
Sericin-Induced Melanogenesis in Cultured Retinal Pigment Epithelial Cells Is Associated with Elevated Levels of Hydrogen Peroxide and Inflammatory Proteins

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
2.3. In-Solution Digestion and Nano LC-Q Exactive Orbitrap Mass Spectrometry
2.4. Data Analysis
2.5. Immunoblotting
2.6. Light Microscopy
2.7. Scanning Electron Microscopy
2.8. Immunofluorescence
2.9. Viability Assay
2.10. Measurement of Superoxide and Hydrogen Peroxide Reactive Oxygen Species
2.11. Quantification of Pigmentation
2.12. Quantification of Microvilli
3. Results
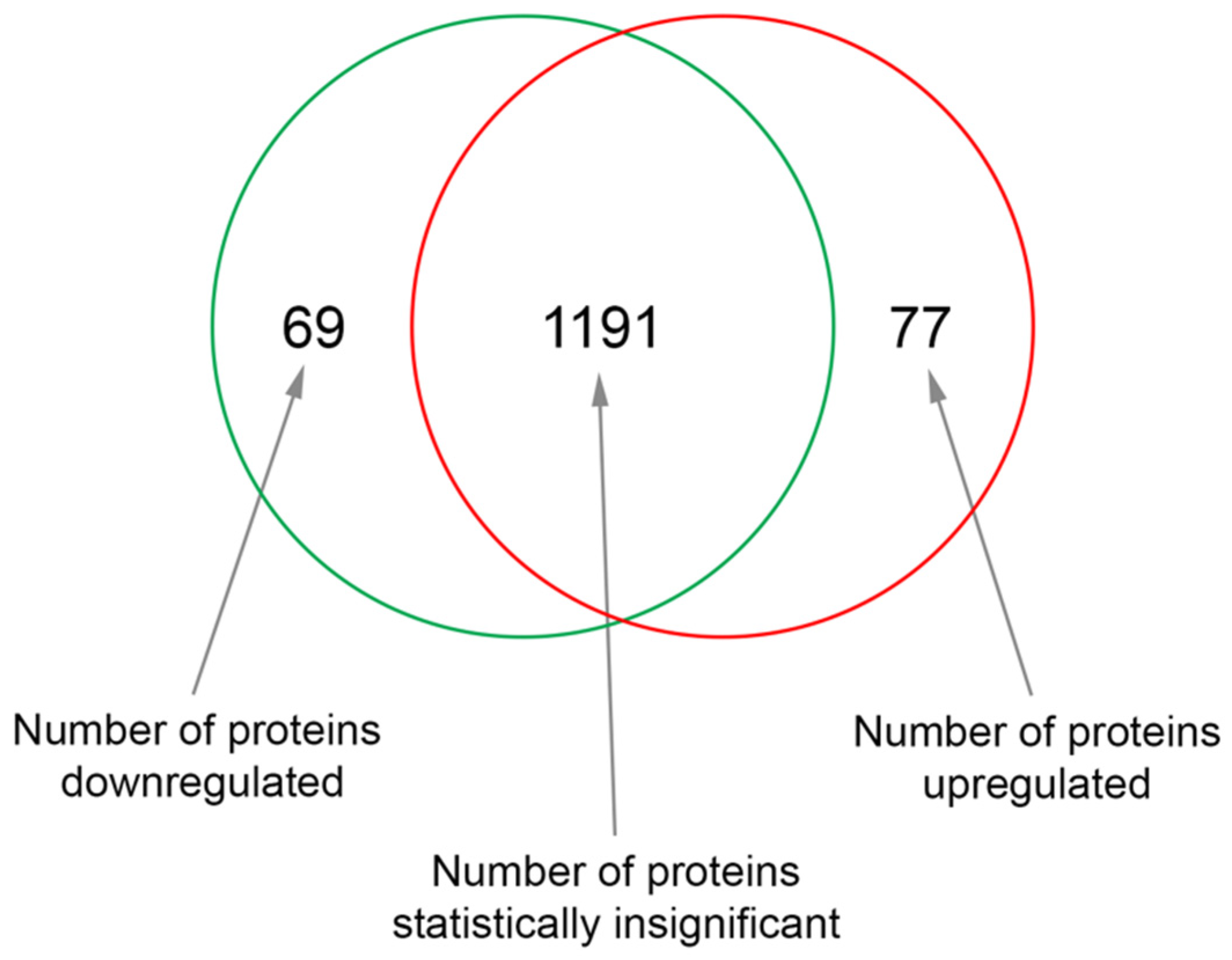
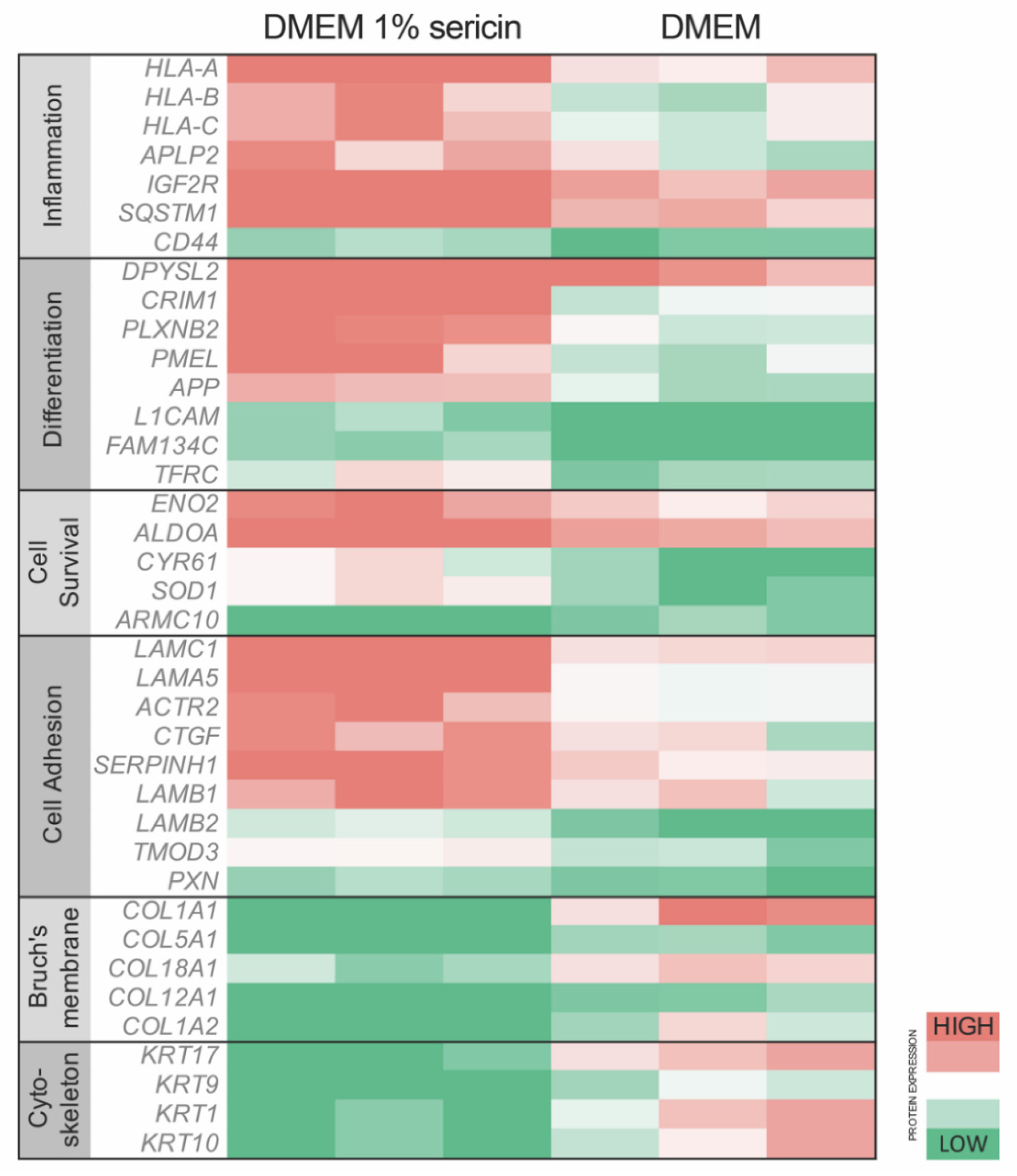
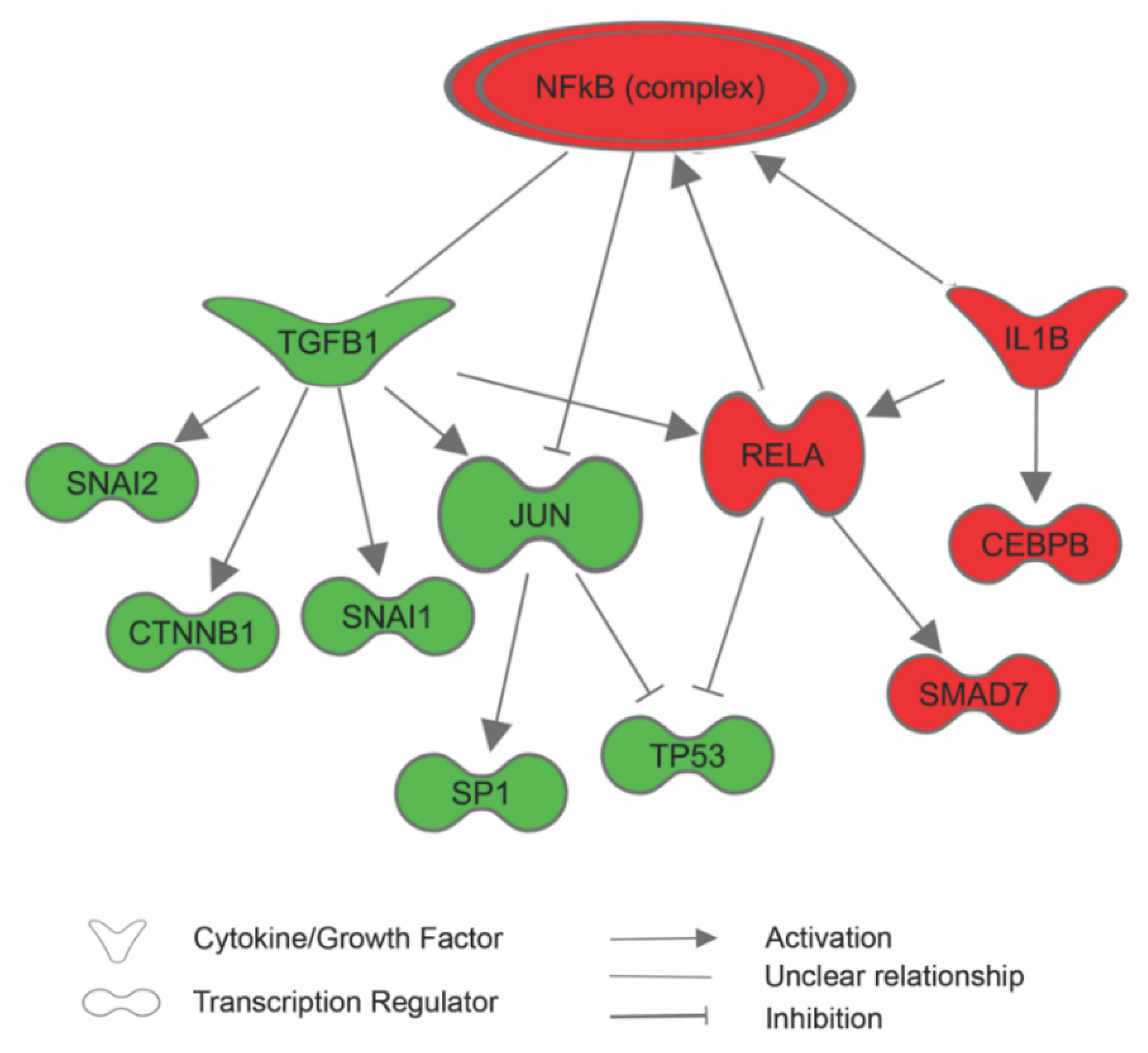
3.1. Effects of Sericin on Protein Expression
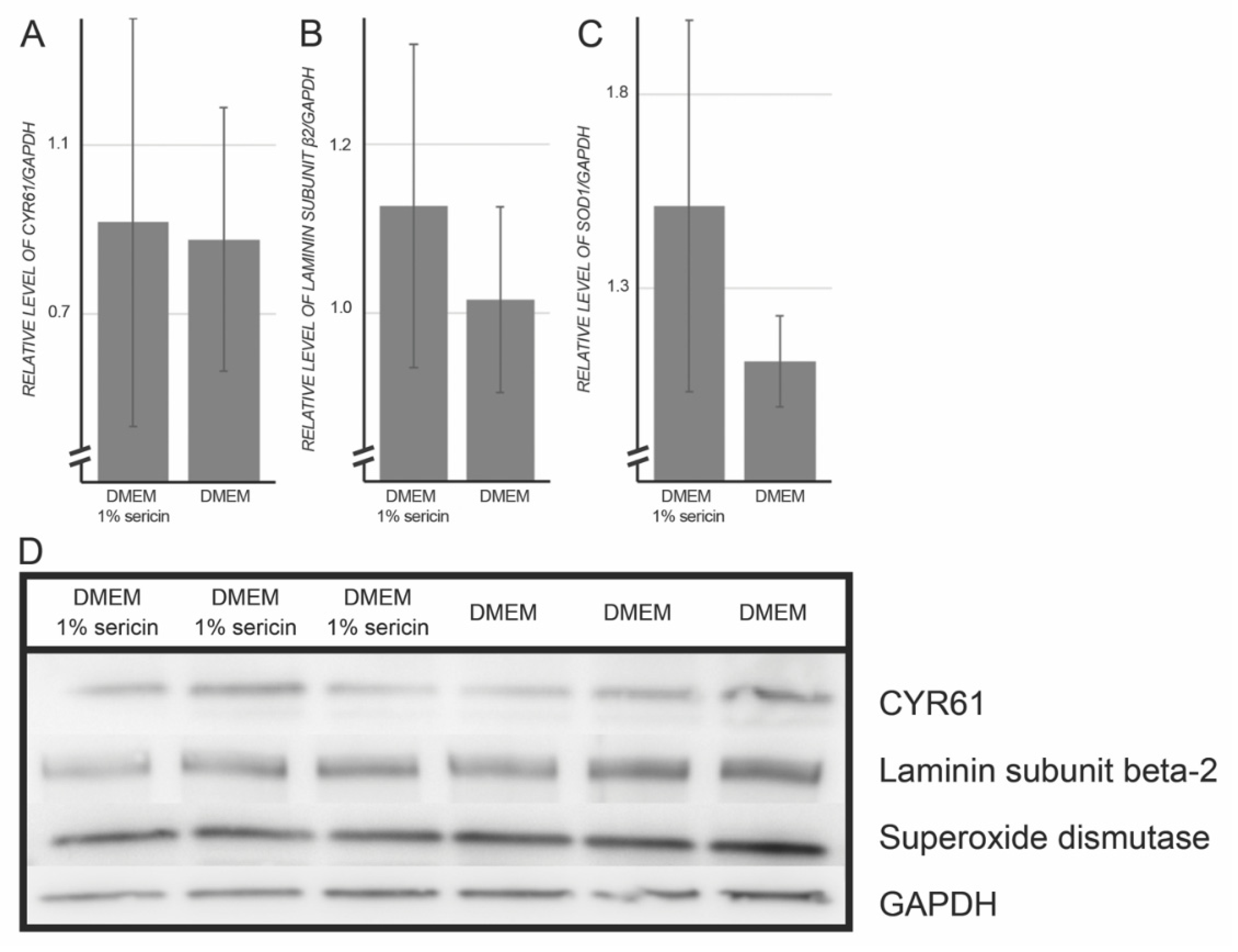
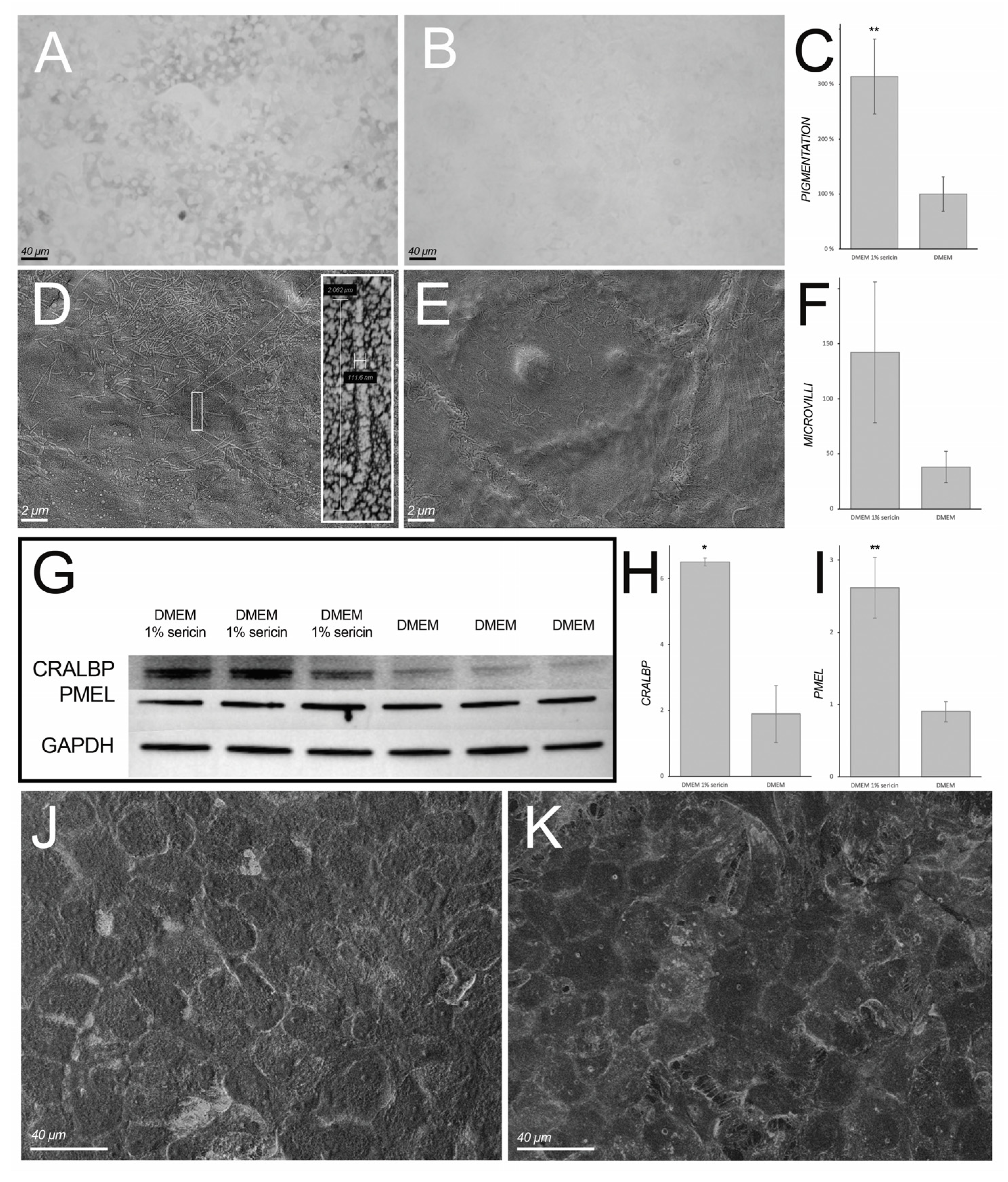
3.2. Immunofluorescence and Immunoblotting
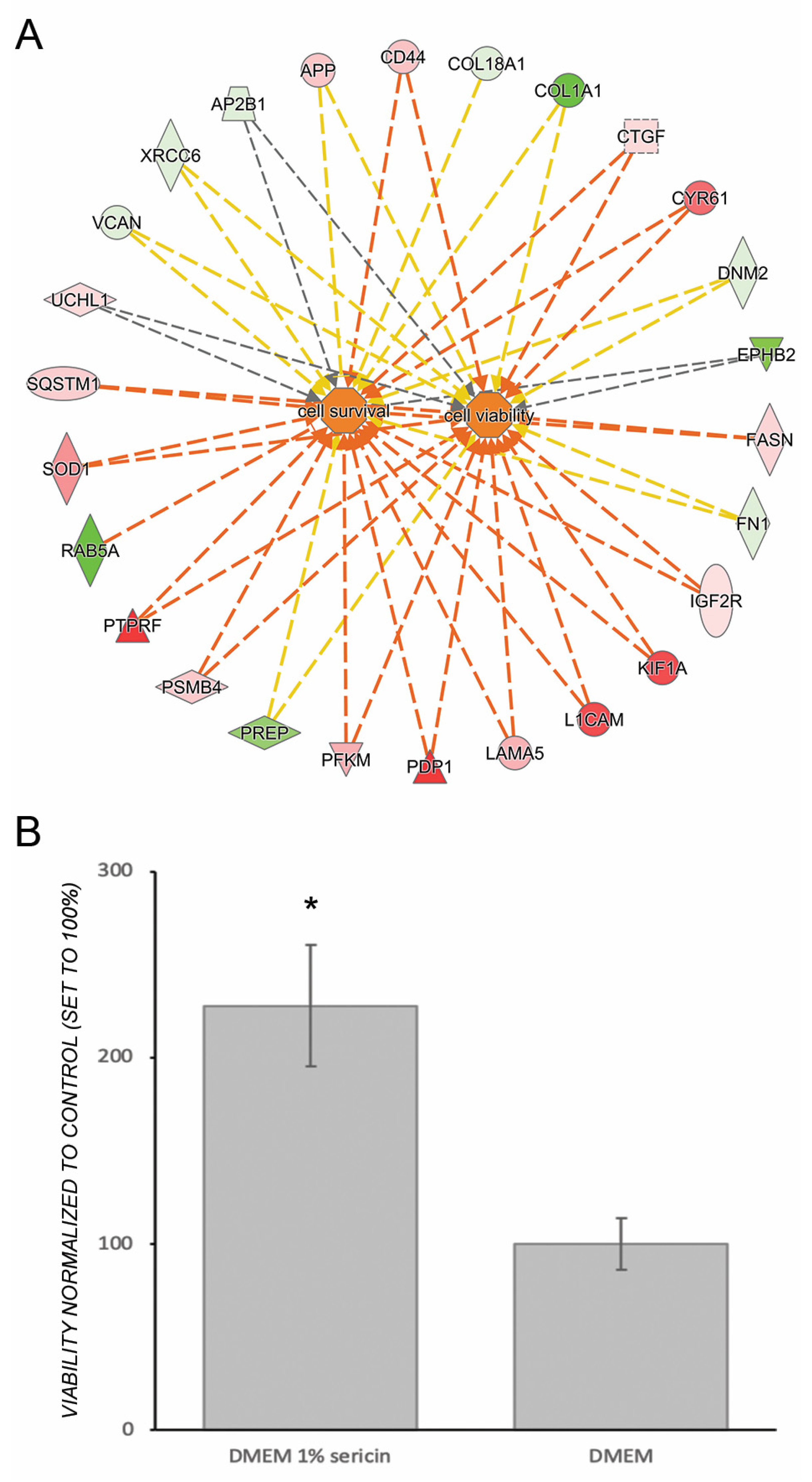
3.3. Sericin Increases Viability Despite Inducing an Inflammatory Response
3.4. Sericin Promotes Pigmentation of Cultured Human Retinal Pigment Epithelial Cells
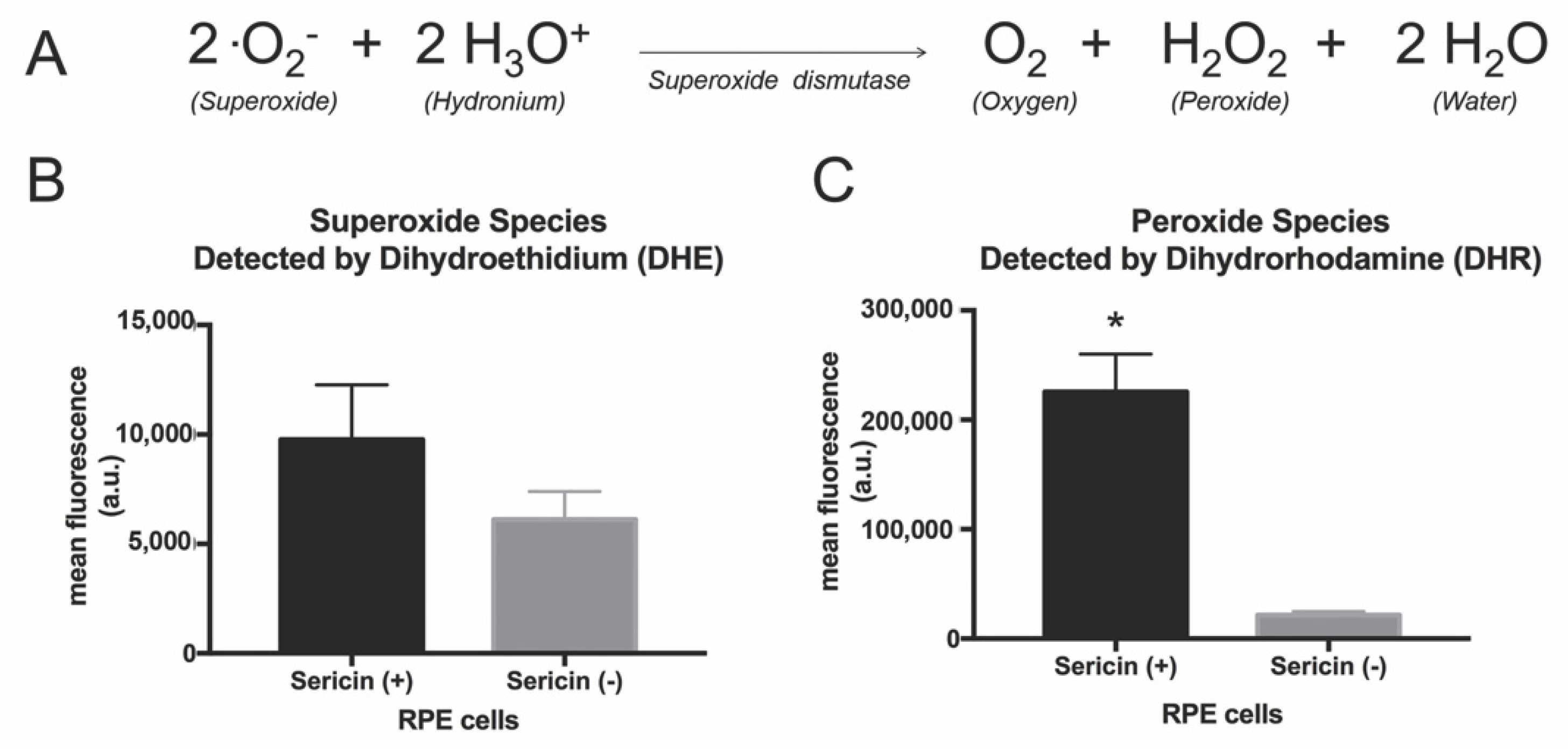
3.5. Effects of Sericin on Superoxide Dismutase Substrate and Product
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Buchholz, D.E.; Hikita, S.T.; Rowland, T.J.; Friedrich, A.M.; Hinman, C.R.; Johnson, L.V.; Clegg, D.O. Derivation of functional retinal pigmented epithelium from induced pluripotent stem cells. Stem Cells 2009, 27, 2427–2434. [Google Scholar] [CrossRef] [PubMed]
- Minoura, N.; Aiba, S.I.; Gotoh, Y.; Tsukada, M.; Imai, Y. Attachment and growth of cultured fibroblast cells on silk protein matrices. J. Biomed. Mater. Res. 1995, 29, 1215–1221. [Google Scholar] [CrossRef] [PubMed]
- Kato, N.; Sato, S.; Yamanaka, A.; Yamada, H.; Fuwa, N.; Nomura, M. Silk protein, sericin, inhibits lipid peroxidation and tyrosinase activity. Biosci. Biotechnol. Biochem. 1998, 62, 145–147. [Google Scholar] [CrossRef]
- Nagai, N.; Murao, T.; Ito, Y.; Okamoto, N.; Sasaki, M. Enhancing effects of sericin on corneal wound healing in rat debrided corneal epithelium. Biol. Pharm. Bull. 2009, 32, 933–936. [Google Scholar] [CrossRef] [PubMed]
- Kunz, R.I.; Brancalhao, R.M.C.; Ribeiro, L.D.C.; Natali, M.R.M. Silkworm sericin: Properties and biomedical applications. Biomed. Res. Int. 2016, 2016, 8175701. [Google Scholar] [CrossRef]
