
Inflammatory Bowel Diseases and Gut Microbiota
by
 , and
, and
Yuri Haneishi
1 ,
,
Yuma Furuya
1,
Mayu Hasegawa
1,
Antonio Picarelli
2,
Mauro Rossi
3 and
Junki Miyamoto
1,* 1
Department of Applied Biological Science, Graduate School of Agriculture, Tokyo University of Agriculture and Technology, Fuchu-shi 183-8509, Tokyo, Japan
2
Department of Translational and Precision Medicine, Sapienza University of Rome, 00185 Rome, Italy
3
Institute of Food Sciences, National Research Council (CNR), Via Roma 64, 83100 Avellino, Italy
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2023, 24(4), 3817; https://doi.org/10.3390/ijms24043817
Submission received: 23 January 2023
/
Revised: 13 February 2023
/
Accepted: 13 February 2023
/
Published: 14 February 2023
(This article belongs to the Special Issue Microbiome and Metabolome in the Gastrointestinal Tract)
Abstract
:Inflammatory bowel disease (IBD) is an inflammatory disease of the gastrointestinal tract, the incidence of which has rapidly increased worldwide, especially in developing and Western countries. Recent research has suggested that genetic factors, the environment, microbiota, and immune responses are involved in the pathogenesis; however, the underlying causes of IBD are unclear. Recently, gut microbiota dysbiosis, especially a decrease in the abundance and diversity of specific genera, has been suggested as a trigger for IBD-initiating events. Improving the gut microbiota and identifying the specific bacterial species in IBD are essential for understanding the pathogenesis and treatment of IBD and autoimmune diseases. Here, we review the different aspects of the role played by gut microbiota in the pathogenesis of IBD and provide a theoretical basis for modulating gut microbiota through probiotics, fecal microbiota transplantation, and microbial metabolites.
1. Introduction
The incidence of immune-mediated diseases, such as autoimmune diseases, continues to increase worldwide [1]. Autoimmune diseases are caused by genetic and environmental factors. It is a general term for diseases in which the immune system, instead of eliminating foreign substances, such as bacteria, viruses, and tumors that differ from itself, malfunctions, overreacts, and attacks its own normal cells and tissues. Additionally, the disruption of the intestinal barrier and changes in the intestinal microbiome are thought to be responsible for various autoimmune and metabolic diseases [2]. Irritable bowel syndrome (IBS), inflammatory bowel diseases (IBD), and celiac disease, as well as obesity and diabetes mellitus, play an important pathogenetic role in these alterations [3,4,5]. During diets conducted in the suspicion of these diseases, dysbiosis seems to play an important role in the emergence of increased intestinal permeability (leaky gut). The human intestinal tract contains approximately 1000 species of intestinal bacteria, totaling 100 trillion bacteria, which are likely to be involved in regulating host homeostasis [6,7]. In recent years, a comprehensive analysis of intestinal bacteria has actively identified gut microbiota species and their metabolites that are directly involved in the pathogenesis of metabolic diseases. These are expected to play an important role in maintaining host homeostasis [8,9,10,11]. Clearly, the gut microbiota influences the pathogenesis and pathology of diseases [12,13,14], and the elucidation of the gut microbiota may contribute to reducing disease risk (Table 1).
IBD is an autoimmune disease. Its incidence continues to rapidly increase worldwide, especially in developing and Western countries [22]. IBD is an inflammatory disease that results from immune disorders arising from complex interactions between genetic and environmental factors such as diet, alcohol, stress, insufficient sleep, and the microbiome [23]. IBD is characterized by the chronic relapse and remission of inflammation in the gastrointestinal tract. It is classified into two main types: ulcerative colitis (UC) and Crohn’s disease (CD). The quality of life in IBD patients has decreased because of symptoms such as diarrhea, abdominal cramps, hematochezia, weight loss, fever, fatigue, and anemia [24]. CD is characterized by transmural inflammation affecting any part of the gastrointestinal tract such as the mouth, anus, and entire intestinal layer, whereas UC typically affects the mucosal layer of the colon [25]. As the species comprising the gut microbiota and the composition vary in a different part of the gastrointestinal tract [26], the impact of UC and CD might be in different areas of inflammation with distinctive intestinal microbiota, which may develop and aggravate the pathological condition. Unlike infectious diseases, IBD is caused by defects in the autoimmune system in the gastrointestinal tract, and a treatment that can completely cure IBD is still lacking [27]. The disruption of the immune system caused by dysbiosis of the gut microbiota increases inflammation in the gastrointestinal tract and leads to the onset of IBD [28]. Therefore, improving the gut microbiota and identifying the specific bacterial species in IBD could be significant for understanding the pathogenesis and treatment of IBD and autoimmune diseases.
Here, we aimed to provide an overview of the interaction between IBD and the gut microbiota, especially the factors, prevention, and therapy for IBD. We have also reviewed the gut microbiota that are associated with IBD and modulate the microbiome using probiotics, fecal microbiota transplantation (FMT), and microbial metabolites.
2. Gut Microbiota and IBD
Currently, in Europe and the United States, 3.6 million people suffer from IBD, including UC and CD, which are becoming increasingly prevalent worldwide [29]. IBD is an autoimmune disease involving intestinal inflammation; however, it has garnered attention due to the crosstalk between the gut microbiota and autoimmune systems. The gut microbiota has been shown to trigger IBD-initiating events. The composition of the gut microbiota is different between healthy participants and IBD patients [30]. The use of antibiotics also leads to the onset of IBD due to the alteration of the composition and function of the gut microbiota [31]. The interaction between the gut microbiota and host plays a critical role in maintaining the function of the host’s immune system [32]. Changing environmental events such as the overuse of antibiotics and changes in diet may enhance autoimmune and inflammatory disorders by modulating the gut microbiota composition, leading to dysbiosis [33]. Thus, a crucial relationship exists between IBD and the microbial communities in the human gut. Although IBD is widely attributed to altered interactions between gut microbes and the intestinal immune system, the exact mechanism underlying gut microbiota dysfunction in IBD remains unclear [34]. Although the pathogenesis of IBD is unknown, the inflamed gastrointestinal tract in patients with IBD is a common feature of an imbalance (dysbiosis) in the gut microbiota. Recently, much evidence has been provided to show that gut dysbiosis leads to the disruption of immune tolerance, which may induce or exaggerate IBD [35].
Studies with human participants have clarified that the composition of the gut microbiota is different in patients with IBD compared with that in healthy subjects [30]. Additionally, the gut microbiota is different between patients with UC and CD [36]. Analyses of the gut microbiota in patients with IBD worldwide has led to the observation that dysbiosis involves an increase or decrease in specific intestinal bacterial species in patients with IBD. Morgan et al. reported that a large, long-term prospective cohort study showed that patients with IBD have characteristic gut microbiota compared with those in healthy participants [37]. The abundance of Roseburia and Phascolarctobacterium was significantly decreased, whereas that of Clostridium was increased in the gut microbiota of patients with UC or CD. Roseburia is associated with anti-inflammatory regulatory T cell production in the intestinal tract [38], and Phascolarctobacterium consumes only succinate and produces propionic acid when co-cultured with Paraprevotella [39]. Propionic acid is a short-chain fatty acid (SCFA) that has anti-inflammatory effects [40]. Patients with IBD have decreased SCFA-producing Phascolarctobacterium, suggesting that the anti-inflammatory effects of SCFAs might be reduced, which could consequently exacerbate IBD symptoms.
