
Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects

 ,
, 
_Cherfan.png)
 ,
,  ,
, 
 ,
, 
 ,
,  and
and 
Abstract
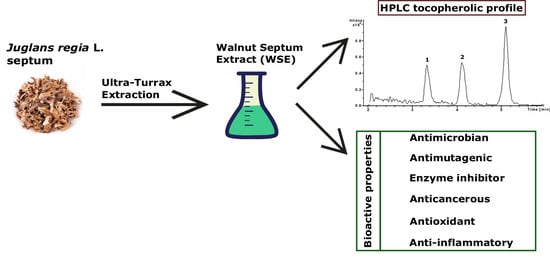
1. Introduction
2. Results and Discussion
2.1. Bioactive Compounds Present in Walnut Septum
2.1.1. Tocopherols
2.1.2. Overview on Phenolic Compounds Identified in WSE and Their Biological Properties
2.2. Biological Activities
2.2.1. Enzyme Inhibitory Activity
Cholinesterase Inhibitory Activity
α-Glucosidase Inhibitory Activity
Lipase Inhibitory Activity
2.2.2. Antibacterial and Antifungal Activity
2.2.3. Antimutagenic Assay
2.2.4. Cytocompatibility
2.2.5. Antioxidant Potential
2.2.6. Anti-Inflammatory Potential
3. Materials and Methods
3.1. Reagents
3.2. Plant Matrices
3.3. Tocopherol Quantification by LC-MS/MS
3.3.1. Chromatographic Analysis
3.3.2. Sample Preparation
3.4. Enzyme Inhibitory Activity
3.4.1. Cholinesterase Inhibition Assay
3.4.2. α-Glucosidase Inhibition Assay
3.4.3. Lipase Inhibition Assay
3.5. Antibacterial Activity
3.5.1. Microorganisms and Culture Conditions
3.5.2. Microdilution Method
3.6. Antifungal Activity
3.7. Antimutagenic Assay
3.8. Biological Activities on Cell Lines
3.8.1. Cell Cultures
3.8.2. Preparation of Extract Solutions
3.8.3. Cytocompatibility
3.8.4. Apoptosis Evaluation Using Flow Cytometry
3.8.5. Antioxidant Potential
3.8.6. Anti-Inflammatory Potential
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Vodnar, D.C.; Călinoiu, L.F.; Dulf, F.V.; Ştefănescu, E.; Crişan, G.; Socaciu, C. Identification of the bioactive compounds and antioxidant, antimutagenic and antimicrobial activities of thermally processed agro-industrial waste. Food Chem. 2017, 231, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Rusu, M.E.; Mocan, A.; Ferreira, I.C.F.R.; Popa, D.-S. Health Benefits of Nut Consumption in Middle-Aged and Elderly Population. Antioxidants (Basel) 2019, 8, 302. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Barros, L.; Calhelha, R.C.; Dueñas, M.; Carvalho, A.M.; Santos-Buelga, C.; Ferreira, I.C.F.R. Leaves and decoction of Juglans regia L.: Different performances regarding bioactive compounds and in vitro antioxidant and antitumor effects. Ind Crop. Prod. 2013, 51, 430–436. [Google Scholar] [CrossRef]
- Rusu, M.E.; Gheldiu, A.-M.; Mocan, A.; Moldovan, C.; Popa, D.-S.; Tomuta, I.; Vlase, L. Process Optimization for Improved Phenolic Compounds Recovery from Walnut (Juglans regia L.) Septum: Phytochemical Profile and Biological Activities. Molecules 2018, 23, 2814. [Google Scholar] [CrossRef]
- Martucci, M.; Ostan, R.; Biondi, F.; Bellavista, E.; Fabbri, C.; Bertarelli, C.; Salvioli, S.; Capri, M.; Franceschi, C.; Santoro, A. Mediterranean diet and inflammaging within the hormesis paradigm. Nutr. Rev. 2017, 75, 442–455. [Google Scholar] [CrossRef]
- Huang, J.; Weinstein, S.J.; Yu, K.; Männistö, S.; Albanes, D. Relationship Between Serum Alpha-Tocopherol and Overall and Cause-Specific Mortality. Circ Res. 2019, 125, 29–40. [Google Scholar] [CrossRef]
- Rusu, M.E.; Simedrea, R.; Gheldiu, A.-M.; Mocan, A.; Vlase, L.; Popa, D.-S.; Ferreira, I.C.F.R. Benefits of tree nut consumption on aging and age-related diseases: Mechanisms of actions. Trends Food Sci. Technol. 2019, 88, 104–120. [Google Scholar] [CrossRef]
