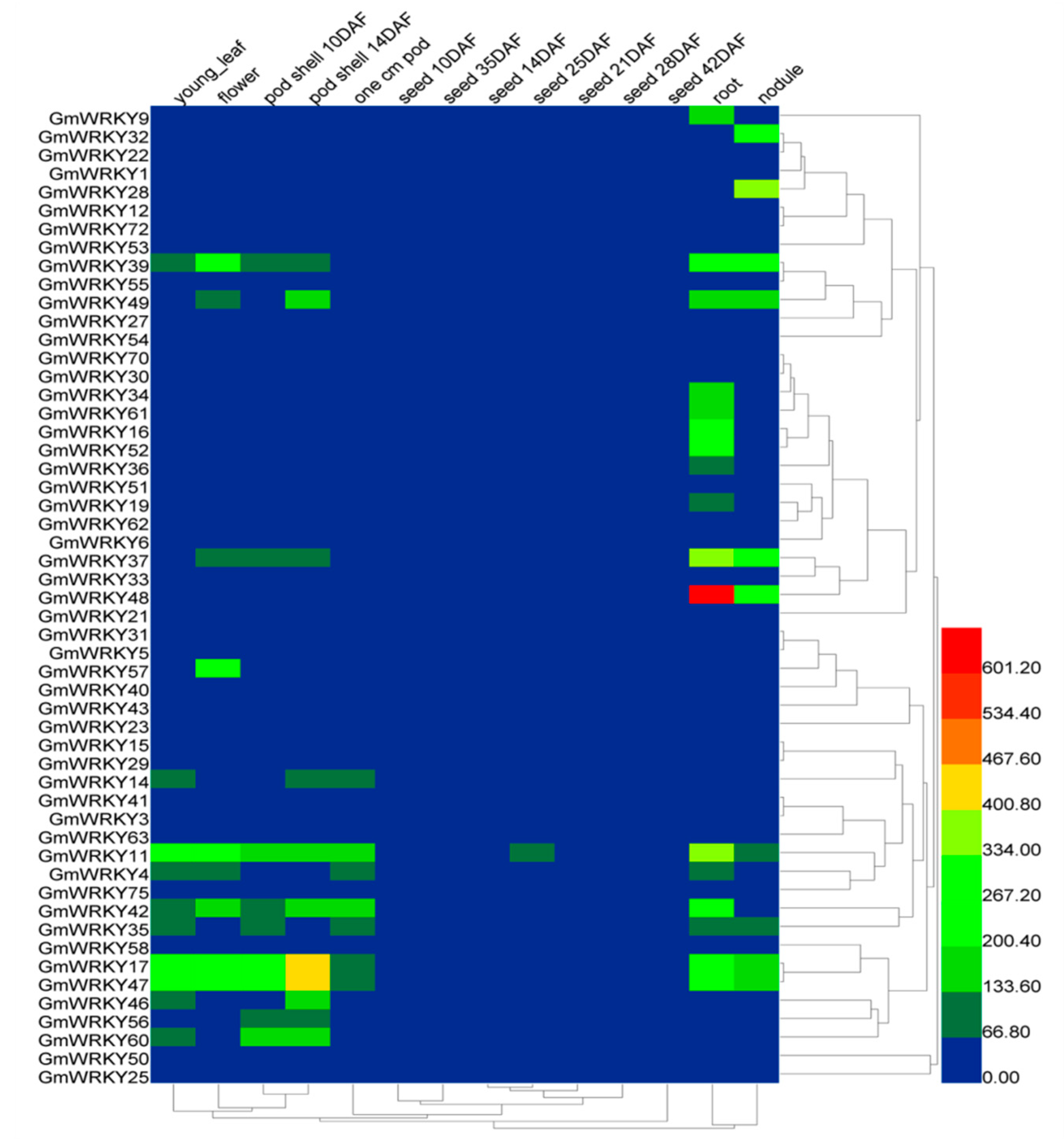
2.2. Tissue-Specific Expression Patterns of GmWRKYs
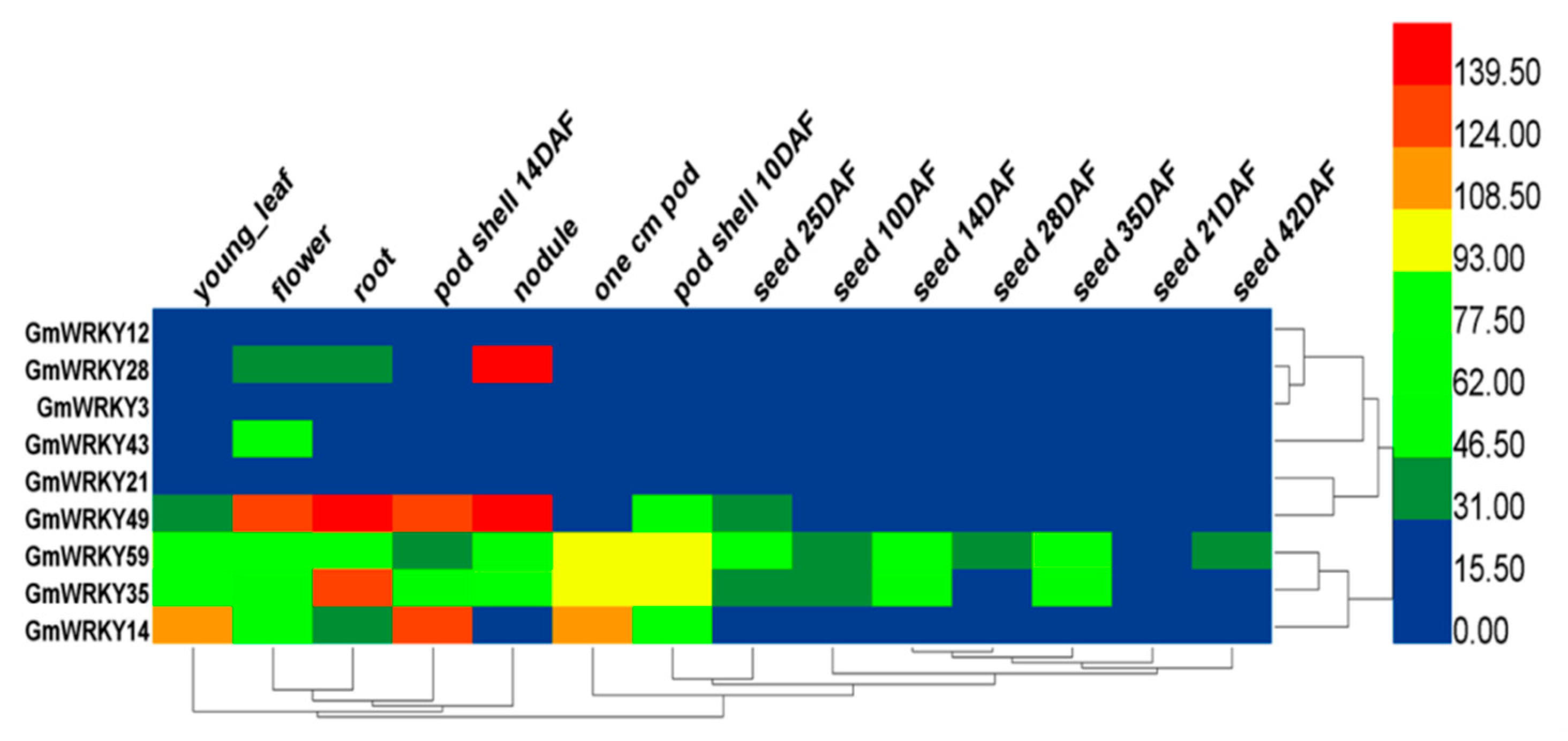
2.3. GmWRKYs Responsive to Both Drought and Salt Treatments
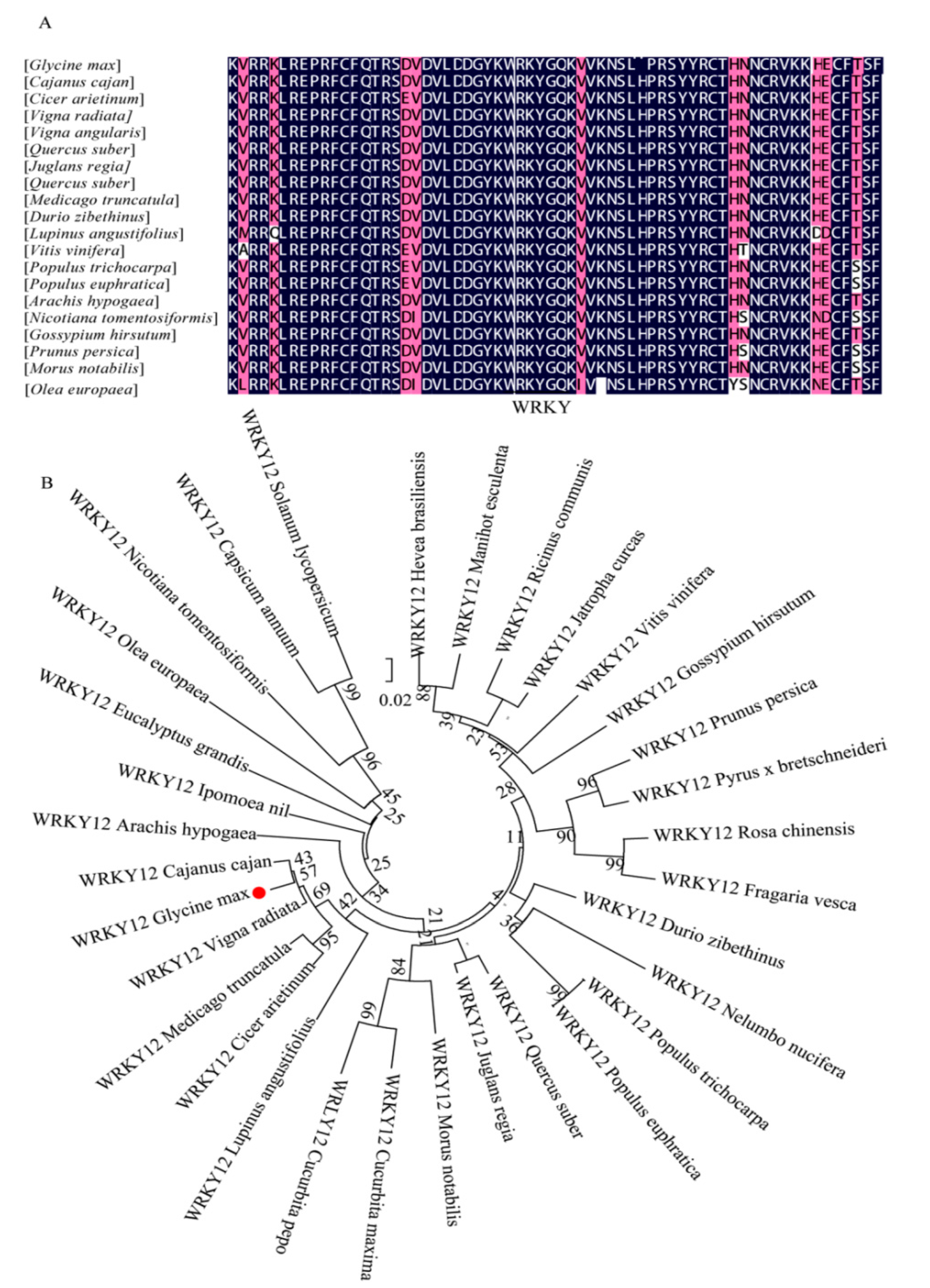
2.4. Multiple Sequence Alignment and Phylogenetic Analysis of GmWRKY12
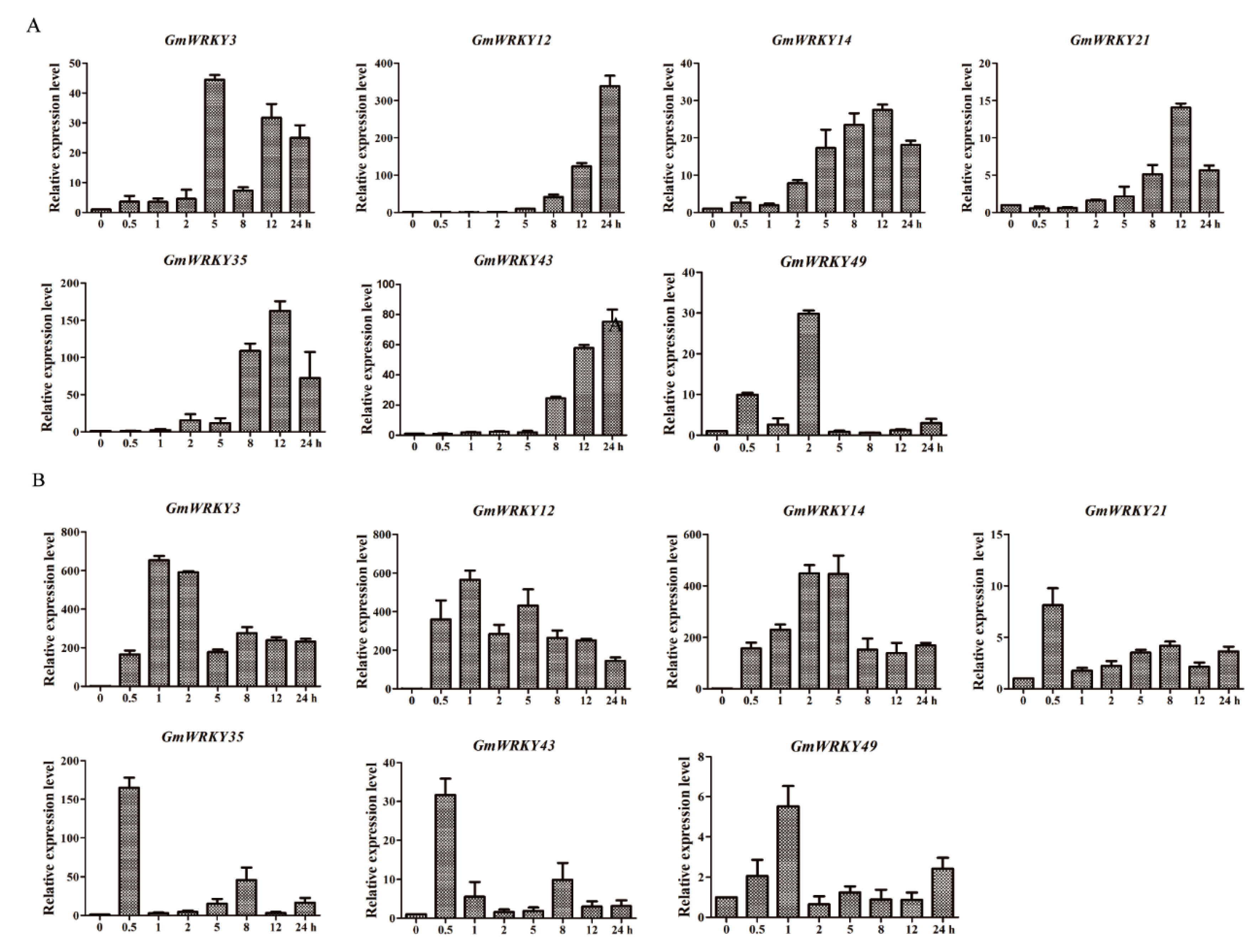
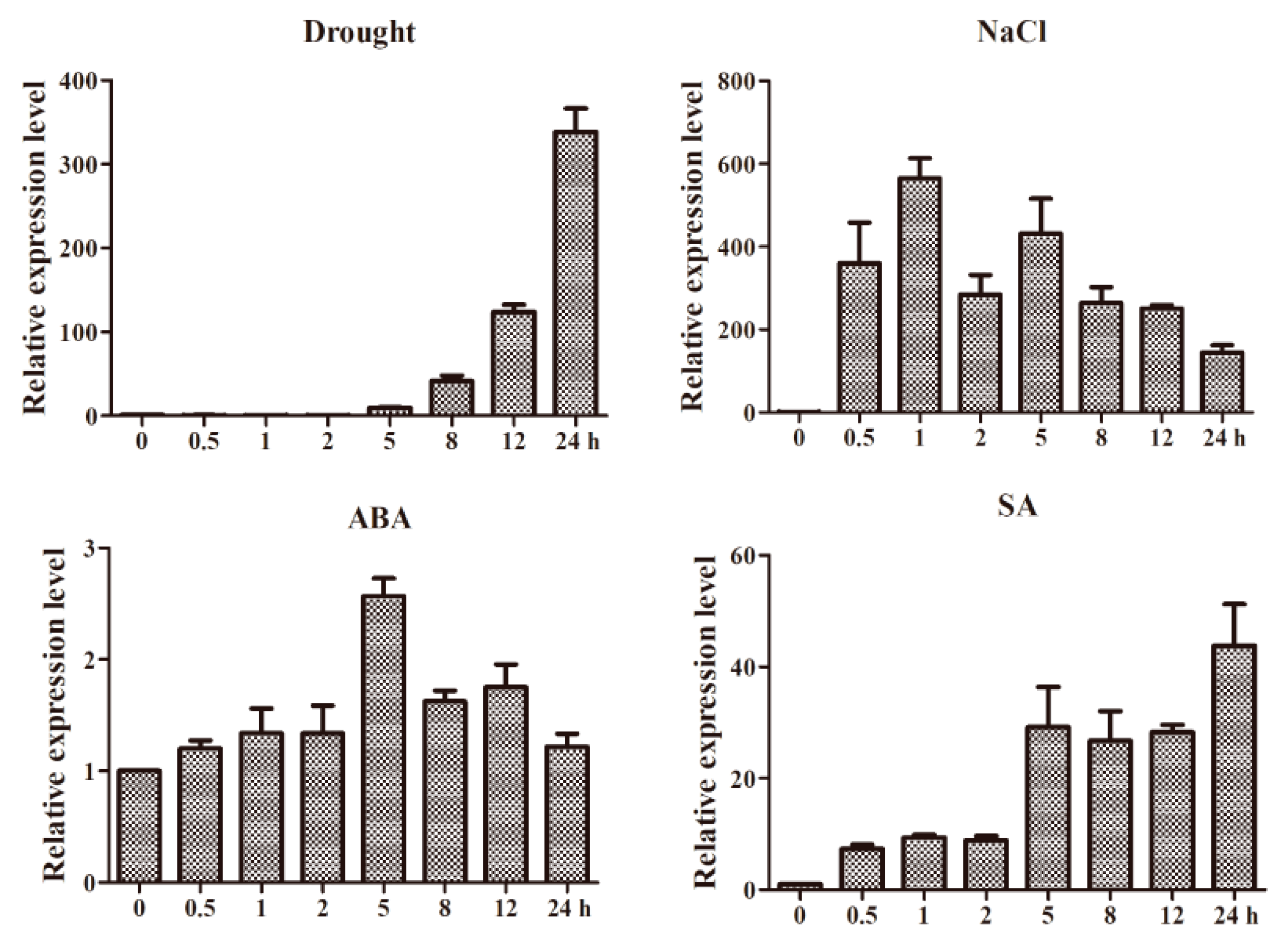
2.5. Expression Patterns of GmWRKY12 under Different Treatments
2.6. Cis-Acting Elements in Promoter
2.7. GmWRKY12 was Located in the Nucleus
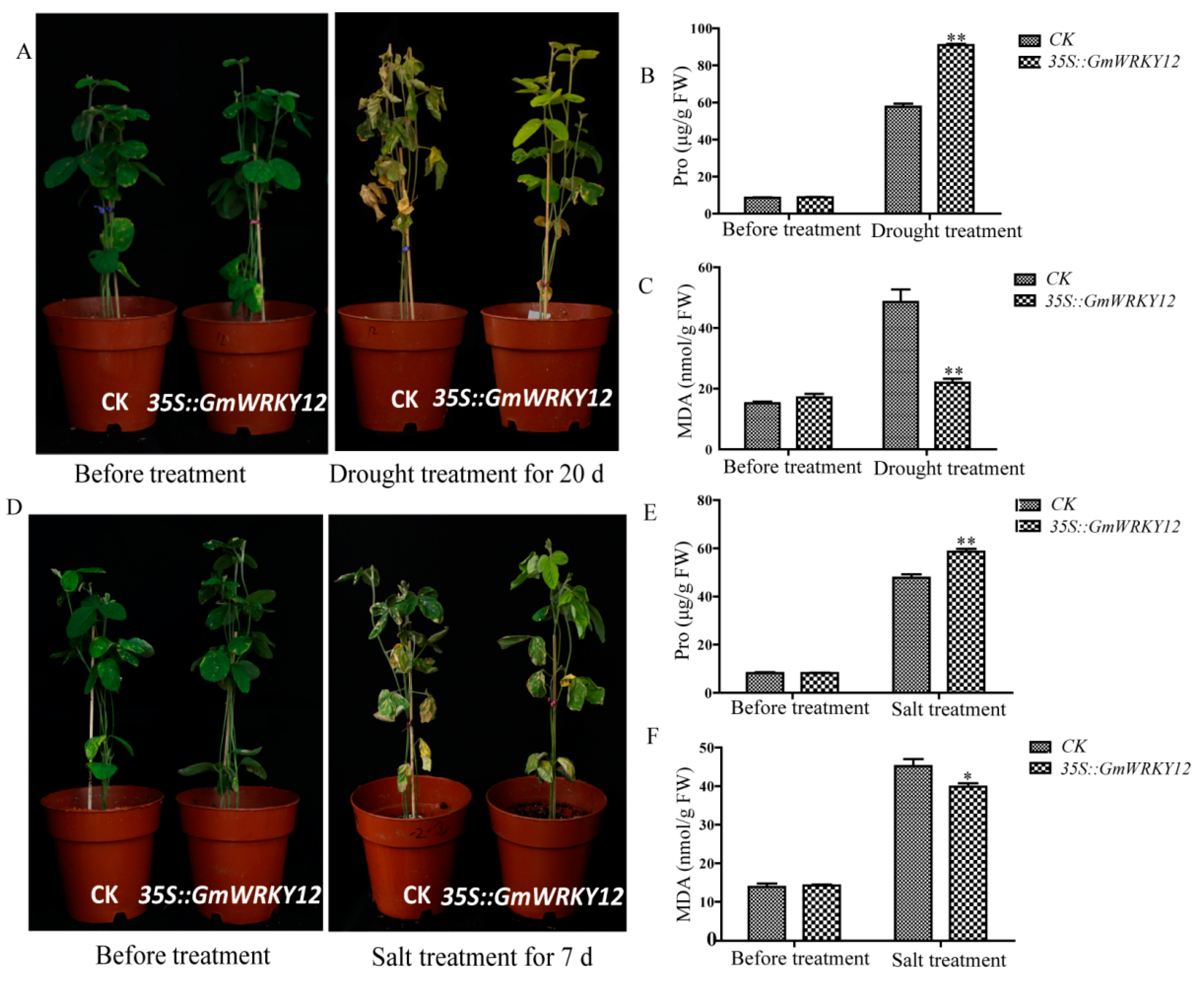
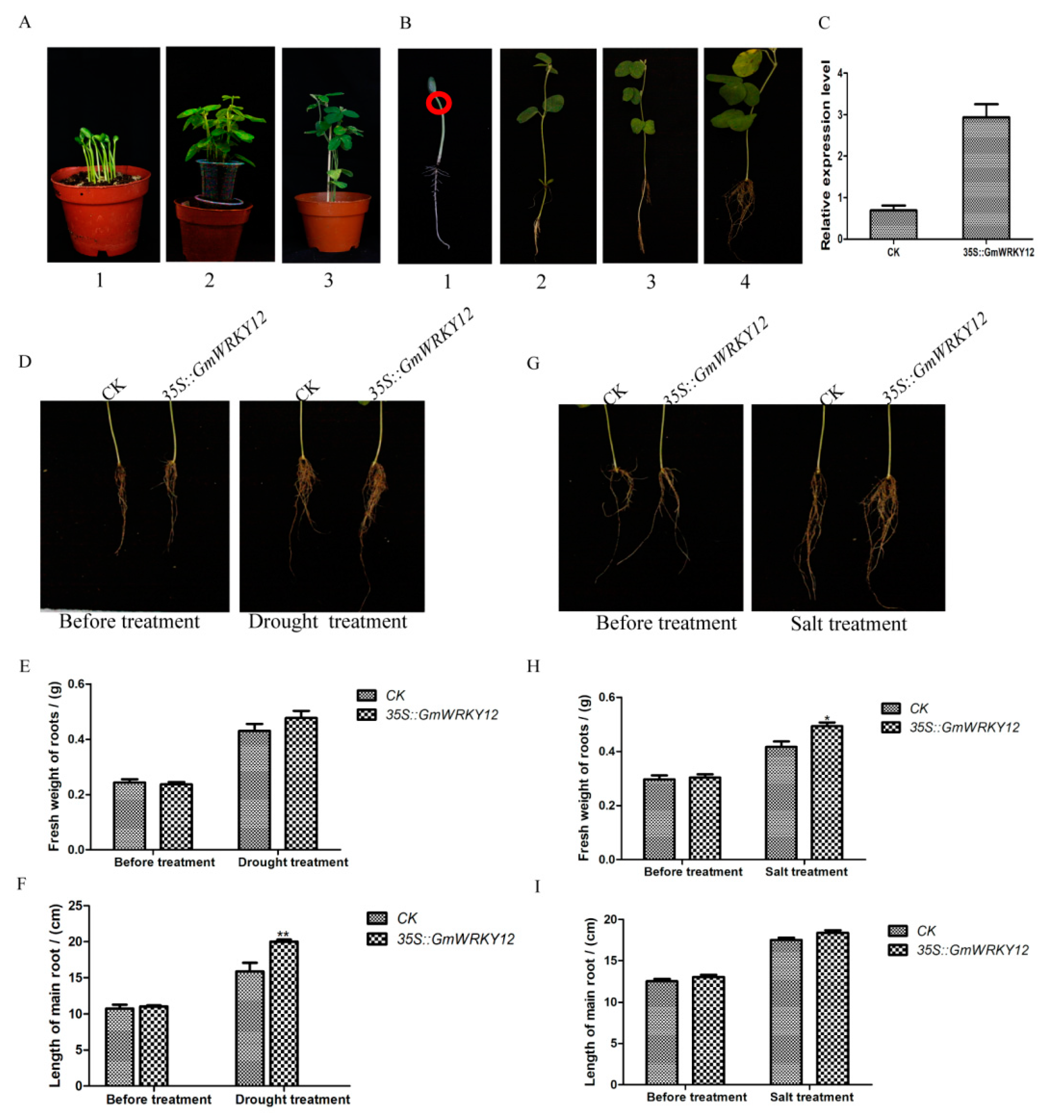
2.8. GmWRKY12 Improved Drought and Salt Tolerance of Soybean
3. Discussion
4. Materials and Methods
4.1. Identification and Annotation of GmWRKYs Response to Drought/Salt Stress
4.2. Tissue-Specific Expression Patterns of GmWRKYs
4.3. RNA Extraction and qRT-PCR
4.4. Gene Isolation and Phylogenetic Analysis of GmWRKY12
4.5. Co-Localization of GmWRKY
4.6. Cis-acting Elements in Promoter
4.7. A. rhizogenes-mediated Drought and Salt Stress Assays
4.8. Measurements of Proline and MDA Contents
4.9. Measurements of Fresh Weight and Main Length
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | abscisic acid |
| ABRE | ABA-responsive element |
| JA | jasmonic acid |
| SA | salicylic acid |
| MeJA | methyl jasmonate |
| qRT-PCR | quantitative real-time PCR |
| Pro | proline |
| MDA | malondialdehyde |
| DAF | days after flowering |
| GFP | green fluorescent protein |
References
- Peng, X.; Ma, X.; Fan, W.; Man, S.; Cheng, L.; Alam, I.; Lee, B.H.; Qi, D.; Shen, S.; Liu, G. Improved drought and salt tolerance of Arabidopsis thaliana by transgenic expression of a novel DREB gene from Leymus chinensis. Plant Cell Rep. 2011, 30, 1493–1502. [Google Scholar]
- Hong, C.; Cheng, D.; Zhang, G.; Zhu, D.; Chen, Y.; Tan, M. The role of ZmWRKY4 in regulating maize antioxidant defense under cadmium stress. Biochem. Bioph. Res. Commun. 2016, 482, 1504–1510. [Google Scholar] [CrossRef]
- Fu, J.; Liu, Q.; Wang, C.; Liang, J.; Liu, L.; Wang, Q. ZmWRKY79 positively regulates maize phytoalexin biosynthetic gene expression and is involved in stress response. J. Exp. Bot. 2017, 69, 497–510. [Google Scholar] [CrossRef]
- Liu, Q.L.; Xu, K.D.; Pan, Y.Z.; Jiang, B.B.; Liu, G.L.; Jia, Y.; Zhang, H.Q. Functional Analysis of a Novel Chrysanthemum WRKY Transcription Factor Gene Involved in Salt Tolerance. Plant Mol. Biol. Rep. 2014, 32, 282–289. [Google Scholar] [CrossRef]
- Zhu, J.K. Cell signaling under salt, water and cold stresses. Curr. Opin. Plant Biol. 2001, 4, 401–406. [Google Scholar] [CrossRef]
- Horie, T.; Hauser, F.; Schroeder, J.I. HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci. 2009, 14, 660–668. [Google Scholar] [CrossRef]
- Takahashi, R.; Liu, S.; Takano, T. Cloning and functional comparison of a high-affinity K+ transporter gene PhaHKT1 of salt-tolerant and salt-sensitive reed plants. J. Exp. Bot. 2007, 58, 4387–4395. [Google Scholar] [CrossRef] [PubMed]
- Guan, R.; Qu, Y.; Guo, Y.; Yu, L.; Liu, Y.; Jiang, J.; Chen, J.; Ren, Y.; Liu, G.; Tian, L. Salinity tolerance in soybean is modulated by natural variation in GmSALT3. Plant J. 2015, 80, 937–950. [Google Scholar] [CrossRef]