The diversity and species richness of the Clostridium leptum group, which is one of the major bacterial groups comprising 16–25% of the human gut microbiota, were different between healthy participants and those with IBD in the remission phase [41]. The C. leptum group, also called Clostridial cluster IV, includes Faecalibacterium prausnitzii, Eubacterium, and Ruminococcus. F. prausnitzii shows a reduced abundance in patients with CD and in mouse models of colitis [42]. The C. leptum group dominated by F. prausnitzii showed a significantly decreased abundance in the fecal microbiota of patients with CD or UC and was detected during both the disease activity and remission stages. This suggests either that the reduction in F. prausnitzii levels leads to the onset of IBD or that IBD onset events may have caused the decrease in F. prausnitzii levels [41]. Additionally, the study showed that a low abundance of F. prausnitzii in the ileum of patients with IBD during surgical resection is a risk factor for endoscopic recurrence. Patients with CD who were treated for endoscopic recurrence at 6 months showed lower levels of Firmicutes, especially those of F. prausnitzii, which is the main bacteria of the C. leptum group. Moreover, F. prausnitzii—a defective commensal bacterium in CD—was found to exert anti-inflammatory effects in vitro and in vivo. This suggests that the use of F. prausnitzii as a probiotic to improve gut dysbiosis leads to the remission of serious CD symptoms [42].
Generally, close relatives living in the same environment have similar gut microbiota, but a study in twins reported that when one twin pair was healthy and the other had IBD, the microbial compositions in the twins were different [43]. As this study included twins, it eliminated the influence of the genetic background on the abundance of F. prausnitzii and other specific genera such as Alistipes, Collinsella, and Ruminococcaceae. This result suggests that the onset of IBD might be more strongly influenced by environmental factors than by genetic factors, especially with regard to the gut microbiota, which were reduced in IBD patients compared to healthy subjects. These results suggested that the onset of IBD might be more strongly influenced by environmental factors, especially gut microbiota, than by genetic factors. Moreover, a multi-omics analysis including metagenomic, metatranscriptomic, metabolomic, and other analyses revealed that the gut microbiota and their metabolites in CD and UC were different during the active and remission phases [44]. Thus, a multi-omics approach identified the characteristics of gut microbiota in IBD and the metabolites produced by the gut microbiota. This may clarify the impact of the crosstalk between the gut microbiota, their metabolites, and the host on the pathogenesis of IBD. Furthermore, follow-up studies that characterize the microbial features in several hosts could provide treatment plans adapted to each individual. However, a larger sample size may be needed. Nevertheless, these molecular biological approaches could be helpful in the development of novel therapies targeting host–microbe interactions as predictive biomarkers of IBD onset, progression, and recovery (Figure 1).
3. Effect of Probiotics, Prebiotics, and Symbiotics on IBD
Despite the lack of definitive therapies for IBD, approaches targeting gut microbiota have been reported in recent years to cure or improve IBD. VSL#3 is one of the most well-known probiotics for IBD treatment. VSL#3 is a probiotic mixture that includes the following eight bacteria at a concentration of 450 billion live bacteria per sachet: Bifidobacterium breve, B. longum, B. infantis, Lactobacillus acidophilus, L. plantarum, L. paracasei, L. delbrueckii subsp. Bulgaricus, and Streptococcus thermophilus [45]. VSL#3 improves IBD symptoms by improving the intestinal environment, enhancing microbial diversity, and modulating the concentration of specific bacteria such as increasing that of Bifidobacterium and decreasing that of Turicibacter in animal models [46,47,48]. Moreover, the administration of VSL#3 improved inflammatory responses in the intestine, suggesting that the remodeling of the composition of the gut microbiota by VSL#3 may have suppressed intestinal inflammation, which ameliorated IBD pathologies. Additionally, several clinical studies have reported that VSL#3 is effective in the improvement of IBD [49,50,51,52]. Although the safety of VSL#3 is recognized [53], the issues surrounding the use of VSL#3 are yet to be elucidated in clinical applications; therefore, the confirmation of the efficacy of VSL#3 could lead to a widely spread use of VSL#3 for IBD treatment.
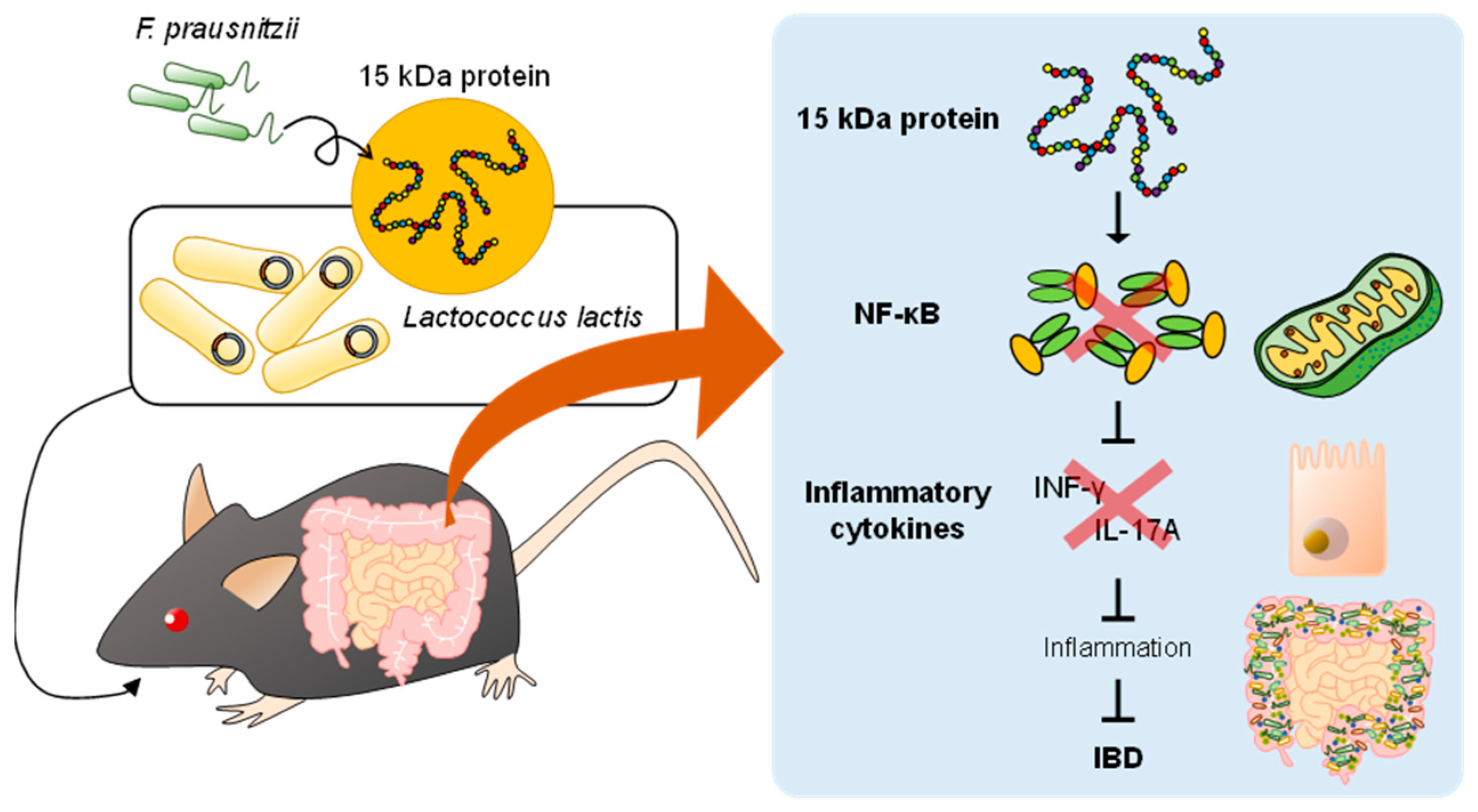
F. prausnitzii decreases pro-inflammatory cytokine levels and promotes the secretion of anti-inflammatory cytokines, suggesting that F. prausnitzii may have the ability to regulate immunity [42]. However, whether the bacterium itself or the metabolites it produced exerted the anti-inflammatory effect is unclear. However, some studies have revealed that the metabolites produced by F. prausnitzii exert anti-inflammatory effects in CD [54]. An unknown bioactive peptide derived from a 15 kDa protein (ZP05614546.1), which has anti-inflammatory effects, was discovered by screening metabolites from F. prausnitzii. Quévrain et al. also found that metabolites produced by F. prausnitzii exerted anti-inflammatory effects by the inhibition of NF-κB signaling in vitro and in vivo and that these metabolites significantly reduced NF-κB pathway activation in a dose-dependent manner. Lactococcus lactis transfected with plasmids encoding anti-inflammatory peptides produced by F. prausnitzii alleviated dinitrobenzene sulfonic acid (DNBS)-induced colitis in mice [54] (Figure 2).
