Variation in Phenolic Compounds Content and Antioxidant Activity of Different Plant Organs from Rumex crispus L. and Rumex obtusifolius L. at Different Growth Stages




Abstract
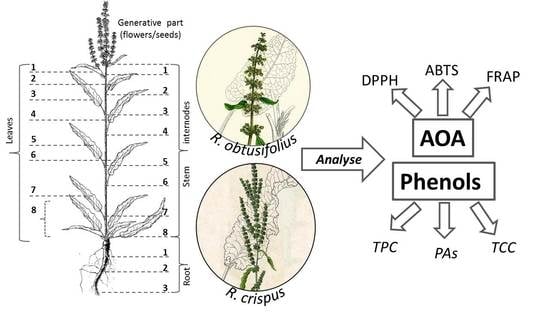
:
1. Introduction
2. Materials and Methods
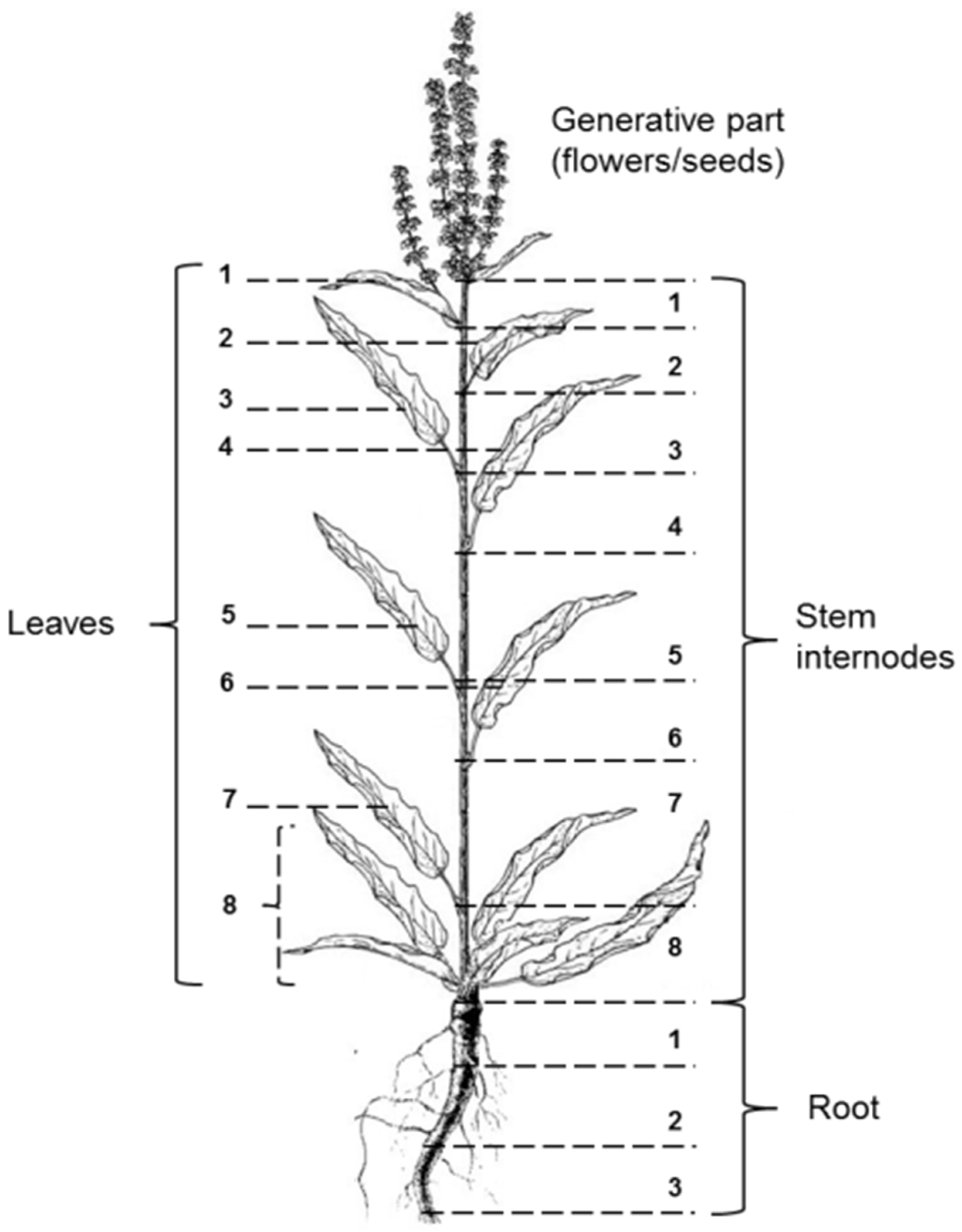
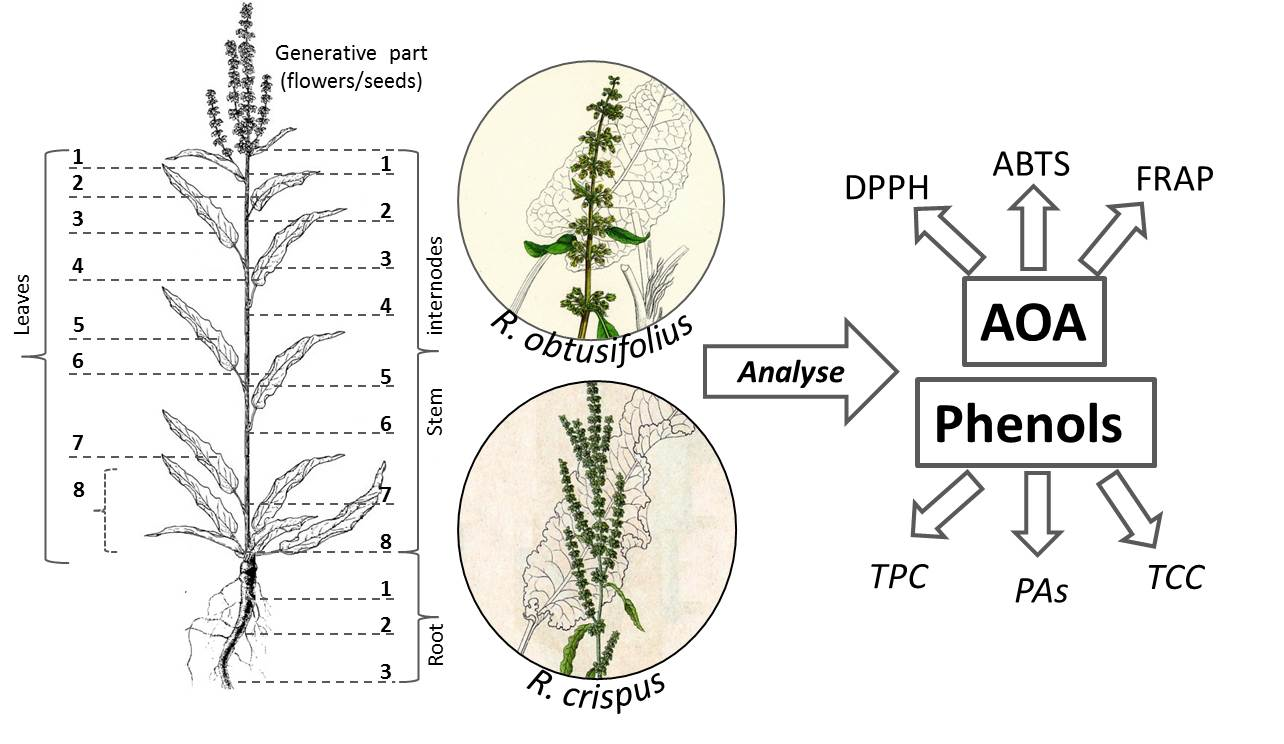
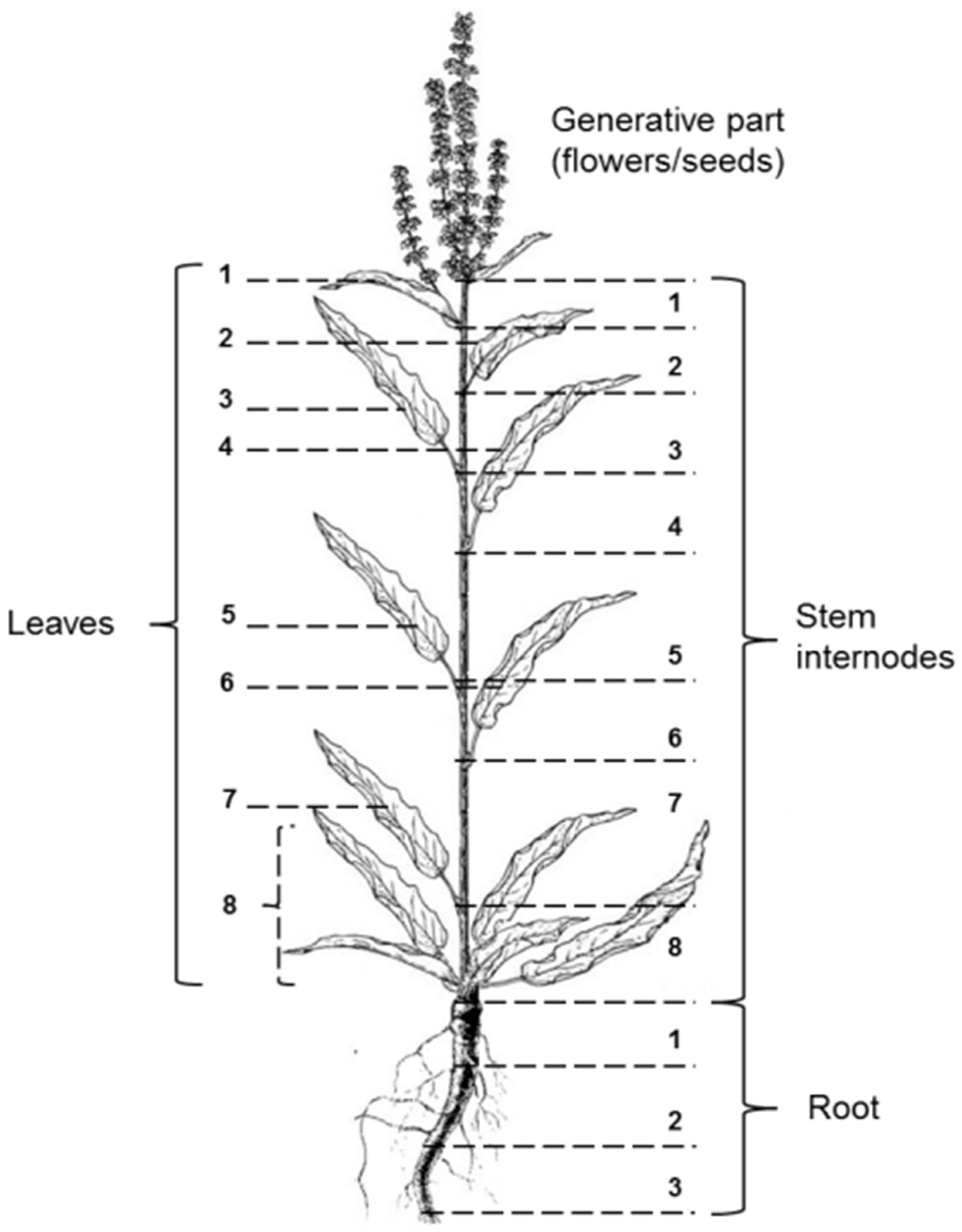
2.1. Plant Material
2.2. Plant Analysis
2.3. Total Catechins Content (TCC)
2.4. Total Proanthocyanidins (PAs) Content
2.5. Total Phenolic Content (TPC)
2.6. Total Antioxidant Activity (AOA)
2.7. Statistical Analysis
3. Results
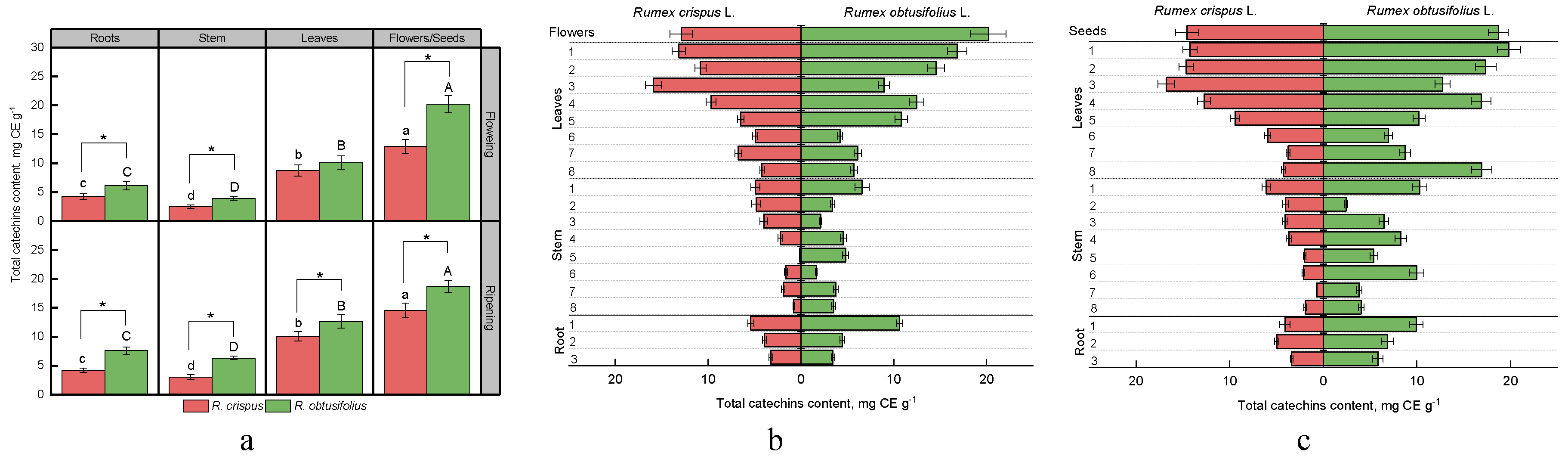
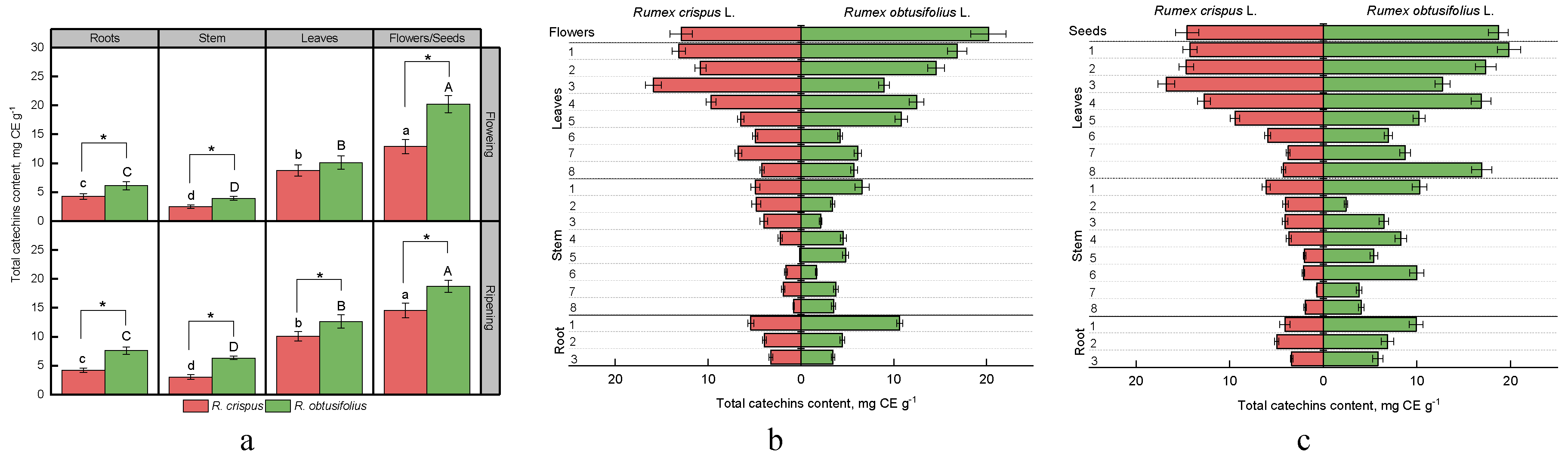
3.1. Total Catechins Content
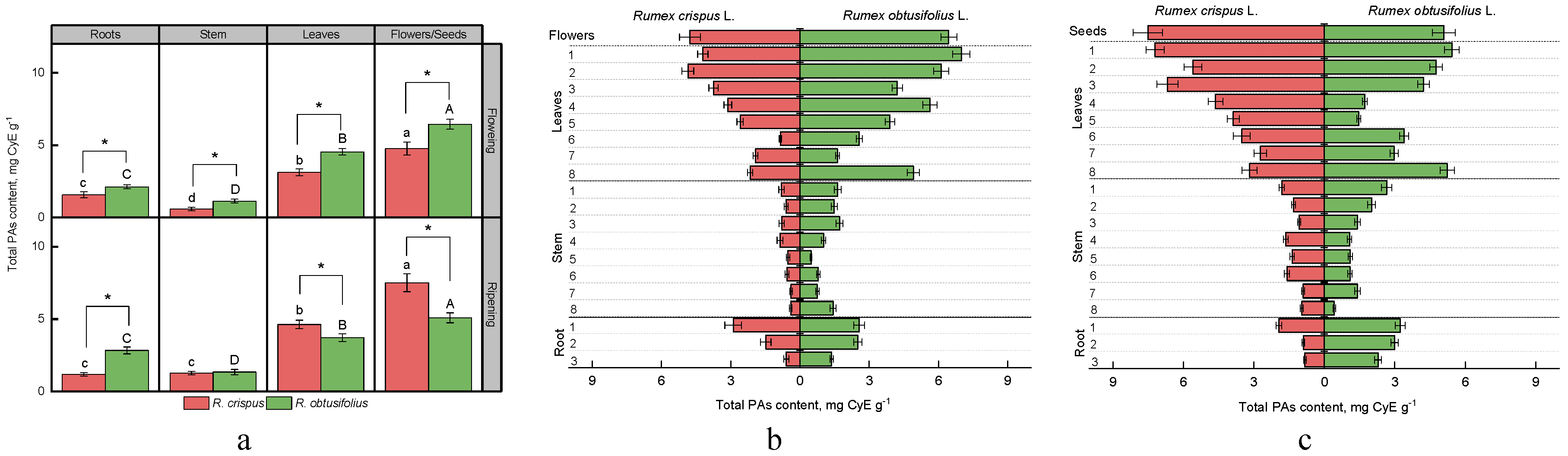
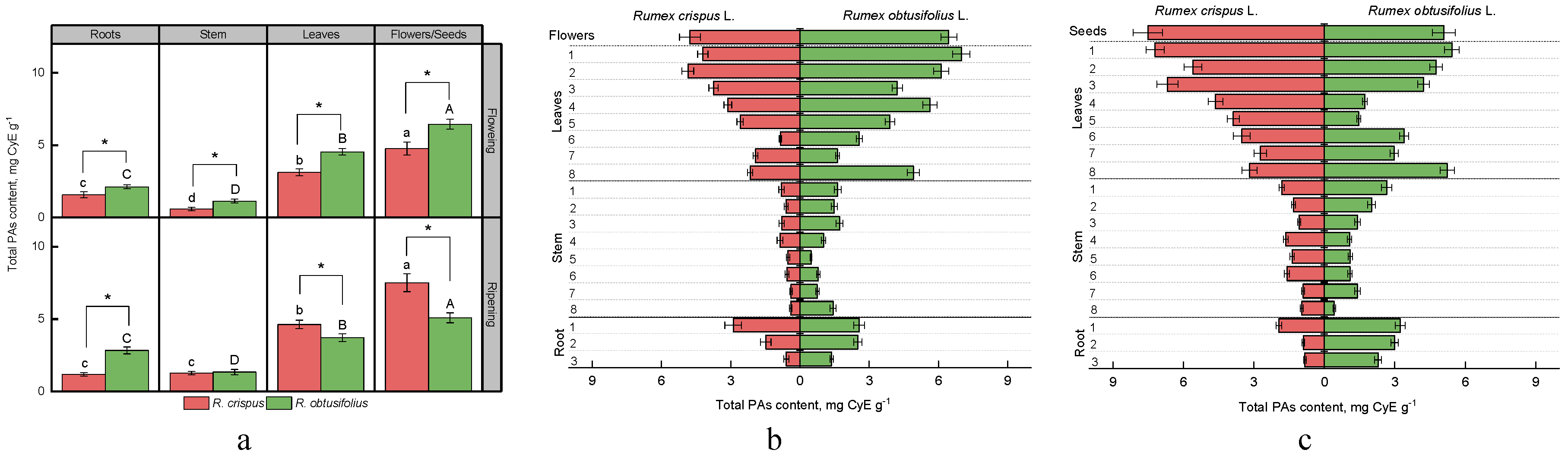
3.2. Total Proanthocyanidins Content (PAs)
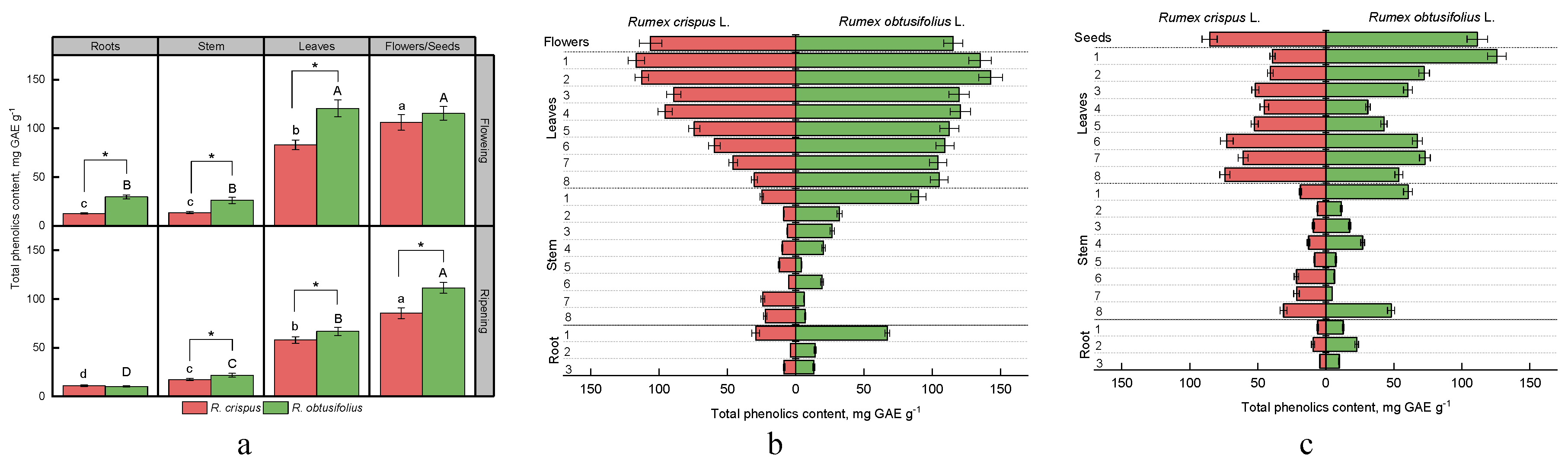
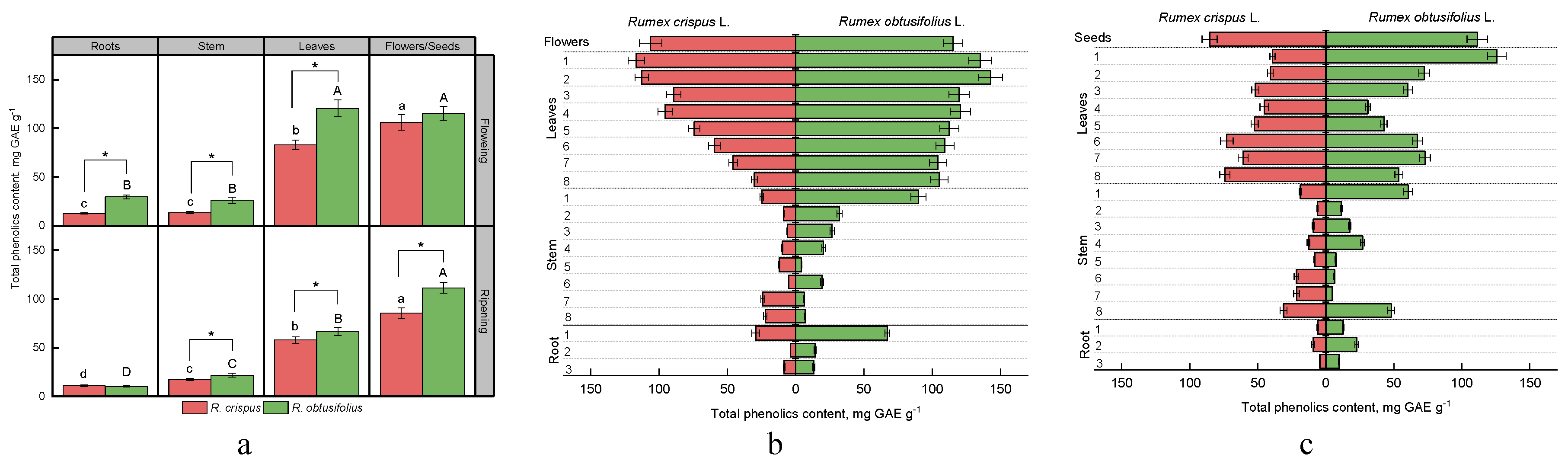
3.3. Total Phenolic Content
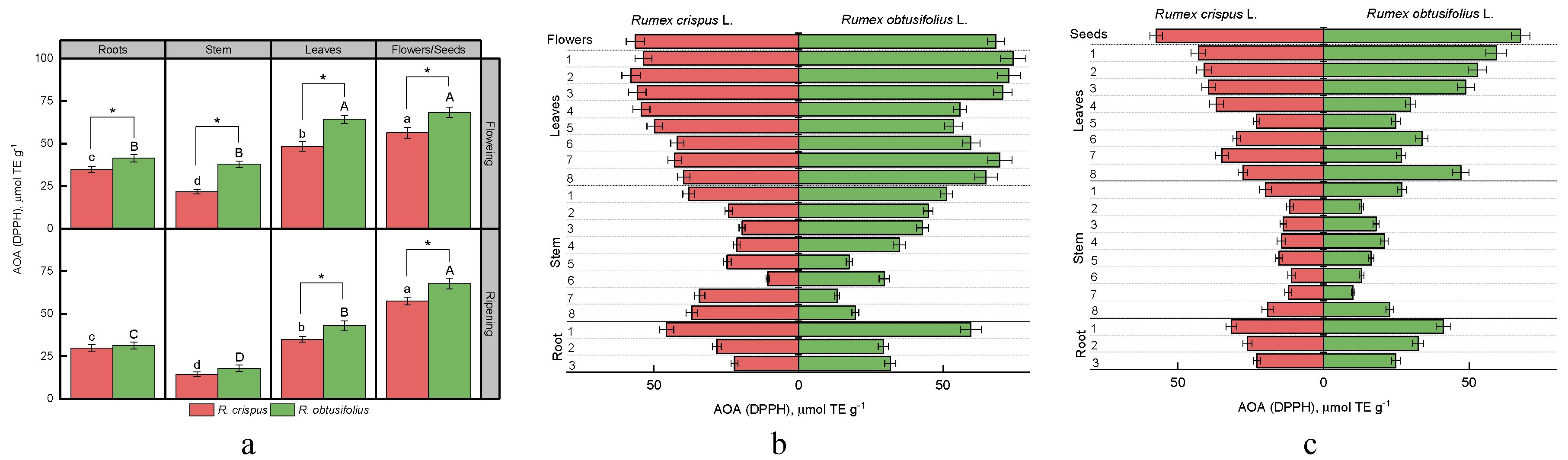
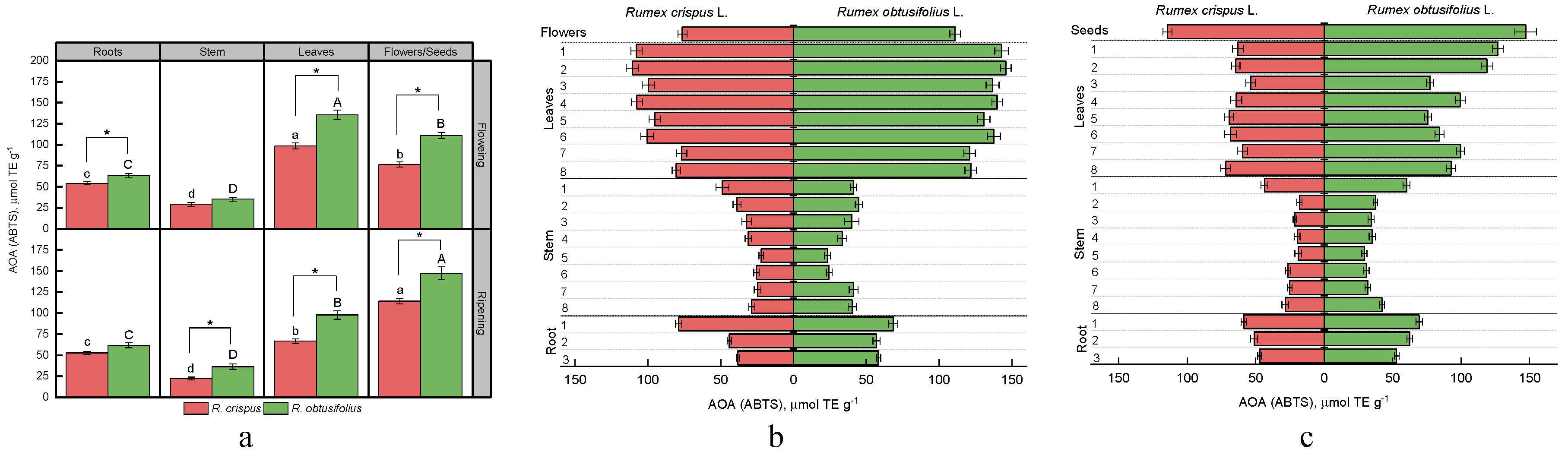
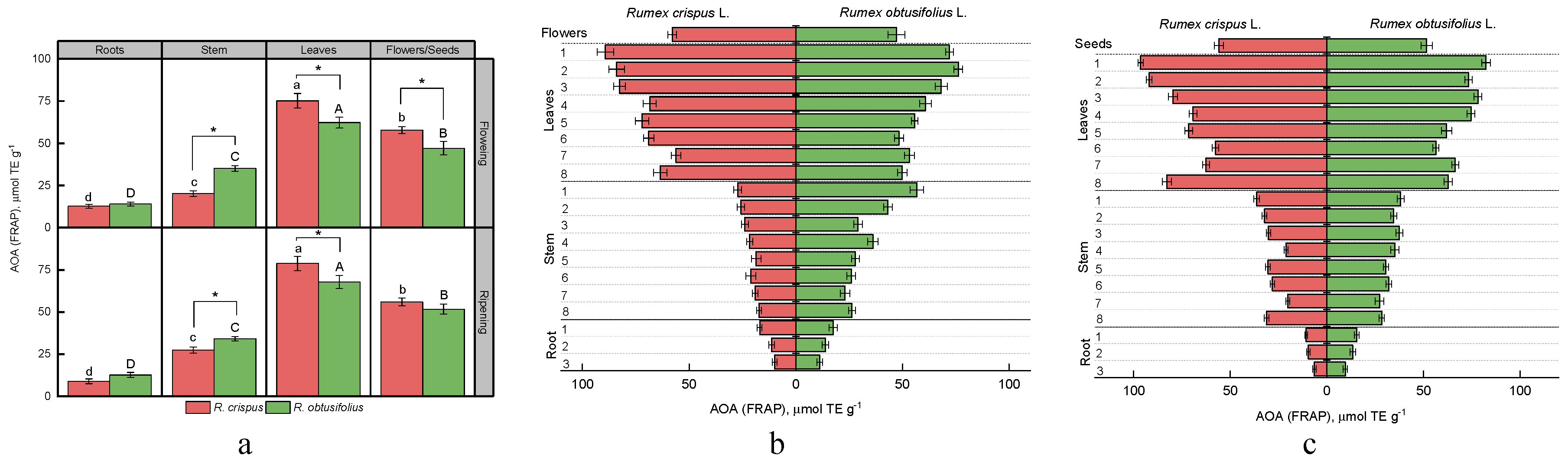
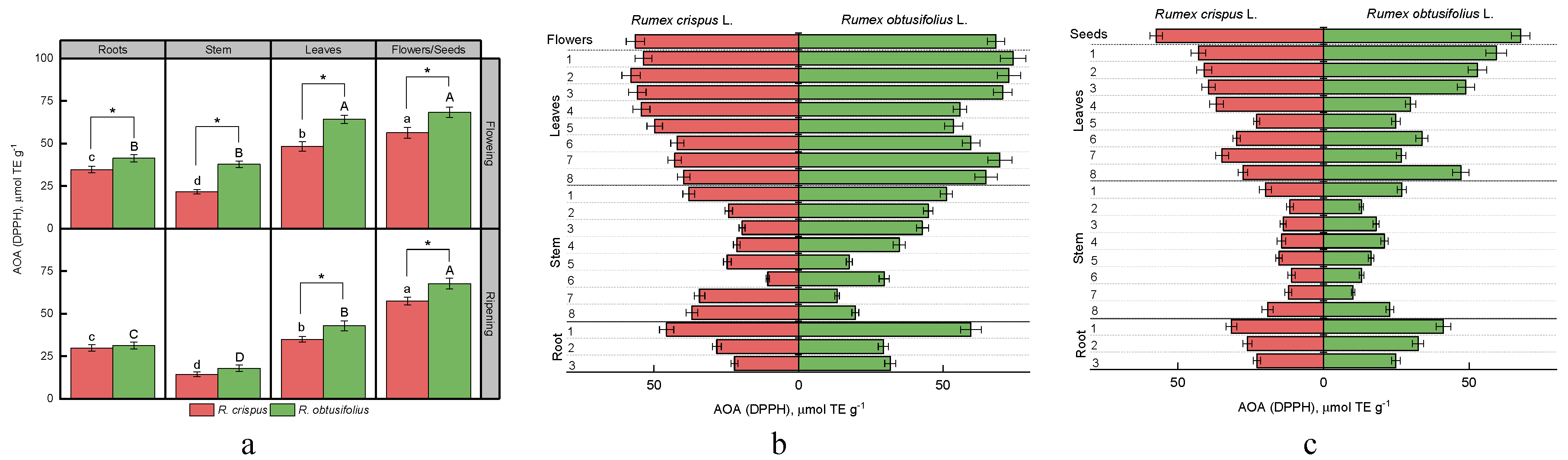
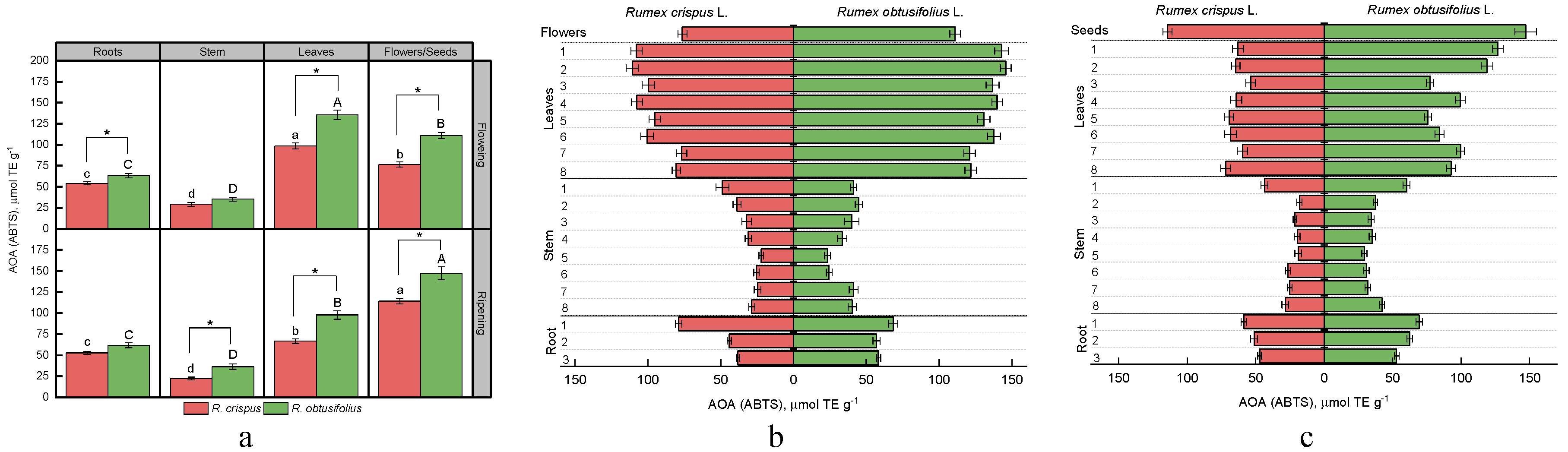
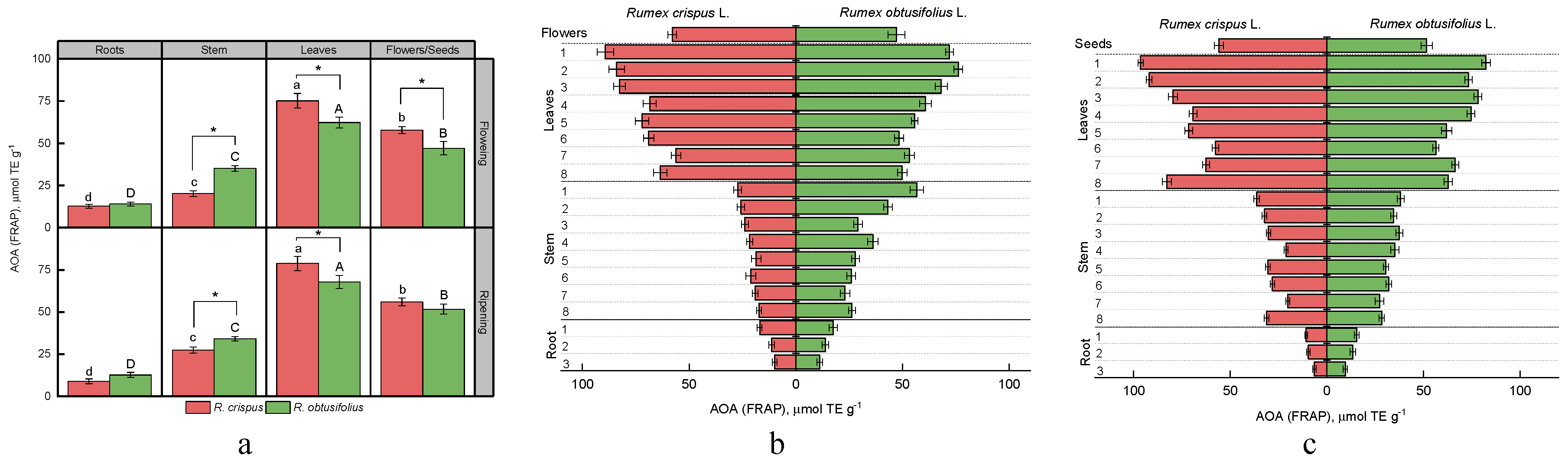
3.4. Total Antioxidant Activity (AOA)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lima, G.P.P.; Vianello, F.; Corrêa, C.R.; Campos, R.A.D.S.; Borguini, M.G. Polyphenols in fruits and vegetables and its effect on human health. Food Nutr. Sci. 2014, 5, 1065–1082. [Google Scholar] [CrossRef]