- Yokoi, S.; Quintero, F.J.; Cubero, B.; Ruiz, M.T.; Bressan, R.A.; Hasegawa, P.M.; Pardo, J.M. Differential expression and function of Arabidopsis thaliana NHX Na+/H+ antiporters in the salt stress response. Plant J. 2010, 30, 529–539. [Google Scholar] [CrossRef]
- Rodríguez-Rosales, M.P.; Gálvez, F.J.; Huertas, R.; Aranda, M.N.; Baghour, M.; Cagnac, O.; Venema, K. Plant NHX cation/proton antiporters. Plant Signal. Behav. 2009, 4, 265–276. [Google Scholar] [CrossRef]
- Shigaki, T.; Hirschi, K. Characterization of CAX-like genes in plants: Implications for functional diversity. Gene 2000, 257, 291–298. [Google Scholar] [CrossRef]
- Shigaki, T.; Hirschi, K.D. Diverse functions and molecular properties emerging for CAX cation/H+ exchangers in plants. Plant Biol. 2006, 8, 419–429. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Li, M.W.; Xie, M.; Liu, X.; Ni, M.; Shao, G.; Song, C.; Kay-Yuen, Y.A.; Tao, Y.; Wong, F.L. Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Yang, F.P.; Chen, X.Q.; Liang, R.Q.; Zhang, L.Q.; Geng, D.M.; Zhang, X.D.; Song, Y.Z.; Zhang, G.S. Induced expression of DREB transcriptional factor and study on its physiological effects of drought tolerance in transgenic wheat. Acta Genetica Sinica 2006, 33, 468–476. [Google Scholar] [CrossRef]
- Morran, S.; Eini, O.; Pyvovarenko, T.; Parent, B.; Singh, R.; Ismagul, A.; Eliby, S.; Shirley, N.; Langridge, P.; Lopato, S. Improvement of stress tolerance of wheat and barley by modulation of expression of DREB/CBF factors. Plant Biotechnol. J. 2011, 9, 230–249. [Google Scholar] [CrossRef] [PubMed]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Trends Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Rushton, P.J.; Robatzek, S.; Somssich, I.E. The WRKY superfamily of plant transcription factors. Trends Plant Sci. 2000, 5, 199–206. [Google Scholar] [CrossRef]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Jin, H.X.; Huang, F.; Cheng, H.; Song, H.N.; Yu, D.Y. Overexpression of the GmNAC2 Gene, an NAC Transcription Factor, Reduces Abiotic Stress Tolerance in Tobacco. Plant Mol. Biol. Rep. 2013, 31, 435–442. [Google Scholar] [CrossRef]
- Guerinot, M.L. The ZIP family of metal transporters. BBA Biomembr. 2000, 1465, 190–198. [Google Scholar] [CrossRef]
- Liao, Y.; Zou, H.F.; Wei, W.; Hao, Y.J.; Tian, A.G.; Huang, J.; Liu, Y.F.; Zhang, J.S.; Chen, S.Y. Soybean GmbZIP44, GmbZIP62 and GmbZIP78 genes function as negative regulator of ABA signaling and confer salt and freezing tolerance in transgenic Arabidopsis. Planta 2008, 228, 225–240. [Google Scholar] [CrossRef]
- Xu, Z.S.; Chen, M.; Ma, L.C.; Ma, Y.Z. Functions and application of the AP2/ERF franscription factor family in crop improvement. Bull Bot. 2011, 61, 570–585. [Google Scholar]
- Xu, Z.S.; Chen, M.; Li, L.C.; Ma, Y.Z. Functions of the ERF transcription factor family in plants. Botany 2008, 86, 969–977. [Google Scholar] [CrossRef]
- Fukuda, A.; Okada, Y.; Suzui, N.; Fujiwara, T.; Yoneyama, T.; Hayashi, H. Cloning and characterization of the gene for a phloem-specific glutathione S -transferase from rice leaves. Physiol. Plant. 2010, 120, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Hincha, D.K. LEA (Late Embryogenesis Abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genomics 2008, 9. [Google Scholar] [CrossRef]