- Siritientong, T.; Angspatt, A.; Ratanavaraporn, J.; Aramwit, P. Clinical potential of a silk sericin-releasing bioactive wound dressing for the treatment of split-thickness skin graft donor sites. Pharm. Res. 2014, 31, 104–116. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.Z.; Utheim, T.P.; Reppe, S.; Sandvik, L.; Lyberg, T.; Roald, B.B.H.; Ibrahim, I.B.; Eidet, J.R. Cultured human retinal pigment epithelial (hRPE) sheets: A search for suitable storage conditions. Microsc. Microanal. 2018, 24, 147–155. [Google Scholar] [CrossRef]
- Eidet, J.R.; Reppe, S.; Pasovic, L.; Olstad, O.K.; Lyberg, T.; Khan, A.Z.; Fostad, I.G.; Chen, D.F.; Utheim, T.P. The silk-protein sericin induces rapid melanization of cultured primary human retinal pigment epithelial cells by activating the NF-κB pathway. Sci. Rep. 2016, 6, 22671. [Google Scholar] [CrossRef]
- Fan, J.B.; Wu, L.P.; Chen, L.S.; Mao, X.Y.; Ren, F.Z. Antioxidant activities of silk sericin from silkworm Bombyx mori. J. Food Biochem. 2009, 33, 74–88. [Google Scholar] [CrossRef]
- Schenk, H.; Klein, M.; Erdbrugger, W.; Droge, W.; Schulzeosthoff, K. Distinct effects of thioredoxin and antioxidants on the activation of transcription factors NF-κB and AP-1. Proc. Natl. Acad. Sci. USA 1994, 91, 1672–1676. [Google Scholar] [CrossRef]
- Ma, Q.; Kinneer, K.; Ye, J.; Chen, B.J. Inhibition of nuclear factor kappaB by phenolic antioxidants: Interplay between antioxidant signaling and inflammatory cytokine expression. Mol. Pharmacol. 2003, 64, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Koehler, C.J.; Thiede, B. Predominant cleavage of proteins N-terminal to serines and threonines using scandium (III) triflate. J. Biol. Inorg. Chem. 2020, 25, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.; Nesvizhskii, A.I.; Kolker, E.; Aebersold, R. Empirical statistical model to estimate the accuracy of peptide identifications made by MS/MS and database search. Anal. Chem. 2002, 74, 5383–5392. [Google Scholar] [CrossRef] [PubMed]
- Nesvizhskii, A.I.; Keller, A.; Kolker, E.; Aebersold, R. A statistical model for identifying proteins by tandem mass spectrometry. Anal. Chem. 2003, 75, 4646–4658. [Google Scholar] [CrossRef]
- Tran, T.T.; Strozynski, M.; Thiede, B. Quantitative phosphoproteome analysis of cisplatin-induced apoptosis in Jurkat T cells. Proteomics 2017, 17, 1600470. [Google Scholar] [CrossRef]