- Valdés, L.; Cuervo, A.; Salazar, N.; Ruas-Madiedo, P.; Gueimonde, M.; González, S. The relationship between phenolic compounds from diet and microbiota: Impact on human health. Food Funct. 2015, 6, 2424–2439. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 1374. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G. The role of polyphenols in modern nutrition. Nutr. Bull. 2017, 42, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Belščak-Cvitanović, A.; Durgo, K.; Huđek, A.; Bačun-Družina, V.; Komes, D. Overview of polyphenols and their properties. In Polyphenols: Properties, Recovery, and Applications; Galanakis, C.M., Ed.; Woodhead Publishing: Sawston, UK, 2018; pp. 3–44. [Google Scholar]
- Ameer, K.; Shahbaz, H.M.; Kwon, J.H. Green extraction methods for polyphenols from plant matrices and their byproducts: A review. Compr. Rev. Food Sci. Food Saf. 2018, 16, 295–315. [Google Scholar] [CrossRef]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxid. Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef]
- Umeno, A.; Horie, M.; Murotomi, K.; Nakajima, Y.; Yoshida, Y. Antioxidative and antidiabetic effects of natural polyphenols and isoflavones. Molecules 2016, 21, 708. [Google Scholar] [CrossRef]
- Zhang, H.; Tsao, R. Dietary polyphenols, oxidative stress and antioxidant and anti-inflammatory effects. Curr. Opin. Food Sci. 2016, 8, 33–42. [Google Scholar] [CrossRef]
- Martins, N.; Barros, L.; Santos-Buelga, C.; Henriques, M.; Silva, S.; Ferreira, I.C. Evaluation of bioactive properties and phenolic compounds in different extracts prepared from Salvia officinalis L. Food Chem. 2015, 170, 378–385. [Google Scholar] [CrossRef]