- Yan, J.; Wang, B.; Jiang, Y.; Cheng, L.; Wu, T. GmFNSII-Controlled Soybean Flavone Metabolism Responds to Abiotic Stresses and Regulates Plant Salt Tolerance. Plant Cell Physiol. 2014, 55, 74–86. [Google Scholar] [CrossRef]
- Fei, C.; Yue, H.; Vannozzi, A.; Wu, K.; Cai, H.; Yuan, Q. The WRKY transcription factor family in model plants and crops. Crit. Rev. Plant Sci. 2018, 36, 1–25. [Google Scholar]
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY transcription factors. Plant Signal. Behav. 2010, 15, 247–258. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, N.; Hu, R.; Xiang, F. Genome-wide identification of soybean WRKY transcription factors in response to salt stress. Springerplus 2016, 5. [Google Scholar] [CrossRef]
- Lagacã, M.; Matton, D.P. Characterization of a WRKY transcription factor expressed in late torpedo-stage embryos of Solanum chacoense. Planta 2004, 219, 185–189. [Google Scholar] [CrossRef] [PubMed]
- Zentella, R.; Zhang, Z.; Park, M.; Thomas, S.; Endo, A.; Murase, K.; Fleet, C.; Jikumaru, Y.; Nambara, E.; Kamiya, Y. Global analysis of della direct targets in early gibberellin signaling in Arabidopsis. Plant Cell 2007, 19, 3037–3057. [Google Scholar] [CrossRef] [PubMed]
- Silke, R.; Imre, E.S. Targets of AtWRKY6 regulation during plant senescence and pathogen defense. Genes Dev. 2002, 16, 1139–1149. [Google Scholar]
- Johnson, C.S.; Kolevski, B.; Smyth, D.R. Transparent TESTA glabra2, a trichome and seed coat development gene of Arabidopsis, encodes a WRKY transcription factor. Plant Cell 2002, 14, 1359–1375. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.P.; Somssich, I.E. The role of WRKY transcription factors in plant immunity. Plant Physiol. 2009, 150, 1648–1655. [Google Scholar] [CrossRef] [PubMed]
- Birkenbihl, R.P.; Diezel, C.; Somssich, I.E. Arabidopsis WRKY33 is a key transcriptional regulator of hormonal and metabolic responses toward Botrytis cinerea infection. Plant Physiol. 2012, 159, 266–285. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Jiang, Y.; Ye, S.; Karim, A.; Ling, Z.; He, Y.; Yang, S.; Luo, K. PtrWRKY73, a salicylic acid-inducible poplar WRKY transcription factor, is involved in disease resistance in Arabidopsis thaliana. Plant Cell Rep. 2015, 34, 831–841. [Google Scholar] [CrossRef]
- Qiu, Y.P.; Yu, D.Q. Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis. Environ. Exp. Bot. 2009, 65, 35–47. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, L.; Ringler, P.; Smith, S.; Rushton, P.J.; Shen, Q.J. Three WRKY transcription factors additively repress abscisic acid and gibberellin signaling in aleurone cells. Int. J. Exp. Plant Biol. 2015, 236, 214–222. [Google Scholar] [CrossRef]
- Zou, X.; Seemann, J.R.; Neuman, D.; Shen, Q.J. A WRKY gene from creosote bush encodes an activator of the abscisic acid signaling pathway. J. Biol. Chem. 2004, 279, 55770–55779. [Google Scholar] [CrossRef]
- Shimono, M.; Sugano, S.; Nakayama, A.; Jiang, C.J.; Ono, K.; Toki, S.; Takatsuji, H. Rice WRKY45 plays a crucial role in benzothiadiazole-inducible blast resistance. Plant Cell 2007, 19, 2064–2076. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Song, Y.; Li, S.; Zhang, L.; Zou, C.; Yu, D. The role of WRKY transcription factors in plant abiotic stresses. BBA Gene Regul. Mech. 2012, 1819, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Nolan, T.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54 and WRKY70 transcription factors are involved in brassinosteroid-regulated plant growth and drought response. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef] [PubMed]