- Tezel, G.; Yang, X.; Luo, C.; Cai, J.; Powell, D.W. An astrocyte-specific proteomic approach to inflammatory responses in experimental rat glaucoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4220–4233. [Google Scholar] [CrossRef]
- Luo, C.; Yang, X.; Kain, A.D.; Powell, D.W.; Kuehn, M.H.; Tezel, G. Glaucomatous tissue stress and the regulation of immune response through glial Toll-like receptor signaling. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5697–5707. [Google Scholar] [CrossRef]
- Tezel, G.; Yang, X.; Luo, C.; Kain, A.D.; Powell, D.W.; Kuehn, M.H.; Kaplan, H.J. Oxidative stress and the regulation of complement activation in human glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 5071–5082. [Google Scholar] [CrossRef]
- Khan, A.Z.; Utheim, T.P.; Jackson, C.J.; Reppe, S.; Lyberg, T.; Eidet, J.R. Nucleus morphometry in cultured epithelial cells correlates with phenotype. Microsc. Microanal. 2016, 22, 612–620. [Google Scholar] [CrossRef]
- The UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Stelzer, G.; Rosen, N.; Plaschkes, I.; Zimmerman, S.; Twik, M.; Fishilevich, S.; Stein, T.I.; Nudel, R.; Lieder, I.; Mazor, Y.; et al. The GeneCards suite: From gene data mining to disease genome sequence analyses. Curr. Protoc. Bioinform. 2016, 54, 1.30.1–1.30.33. [Google Scholar] [CrossRef] [PubMed]
- Handler, D.C.L.; Pascovici, D.; Mirzaei, M.; Gupta, V.; Salekdeh, G.H.; Haynes, P.A. The art of validating quantitative proteomics data. Proteomics 2018, 18, 1800222. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Hayyan, M.; Hashim, M.A.; AlNashef, I.M. Superoxide ion: Generation and chemical implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef] [PubMed]
- Ersel, M.; Uyanikgil, Y.; Akarca, F.K.; Ozcete, E.; Altunci, Y.A.; Karabey, F.; Cavusoglu, T.; Meral, A.; Yigitturk, G.; Cetin, E.O. Effects of silk sericin on incision wound healing in a dorsal skin flap wound healing rat model. Med. Sci. Monit. 2016, 22, 1064–1078. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kumar, D.; Sikka, P.; Singh, P. Sericin supplementation improves semen freezability of buffalo bulls by minimizing oxidative stress during cryopreservation. Anim. Reprod. Sci. 2015, 152, 26–31. [Google Scholar] [CrossRef]