- Xu, Y.; Burton, S.; Kim, C.; Sismour, E. Phenolic compounds, antioxidant, and antibacterial properties of pomace extracts from four Virginia-grown grape varieties. Food Sci. Nutr. 2016, 4, 125–133. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Fu, S.G.; Wang, S.Y.; Yang, D.J.; Wu, Y.H.S.; Chen, Y.C. Effects of a natural antioxidant, polyphenol-rich rosemary (Rosmarinus officinalis L.) extract, on lipid stability of plant-derived omega-3 fatty-acid rich oil. Food Sci. Technol. 2018, 89, 210–216. [Google Scholar] [CrossRef]
- Chobot, V.; Hadacek, F.; Bachmann, G.; Weckwerth, W.; Kubicova, L. Pro-and antioxidant activity of three selected flavan type flavonoids: Catechin, eriodictyol and taxifolin. Int. J. Mol. Sci. 2016, 17, 1986. [Google Scholar] [CrossRef] [PubMed]
- Amorati, R.; Baschieri, A.; Cowden, A.; Valgimigli, L. The antioxidant activity of quercetin in water solution. Biomimetics 2017, 2, 9. [Google Scholar] [CrossRef] [PubMed]
- Vignault, A.; González-Centeno, M.R.; Pascual, O.; Gombau, J.; Jourdes, M.; Moine, V.; Teissedre, P.L. Chemical characterization, antioxidant properties and oxygen consumption rate of 36 commercial oenological tannins in a model wine solution. Food Chem. 2018, 268, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Wu, G.; Johnson, S.K.; Blanchard, C.L. Characterization of phenolic compounds and antioxidant activity in sorghum grains. J. Cereal Sci. 2018, 84, 103–111. [Google Scholar] [CrossRef]