- Erpen, L.; Devi, H.S.; Grosser, J.W.; Dutt, M. Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants. Plant Cell Tissue Organ 2018, 132, 1–25. [Google Scholar] [CrossRef]
- Taji, T.; Ohsumi, C.; Iuchi, S.; Seki, M.; Kasuga, M.; Kobayashi, M.; YamaguchiShinozaki, K.; Shinozaki, K. Important roles of drought- and cold-inducible genes for galactinol synthase in stress tolerance in Arabidopsis thaliana. Plant J. 2010, 29, 417–426. [Google Scholar] [CrossRef]
- Wang, C.T.; Ru, J.N.; Liu, Y.W.; Li, M.; Zhao, D.; Yang, J.F.; Fu, J.D.; Xu, Z.S. Maize WRKY Transcription Factor ZmWRKY106 Confers Drought and Heat Tolerance in Transgenic Plants. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- He, G.H.; Xu, J.Y.; Wang, Y.X.; Liu, J.M.; Li, P.S.; Ming, C.; Ma, Y.Z.; Xu, Z.S. Drought-responsive WRKY transcription factor genes TaWRKY1 and TaWRKY33 from wheat confer drought and/or heat resistance in Arabidopsis. BMC Plant Biol. 2016, 16. [Google Scholar] [CrossRef]
- Wang, C.T.; Ru, J.N.; Liu, Y.W.; Yang, J.F.; Li, M.; Xu, Z.S.; Fu, J.D. The Maize WRKY transcription factor ZmWRKY40 confers drought resistance in transgenic Arabidopsis. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef]
- Ulker, B.; Somssich, I.E. WRKY transcription factors: From DNA binding towards biological function. Curr. Opin. Plant Biol. 2004, 7, 491–498. [Google Scholar] [CrossRef]
- Jiang, Y.; Qiu, Y.; Hu, Y.; Yu, D. Heterologous expression of AtWRKY57 confers drought tolerance in Oryza sativa. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Niu, C.F.; Wei, W.; Zhou, Q.Y.; Tian, A.G.; Hao, Y.J.; Zhang, W.K.; Ma, B.; Lin, Q.; Zhang, Z.B.; Zhang, J.S. Wheat WRKY genes TaWRKY2 and TaWRKY19 regulate abiotic stress tolerance in transgenic Arabidopsis plants. Plant Cell Environ. 2012, 35, 1156–1170. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shiroto, Y.; Kishitani, S.; Ito, Y.; Toriyama, K. Enhanced heat and drought tolerance in transgenic rice seedlings overexpressing OsWRKY11 under the control of HSP101 promoter. Plant Cell Rep. 2009, 28, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Cha, J.; Choi, C.; Choi, N.; Ji, H.S.; Park, S.R.; Lee, S.; Hwang, D.J. Rice WRKY11 plays a role in pathogen defense and drought tolerance. Rice 2018, 11. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Zhao, Y.; Wang, Y.; Lin, Y.; Peng, X.; Li, Q.; Chang, Y.; Jiang, H.; Xiang, Y.; Cheng, B. Overexpression of a maize WRKY58 gene enhances drought and salt tolerance in transgenic rice. Plant Cell Tissue Organ 2014, 119, 565–577. [Google Scholar] [CrossRef]
- Li, H.; Gao, Y.; Xu, H.; Dai, Y.; Deng, D.; Chen, J. ZmWRKY33, a WRKY maize transcription factor conferring enhanced salt stress tolerances in Arabidopsis. Plant Growth Regul. 2013, 70, 207–216. [Google Scholar] [CrossRef]