Several reports have indicated that a certain probiotic bacterial strain improved IBD or colitis. Escherichia coli Nissle 1917 (EcN) has been proposed as an efficient probiotic for UC in a clinical trial [55]. In a double-blind, double-dummy study, patients with UC in remission received a bacterial preparation containing viable EcN or mesalazine in tablet form for 5 days. The formulation was meant to prevent UC relapse. After a 12-month follow-up, no difference was observed in the relapse of UC between the EcN-treated and mesalazine-treated groups. Hence, EcN could be a new probiotic formulation to replace mesalazine. Some Lactobacillus species are useful for the treatment of IBD. The oral administration of L. rhamnosus GG reduced inflammatory cytokine levels and improved colonic histology scores, which prevented colitis relapse in rats during antibiotic treatment [56]. Additionally, a study using mouse models of colitis showed that Enterobacter ludwigii was effective in alleviating the symptoms of colitis [57]. In a dextran sulfate sodium (DSS)-induced colitis mouse model, compared with that of other antibiotic treatments, metronidazole had the best effect in reducing colitis, which was related to an increase in the abundance of the gut microbiota species E. ludwigii. The administration of E. ludwigii induced Treg differentiation via metabolites from E. ludwigii and increased the immune tolerance response, which reduced the susceptibility to DSS-induced colitis in mice [57].
Prebiotics are non-digestible food ingredients that exert beneficial effects by selectively stimulating the growth of bacteria that can improve host metabolism in the gastrointestinal tract. The abundance of Bifidobacteria increased in patients with CD following prebiotic oligofructose and inulin ingestion [58]. The improvement in the intestinal environment through prebiotic administration is well documented in not only animal models but also humans. Therefore, these results suggest that prebiotics may be effective food ingredients for improving the intestinal environment in IBD pathology. Symbiotics are microbial therapeutics that, when administered in combination with prebiotics and probiotics, can benefit the host health to a greater degree than prebiotics or probiotics alone. In the meta-analysis, the administration of probiotics, prebiotics, or symbiotics to patients with IBD alleviated or improved IBD symptoms in all groups and increased the abundance of beneficial Bifidobacteria [59]. In particular, the symbiotic group showed the highest improvement, indicating that symbiotics may be more effective than prebiotics or probiotics in improving the pathophysiology of IBD.
4. FMT for IBD
FMT is an emerging therapy that may be a potential therapeutic approach for IBD. Based on the key role of gut microbiota in the pathogenesis of IBD, FMT can restore intestinal mucosal immune homeostasis in patients with IBD, which is a current research hotspot. FMT can be used to treat moderate to severe IBD complicated by recurrent or refractory Clostridium difficile infection [60,61,62,63].
FMT may be an efficient therapy for IBD in animal models. In mice with DSS-induced colitis, FMT suppressed colon inflammation and restored intestinal barrier functions and the amelioration of colitis by suppressing colon damage and recovering colon length [64,65]. Moreover, FMT-treated mice showed a decrease in the abundance of specific gut microbiota, such as Bacteroides acidifaciens, Escherichia-Shigella, and Blautia, which induce inflammation. This suggests that FMT mitigated colitis in mice by suppressing colitis-associated inflammation by improving the intestinal microbiota. Zhang et al. reported that FMT, in addition to improving colitis pathogenesis in mice with gut dysbiosis, also significantly increased the intestinal bacterial metabolite SCFA levels [66]. SCFAs regulate inflammatory responses and restore immune function [67], and Zhang et al. demonstrated that FMT may alleviate IBD by altering the composition of gut microbiota by increasing the SCFAs produced by gut microbiota. FMT is currently being investigated in registered and ethical clinical research studies and has not been approved for clinical application. Further research is needed to better understand the mechanisms underlying the effects of FMT to develop more effective regimens for treating IBD [68,69,70].
A double-blind study in patients with UC investigated the association between the remission of UC symptoms and gut microbiota [71]. Following treatment with FMT or a placebo for active UC, the gut microbiota was altered in patients in the FMT group. Additionally, increased α- and β-diversity of the gut microbiota and an increased abundance of Eubacterium hallii, Roseburia inulivorans, Eggerthella, and Ruminococcus bromii were observed in patients with UC remission in the FMT group. Moreover, increased levels of gut microbial metabolites (such as SCFAs and secondary bile acids) in feces were observed in patients with UC remission in the FMT group [72]. Furthermore, the treatment of FMT in patients with active CD also showed improvements in symptoms. Similarly, an increase in α-diversity was observed in patients with CD remission. Therefore, the FMT-induced alteration of the gut microbial composition may at least partially alleviate the signs and symptoms of IBD.
5. Microbial Metabolites and IBD
The Westernization of diets, such as an increased prevalence of high-fat and high-sugar diets, has induced both non-communicable diseases such as obesity, diabetes, and cardiovascular diseases and immune-related diseases such as IBD, hay fever, and celiac disease [33,73]. As diet could have a significant influence on the host immune system, developing methods for preventing and ameliorating autoimmune diseases using diet therapy is essential [74,75,76]. Celiac disease is another autoimmune disease that causes inflammation of the intestinal tract similar to IBD, but celiac disease symptoms are dramatically alleviated by the removal of gluten contained in wheat from the diet [77]. IBD does not have an established diet therapy; however, in recent years, the quantity and quality of the diet has been suggested to alter the host’s intestinal microbiota and influence the development of IBD [78]. For example, food emulsifiers, which are widely contained in processed foods in Western diets, increase bacterial intestinal permeability in vitro and promote early lesions of inflammation in IBD, whereas dietary fiber, which is scarce in Western diets, inhibits these responses [79]. The administration of food emulsifiers to mice promoted obesity and inflammation and exacerbated the symptoms of colitis. In addition, the gut microbiota composition was dramatically altered in mice treated with food emulsifiers compared with that in the controls [80]. Additionally, the quantity of SCFAs and bile acids produced by gut microbiota was altered in emulsifier-supplemented mice, indicating that emulsifiers may affect the gut microbiota and its metabolites, which may induce obesity and colitis [80].
SCFAs are fatty acids with two to six carbon atoms. The major SCFAs are acetic, propionic, and butyric acid. SCFAs are major intestinal bacterial metabolites produced by the fermentation of non-digestible polysaccharides, such as dietary fiber, by intestinal bacteria. SCFAs contribute to biological homeostasis by being used as an energy source for colonic epithelial cells and fatty acid synthesis in peripheral tissues [81]. SCFAs play an important role in host metabolic regulation and may contribute to improving the amelioration of immune disorders such as IBD [82]. SCFAs were decreased in the feces of patients with IBD compared with that of healthy subjects, which might be attributed to dietary differences [83]. The SCFA levels in the fecal samples of patients with UC were significantly lower compared with those of healthy participants, but the SCFA levels increased during the remission of UC, indicating that SCFAs could change with the stage of disease activity [84]. Butyrate derived from commensal gut microbes effectively facilitated anti-inflammatory M2 macrophage polarization, which influenced the immune system and improved intestinal inflammation in a mouse DSS-induced colitis model. Thus, SCFAs from gut microbiota may be a novel activator with anti-inflammatory properties and a therapeutic target for IBD [85]. The administration of Pediococcus pentosaceus LI05 changed gut microbial profiles by promoting SCFA production, modulating the gut microbial composition, and increasing the abundance of specific genera (Akkermansia, Faecalibacterium, and Ruminiclostridium_5), which ameliorated DSS-induced colitis [86]. Moreover, the ingestion of β-glucans, which are non-digestible complex dietary polysaccharides commonly contained in barley, increased SCFA levels, which also ameliorated DSS-induced colitis [87]. Mice fed saturated fatty acids, which are abundant in dairy products, meat, and palm oil, showed increased concentrations of the sulfite-reducing pathogen Bilophila wadsworthia hydrogen sulfide and secondary bile acids produced by B. wadsworthia, which promoted the onset of colitis [88]. In a clinical study, the short-term consumption of an animal-based diet significantly increased the B. wadsworthia abundance and the expression level of gut bacterial genes encoding bile acid and sulfite reductase, suggesting that dietary fat is abundant in the Western diet and may be capable of triggering IBD by altering the intestinal environment [89].