- Mut-Salud, N.; Álvarez, P.J.; Garrido, J.M.; Carrasco, E.; Aránega, A.; Rodríguez-Serrano, F. Antioxidant intake and antitumor therapy: Toward nutritional recommendations for optimal results. Oxid. Med. Cell. Longev. 2016, 2016, 6719534. [Google Scholar] [CrossRef]
- Khurana, R.K.; Jain, A.; Jain, A.; Sharma, T.; Singh, B.; Kesharwani, P. Administration of antioxidants in cancer: Debate of the decade. Drug Discov. Today 2018, 23, 763–770. [Google Scholar] [CrossRef]
- Matthew, S.; Cullen, A.J.; Cullen, J.J. Treating pancreatic cancer: More antioxidants more problems? Expert Rev. Gastroenterol. Hepatol. 2018, 12, 849–851. [Google Scholar]
- Chupakhina, G.N.; Maslennikov, P.V.; Skrypnik, L.N.; Chupakhina, N.Y.; Poltavskaya, R.L.; Feduraev, P.V. The influence of the Baltic regional conditions on the accumulation of water-soluble antioxidants in plants. Russ. Chem. Bull. 2014, 63, 1946–1953. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Blekkenhorst, L.; Sim, M.; Bondonno, C.; Bondonno, N.; Ward, N.; Prince, R.; Hodgson, J. Cardiovascular health benefits of specific vegetable types: A narrative review. Nutrients 2018, 10, 595. [Google Scholar] [CrossRef] [PubMed]
- Morris, M.C.; Wang, Y.; Barnes, L.L.; Bennett, D.A.; Dawson-Hughes, B.; Booth, S.L. Nutrients and bioactives in green leafy vegetables and cognitive decline: Prospective study. Neurology 2018, 90, e214–e222. [Google Scholar] [CrossRef] [PubMed]