- Ullah, A.; Sun, H.; Hakim; Yang, X.; Zhang, X. A novel cotton WRKY-gene, GhWRKY6-like, improves salt tolerance by activating the ABA signalling pathway and scavenging of reactive oxygen species. Physiol. Plant 2017, 162, 439–454. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, P.; Hou, L.; Zhao, S.; Zhao, C.; Xia, H.; Li, P.; Zhang, Y.; Bian, X.; Wang, X. Global analysis of WRKY genes and their response to dehydration and salt stress in soybean. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Tian, A.G.; Zou, H.F.; Xie, Z.M.; Lei, G.; Huang, J.; Wang, C.M.; Wang, H.W.; Zhang, J.S.; Chen, S.Y. Soybean WRKY-type transcription factor genes, GmWRKY13, GmWRKY21 and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants. Plant Biotechnol. J. 2010, 6, 486–503. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Wang, N.; Guo, X.; Gao, Z. GhWRKY44, a WRKY transcription factor of cotton, mediates defense responses to pathogen infection in transgenic Nicotiana benthamiana. Plant Cell Tissue Organ 2015, 121, 127–140. [Google Scholar] [CrossRef]
- Liu, X.; Song, Y.; Xing, F.; Wang, N.; Wen, F.; Zhu, C. GhWRKY25, a group I WRKY gene from cotton, confers differential tolerance to abiotic and biotic stresses in transgenic Nicotiana benthamiana. Protoplasma 2015, 253, 1–17. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, L.; Chen, J.; Tao, L.; An, Y.; Cai, H. Overexpression of the alfalfa WRKY11 gene enhances salt tolerance in soybean. PLoS ONE 2018, 13, e0192382. [Google Scholar] [CrossRef]
- Mehdi, P.; Vijayaraj, N.; Youping, D. GeneVenn—A web application for comparing gene lists using Venn diagrams. Bioinformation 2007, 1, 420–422. [Google Scholar]
- Berri, S.; Abbruscato, P.; FaivreRampant, O.; Brasileiro, A.C.; Fumasoni, I.; Satoh, K.; Kikuchi, S.; Mizzi, L.; Morandini, P.; Pè, M.E. Characterization of WRKY co-regulatory networks in rice and Arabidopsis. BMC Plant Biol. 2009, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Guo, R.; Xu, X.; Gao, M.; Li, X.; Song, J.; Zheng, Y.; Wang, X. Evolution and expression analysis of the grape (Vitis vinifera L.) WRKY gene family. J. Exp. Bot. 2014, 65, 1513–1528. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z.; Cao, S.; Liu, Y. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol. Genet. Genomics 2012, 287, 495–513. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.J.; Barrett-Wilt, G.A.; Zhihua, H.; Vierstra, R.D. Proteomic analyses identify a diverse array of nuclear processes affected by small ubiquitin-like modifier conjugation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 107, 16512–16517. [Google Scholar] [CrossRef]
- Du, Y.T.; Zhao, M.J.; Wang, C.T.; Gao, Y.; Wang, Y.X.; Liu, Y.W.; Chen, M.; Chen, J.; Zhou, Y.B.; Xu, Z.S. Identification and characterization of GmMYB118 responses to drought and salt stress. BMC Plant Biol. 2018, 18. [Google Scholar] [CrossRef]
- Kereszt, A.; Li, D.; Indrasumunar, A.; Nguyen, C.D.; Nontachaiyapoom, S.; Kinkema, M.; Gresshoff, P.M. Agrobacterium rhizogenes-mediated transformation of soybean to study root biology. Nat. Protoc. 2007, 2, 948–952. [Google Scholar] [CrossRef]
- Cao, D.; Hou, W.S.; Song, S.K.; Sun, H.B.; Wu, C.X.; Gao, Y.S.; Han, T.F. Assessment of conditions affecting Agrobacterium rhizogenes-mediated transformation of soybean. Plant Cell Tissue Organ 2009, 96, 45–52. [Google Scholar] [CrossRef]