- Galmés, S.; Serra, F.; Palou, A. Vitamin E metabolic effects and genetic variants: A challenge for precision nutrition in obesity and associated disturbances. Nutrients 2018, 10, 1919. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the substantiation of health claims related to vitamin E and protection of DNA, proteins and lipids from oxidative damage, maintenance of the normal function of the immune system. EFSA J. 2010, 8, 1816. [Google Scholar] [CrossRef]
- Kornsteiner, M.; Wagner, K.H.; Elmadfa, I. Tocopherols and total phenolics in 10 different nut types. Food Chem. 2006, 98, 381–387. [Google Scholar] [CrossRef]
- Amaral, J.S.; Alves, M.R.; Seabra, R.M.; Oliveira, B.P. Vitamin E composition of walnuts (Juglans regia L.): A 3-year comparative study of different cultivars. J. Agric. Food Chem. 2005, 53, 5467–5472. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Tsao, R.; Yang, R.; Kramer, J.; Hernandez, M. Fatty acid profiles, tocopherol contents, and antioxidant activities of heartnut (Juglans ailanthifolia var. cordiformis) and Persian walnut (Juglans regia L.). J. Agric. Food Chem. 2007, 55, 1164–1169. [Google Scholar] [CrossRef] [PubMed]
- Pycia, K.; Kapusta, I.; Jaworska, G. Impact of the Degree of Maturity of Walnuts (Juglans regia L.) and Their Variety on the Antioxidant Potential and the Content of Tocopherols and Polyphenols. Molecules 2019, 24, 2936. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Li, L.; Song, L.; Sun, X.; Yan, S.; Huang, W. Characterisation of phenolics in fruit septum of Juglans regia Linn. by ultra performance liquid chromatography coupled with Orbitrap mass spectrometer. Food Chem. 2019, 286, 669–677. [Google Scholar] [CrossRef]
- Pereira, J.A.; Oliveira, I.; Sousa, A.; Valentão, P.; Andrade, P.B.; Ferreira, I.C.F.R.; Ferreres, F.; Bento, A.; Seabra, R.; Estevinho, L. Walnut (Juglans regia L.) leaves: Phenolic compounds, antibacterial activity and antioxidant potential of different cultivars. Food Che Toxicol. 2007, 45, 2287–2295. [Google Scholar] [CrossRef]
- Cosmulescu, S.; Trandafir, I.; Violeta, N.; Ionica, M.; Tutulescu, F. Phenolics content, antioxidant activity and color of green walnut extracts for preparing walnut liquor. Not. Bot. Horti. Agrobot. Cluj-Napoca 2014, 42, 551–555. [Google Scholar] [CrossRef]
- Slatnar, A.; Mikulic-Petkovsek, M.; Stampar, F.; Veberic, R.; Solar, A. Identification and quantification of phenolic compounds in kernels, oil and bagasse pellets of common walnut (Juglans regia L.). Food Res. Int. 2015, 67, 255–263. [Google Scholar] [CrossRef]
- Fernández-Agulló, A.; Castro-Iglesias, A.; Freire, M.S.; González-Álvarez, J. Optimization of the Extraction of Bioactive Compounds from Walnut (Juglans major 209 x Juglans regia) Leaves: Antioxidant Capacity and Phenolic Profile. Antioxidants 2020, 9, 18. [Google Scholar] [CrossRef]
- Mollica, A.; Zengin, G.; Stefanucci, A.; Ferrante, C.; Menghini, L.; Orlando, G.; Brunetti, L.; Locatelli, M.; Dimmito, M.; Novellino, E.; et al. Nutraceutical potential of Corylus avellana daily supplements for obesity and related dysmetabolism. J. Funct. Foods 2018, 47, 562–574. [Google Scholar] [CrossRef]