- Olas, B. Berry Phenolic Antioxidants—Implications for Human Health? Front. Pharmacol. 2018, 9, 78. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.X.; Dai, H.Q.; Fu, H.A.; Dong, X.P.; Adebayo, A.H.; Zhang, L.X.; Cheng, Y.X. Bioactive compounds from Rumex plants. Phytochem. Lett. 2010, 3, 181–184. [Google Scholar] [CrossRef]
- Vasas, A.; Orbán-Gyapai, O.; Hohmann, J. The Genus Rumex: Review of traditional uses, phytochemistry and pharmacology. J. Ethnopharmacol. 2015, 175, 198–228. [Google Scholar] [CrossRef] [PubMed]
- Bélanger, J.; Balakrishna, M.; Latha, P.; Katumalla, S.; Johns, T. Contribution of selected wild and cultivated leafy vegetables from South India to lutein and β-carotene intake. Asia Pac. J. Clin. Nutr. 2010, 19, 417–424. [Google Scholar] [PubMed]
- Maksimović, Z.; Kovačević, N.; Lakušić, B.; Ćebović, T. Antioxidant activity of yellow dock (Rumex crispus L., Polygonaceae) fruit extract. Phytother. Res. 2011, 25, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Van Evert, F.K.; Samsom, J.; Polder, G.; Vijn, M.; Dooren, H.J.V.; Lamaker, A.; Lotz, L.A. A robot to detect and control broad-leaved dock (Rumex obtusifolius L.) in grassland. J. Field Robot. 2011, 28, 264–277. [Google Scholar] [CrossRef]
- Cavers, P.B.; Harper, J.L. Rumex obtusifolius L. and R. crispus L. J. Ecol. 1964, 52, 737–766. [Google Scholar]
- Coruh, I.; Gormez, A.; Ercisli, S.; Sengul, M. Total phenolic content, antioxidant, and antibacterial activity of Rumex crispus grown wild in Turkey. Pharm. Biol. 2008, 46, 634–638. [Google Scholar] [CrossRef]