- Wang, F.; Chen, H.W.; Li, Q.T.; Wei, W.; Li, W.; Zhang, W.K.; Ma, B.; Bi, Y.D.; Lai, Y.C.; Liu, X.L. GmWRKY27 interacts with GmMYB174 to reduce expression of GmNAC29 for stress tolerance in soybean plants. Plant J. 2015, 83, 224–236. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, S.; Nakamura, K. Characterization of a cDNA encoding a novel DNA-binding protein, SPF1, that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and β-amylase from sweet potato. Mol. Genet. Genomics 1994, 244, 563–571. [Google Scholar] [CrossRef]
- Wei, K.F.; Chen, J.; Chen, Y.F.; Wu, L.J.; Xie, D.X. Molecular phylogenetic and expression analysis of the complete WRKY transcription factor family in Maize. DNA Res. 2012, 19, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.L. The WRKY family of transcription factors in rice and Arabidopsis and their origins. DNA Res. 2005, 12, 9–26. [Google Scholar] [CrossRef]
- Jue, D.; Sang, X.; Liu, L.; Shu, B.; Wang, Y.; Liu, C.; Xie, J.; Shi, S. Identification of WRKY Gene Family from Dimocarpus longan and its expression analysis during flower induction and abiotic stress responses. Int. J. Mol. Sci. 2018, 19. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Vo, K.T.X.; Cong, D.N.; Jeong, D.H.; Lee, S.K.; Kumar, M.; Kim, S.R.; Park, S.H.; Kim, J.K.; Jeon, J.S. Functional analysis of a cold-responsive rice WRKY gene, OsWRKY71. Plant Biotechnol. Rep. 2016, 10, 13–23. [Google Scholar] [CrossRef]
- Ding, M.; Chen, J.; Jiang, Y.; Lin, L.; Cao, Y.; Wang, M.; Zhang, Y.; Rong, J.; Ye, W. Genome-wide investigation and transcriptome analysis of the WRKY gene family in Gossypium. Mol. Genet. Genomics 2015, 290, 151–171. [Google Scholar] [CrossRef]
- Shi, J.N.; Li-Yun, L.I.; Wen-Jing, X.U.; Guan, M.L.; Xue-Jiao, L.I.; Niu, D.D.; Lan, J.P.; Dou, S.J.; Liu, L.J.; Liu, G.Z. Expression analysis of eight WRKY transcription factors in rice leaf growth and disease resistance response. Acta Phytopathol. Sin. 2014, 44, 54–64. [Google Scholar]
- Yu, Y.; Hu, R.; Wang, H.; Cao, Y.; He, G.; Fu, C.; Zhou, G. MlWRKY12, a novel Miscanthus transcription factor, participates in pith secondary cell wall formation and promotes flowering. Int. J. Exp. Plant Biol. 2013, 212, 1–9. [Google Scholar] [CrossRef]
- Li, W.; Wang, H.; Yu, D. Arabidopsis WRKY Transcription Factors WRKY12 and WRKY13 oppositely regulate flowering under short-day conditions. Mol. Plant 2016, 9, 1492–1503. [Google Scholar] [CrossRef]
- Li, X.J.; Guo, C.J.; Lu, W.J.; Duan, W.W.; Zhao, M.; Ma, C.Y.; Gu, J.T.; Xiao, K. expression pattern analysis of Zinc finger protein genes in wheat (Triticum aestivum L.) under phosphorus deprivation. J. Integr. Agric. 2014, 13, 1621–1633. [Google Scholar] [CrossRef]
- Van Verk, M.C.; Neeleman, L.; Bol, J.F.; Linthorst, H.J. Tobacco transcription factor NtWRKY12 interacts with TGA2.2 in vitro and in vivo. Front. Plant Sci. 2011, 2, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Thurow, C.; Schiermeyer, A.; Krawczyk, S.; Butterbrodt, T.; Nickolov, K.; Gatz, C. Tobacco bZIP transcription factor TGA2.2 and related factor TGA2.1 have distinct roles in plant defense responses and plant development. Plant J. 2010, 44, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Park, Y.H.; Nam, H.; Lee, Y.M.; Song, K.; Choi, C.; Ahn, I.; Park, S.R.; Lee, Y.H.; Hwang, D.J. Overexpression of the Brassica rapa transcription factor WRKY12 results in reduced soft rot symptoms caused by Pectobacterium carotovorum in Arabidopsis and Chinese cabbage. Plant Biol. 2015, 16, 973–981. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Yan, X.; Gao, X.; Zhang, D.M.; He, H.B.; Jia, G.X. Analysis of WRKY transcription factors and characterization of two Botrytis cinerea-responsive LrWRKY genes from Lilium regale. Plant Physiol. Biochem. 2018, 525–536. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Tang, Y.K.; Liu, Q.; Liang, C.Q.; Wang, Y.C. Cloning and expression analysis of THWRKY12 gene from Tamarix hispida. Chin. J. Agric. Biotechol. 2013, 5, 55–57. [Google Scholar]