- Zhang, J.; Johnston, G.; Stebler, B.; Keller, E.T. Hydrogen peroxide activates NFκB and the interleukin-6 promoter through NFκB-inducing kinase. Antioxid. Redox Signal. 2001, 3, 493–504. [Google Scholar] [CrossRef]
- Pelle, E.; Mammone, T.; Maes, D.H.; Frenkel, K. Keratinocytes act as a source of reactive oxygen species by transferring hydrogen peroxide to melanocytes. J. Investig. Dermatol. 2005, 124, 793–797. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Li, J.; Lin, X.; Wu, W.; Kang, K.; Fu, W. Oxidation levels differentially impact melanocytes: Low versus high concentration of hydrogen peroxide promotes melanin synthesis and melanosome transfer. Dermatology 2012, 224, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.E.; Lee, S.G. Induction of ATP synthase β by H2O2 induces melanogenesis by activating PAH and cAMP/CREB/MITF signaling in melanoma cells. Int. J. Biochem. Cell Biol. 2013, 45, 1217–1222. [Google Scholar] [CrossRef] [PubMed]
- Perelman, A.; Dubinsky, Z.; Martínez, R. Temperature dependence of superoxide dismutase activity in plankton. J. Exp. Mar. Biol. Ecol. 2006, 334, 229–235. [Google Scholar]
- Rak, D.J.; Hardy, K.M.; Jaffe, G.J.; McKay, B.S. Ca++-switch induction of RPE differentiation. Exp. Eye Res. 2006, 82, 648–656. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Gene Symbol | FC | p-Value |
|---|---|---|---|
| Hydroxyacylglutathione hydrolase, mitochondrial | HAGH | 15.376 | 0.039 |
| Low-density lipoprotein receptor | LDLR | 13.523 | 0.003 |
| (Pyruvate dehydrogenase (acetyl-transferring))-phosphatase 1, mitochondrial | PDP1 | 12.853 | 0.010 |
| Receptor-type tyrosine-protein phosphatase F | PTPRF | 11.652 | 0.030 |
| Laminin subunit β-2 | LAMB2 | 10.453 | 0.001 |
| Translational activator GCN1 | GCN1L1 | 9.168 | 0.003 |
| Kinesin-like protein KIF1A | KIF1A | 9.129 | 0.029 |
| Neural cell adhesion molecule L1 | L1CAM | 9.129 | 0.029 |
| Protein FAM134C | FAM134C | 8.789 | 0.009 |
| Pyridoxine-5′-phosphate oxidase | PNPO | 8.789 | 0.009 |
| Hsc70-interacting protein | ST13 | 7.696 | 0.039 |
| Protein CYR61 | CYR61 | 7.346 | 0.020 |
| Isocitrate dehydrogenase (NAD) subunit α, mitochondrial | IDH3A | 6.797 | 0.047 |
| Leucine-rich repeat neuronal protein 1 | LRRN1 | 5.691 | 0.029 |
| Glycine cleavage system H protein, mitochondrial | GCSH | 5.495 | 0.020 |