- Wegiera, M.; Grabarczyk, P.; Baraniak, B.; Smolarz, H. Antiradical properties of extracts from roots, leaves and fruits of six Rumex L. species. Acta Biol. Crac. Ser. Bot. 2011, 53, 125–131. [Google Scholar] [CrossRef]
- Balog, K.; Svirčev, E.; Lesjak, M.; Orcic, D.; Beara, I.; Francišković, M.; Simin, N. Phenolic profiling of Rumex L. species by means of the LC-MS/MS. Planta Med. 2012, 78, PL12. [Google Scholar] [CrossRef]
- Mishra, A.P.; Sharifi-Rad, M.; Shariati, M.A.; Mabkhot, Y.N.; Al-Showiman, S.S.; Rauf, A.; Sharifi-Rad, J. Bioactive compounds and health benefits of edible Rumex species—A review. Cell. Mol. Biol. 2018, 64, 27–34. [Google Scholar] [CrossRef]
- Harshaw, D.; Nahar, L.; Vadla, B.; Sarker, S.D. Bioactivity of Rumex obtusifolius (Polygonaceae). Arch. Biol. Sci. 2010, 62, 387–392. [Google Scholar] [CrossRef]
- Aghajanyan, A.; Nikoyan, A.; Trchounian, A. Biochemical Activity and Hypoglycemic Effects of Rumex obtusifolius L. Seeds Used in Armenian Traditional Medicine. BioMed Res. Int. 2018, 2018, 4526352. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Lattanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef]
- Jiang, X.; Liu, Y.; Li, W.; Zhao, L.; Meng, F.; Wang, Y.; Gao, L. Tissue-specific, development-dependent phenolic compounds accumulation profile and gene expression pattern in tea plant (Camellia sinensis). PLoS ONE 2013, 8, e62315. [Google Scholar] [CrossRef]