- Mandalari, G.; Bisignano, C.; Filocamo, A.; Chessa, S.; Sarò, M.; Torre, G.; Faulks, R.M.; Dugo, P. Bioaccessibility of pistachio polyphenols, xanthophylls, and tocopherols during simulated human digestion. Nutrition 2013, 29, 338–344. [Google Scholar] [CrossRef]
- Bottone, A.; Cerulli, A.; D’Urso, G.; Masullo, M.; Montoro, P.; Napolitano, A.; Piacente, S. Plant Specialized Metabolites in Hazelnut (Corylus avellana) Kernel and Byproducts: An Update on Chemistry, Biological Activity, and Analytical Aspects. Planta Med. 2019, 85, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Kulišić-Bilušić, T.; Katalinić, V.; Dragović-Uzelac, V.; Ljubenkov, I.; Kriško, A.; Dejanović, B.; Jukić, M.; Politeo, O.; Pifat, G.; Miloš, M. Antioxidant and Acetylcholinesterase Inhibiting Activity of Several Aqueous Tea Infusions in vitro. Food Technol. Biotechnol. 2008, 46, 368–375. [Google Scholar]
- Orhan, I.E.; Suntar, I.P.; Akkol, E.K. In vitro neuroprotective effects of the leaf and fruit extracts of Juglans regia L. (walnut) through enzymes linked to Alzheimer’s disease and antioxidant activity. Int. J. Food Sci. Nutr. 2011, 62, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Mocan, A.; Diuzheva, A.; Carradori, S.; Andruch, V.; Massafra, C.; Moldovan, C.; Sisea, C.; Petzer, J.P.; Petzer, A.; Zara, S.; et al. Development of novel techniques to extract phenolic compounds from Romanian cultivars of Prunus domestica L. and their biological properties. Food Chem. Toxicol. 2018, 119, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Tsujita, T.; Shintani, T.; Sato, H. α-Amylase inhibitory activity from nut seed skin polyphenols. 1. Purification and characterization of almond seed skin polyphenols. J. Agric. Food Chem. 2013, 61, 4570–4576. [Google Scholar] [CrossRef]
- Rusu, M.E.; Fizeșan, I.; Pop, A.; Gheldiu, A.-M.; Mocan, A.; Crișan, G.; Vlase, L.; Loghin, F.; Popa, D.-S.; Tomuta, I. Enhanced Recovery of Antioxidant Compounds from Hazelnut (Corylus avellana L.) Involucre Based on Extraction Optimization: Phytochemical Profile and Biological Activities. Antioxidants (Basel) 2019, 8, 460. [Google Scholar] [CrossRef]
- Dehghani, F.; Mashhoody, T.; Panjehshahin, M. Effect of aqueous extract of walnut septum on blood glucose and pancreatic structure in streptozotocin-induced diabetic mouse. Iran J. Pharmacol. Ther. 2012, 11, 10–14. [Google Scholar]
- Yun, J.W. Possible anti-obesity therapeutics from nature—A review. Phytochemistry 2010, 71, 1625–1641. [Google Scholar] [CrossRef]
- Shi, D.; Chen, C.; Zhao, S.; Ge, F.; Liu, D.; Song, H. Walnut Polyphenols Inhibit Pancreatic Lipase Activity in Vitro and Have Hypolipidemic Effect on High-Fat Diet-Induced Obese Mice. J. Food Nutr. Res. 2014, 2, 757–763. [Google Scholar] [CrossRef]
- Salvat, A.; Antonacci, L.; Fortunato, R.; Suarez, E.; Godoy, H. Antimicrobial activity in methanolic extracts of several plant species from northern Argentina. Phytomedicine 2004, 11, 230–234. [Google Scholar] [CrossRef]