| Protein arginine N-methyltransferase 5 | PRMT5 | 5.495 | 0.020 |
| Superoxide dismutase (Cu-Zn) | SOD1 | 5.443 | 0.003 |
| Seizure 6-like protein 2 | SEZ6L2 | 4.820 | 0.002 |
| Tetraspanin-10 | TSPAN10 | 4.669 | 0.040 |
| Sortilin | SORT1 | 4.492 | 0.041 |
| Cysteine-rich motor neuron 1 protein | CRIM1 | 4.437 | 0.009 |
| Sodium-coupled neutral amino acid transporter 2 | SLC38A2 | 4.106 | 0.021 |
| 6-phosphofructokinase, muscle type | PFKM | 3.903 | 0.015 |
| Laminin subunit α-5 | LAMA5 | 3.788 | 0.011 |
| Tubulin--tyrosine ligase-like protein 12 | TTLL12 | 3.621 | 0.035 |
| Melanocyte protein PMEL | PMEL | 3.123 | 0.021 |
| Paxillin | PXN | 3.097 | 0.029 |
| 3-hydroxyisobutyrate dehydrogenase, mitochondrial | HIBADH | 3.056 | 0.030 |
| Plexin-B2 | PLXNB2 | 3.049 | 0.011 |
| 40S ribosomal protein S28 | RPS28 | 2.956 | 0.034 |
| CD44 antigen | CD44 | 2.956 | 0.034 |
| Cullin-associated NEDD8-dissociated protein 1 | CAND1 | 2.951 | 0.049 |
| Transferrin receptor protein 1 | TFRC | 2.847 | 0.029 |
| Amyloid β A4 protein | APP | 2.838 | 0.001 |
| Calpain-1 catalytic subunit | CAPN1 | 2.751 | 0.026 |
| 60S ribosomal protein L18a | RPL18A | 2.704 | 0.007 |
| Proteasome subunit β type-4 | PSMB4 | 2.692 | 0.026 |
| 7-dehydrocholesterol reductase | DHCR7 | 2.600 | 0.048 |
| 60S ribosomal protein L13 | RPL13 | 2.551 | 0.009 |
| Small nuclear ribonucleoprotein G | SNRPG | 2.551 | 0.009 |
| NADH dehydrogenase (ubiquinone) iron-sulfur protein 2, mitochondrial | NDUFS2 | 2.546 | 0.018 |
| Sequestosome-1 | SQSTM1 | 2.487 | 0.001 |
| Heterogeneous nuclear ribonucleoprotein A1 | HNRNPA1 | 2.416 | 0.023 |
| HLA class I histocompatibility antigen, B-8 α chain | HLA-B | 2.365 | 0.031 |
| Laminin subunit γ-1 | LAMC1 | 2.334 | 0.006 |
| Eukaryotic translation initiation factor 4H | EIF4H | 2.252 | 0.019 |
| Rab GDP dissociation inhibitor β | GDI2 | 2.245 | 0.028 |
| Actin-related protein 2 | ACTR2 | 2.228 | 0.022 |
| Amyloid-like protein 2 | APLP2 | 2.203 | 0.042 |
| GTP-binding nuclear protein Ran | RAN | 2.202 | 0.009 |
| Syntenin-1 | SDCBP | 2.173 | 0.006 |
| Integrin α-3 | ITGA3 | 2.142 | 0.027 |
| Tropomodulin-3 | TMOD3 | 2.131 | 0.016 |
| HLA class I histocompatibility antigen, Cw-12 α chain | HLA-C | 2.065 | 0.012 |
| Fatty acid synthase | FASN | 2.050 | 0.025 |