- Baranwal, V.K.; Negi, N.; Khurana, P. Genome-wide identification and structural, functional and evolutionary analysis of WRKY components of Mulberry. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Tao, X.; Chen, C.; Li, C.; Liu, J.; Liu, C.; He, Y. Genome-wide investigation of WRKY gene family in pineapple: Evolution and expression profiles during development and stress. BMC Genomics 2018, 19. [Google Scholar] [CrossRef]
- Tan, C.K.; Carey, A.J.; Cui, X.; Webb, R.I.; Ipe, D.; Crowley, M.; Cripps, A.W.; Benjamin, B.W., Jr.; Ulett, K.B.; Schembri, M.A. Genome-wide mapping of cystitis due to Streptococcus agalactiae and Escherichia coli in mice identifies a unique bladder transcriptome that signifies pathogen-specific antimicrobial defense against urinary tract infection. Infect. Immun. 2012, 80, 3145–3160. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID a | Name b | Chr | CDS (bp) | Protein (aa) | Group c |
|---|---|---|---|---|---|
| GLYMA_14G103100 | GmWRKY40 | 14 | 849 | 282 | IIb |
| GLYMA_18G056600 | GmWRKY62 | 18 | 1689 | 542 | IIb |
| GLYMA_17G042300 | GmWRKY6 | 17 | 1173 | 390 | IIe |
| GLYMA_04G054200 | GmWRKY50 | 4 | 486 | 161 | IIe |
| GLYMA_01G222300 | GmWRKY22 | 1 | 738 | 245 | IIc |
| GLYMA_02G293400 | GmWRKY31 | 2 | 1278 | 425 | IIa |
| GLYMA_04G218700 | GmWRKY21 | 4 | 591 | 196 | I |
| GLYMA_06G147100 | GmWRKY51 | 6 | 591 | 196 | III |
| GLYMA_01G224800 | GmWRKY12 | 1 | 714 | 237 | IIc |
| GLYMA_11G163300 | GmWRKY19 | 11 | 1647 | 548 | I |
| GLYMA_06G061900 | GmWRKY17 | 6 | 885 | 294 | IIb |
| GLYMA_10G011300 | GmWRKY54 | 10 | 972 | 323 | IIa |
| GLYMA_04G223300 | GmWRKY58 | 4 | 954 | 317 | III |
| GLYMA_18G213200 | GmWRKY57 | 18 | 900 | 299 | III |
| GLYMA_06G125600 | GmWRKY53 | 6 | 1095 | 364 | IIa |
| GLYMA_19G217800 | GmWRKY23 | 19 | 873 | 290 | IId |
| GLYMA_09G280200 | GmWRKY33 | 9 | 1632 | 543 | I |
| GLYMA_03G002300 | GmWRKY70 | 3 | 747 | 248 | IIc |
| GLYMA_13G310100 | GmWRKY36 | 13 | 1845 | 614 | IIc |
| GLYMA_14G200200 | GmWRKY49 | 14 | 1728 | 575 | IIc |
| GLYMA_16G026400 | GmWRKY60 | 16 | 1122 | 373 | IIc |
| GLYMA_16G0544001 | GmWRKY75 | 16 | 588 | 195 | IIb |
| GLYMA_04G223200 | GmWRKY55 | 4 | 1020 | 339 | IId |
| GLYMA_02G232600 | GmWRKY39 | 2 | 1743 | 580 | III |
| GLYMA_05G0290001 | GmWRKY72 | 5 | 1785 | 594 | I |
| GLYMA_03G220100 | GmWRKY41 | 5 | 762 | 253 | IIe |
| GLYMA_08G021900 | GmWRKY46 | 8 | 1080 | 356 | III |
| GLYMA_15G003300 | GmWRKY27 | 15 | 921 | 306 | IIb |
| GLYMA_17G097900 | GmWRKY61 | 17 | 1803 | 600 | IIc |
| GLYMA_01G128100 | GmWRKY5 | 1 | 1527 | 508 | IId |
| GLYMA_12G212300 | GmWRKY16 | 12 | 792 | 263 | IIc |
| GLYMA_08G082400 | GmWRKY28 | 8 | 881 | 293 | III |
| GLYMA_07G227200 | GmWRKY3 | 7 | 1602 | 533 | IIc |
| GLYMA_03G256700 | GmWRKY43 | 66 | 1089 | 362 | IIe |
| GLYMA_15G168200 | GmWRKY42 | 15 | 882 | 293 | IIb |