- Oliveira, I.; Sousa, A.; Ferreira, I.C.F.R.; Bento, A.; Estevinho, L.; Pereira, J.A. Total phenols, antioxidant potential and antimicrobial activity of walnut (Juglans regia L.) green husks. Food Chem. Toxicol. 2008, 46, 2326–2331. [Google Scholar] [CrossRef] [PubMed]
- Mocan, A.; Zengin, G.; Uysal, A.; Gunes, E.; Mollica, A.; Degirmenci, N.S.; Alpsoy, L.; Aktumsek, A. Biological and chemical insights of Morina persica L.: A source of bioactive compounds with multifunctional properties. J. Func.t Foods. 2016, 25, 94–109. [Google Scholar] [CrossRef]
- Ikken, Y.; Morales, P.; Martínez, A.; Marín, M.; Haza, A.; Cambero, M. Antimutagenic effect of fruit and vegetable ethanolic extracts against N-nitrosamines evaluated by the Ames test. J. Agric. Food Chem. 1999, 47, 3257–3264. [Google Scholar] [CrossRef]
- Mocan, A.; Zengin, G.; Simirgiotis, M.; Schafberg, M.; Mollica, A.; Vodnar, D.C.; Crişan, G.; Rohn, S. Functional constituents of wild and cultivated Goji (L. barbarum L.) leaves: Phytochemical characterization, biological profile, and computational studies. J. Enzym. Inhib. Med. Chem. 2017, 32, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Hasan, T.; Beslin, L.G.; Shafi, G.; Al-Hazzani, A.; Alshatwi, A. Anti-proliferative effects of organic extracts from root bark of Juglans regia L. (RBJR) on MDA-MB-231 human breast cancer cells: Role of Bcl-2/Bax, caspases and Tp53. Asian Pac. J. Cancer Prev. 2011, 12, 525–530. [Google Scholar] [PubMed]
- Carvalho, M.; Ferreira, P.J.; Mendes, V.S.; Silva, R.; Pereira, J.A.; Jerónimo, C.; Silva, B.M. Human cancer cell antiproliferative and antioxidant activities of Juglans regia L. Food Chem. Toxicol. 2010, 48, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.J.; Teuber, S.S.; Gobeille, A.; Cremin, P.; Waterhouse, A.L.; Steinberg, F.M. Walnut polyphenolics inhibit in vitro human plasma and LDL oxidation. J. Nutr. 2001, 131, 2837–2842. [Google Scholar] [CrossRef]
- Jiang, Q. Natural Forms of Vitamin E as Effective Agents for Cancer Prevention and Therapy. Adv. Nutr. 2017, 8, 850–867. [Google Scholar] [CrossRef]
- Tucker, L.A. Alpha- and gamma-tocopherol and telomere length in 5768 US men and women: A NHANES study. Nutrients 2017, 9, 601. [Google Scholar] [CrossRef]
- Browne, D.; McGuinness, B.; Woodside, J.V.; McKay, G.J. Vitamin E and Alzheimer’s disease: What do we know so far? Clin. Interv. Aging 2019, 14, 1303–1317. [Google Scholar] [CrossRef]
- Muthaiyah, B.; Essa, M.M.; Chauhan, V.; Chauhan, A. Protective effects of walnut extract against amyloid beta peptide-induced cell death and oxidative stress in PC12 cells. Neurochem. Res. 2011, 36, 2096–2103. [Google Scholar] [CrossRef] [PubMed]
- Muzaffer, U.; Paul, V.; Prasad, N.R.; Karthikeyan, R.; Agilan, B. Protective effect of Juglans regia L. against ultraviolet B radiation induced inflammatory responses in human epidermal keratinocytes. Phytomedicine 2018, 42, 100–111. [Google Scholar] [CrossRef]
- Anjum, S.; Gani, A.; Ahmad, M.; Shah, A.; Masoodi, F.A.; Shah, Y.; Gani, A. Antioxidant and antiproliferative activity of walnut extract (Juglans regia L.) processed by different methods and identification of compounds using GC/MS and LC/MS technique. J. Food Process. Pres. 2017, 41, e12756. [Google Scholar] [CrossRef]