| HLA class I histocompatibility antigen, A-30 α chain | HLA-A | 2.014 | 0.006 |
| Guanine nucleotide-binding protein G(s) subunit α isoforms XLas | GNAS | 1.995 | 0.010 |
| Catechol O-methyltransferase | COMT | 1.899 | 0.045 |
| Connective tissue growth factor | CTGF | 1.881 | 0.049 |
| Ubiquitin carboxyl-terminal hydrolase isozyme L1 | UCHL1 | 1.861 | 0.036 |
| Serpin H1 | SERPINH1 | 1.851 | 0.003 |
| Thioredoxin reductase 1, cytoplasmic | TXNRD1 | 1.807 | 0.049 |
| Pyruvate dehydrogenase E1 component subunit β, mitochondrial | PDHB | 1.784 | 0.004 |
| WD repeat-containing protein 1 | WDR1 | 1.776 | 0.009 |
| Laminin subunit β-1 | LAMB1 | 1.768 | 0.049 |
| Importin subunit β-1 | KPNB1 | 1.763 | 0.048 |
| Ribonuclease inhibitor | RNH1 | 1.744 | 0.023 |
| T-complex protein 1 subunit eta | CCT7 | 1.680 | 0.011 |
| Fructose-bisphosphate aldolase A | ALDOA | 1.643 | 0.023 |
| γ-enolase | ENO2 | 1.598 | 0.017 |
| Cation-independent mannose-6-phosphate receptor | IGF2R | 1.575 | 0.009 |
| S-formylglutathione hydrolase | ESD | 1.573 | 0.025 |
| 14-3-3 protein β/α | YWHAB | 1.515 | 0.037 |
| Heat shock protein HSP 90-α | HSP90AA1 | 1.460 | 0.025 |
| Dihydropyrimidinase-related protein 2 | DPYSL2 | 1.452 | 0.033 |
| Myoferlin | MYOF | 1.349 | 0.044 |
| Coronin-1C | CORO1C | 1.235 | 0.030 |
| Protein Name | Gene Symbol | FC | p-Value |
|---|---|---|---|
| Ras-related protein Rab-5A | RAB5A | 0.000 | 0.047 |
| Collagen α-2(I) chain | COL1A2 | 0.000 | 0.041 |
| Myosin phosphatase Rho-interacting protein | MPRIP | 0.000 | 0.024 |
| Collagen α-1(XII) chain | COL12A1 | 0.000 | 0.021 |
| Fibulin-1 | FBLN1 | 0.000 | 0.021 |
| Lamina-associated polypeptide 2, isoforms β/γ | TMPO | 0.000 | 0.021 |
| Zinc finger protein ubi-d4 | DPF2 | 0.000 | 0.021 |
| Lysyl oxidase homolog 1 | LOXL1 | 0.000 | 0.019 |
| Armadillo repeat-containing protein 10 | ARMC10 | 0.000 | 0.018 |
| Heterogeneous nuclear ribonucleoprotein D-like | HNRNPDL | 0.000 | 0.018 |
| Tetraspanin-6 | TSPAN6 | 0.000 | 0.018 |
| Collagen α-1(I) chain | COL1A1 | 0.000 | 0.012 |
| Cysteine-rich with EGF-like domain protein 1 | CRELD1 | 0.000 | 0.010 |
| Fragile X mental retardation syndrome-related protein 2 | FXR2 | 0.000 | 0.010 |
| Prolyl endopeptidase | PREP | 0.000 | 0.010 |
| Ras-related protein Rab-13 | RAB13 | 0.000 | 0.010 |
| RNA-binding protein 42 | RBM42 | 0.000 | 0.010 |