The gut microbiota metabolizes primary bile acids synthesized from cholesterol in the liver into secondary bile acids. Recently, it has been reported that the number of secondary bile acids in plasma and feces decreases in patients with IBD [90]. The administration of human microbiota decreases the production of secondary bile acids in donor mice, which may disrupt the immune response and induce IBD onset [91]. Tryptophan, an essential amino acid, is converted by the gut microbiota into quinolinic acid, which has been associated with IBD [92]. Quinolinic acid was significantly increased in patients with IBD, and gut microbial enzymes involved in the tryptophan metabolism were activated in a large cohort study of 535 IBD patients in Germany, including patients with UC and CD. This result suggests that gut dysbiosis may induce IBD by promoting the conversion of tryptophan to quinolinic acid. Aryl hydrocarbon receptor (AhR) ligands include well-known chemicals such as dioxin and benzopyrene. AhR ligands exert anti-inflammatory effects by regulating various immunological responses, and, recently, it has been shown that the administration of AhR ligands improves gastrointestinal symptoms in patients with UC [93]. These results suggest that diet-induced changes in the gut microbiota and their metabolites alter the host immune system and metabolism, which may contribute to the development of IBD. However, the immune mechanism underlying the role of the metabolites in IBD remains unclear. Therefore, understanding the interaction between food, gut microbiota, and bacterial metabolites may play a key role in diet therapy for IBD.
6. A Potential Innovative Strategy by Postbiotic Treatment
Impairments in IBD immune regulation have been described at different levels. Regarding the lymphocytic response, CD has been mainly associated with a Th (T helper)1/Th17 condition, whereas UC is characterized by an altered Th2 response [94]. An association of dysbiosis in IBD pathogenesis is also evident. Consequently, the administration of probiotics and their derivatives might be considered for reducing the inflammation or even strengthening the intestinal epithelial barrier [95]. Postbiotics mainly refer to biologically active components secreted by bacteria [96]. Their advantages over probiotics include a reduced risk of infection or potential side effects triggered by the administration of viable microorganisms to immunocompromised individuals. Among postbiotics, self-assembling protein subunits surface-layer proteins (Slp) have been recently considered. Slp constitute the first line of contact and are directly involved in several molecular mechanisms responsible for beneficial health effects [97]. Slp subunits are held together and are attached to cell wall carbohydrates by non-covalent interactions [98]. Moreover, Slp are associated with additional cell surface proteins, named S-layer-associated proteins (SLAP). These proteins exhibit immunomodulatory abilities as well as other biological functions [99]. Slp and phagocytic cells expressing sensing receptors synergistically interact to fine-tune the T cell signaling that is critical for protecting the host against pathogenic inflammation [100]. Pretreatment with Slp from Lactobacillus acidophilus NCFM reduced proinflammatory cytokines [101]. In vivo, SlpA from L. acidophilus NCFM reduced intestinal inflammatory cytokines and TLR4 and COX2 expression in a DSS-induced colitis mouse model [102]. More recently, a protective role of glycosylated SlpA expressed by the Propionibacterium strain P. UF1 in chemically induced colitis has been shown [103]. Histological analysis demonstrated severe crypt loss with associated mucosal and submucosal inflammation in the colonic tissues of mice gavaged with PBS or ΔlpsA P. UF1, while DSS-induced colitis was substantially mitigated in mice gavaged with wild type P. UF1. In vitro results suggested that glycosylated LpsA regulated colonic DC metabolic function during intestinal inflammation. In particular, the glycosylation of LpsA was shown to regulate the interaction with the sensing receptor SIGNR1. Activating critical signals involving SIGNR1 and its human homolog, DC-SIGN, may have relevance for developing therapeutic strategies for mitigating intestinal inflammation. Taken together, these reported preclinical studies are pivotal for a deeper understanding of beneficial Slp and pave the way for the development of new postbiotic therapeutic strategies for potentially treating IBD.
Biomarkers targeting the gut microbiota represent noninvasive and reasonable methods that may enable the early detection of disease, the prognosis of the disease course, and even personalized treatments for individual pathogenesis. Many scientists have evaluated biomarkers for IBD targeting the gut microbiota in recent years [104,105,106]. Klebsiella oxytoca, Morganella morganii, and Citrobacter amalonaticus increased in patients with CD, and C. portucalensis, C. pasteurii, C. werkmanii, and Proteus hauseri increased in patients with UC, indicating that specific gut microbial species can be used to distinguish the pathologies of CD and UC [107]. Additional research on the species of gut microbiota that are associated with the onset, pathogenesis, or pathophysiology of IBD will help in diagnosing IBD, as well as determining its prognosis and developing treatment options.
7. Conclusions
A massive effort in recent years to elucidate microbiome–IBD interactions has led to a better understanding of the pathophysiology of IBD. Patients with IBD and colitis model mice showed altered gut microbial profiles compared with those of healthy participants and mice, suggesting that treatment with probiotics and FMT may ameliorate IBD symptoms by targeting the gut microbiota. Moreover, since dysbiosis of the gut microbiota might result from changes in the diet, qualitative differences in the diet could be expected to restore the intestinal microbiota in patients with IBD and contribute to the improvement of their pathophysiology. Therefore, the quality of the daily diet contributes to improving the gut microbiota and may ameliorate the pathogenesis of IBD.
Author Contributions
Y.H., Y.F., M.H., A.P., M.R. and J.M. contributed equally to all aspects of the article. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by research grants from the JSPS KAKENHI (JP22K17771), the Uehara Memorial Foundation, the Lotte Foundation, the Astellas Foundation for Research on Metabolic Disorders, the Asahi Glass Foundation, and Technologically Advanced research through Marriage of Agriculture and engineering as Groundbreaking Organization (TAMAGO) to J.M.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Prescott, S.L. Early-life environmental determinants of allergic diseases and the wider pandemic of inflammatory noncommunicable diseases. J. Allergy Clin. Immunol. 2013, 131, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Al Bander, Z.; Nitert, M.D.; Mousa, A.; Naderpoor, N. The Gut Microbiota and Inflammation: An Overview. Int. J. Environ. Res. Public Health 2020, 17, 7618. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Warmbrunn, M.V.; Nieuwdorp, M.; Clément, K. Metabolism and Metabolic Disorders and the Microbiome: The Intestinal Microbiota Associated With Obesity, Lipid Metabolism, and Metabolic Health-Pathophysiology and Therapeutic Strategies. Gastroenterology 2021, 160, 573–599. [Google Scholar] [CrossRef] [PubMed]