- Gupta, S.; Suh, N. Tocopherols in cancer: An update. Mol. Nutr. Food Res. 2016, 60, 1354–1363. [Google Scholar] [CrossRef] [PubMed]
- Nardini, M.; Finkelstein, E.I.; Reddy, S.; Valacchi, G.; Traber, M.; Cross, C.E.; Van der Vliet, A. Acrolein-induced cytotoxicity in cultured human bronchial epithelial cells. Modulation by alpha-tocopherol and ascorbic acid. Toxicology 2002, 170, 173–185. [Google Scholar] [CrossRef]
- Luan, S.; Muhayimana, S.; Xu, J.; Zhang, X.; Xiao, C.; Huang, Q. The effect of α-tocopherol and dithiothreitol in ameliorating emamectin benzoate cytotoxicity in human K562 cells involving the modulation of ROS accumulation and NF-κB signaling. Ecotoxicol. Env. Saf. 2019, 167, 114–121. [Google Scholar] [CrossRef]
- Zarogoulidis, P.; Cheva, A.; Zarampouka, K.; Huang, H.; Li, C.; Huang, Y.; Katsikogiannis, N.; Zarogoulidis, K. Tocopherols and tocotrienols as anticancer treatment for lung cancer: Future nutrition. J. Thorac. Dis. 2013, 5, 349–352. [Google Scholar] [CrossRef]
- Azzi, A. Many tocopherols, one vitamin E. Mol. Asp. Med. 2018, 61, 92–103. [Google Scholar] [CrossRef]
- Boots, A.W.; Haenen, G.R.; Bast, A. Health effects of quercetin: From antioxidant to nutraceutical. Eur. J. Pharmacol. 2008, 585, 325–337. [Google Scholar] [CrossRef]
- Shirai, M.; Yamanishi, R.; Moon, J.-H.; Murota, K.; Terao, J. Effect of quercetin and its conjugated metabolite on the hydrogen peroxide-induced intracellular production of reactive oxygen species in mouse fibroblasts. Biosci. Biotechnol. Biochem. 2002, 66, 1015–1021. [Google Scholar] [CrossRef]
- Choi, S.-J.; Tai, B.H.; Cuong, N.M.; Kim, Y.-H.; Jang, H.-D. Antioxidative and anti-inflammatory effect of quercetin and its glycosides isolated from mampat (Cratoxylum formosum). Food Sci. Biotechnol. 2012, 21, 587–595. [Google Scholar] [CrossRef]
- Tanigawa, S.; Fujii, M.; Hou, D.-X. Action of Nrf2 and Keap1 in ARE-mediated NQO1 expression by quercetin. Free Radic. Biol. Med. 2007, 42, 1690–1703. [Google Scholar] [CrossRef] [PubMed]
- Yao, P.; Nussler, A.; Liu, L.; Hao, L.; Song, F.; Schirmeier, A.; Nussler, N. Quercetin protects human hepatocytes from ethanol-derived oxidative stress by inducing heme oxygenase-1 via the MAPK/Nrf2 pathways. J. Hepatol. 2007, 47, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Han, X.; Piao, M.; Oh, M.; Fernando, P.; Kang, K.; Ryu, Y.; Jung, U.; Kim, I.; Hyun, J. Hyperoside Induces Endogenous Antioxidant System to Alleviate Oxidative Stress. J. Cancer Prev. 2016, 21, 41–47. [Google Scholar] [CrossRef]
- Wardyn, J.D.; Ponsford, A.H.; Sanderson, C.M. Dissecting molecular cross-talk between Nrf2 and NF-κB response pathways. Biochem. Soc. Trans. 2015, 43, 621–626. [Google Scholar] [CrossRef]
- Vieira, V.; Pereira, C.; Abreu, R.M.V.; Calhelha, R.C.; Alves, J.M.; Coutinho, J.A.P.; Ferreira, O.; Barros, L.; Ferreira, I.C.F.R. Hydroethanolic extract of Juglans regia L. green husks: A source of bioactive phytochemicals. Food Chem. Toxicol. 2020, 137, 111189. [Google Scholar] [CrossRef]
- Papoutsi, Z.; Kassi, E.; Chinou, I.; Halabalaki, M.; Skaltsounis, L.; Moutsatsou, P. Walnut extract (Juglans regia L.) and its component ellagic acid exhibit anti-inflammatory activity in human aorta endothelial cells and osteoblastic activity in the cell line KS483. Br. J. Nutr. 2008, 99, 715–722. [Google Scholar] [CrossRef]