| GLYMA_13G289400 | GmWRKY52 | 13 | 798 | 265 | IIc |
| GLYMA_08G011300 | GmWRKY25 | 8 | 444 | 147 | IId |
| GLYMA_09G061900 | GmWRKY47 | 19 | 1573 | 296 | IIc |
| GLYMA_17G222300 | GmWRKY30 | 4 | 555 | 184 | IIa |
| GLYMA_01G053800 | GmWRKY9 | 1 | 1368 | 455 | IIc |
| GLYMA_08G118200 | GmWRKY48 | 7 | 789 | 262 | IIc |
| GLYMA_01G056800 | GmWRKY32 | 1 | 894 | 297 | IId |
| GLYMA_08G218600 | GmWRKY56 | 8 | 942 | 313 | III |
| GLYMA_07G262700 | GmWRKY34 | 7 | 1554 | 517 | IIb |
| GLYMA_03G159700 | GmWRKY15 | 1 | 1017 | 338 | I |
| GLYMA_11G053100 | GmWRKY14 | 11 | 963 | 320 | I |
| GLYMA_05G096500 | GmWRKY11 | 17 | 1050 | 334 | I |
| GLYMA_17G222500 | GmWRKY63 | 17 | 849 | 278 | IIa |
| GLYMA_08G240800 | GmWRKY4 | 2 | 1572 | 523 | I |
| GLYMA_03G176600 | GmWRKY29 | 5 | 1308 | 436 | IIc |
| GLYMA_08G325800 | GmWRKY35 | 8 | 1734 | 577 | IIc |
| GLYMA_10G138300 | GmWRKY1 | 14 | 1449 | 482 | IIb |
| GLYMA_06G077400 | GmWRKY37 | 6 | 903 | 300 | III |
| Gene ID a | Name b | Chr | CDS (pb) | Protein (aa) | Group c |
|---|---|---|---|---|---|
| GLYMA_11G053100 | GmWRKY14 | 9 | 963 | 320 | I |
| GLYMA_08G325800 | GmWRKY35 | 8 | 1734 | 577 | IIc |
| GLYMA_04G218700 | GmWRKY21 | 10 | 591 | 196 | I |
| GLYMA_14G200200 | GmWRKY49 | 18 | 1728 | 575 | IIc |
| GLYMA_07G227200 | GmWRKY3 | 18 | 1602 | 533 | IIc |
| GLYMA_02G115200 | GmWRKY28 | 8 | 881 | 293 | III |
| GLYMA_03G256700 | GmWRKY43 | 16 | 1089 | 362 | III |
| GLYMA_06G320700 | GmWRKY59 | 6 | 2331 | 776 | IIc |
| GLYMA_01G224800 | GmWRKY12 | 7 | 714 | 237 | IIc |
| Cis-Elements | Numbers | Target Sequences | Functions |
|---|---|---|---|
| MYC | 32 | CANNTG | ABA and wound responsive element |
| W-box | 21 | TTGAC/TTTGACY/TGACY | SA responsive element |
| ABER4 | 18 | ACGT | ABA responsive element |
| MYB | 14 | C/TAACNA/G | Drought responsive element |
| CCAATB | 10 | CCAAT | Heat-responsive element |
| GT-1 | 7 | GAAAAA | Salt stress responsive element |
| DPBF | 6 | ACACNNG | Dehydration-responsive element |
| GARE | 2 | TAACAAR | GA-responsive element |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, W.-Y.; Du, Y.-T.; Ma, J.; Min, D.-H.; Jin, L.-G.; Chen, J.; Chen, M.; Zhou, Y.-B.; Ma, Y.-Z.; Xu, Z.-S.; et al. The WRKY Transcription Factor GmWRKY12 Confers Drought and Salt Tolerance in Soybean. Int. J. Mol. Sci. 2018, 19, 4087. https://doi.org/10.3390/ijms19124087
Shi W-Y, Du Y-T, Ma J, Min D-H, Jin L-G, Chen J, Chen M, Zhou Y-B, Ma Y-Z, Xu Z-S, et al. The WRKY Transcription Factor GmWRKY12 Confers Drought and Salt Tolerance in Soybean. International Journal of Molecular Sciences. 2018; 19(12):4087. https://doi.org/10.3390/ijms19124087
Chicago/Turabian StyleShi, Wen-Yan, Yong-Tao Du, Jian Ma, Dong-Hong Min, Long-Guo Jin, Jun Chen, Ming Chen, Yong-Bin Zhou, You-Zhi Ma, Zhao-Shi Xu, and et al. 2018. "The WRKY Transcription Factor GmWRKY12 Confers Drought and Salt Tolerance in Soybean" International Journal of Molecular Sciences 19, no. 12: 4087. https://doi.org/10.3390/ijms19124087
APA StyleShi, W.-Y., Du, Y.-T., Ma, J., Min, D.-H., Jin, L.-G., Chen, J., Chen, M., Zhou, Y.-B., Ma, Y.-Z., Xu, Z.-S., & Zhang, X.-H. (2018). The WRKY Transcription Factor GmWRKY12 Confers Drought and Salt Tolerance in Soybean. International Journal of Molecular Sciences, 19(12), 4087. https://doi.org/10.3390/ijms19124087