| Keratin, type I cytoskeletal 9 | KRT9 | 0.000 | 0.008 |
| Ephrin type-B receptor 2 | EPHB2 | 0.000 | 0.007 |
| Single-stranded DNA-binding protein, mitochondrial | SSBP1 | 0.000 | 0.007 |
| Transformer-2 protein homolog α | TRA2A | 0.000 | 0.007 |
| Collagen α-1(V) chain | COL5A1 | 0.000 | 0.006 |
| Hydroxyacyl-coenzyme A dehydrogenase, mitochondrial | HADH | 0.000 | 0.006 |
| Prolyl 4-hydroxylase subunit α-1 | P4HA1 | 0.000 | 0.004 |
| Keratin, type I cytoskeletal 17 | KRT17 | 0.049 | 0.001 |
| Keratin, type II cytoskeletal 1 | KRT1 | 0.063 | 0.013 |
| Keratin, type I cytoskeletal 10 | KRT10 | 0.075 | 0.038 |
| Septin-11 | SEPT11 | 0.114 | 0.002 |
| AP-2 complex subunit β | AP2B1 | 0.121 | 0.043 |
| Prolyl 3-hydroxylase 3 | LEPREL2 | 0.136 | 0.021 |
| Putative tropomyosin α-3 chain-like protein | TPM3L | 0.146 | 0.018 |
| Serine/arginine-rich splicing factor 1 | SRSF1 | 0.173 | 0.026 |
| Septin-7 | SEPT7 | 0.174 | 0.038 |
| S-phase kinase-associated protein 1 | SKP1 | 0.207 | 0.047 |
| Myristoylated alanine-rich C-kinase substrate | MARCKS | 0.217 | 0.020 |
| Dynactin subunit 1 | DCTN1 | 0.228 | 0.039 |
| Epiplakin | EPPK1 | 0.250 | 0.015 |
| Versican core protein | VCAN | 0.259 | 0.016 |
| EGF-containing fibulin-like extracellular matrix protein 1 | EFEMP1 | 0.271 | 0.016 |
| Y-box-binding protein 3 | YBX3 | 0.294 | 0.026 |
| Basement membrane-specific heparan sulfate proteoglycan core protein | HSPG2 | 0.300 | 0.004 |
| Myosin regulatory light polypeptide 9 | MYL9 | 0.331 | 0.032 |
| Collagen α-1(XVIII) chain | COL18A1 | 0.338 | 0.004 |
| Fibronectin | FN1 | 0.365 | 0.007 |
| U1 small nuclear ribonucleoprotein 70 kDa | SNRNP70 | 0.374 | 0.031 |
| Fibrillin-2 | FBN2 | 0.390 | 0.001 |
| X-ray repair cross-complementing protein 6 | XRCC6 | 0.397 | 0.021 |
| Fibulin-2 | FBLN2 | 0.398 | 0.005 |
| Fibrillin-1 | FBN1 | 0.419 | 0.001 |
| Histone H4 | HIST1H4A | 0.442 | 0.007 |
| β-galactosidase | GLB1 | 0.452 | 0.026 |
| Nodal modulator 3 | NOMO3 | 0.456 | 0.021 |
| Drebrin | DBN1 | 0.472 | 0.037 |
| Histone H3.1 | HIST1H3A | 0.481 | 0.015 |
| Calmodulin | CALM1 | 0.487 | 0.022 |
| General transcription factor II-I | GTF2I | 0.551 | 0.028 |
| Myosin regulatory light chain 12B | MYL12B | 0.563 | 0.001 |
| Spectrin α chain, nonerythrocytic 1 | SPTAN1 | 0.613 | 0.024 |