- Poulose, S.; Bielinski, D.; Shukitt-Hale, B. Walnut diet reduces accumulation of polyubiquitinated proteins and inflammation in the brain of aged rats. J. Nutr. Biochem. 2013, 24, 912–919. [Google Scholar] [CrossRef]
- Eidi, A.; Moghadam, J.Z.; Mortazavi, P.; Rezazadeh, S.; Olamafar, S. Hepatoprotective effects of Juglans regia extract against CCl4-induced oxidative damage in rats. Pharm. Biol. 2013, 51, 558–565. [Google Scholar] [CrossRef]
- Qamar, W.; Sultana, S. Polyphenols from Juglans regia L.(walnut) kernel modulate cigarette smoke extract induced acute inflammation, oxidative stress and lung injury in Wistar rats. Hum. Exp. Toxicol. 2011, 30, 499–506. [Google Scholar] [CrossRef]
- Hosseinzadeh, H.; Zarei, H.; Taghiabadi, E. Antinociceptive, anti-inflammatory and acute toxicity effects of Juglans regia L. leaves in mice. Iran Red Crescent Med. J. 2011, 13, 27–33. [Google Scholar] [PubMed]
- Meng, Q.; Wang, Y.; Chen, F.; Xiao, T.; Zhang, L. Polysaccharides from diaphragma juglandis fructus: Extraction optimization, antitumor, and immune-enhancement effects. Int. J. Biol. Macromol. 2018, 115, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Coetzee, D.D.; López, V.; Smith, C. High-mesembrine Sceletium extract (TrimesemineTM) is a monoamine releasing agent, rather than only a selective serotonin reuptake inhibitor. J. Ethnopharmacol. 2016, 177, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Lee, Y.M.; Kim, H.; Kim, J.; Jang, D.S.; Kim, J.H.; Kim, J.S. Anti-obesity effect of Morus bombycis root extract: Anti-lipase activity and lipolytic effect. J. Ethnopharmacol. 2010, 130, 621–624. [Google Scholar] [CrossRef]
- Mortelmans, K.; Zeiger, E. The Ames Salmonella/microsome mutagenicity assay. Mutat. Res. 2000, 455, 29–60. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds are not available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | RT (min) | M | [M − H+] | m/z Ions |
|---|---|---|---|---|
| δ-tocopherol | 3.3 | 402.6 | 401.6 | 401→386 |
| γ-tocopherol | 4.1 | 416.7 | 415.7 | 415→400 |
| α-tocopherol | 5.1 | 430.7 | 429.7 | 429→163 |
| α-Tocopherol | γ/β-Tocopherols | δ-Tocopherol | Total-Tocopherols | |
|---|---|---|---|---|
| WSE | 3.35 ± 0.04 | 1.73 ± 0.01 | 1.47 ± 0.02 | 6.55 ± 0.07 |
| WE | 0.78 ± 0.01 | 16.72 ± 0.63 | 2.06 ± 0.11 | 19.56 ± 0.74 |
| WSE/WE | 4.295 | 0.103 | 0.714 |
| Peak No. | Phenolic Compound | RT (min) | m/z | Analysis (mg/100 g) |
|---|---|---|---|---|
| 1 | Gallic acid | 1.5 | 169.1 | 7.96 |
| 2 | Protocatechuic acid | 2.8 | 153.1 | 0.99 |
| 3 | Caftaric acid | 3.5 | 311.2 | ˂ LOD |
| 4 | Gentisic acid | 3.5 | 153.1 | ˂ LOQ |
| 5 | Caffeic acid | 5.6 | 179.1 | ˂ LOD |
| 6 | Chlorogenic acid | 5.6 | 353.3 | ˂ LOQ |
| 7 | Catechin | 6.0 | 289.2 | 59.76 |
| 8 | Vanillic acid | 6.7 | 167.1 | 0.56 |
| 9 | Syringic acid | 8.4 | 197.2 | 0.52 |
| 10 | Epicatechin | 9.0 | 289.2 | 1.25 |
| 11 | p-Coumaric acid | 9.5 | 163.0 | ˂ LOQ |
| 12 | Ferulic acid | 12.8 | 193.2 | ˂ LOQ |
| 13 | Sinapic acid | 15.0 | 223.2 | ˂ LOD |
| 14 | Hyperoside | 18.6 | 463.4 | 6.73 |