- Sekirov, I.; Russell, S.L.; Antunes, L.C.; Finlay, B.B. Gut microbiota in health and disease. Physiol. Rev. 2010, 90, 859–904. [Google Scholar] [CrossRef] [PubMed]
- Maloy, K.J.; Powrie, F. Intestinal homeostasis and its breakdown in inflammatory bowel disease. Nature 2011, 474, 298–306. [Google Scholar] [CrossRef]
- Honda, K.; Littman, D.R. The microbiota in adaptive immune homeostasis and disease. Nature 2016, 535, 75–84. [Google Scholar] [CrossRef]
- Hooper, L.V.; Littman, D.R.; Macpherson, A.J. Interactions between the microbiota and the immune system. Science 2012, 336, 1268–1273. [Google Scholar] [CrossRef]
- Nieuwdorp, M.; Gilijamse, P.W.; Pai, N.; Kaplan, L.M. Role of the microbiome in energy regulation and metabolism. Gastroenterology 2014, 146, 1525–1533. [Google Scholar] [CrossRef]
- Gomes, A.C.; Hoffmann, C.; Mota, J.F. The human gut microbiota: Metabolism and perspective in obesity. Gut Microbes 2018, 9, 308–325. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-talk between Akkermansia muciniphila and intestinal epithelium controls diet-induced obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [Green Version]
- Brestoff, J.R.; Artis, D. Commensal bacteria at the interface of host metabolism and the immune system. Nat. Immunol. 2013, 14, 676–684. [Google Scholar] [CrossRef] [PubMed]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut microbiota from twins discordant for obesity modulate metabolism in mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Ivanov, I.I.; Honda, K. Intestinal commensal microbes as immune modulators. Cell Host Microbe 2012, 12, 496–508. [Google Scholar] [CrossRef] [PubMed]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.; Wong, F.S.; Szot, G.L.; Bluestone, J.A.; et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Shimokawa, C.; Kato, T.; Takeuchi, T.; Ohshima, N.; Furuki, T.; Ohtsu, Y.; Suzue, K.; Imai, T.; Obi, S.; Olia, A.; et al. CD8+ Regulatory T Cells Are Critical in Prevention of Autoimmune-Mediated Diabetes. Nat. Commun. 2020, 11, 1922. [Google Scholar] [CrossRef]
- Cignarella, F.; Cantoni, C.; Ghezzi, L.; Salter, A.; Dorsett, Y.; Chen, L.; Phillips, D.; Weinstock, G.M.; Fontana, L.; Cross, A.H.; et al. Intermittent Fasting Confers Protection in CNS Autoimmunity by Altering the Gut Microbiota. Cell Metab. 2018, 27, 1222–1235.e6. [Google Scholar] [CrossRef]
- Berer, K.; Gerdes, L.A.; Cekanaviciute, E.; Jia, X.; Xiao, L.; Xia, Z.; Liu, C.; Klotz, L.; Stauffer, U.; Baranzini, S.E.; et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc. Natl. Acad. Sci. USA 2017, 114, 10719–10724. [Google Scholar] [CrossRef]
- Miyauchi, E.; Kim, S.W.; Suda, W.; Kawasumi, M.; Onawa, S.; Taguchi-Atarashi, N.; Morita, H.; Taylor, T.D.; Hattori, M.; Ohno, H. Gut microorganisms act together to exacerbate inflammation in spinal cords. Nature 2020, 585, 102–106. [Google Scholar] [CrossRef]
- Chen, J.; Wright, K.; Davis, J.M.; Jeraldo, P.; Marietta, E.V.; Murray, J.; Nelson, H.; Matteson, E.L.; Taneja, V. An expansion of rare lineage intestinal microbes characterizes rheumatoid arthritis. Genome Med. 2016, 8, 43. [Google Scholar] [CrossRef] [Green Version]
- Leonard, M.M.; Valitutti, F.; Karathia, H.; Pujolassos, M.; Kenyon, V.; Fanelli, B.; Troisi, J.; Subramanian, P.; Camhi, S.; Colucci, A.; et al. Microbiome signatures of progression toward celiac disease onset in at-risk children in a longitudinal prospective cohort study. Proc. Natl. Acad. Sci. USA 2021, 118, e2020322118. [Google Scholar] [CrossRef] [PubMed]
- Windsor, J.W.; Kaplan, G.G. Evolving Epidemiology of IBD. Curr. Gastroenterol. Rep. 2019, 21, 40. [Google Scholar] [CrossRef] [PubMed]
- Kaser, A.; Zeissig, S.; Blumberg, R.S. Genes and environment: How will our concepts on the pathophysiology of IBD develop in the future? Dig. Dis. 2010, 28, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Knowles, S.R.; Graff, L.A.; Wilding, H.; Hewitt, C.; Keefer, L.; Mikocka-Walus, A. Quality of Life in Inflammatory Bowel Disease: A Systematic Review and Meta-analyses-Part I. Inflamm. Bowel Dis. 2018, 24, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Dorofeyev, A.E.; Vasilenko, I.V.; Rassokhina, O.A.; Kondratiuk, R.B. Mucosal barrier in ulcerative colitis and Crohn’s disease. Gastroenterol. Res. Pract. 2013, 2013, 431231. [Google Scholar] [CrossRef]
- Peterson, D.A.; Frank, D.N.; Pace, N.R.; Gordon, J.I. Metagenomic approaches for defining the pathogenesis of inflammatory bowel diseases. Cell Host Microbe 2008, 3, 417–427. [Google Scholar] [CrossRef]
- Yeshi, K.; Ruscher, R.; Hunter, L.; Daly, N.L.; Loukas, A.; Wangchuk, P. Revisiting Inflammatory Bowel Disease: Pathology, Treatments, Challenges and Emerging Therapeutics Including Drug Leads from Natural Products. J. Clin. Med. 2020, 9, 1273. [Google Scholar] [CrossRef]
- Santana, P.T.; Rosas, S.L.B.; Ribeiro, B.E.; Marinho, Y.; de Souza, H.S.P. Dysbiosis in Inflammatory Bowel Disease: Pathogenic Role and Potential Therapeutic Targets. Int. J. Mol. Sci. 2022, 23, 3464. [Google Scholar] [CrossRef]
- Loftus, E.V., Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2004, 126, 1504–1517. [Google Scholar] [CrossRef]
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The microbiome and inflammatory bowel disease. J. Allergy Clin. Immunol. 2020, 145, 16–27. [Google Scholar] [CrossRef] [Green Version]
- Aniwan, S.; Tremaine, W.J.; Raffals, L.E.; Kane, S.V.; Loftus, E.V., Jr. Antibiotic Use and New-Onset Inflammatory Bowel Disease in Olmsted County, Minnesota: A Population-Based Case-Control Study. J. Crohns Colitis 2018, 12, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Rohr, M.W.; Narasimhulu, C.A.; Rudeski-Rohr, T.A.; Parthasarathy, S. Negative Effects of a High-Fat Diet on Intestinal Permeability: A Review. Adv. Nutr. 2020, 11, 77–91. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the Microbiota in Immunity and Inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Fava, F.; Danese, S. Intestinal microbiota in inflammatory bowel disease: Friend of foe? World J. Gastroenterol. 2011, 17, 557–566. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef]
- Atarashi, K.; Tanoue, T.; Shima, T.; Imaoka, A.; Kuwahara, T.; Momose, Y.; Cheng, G.; Yamasaki, S.; Saito, T.; Ohba, Y.; et al. Induction of colonic regulatory T cells by indigenous Clostridium species. Science 2011, 331, 337–341. [Google Scholar] [CrossRef]