- Jeelani, S.M.; Farooq, U.; Gupta, A.P.; Lattoo, S.K. Phytochemical evaluation of major bioactive compounds in different cytotypes of five species of Rumex L. Ind. Crop. Prod. 2017, 109, 897–904. [Google Scholar] [CrossRef]
- Munakata, R.; Larbat, R.; Duriot, L.; Olry, A.; Gavira, C.; Mignard, B.; Bourgaud, F. Polyphenols from Plant Roots. Recent Adv. Polyphen. Res. 2019, 6, 207–237. [Google Scholar]
- Gubanov, I.A.; Kiseleva, K.V.; Novikov, V.S.; Tikhomirov, V.N. Illyustrirovannyy Opredelitel’ Rasteniy Sredney Rossii; Tovarishchestvo Nauchnykh Izdaniy KMK Publ.: Moscow, Russia, 2003; pp. 113–115. [Google Scholar]
- He, Q.; Yao, K.; Jia, D.; Fan, H.; Liao, X.; Shi, B. Determination of total catechins in tea extracts by HPLC and spectrophotometry. Nat. Prod. Res. 2009, 23, 93–100. [Google Scholar] [CrossRef]
- Maksimović, Z.; Malenčić, Đ.; Kovačević, N. Polyphenol contents and antioxidant activity of Maydis stigma extracts. Bioresour. Technol. 2005, 96, 873–877. [Google Scholar] [CrossRef] [PubMed]
- Padhi, E.M.; Liu, R.; Hernandez, M.; Tsao, R.; Ramdath, D.D. Total polyphenol content, carotenoid, tocopherol and fatty acid composition of commonly consumed Canadian pulses and their contribution to antioxidant activity. J. Funct. Foods 2017, 38, 602–611. [Google Scholar] [CrossRef]
- Tõnutare, T. Possibilities to Affect Antioxidant Properties of Strawberries and Some Methodical Aspects in Their Determination. Ph.D. Thesis, Eesti Maaülikool, Tartu, Estonia, 2015. [Google Scholar]