| 15 | Isoquercitrin | 19.6 | 463.1 | 10.36 |
| 16 | Rutoside | 20.2 | 609.5 | ˂ LOD |
| 17 | Myricetol | 21.1 | 317.2 | ˂ LOD |
| 18 | Fisetin | 22.9 | 285.2 | ˂ LOD |
| 19 | Quercitrin | 23.6 | 447.4 | 107.3 |
| 20 | Quercetin | 26.8 | 301.2 | ˂ LOD |
| 21 | Patuletin | 28.7 | 331.3 | ˂ LOD |
| 22 | Luteolin | 29.1 | 285.2 | ˂ LOD |
| 23 | Kaempferol | 32.5 | 285.2 | ˂ LOD |
| 24 | Apigenin | 33.1 | 269.2 | ˂ LOD |
| Samples | Staphylococcus aureus | Escherichia coli | Pseudomonas aeruginosa | Salmonella enteritidis | Candida albicans | Candida parapsilosis |
|---|---|---|---|---|---|---|
| WSE (mg/mL) | 0.098 | 1.56 | 0.012 | 0.098 | 3.12 | 3.12 |
| Gentamycin/ Fluconazole (μg/mL) | 0.038 | 1.2 | 1.2 | 2.4 | 0.1 | 0.1 |
| Test Item | Number of Revertants | |||
|---|---|---|---|---|
| TA 98 | TA 100 | |||
| Mean (±SD) | Inhibition (%) | Mean (±SD) | Inhibition (%) | |
| Negative Control | 9.25 ± 3.6 | - | 9.25 ± 2.4 | - |
| WSE | 124 ± 4.4 | 36.08 | 198 ± 6.3 | 43.27 |
| 4-NPD/NaN3 | 194 ± 3.3 | 0 | 349 ± 15.22 | 0 |
| Cell Lines | Exposure Duration (h) | |
|---|---|---|
| 24 h | 48 h | |
| A549 | 80.02 ± 4.33 | 70.79 ± 1.93 |
| T47D-KBluc | 265.60 ± 53.79 | 112.75 ± 6.38 |
| MCF-7 | >400 | >400 |
| HGF | >400 | 254.25 ± 3.56 |
| Cell Type | Dose (µg/mL) | Viable Cells (%) | Necrotic Cells (%) | Early Apoptotic (%) | Late Apoptotic (%) |
|---|---|---|---|---|---|
| A549 | NC | 98.05 ± 0.95 | 0.43 ± 0.15 | 0.93 ± 0.57 | 0.53 ± 0.41 |
| 100 | 58.23 ± 2.51 * | 30.13 ± 5.10 * | 6.1 ± 1.15 * | 5.66 ± 2.53 * | |
| 200 | 49.65 ± 3.63 * | 41.66 ± 7.80 * | 3.36 ± 0.55 * | 5.8 ± 4.07 * | |
| 400 | 28.96 ± 2.75 * | 69.83 ± 2.65 * | 0.03 ± 0.05 | 0.61 ± 0.26 | |
| T47D-KBluc | NC | 98.30 ± 0.43 | 0.50 ± 0.17 | 0.63 ± 0.15 | 0.53 ± 0.40 |
| 100 | 80.03 ± 5.12 * | 12.13 ± 8.35 * | 1.73 ± 1.79 | 6.03 ± 5.4 | |
| 200 | 42.73 ± 4.05 * | 56.53 ± 4.18 * | 0 | 0.40 ± 0.26 | |
| 400 | 12.45 ± 3.25 * | 84.85 ± 3.75 * | 0.1 ± 0.1 | 2.65 ± 1.45 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rusu, M.E.; Fizesan, I.; Pop, A.; Mocan, A.; Gheldiu, A.-M.; Babota, M.; Vodnar, D.C.; Jurj, A.; Berindan-Neagoe, I.; Vlase, L.; et al. Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects. Molecules 2020, 25, 2187. https://doi.org/10.3390/molecules25092187
Rusu ME, Fizesan I, Pop A, Mocan A, Gheldiu A-M, Babota M, Vodnar DC, Jurj A, Berindan-Neagoe I, Vlase L, et al. Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects. Molecules. 2020; 25(9):2187. https://doi.org/10.3390/molecules25092187
Chicago/Turabian StyleRusu, Marius Emil, Ionel Fizesan, Anca Pop, Andrei Mocan, Ana-Maria Gheldiu, Mihai Babota, Dan Cristian Vodnar, Ancuta Jurj, Ioana Berindan-Neagoe, Laurian Vlase, and et al. 2020. "Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects" Molecules 25, no. 9: 2187. https://doi.org/10.3390/molecules25092187
APA StyleRusu, M. E., Fizesan, I., Pop, A., Mocan, A., Gheldiu, A.-M., Babota, M., Vodnar, D. C., Jurj, A., Berindan-Neagoe, I., Vlase, L., & Popa, D.-S. (2020). Walnut (Juglans regia L.) Septum: Assessment of Bioactive Molecules and In Vitro Biological Effects. Molecules, 25(9), 2187. https://doi.org/10.3390/molecules25092187