- Watanabe, Y.; Nagai, F.; Morotomi, M. Characterization of Phascolarctobacterium succinatutens sp. nov., an asaccharolytic, succinate-utilizing bacterium isolated from human feces. Appl. Environ. Microbiol. 2012, 78, 511–518. [Google Scholar] [CrossRef]
- Eslick, S.; Thompson, C.; Berthon, B.; Wood, L. Short-chain fatty acids as anti-inflammatory agents in overweight and obesity: A systematic review and meta-analysis. Nutr. Rev. 2022, 80, 838–856. [Google Scholar] [CrossRef]
- Kabeerdoss, J.; Sankaran, V.; Pugazhendhi, S.; Ramakrishna, B.S. Clostridium leptum group bacteria abundance and diversity in the fecal microbiota of patients with inflammatory bowel disease: A case-control study in India. BMC Gastroenterol. 2013, 13, 20. [Google Scholar] [CrossRef]
- Sokol, H.; Pigneur, B.; Watterlot, L.; Lakhdari, O.; Bermúdez-Humarán, L.G.; Gratadoux, J.J.; Blugeon, S.; Bridonneau, C.; Furet, J.P.; Corthier, G.; et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc. Natl. Acad. Sci. USA 2008, 105, 16731–16736. [Google Scholar] [CrossRef]
- Willing, B.P.; Dicksved, J.; Halfvarson, J.; Andersson, A.F.; Lucio, M.; Zheng, Z.; Järnerot, G.; Tysk, C.; Jansson, J.K.; Engstrand, L. A pyrosequencing study in twins shows that gastrointestinal microbial profiles vary with inflammatory bowel disease phenotypes. Gastroenterology 2010, 139, 1844–1854.e1. [Google Scholar] [CrossRef]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Selinger, C.P.; Bell, A.; Cairns, A.; Lockett, M.; Sebastian, S.; Haslam, N. Probiotic VSL#3 prevents antibiotic-associated diarrhoea in a double-blind, randomized, placebo-controlled clinical trial. J. Hosp. Infect. 2013, 84, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.S.; Li, W.B.; Wang, H.Y.; Ma, Y.M.; Zhao, X.H.; Yang, H.; Qian, J.M.; Li, J.N. VSL#3 can prevent ulcerative colitis-associated carcinogenesis in mice. World J. Gastroenterol. 2018, 24, 4254–4262. [Google Scholar] [CrossRef] [PubMed]
- Rossi, G.; Pengo, G.; Caldin, M.; Palumbo Piccionello, A.; Steiner, J.M.; Cohen, N.D.; Jergens, A.E.; Suchodolski, J.S. Comparison of microbiological, histological, and immunomodulatory parameters in response to treatment with either combination therapy with prednisone and metronidazole or probiotic VSL#3 strains in dogs with idiopathic inflammatory bowel disease. PLoS ONE 2014, 9, e94699. [Google Scholar] [CrossRef]
- Uronis, J.M.; Arthur, J.C.; Keku, T.; Fodor, A.; Carroll, I.M.; Cruz, M.L.; Appleyard, C.B.; Jobin, C. Gut microbial diversity is reduced by the probiotic VSL#3 and correlates with decreased TNBS-induced colitis. Inflamm. Bowel Dis. 2011, 17, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Carnero-Gregorio, M.; Molares-Vila, A.; Corbalán-Rivas, A.; Villaverde-Taboada, C.; Rodríguez-Cerdeira, C. Effect of VSL#3 Probiotic in a Patient with Glycogen Storage Disease Type Ia and Irritable Bowel Disease-like Disease. Probiotics Antimicrob. Proteins 2019, 11, 143–149. [Google Scholar] [CrossRef]
- Gionchetti, P.; Rizzello, F.; Helwig, U.; Venturi, A.; Lammers, K.M.; Brigidi, P.; Vitali, B.; Poggioli, G.; Miglioli, M.; Campieri, M. Prophylaxis of pouchitis onset with probiotic therapy: A double-blind, placebo-controlled trial. Gastroenterology 2003, 124, 1202–1209. [Google Scholar] [CrossRef] [PubMed]
- Gionchetti, P.; Rizzello, F.; Venturi, A.; Brigidi, P.; Matteuzzi, D.; Bazzocchi, G.; Poggioli, G.; Miglioli, M.; Campieri, M. Oral bacteriotherapy as maintenance treatment in patients with chronic pouchitis: A double-blind, placebo-controlled trial. Gastroenterology 2000, 119, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Kühbacher, T.; Ott, S.J.; Helwig, U.; Mimura, T.; Rizzello, F.; Kleessen, B.; Gionchetti, P.; Blaut, M.; Campieri, M.; Fölsch, U.R.; et al. Bacterial and fungal microbiota in relation to probiotic therapy (VSL#3) in pouchitis. Gut 2006, 55, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Dang, X.; Xu, M.; Liu, D.; Zhou, D.; Yang, W. Assessing the efficacy and safety of fecal microbiota transplantation and probiotic VSL#3 for active ulcerative colitis: A systematic review and meta-analysis. PLoS ONE 2020, 15, e0228846. [Google Scholar] [CrossRef]
- Quévrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B.; et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2016, 65, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Kruis, W.; Fric, P.; Pokrotnieks, J.; Lukás, M.; Fixa, B.; Kascák, M.; Kamm, M.A.; Weismueller, J.; Beglinger, C.; Stolte, M.; et al. Maintaining remission of ulcerative colitis with the probiotic Escherichia coli Nissle 1917 is as effective as with standard mesalazine. Gut 2004, 53, 1617–1623. [Google Scholar] [CrossRef]
- Dieleman, L.A.; Goerres, M.S.; Arends, A.; Sprengers, D.; Torrice, C.; Hoentjen, F.; Grenther, W.B.; Sartor, R.B. Lactobacillus GG prevents recurrence of colitis in HLA-B27 transgenic rats after antibiotic treatment. Gut 2003, 52, 370–376. [Google Scholar] [CrossRef]
- Li, Q.; Sun, X.; Yu, K.; Lv, J.; Miao, C.; Yang, J.; Wang, S.; Fu, Z.; Sun, Y.; Zhang, H.; et al. Enterobacter ludwigii protects DSS-induced colitis through choline-mediated immune tolerance. Cell Rep. 2022, 40, 111308. [Google Scholar] [CrossRef]
- Hedin, C.R.; McCarthy, N.E.; Louis, P.; Farquharson, F.M.; McCartney, S.; Stagg, A.J.; Lindsay, J.O.; Whelan, K. Prebiotic Fructans Have Greater Impact on Luminal Microbiology and CD3+ T Cells in Healthy Siblings than Patients with Crohn’s Disease: A Pilot Study Investigating the Potential for Primary Prevention of Inflammatory Bowel Disease. Clin. Nutr. 2021, 40, 5009–5019. [Google Scholar] [CrossRef]
- Zhang, X.F.; Guan, X.X.; Tang, Y.J.; Sun, J.F.; Wang, X.K.; Wang, W.D.; Fan, J.M. Clinical Effects and Gut Microbiota Changes of Using Probiotics, Prebiotics or Synbiotics in Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. Eur. J. Nutr. 2021, 60, 2855–2875. [Google Scholar] [CrossRef]
- Colman, R.J.; Rubin, D.T. Fecal microbiota transplantation as therapy for inflammatory bowel disease: A systematic review and meta-analysis. J. Crohns Colitis 2014, 8, 1569–1581. [Google Scholar] [CrossRef] [Green Version]
- Nishida, A.; Imaeda, H.; Ohno, M.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Andoh, A. Efficacy and safety of single fecal microbiota transplantation for Japanese patients with mild to moderately active ulcerative colitis. J. Gastroenterol. 2017, 52, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Goyal, A.; Yeh, A.; Bush, B.R.; Firek, B.A.; Siebold, L.M.; Rogers, M.B.; Kufen, A.D.; Morowitz, M.J. Safety, Clinical Response, and Microbiome Findings Following Fecal Microbiota Transplant in Children With Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 410–421. [Google Scholar] [CrossRef] [PubMed]