| Golgi-associated plant pathogenesis-related protein 1 | GLIPR2 | 0.638 | 0.013 |
| N-acetylglucosamine-6-sulfatase | GNS | 0.667 | 0.021 |
| Calnexin | CANX | 0.675 | 0.028 |
| Protein disulfide-isomerase A3 | PDIA3 | 0.681 | 0.006 |
| Voltage-dependent anion-selective channel protein 3 | VDAC3 | 0.685 | 0.007 |
| Actin, cytoplasmic 1 | ACTB | 0.690 | 0.013 |
| Actin, cytoplasmic 2 | ACTG1 | 0.690 | 0.012 |
| Prelamin-A/C | LMNA | 0.698 | 0.013 |
| Calreticulin | CALR | 0.740 | 0.020 |
| Tubulin β-4A chain | TUBB4A | 0.836 | 0.040 |
| Tubulin β-4B chain | TUBB4B | 0.892 | 0.031 |
| Protein Name | MS/MS | Immunofluorescence | Immunoblotting | |||
|---|---|---|---|---|---|---|
| FC | p-Value | FC | p-Value | FC | p-Value | |
| CYR61 | 7.3 | 0.02 | 1.10 | 0.17 | 1.05 | 0.90 |
| Laminin subunit β-2 | 10.5 | 0.001 | 1.91 | 0.007 | 1.11 | 0.43 |
| Superoxide dismutase | 5.4 | 0.003 | 1.22 | 0.25 | 1.36 | 0.23 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.Z.; Jackson, C.J.; Utheim, T.P.; Reppe, S.; Sapkota, D.; Olstad, O.K.; Thiede, B.; Eidet, J.R. Sericin-Induced Melanogenesis in Cultured Retinal Pigment Epithelial Cells Is Associated with Elevated Levels of Hydrogen Peroxide and Inflammatory Proteins. Molecules 2020, 25, 4395. https://doi.org/10.3390/molecules25194395
Khan AZ, Jackson CJ, Utheim TP, Reppe S, Sapkota D, Olstad OK, Thiede B, Eidet JR. Sericin-Induced Melanogenesis in Cultured Retinal Pigment Epithelial Cells Is Associated with Elevated Levels of Hydrogen Peroxide and Inflammatory Proteins. Molecules. 2020; 25(19):4395. https://doi.org/10.3390/molecules25194395
Chicago/Turabian StyleKhan, Ayyad Zartasht, Catherine Joan Jackson, Tor Paaske Utheim, Sjur Reppe, Dipak Sapkota, Ole Kristoffer Olstad, Bernd Thiede, and Jon Roger Eidet. 2020. "Sericin-Induced Melanogenesis in Cultured Retinal Pigment Epithelial Cells Is Associated with Elevated Levels of Hydrogen Peroxide and Inflammatory Proteins" Molecules 25, no. 19: 4395. https://doi.org/10.3390/molecules25194395
APA StyleKhan, A. Z., Jackson, C. J., Utheim, T. P., Reppe, S., Sapkota, D., Olstad, O. K., Thiede, B., & Eidet, J. R. (2020). Sericin-Induced Melanogenesis in Cultured Retinal Pigment Epithelial Cells Is Associated with Elevated Levels of Hydrogen Peroxide and Inflammatory Proteins. Molecules, 25(19), 4395. https://doi.org/10.3390/molecules25194395