- Lewis, J.D.; Sandler, R.S.; Brotherton, C.; Brensinger, C.; Li, H.; Kappelman, M.D.; Daniel, S.G.; Bittinger, K.; Albenberg, L.; Valentine, J.F.; et al. A Randomized Trial Comparing the Specific Carbohydrate Diet to a Mediterranean Diet in Adults With Crohn’s Disease. Gastroenterology 2021, 161, 837–852.e9. [Google Scholar] [CrossRef] [PubMed]
- Burrello, C.; Giuffrè, M.R.; Macandog, A.D.; Diaz-Basabe, A.; Cribiù, F.M.; Lopez, G.; Borgo, F.; Nezi, L.; Caprioli, F.; Vecchi, M.; et al. Fecal Microbiota Transplantation Controls Murine Chronic Intestinal Inflammation by Modulating Immune Cell Functions and Gut Microbiota Composition. Cells 2019, 8, 517. [Google Scholar] [CrossRef]
- Wen, X.; Wang, H.G.; Zhang, M.N.; Zhang, M.H.; Wang, H.; Yang, X.Z. Fecal microbiota transplantation ameliorates experimental colitis via gut microbiota and T-cell modulation. World J. Gastroenterol. 2021, 27, 2834–2849. [Google Scholar] [CrossRef]
- Zhang, W.; Zou, G.; Li, B.; Du, X.; Sun, Z.; Sun, Y.; Jiang, X. Fecal Microbiota Transplantation (FMT) Alleviates Experimental Colitis in Mice by Gut Microbiota Regulation. J. Microbiol. Biotechnol. 2020, 30, 1132–1141. [Google Scholar] [CrossRef]
- Maslowski, K.M.; Vieira, A.T.; Ng, A.; Kranich, J.; Sierro, F.; Yu, D.; Schilter, H.C.; Rolph, M.S.; Mackay, F.; Artis, D.; et al. Regulation of inflammatory responses by gut microbiota and chemoattractant receptor GPR43. Nature 2009, 461, 1282–1286. [Google Scholar] [CrossRef]
- Allegretti, J.R.; Kassam, Z.; Carrellas, M.; Mullish, B.H.; Marchesi, J.R.; Pechlivanis, A.; Smith, M.; Gerardin, Y.; Timberlake, S.; Pratt, D.S.; et al. Fecal Microbiota Transplantation in Patients With Primary Sclerosing Cholangitis: A Pilot Clinical Trial. Am. J. Gastroenterol. 2019, 114, 1071–1079. [Google Scholar] [CrossRef]
- Caruso, R.; Lo, B.C.; Núñez, G. Host-microbiota interactions in inflammatory bowel disease. Nat. Rev. Immunol. 2020, 20, 411–426. [Google Scholar] [CrossRef]
- Schmidt, T.S.B.; Li, S.S.; Maistrenko, O.M.; Akanni, W.; Coelho, L.P.; Dolai, S.; Fullam, A.; Glazek, A.M.; Hercog, R.; Herrema, H.; et al. Drivers and determinants of strain dynamics following fecal microbiota transplantation. Nat. Med. 2022, 28, 1902–1912. [Google Scholar] [CrossRef]
- Paramsothy, S.; Nielsen, S.; Kamm, M.A.; Deshpande, N.P.; Faith, J.J.; Clemente, J.C.; Paramsothy, R.; Walsh, A.J.; van den Bogaerde, J.; Samuel, D.; et al. Specific Bacteria and Metabolites Associated With Response to Fecal Microbiota Transplantation in Patients With Ulcerative Colitis. Gastroenterology 2019, 156, 1440–1454.e2. [Google Scholar] [CrossRef] [PubMed]
- Vaughn, B.P.; Vatanen, T.; Allegretti, J.R.; Bai, A.; Xavier, R.J.; Korzenik, J.; Gevers, D.; Ting, A.; Robson, S.C.; Moss, A.C. Increased Intestinal Microbial Diversity Following Fecal Microbiota Transplant for Active Crohn’s Disease. Inflamm. Bowel Dis. 2016, 22, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Christ, A.; Lauterbach, M.; Latz, E. Western Diet and the Immune System: An Inflammatory Connection. Immunity 2019, 51, 794–811. [Google Scholar] [CrossRef]
- Moszak, M.; Szulińska, M.; Bogdański, P. You Are What You Eat—The Relationship between Diet, Microbiota, and Metabolic Disorders—A Review. Nutrients 2020, 12, 1096. [Google Scholar] [CrossRef] [PubMed]
- Mentella, M.C.; Scaldaferri, F.; Pizzoferrato, M.; Gasbarrini, A.; Miggiano, G.A.D. Nutrition, IBD and Gut Microbiota: A Review. Nutrients 2020, 12, 944. [Google Scholar] [CrossRef]
- Sugihara, K.; Kamada, N. Diet–Microbiota Interactions in Inflammatory Bowel Disease. Nutrients 2021, 13, 1533. [Google Scholar] [CrossRef]
- Tursi, A.; Brandimarte, G. The symptomatic and histologic response to a gluten-free diet in patients with borderline enteropathy. J. Clin. Gastroenterol. 2003, 36, 13–17. [Google Scholar] [CrossRef]
- Adolph, T.E.; Zhang, J. Diet fuelling inflammatory bowel diseases: Preclinical and clinical concepts. Gut 2022, 71, 2574–2586. [Google Scholar] [CrossRef]
- Roberts, C.L.; Keita, A.V.; Duncan, S.H.; O’Kennedy, N.; Söderholm, J.D.; Rhodes, J.M.; Campbell, B.J. Translocation of Crohn’s disease Escherichia coli across M-cells: Contrasting effects of soluble plant fibres and emulsifiers. Gut 2010, 59, 1331–1339. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Koren, O.; Goodrich, J.K.; Poole, A.C.; Srinivasan, S.; Ley, R.E.; Gewirtz, A.T. Dietary emulsifiers impact the mouse gut microbiota promoting colitis and metabolic syndrome. Nature 2015, 519, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Chen, W.D.; Wang, Y.D. Nuclear Receptors: A Bridge Linking the Gut Microbiome and the Host. Mol. Med. 2021, 27, 144. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, Z.; Xu, C.; Kan, S.; Chen, D. Disturbances of the Gut Microbiota and Microbiota-Derived Metabolites in Inflammatory Bowel Disease. Nutrients 2022, 14, 5140. [Google Scholar] [CrossRef] [PubMed]
- Huda-Faujan, N.; Abdulamir, A.S.; Fatimah, A.B.; Anas, O.M.; Shuhaimi, M.; Yazid, A.M.; Loong, Y.Y. The Impact of the Level of the Intestinal Short Chain Fatty Acids in Inflammatory Bowel Disease Patients Versus Healthy Subjects. Open Biochem. J. 2010, 4, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Kumari, R.; Ahuja, V.; Paul, J. Fluctuations in butyrate-producing bacteria in ulcerative colitis patients of North India. World J. Gastroenterol. 2013, 19, 3404–3414. [Google Scholar] [CrossRef]
- Ji, J.; Shu, D.; Zheng, M.; Wang, J.; Luo, C.; Wang, Y.; Guo, F.; Zou, X.; Lv, X.; Li, Y.; et al. Microbial metabolite butyrate facilitates M2 macrophage polarization and function. Sci. Rep. 2016, 6, 24838. [Google Scholar] [CrossRef]
- Bian, X.; Yang, L.; Wu, W.; Lv, L.; Jiang, X.; Wang, Q.; Wu, J.; Li, Y.; Ye, J.; Fang, D.; et al. Pediococcus pentosaceus LI05 alleviates DSS-induced colitis by modulating immunological profiles, the gut microbiota and short-chain fatty acid levels in a mouse model. Microb. Biotechnol. 2020, 13, 1228–1244. [Google Scholar] [CrossRef]
- Gudi, R.; Suber, J.; Brown, R.; Johnson, B.M.; Vasu, C. Pretreatment with Yeast-Derived Complex Dietary Polysaccharides Suppresses Gut Inflammation, Alters the Microbiota Composition, and Increases Immune Regulatory Short-Chain Fatty Acid Production in C57BL/6 Mice. J. Nutr. 2020, 150, 1291–1302. [Google Scholar] [CrossRef]
- Devkota, S.; Wang, Y.; Musch, M.W.; Leone, V.; Fehlner-Peach, H.; Nadimpalli, A.; Antonopoulos, D.A.; Jabri, B.; Chang, E.B. Dietary-fat-induced taurocholic acid promotes pathobiont expansion and colitis in Il10-/- mice. Nature 2012, 487, 104–108. [Google Scholar] [CrossRef]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Duboc, H.; Rajca, S.; Rainteau, D.; Benarous, D.; Maubert, M.A.; Quervain, E.; Thomas, G.; Barbu, V.; Humbert, L.; Despras, G.; et al. Connecting Dysbiosis, Bile-Acid Dysmetabolism and Gut Inflammation in Inflammatory Bowel Diseases. Gut 2013, 62, 531–539. [Google Scholar] [CrossRef]
- Paik, D.; Yao, L.; Zhang, Y.; Bae, S.; D’Agostino, G.D.; Zhang, M.; Kim, E.; Franzosa, E.A.; Avila-Pacheco, J.; Bisanz, J.E.; et al. Human Gut Bacteria Produce TH17-Modulating Bile Acid Metabolites. Nature 2022, 603, 907–912. [Google Scholar] [CrossRef] [PubMed]
- Nikolaus, S.; Schulte, B.; Al-Massad, N.; Thieme, F.; Schulte, D.M.; Bethge, J.; Rehman, A.; Tran, F.; Aden, K.; Häsler, R.; et al. Increased Tryptophan Metabolism Is Associated With Activity of Inflammatory Bowel Diseases. Gastroenterology 2017, 153, 1504–1516.e2. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, M.; Sugimoto, S.; Mitsuyama, K.; Kobayashi, T.; Yoshimura, N.; Ohi, H.; Tanaka, S.; Andoh, A.; Ohmiya, N.; Saigusa, K.; et al. Efficacy of Indigo Naturalis in a Multicenter Randomized Controlled Trial of Patients With Ulcerative Colitis. Gastroenterology 2018, 154, 935–947. [Google Scholar] [CrossRef] [PubMed]
- Mayne, C.G.; Williams, C.B. Induced and natural regulatory T cells in the development of inflammatory bowel disease. Inflamm. Bowel Dis. 2013, 19, 1772–1788. [Google Scholar] [CrossRef] [PubMed]
- Lê, A.; Mantel, M.; Marchix, J.; Bodinier, M.; Jan, G.; Rolli-Derkinderen, M. Inflammatory bowel disease therapeutic strategies by modulation of the microbiota: How and when to introduce pre-, pro-, syn-, or postbiotics? Am. J. Physiol. Gastrointest. Liver Physiol. 2022, 323, G523–G553. [Google Scholar] [CrossRef]
- Aguilar-Toalá, J.E.; Garcia-Varela, R.; Garcia, H.S.; Mata-Haro, V.; González-Córdova, A.F.; Vallejo-Cordoba, B.; Hernández-Mendoza, A. Postbiotics: An Evolving Term within the Functional Foods Field. Trends Food Sci. Technol. 2018, 75, 105–114. [Google Scholar] [CrossRef]
- Lebeer, S.; Vanderleyden, J.; De Keersmaecker, S.C. Host interactions of probiotic bacterial surface molecules: Comparison with commensals and pathogens. Nat. Rev. Microbiol. 2010, 8, 171–184. [Google Scholar] [CrossRef]
- Hynönen, U.; Palva, A. Lactobacillus surface layer proteins: Structure, function and applications. Appl. Microbiol. Biotechnol. 2013, 97, 5225–5243. [Google Scholar] [CrossRef]
- Klotz, C.; Goh, Y.J.; O’Flaherty, S.; Barrangou, R. S-layer associated proteins contribute to the adhesive and immunomodulatory properties of Lactobacillus acidophilus NCFM. BMC Microbiol. 2020, 20, 248. [Google Scholar] [CrossRef]
- Lathrop, S.K.; Bloom, S.M.; Rao, S.M.; Nutsch, K.; Lio, C.W.; Santacruz, N.; Peterson, D.A.; Stappenbeck, T.S.; Hsieh, C.S. Peripheral education of the immune system by colonic commensal microbiota. Nature 2011, 478, 250–254. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Zhang, L.; Xu, S.; Pan, J.; Zhang, Q.; Lu, R. Surface-Layer Protein from Lactobacillus acidophilus NCFM Inhibits Lipopolysaccharide-Induced Inflammation through MAPK and NF-κB Signaling Pathways in RAW264.7 Cells. J. Agric. Food Chem. 2018, 66, 7655–7662. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Xu, P.; Wu, Z.; Pan, D. Anti-Inflammatory Activity of Surface Layer Protein SlpA of Lactobacillus Acidophilus CICC 6074 in LPS-Induced RAW 264.7 Cells and DSS-Induced Mice Colitis. J. Funct. Foods 2018, 51, 16–27. [Google Scholar] [CrossRef]
- Ge, Y.; Gong, M.; Zadeh, M.; Li, J.; Abbott, J.R.; Li, W.; Morel, L.; Sonon, R.; Supekar, N.T.; Azadi, P.; et al. Regulating colonic dendritic cells by commensal glycosylated large surface layer protein A to sustain gut homeostasis against pathogenic inflammation. Mucosal. Immunol. 2020, 13, 34–46. [Google Scholar] [CrossRef]
- Aldars-García, L.; Chaparro, M.; Gisbert, J.P. Systematic Review: The Gut Microbiome and Its Potential Clinical Application in Inflammatory Bowel Disease. Microorganisms 2021, 9, 977. [Google Scholar] [CrossRef]
- Sudhakar, P.; Andrighetti, T.; Verstockt, S.; Caenepeel, C.; Ferrante, M.; Sabino, J.; Verstockt, B.; Vermeire, S. Integrated Analysis of Microbe-Host Interactions in Crohn’s Disease Reveals Potential Mechanisms of Microbial Proteins on Host Gene Expression. iScience 2022, 25, 103963. [Google Scholar] [CrossRef]
- Aleksandrova, K.; Romero-Mosquera, B.; Hernandez, V. Diet, Gut Microbiome and Epigenetics: Emerging Links with Inflammatory Bowel Diseases and Prospects for Management and Prevention. Nutrients 2017, 9, 962. [Google Scholar] [CrossRef]
- Khorsand, B.; Asadzadeh Aghdaei, H.; Nazemalhosseini-Mojarad, E.; Nadalian, B.; Nadalian, B.; Houri, H. Overrepresentation of Enterobacteriaceae and Escherichia Coli Is the Major Gut Microbiome Signature in Crohn’s Disease and Ulcerative Colitis; A Comprehensive Metagenomic Analysis of IBDMDB Datasets. Front. Cell. Infect. Microbiol. 2022, 12, 1015890. [Google Scholar] [CrossRef]
Figure 1.
The gut microbiota is altered in IBD patients compared to healthy subjects. Up-arrows indicate an increase, down-arrows indicate a decrease, and horizon-arrows indicate no changes.
Figure 1.
The gut microbiota is altered in IBD patients compared to healthy subjects. Up-arrows indicate an increase, down-arrows indicate a decrease, and horizon-arrows indicate no changes.

Figure 2.
Improved intestinal environment alleviates IBD. Modulating gut microbiota, such as probiotics, fecal microbiota transplantation (FMT), and gut microbial metabolites, can alleviate IBD symptoms by suppressing gastrointestinal inflammation.
Figure 2.
Improved intestinal environment alleviates IBD. Modulating gut microbiota, such as probiotics, fecal microbiota transplantation (FMT), and gut microbial metabolites, can alleviate IBD symptoms by suppressing gastrointestinal inflammation.

{kind=link}
{kind=link}
Table 1.
Autoimmune diseases and gut microbiota.
| Diseases | Gut Microbiota | References |
|---|---|---|
| Type 1 diabetes | Lactobacillaceae ↓ Rikenellaceae ↓ Porphoromadaceae ↓ | [15] |
| Ruminococcaceae ↓ Ruminococcus ↓ | [16] | |
| Multiple sclerosis | Lactobacillaceae ↓ Bacteroidaceae ↓ Prevotellaceae ↓ | [17] |
| Akkermansia ↑ | [18,19] | |
| Erysipelotrichaceae ↑ Lactobacillus reuteri ↑ | ||
| Rheumatoid arthritis | Collinsella ↑ Faecalibacterium ↑ Eggerthella ↑ | [20] |
| Celiac disease | Dialister invisus ↑ Parabacteroides ↑ Lachnospiraceae bacterium ↑ | [21] |
↓; indicates the decrease. ↑; indicates the increase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Haneishi, Y.; Furuya, Y.; Hasegawa, M.; Picarelli, A.; Rossi, M.; Miyamoto, J. Inflammatory Bowel Diseases and Gut Microbiota. Int. J. Mol. Sci. 2023, 24, 3817. https://doi.org/10.3390/ijms24043817
AMA Style
Haneishi Y, Furuya Y, Hasegawa M, Picarelli A, Rossi M, Miyamoto J. Inflammatory Bowel Diseases and Gut Microbiota. International Journal of Molecular Sciences. 2023; 24(4):3817. https://doi.org/10.3390/ijms24043817
Chicago/Turabian StyleHaneishi, Yuri, Yuma Furuya, Mayu Hasegawa, Antonio Picarelli, Mauro Rossi, and Junki Miyamoto. 2023. "Inflammatory Bowel Diseases and Gut Microbiota" International Journal of Molecular Sciences 24, no. 4: 3817. https://doi.org/10.3390/ijms24043817
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.