- Taneva, I.; Petkova, N.; Dimov, I.; Ivanov, I.; Denev, P. Characterization of rose hip (Rosa canina L.) fruits extracts and evaluation of their in vitro antioxidant activity. J. Pharmacogn. Phytochem. 2016, 5, 35–38. [Google Scholar]
- Fedurayev, P.V.; Chupakhina, G.N.; Skrypnik, L.N. Dinamika nakopleniya katekhinov shchavelem kurchavym (Rumex crispus L.) superprodutsentom fenol’nykh soyedineniy proantotsianidinovogo ryada. Khimiya Rastitel’nogo Syr’ya 2011, 4, 205–208. [Google Scholar]
- Feduraev, P.V.; Skrypnik, L.N.; Maslennikov, P.V.; Tchoupakhina, G.N.; Tacenko, N.A. The accumulation features of phenolic compounds in some species of genus Rumex L. Khimiya Rastitel’nogo Syr’ya 2017, 3, 123–130. [Google Scholar]
- Sellami, I.H.; Maamouri, E.; Chahed, T.; Wannes, W.A.; Kchouk, M.E.; Marzouk, B. Effect of growth stage on the content and composition of the essential oil and phenolic fraction of sweet marjoram (Origanum majorana L.). Ind. Crop. Prod. 2009, 30, 395–402. [Google Scholar] [CrossRef]
- Ammar, I.; Ennouri, M.; Khemakhem, B.; Yangui, T.; Attia, H. Variation in chemical composition and biological activities of two species of Opuntia flowers at four stages of flowering. Ind. Crop. Prod. 2012, 37, 34–40. [Google Scholar] [CrossRef]
- Fernando, I.D.N.S.; Abeysinghe, D.C.; Dharmadasa, R.M. Determination of phenolic contents and antioxidant capacity of different parts of Withania somnifera (L.) Dunal. from three different growth stages. Ind. Crop. Prod. 2013, 50, 537–539. [Google Scholar] [CrossRef]
- Andreotti, C.; Costa, G.; Treutter, D. Composition of phenolic compounds in pear leaves as affected by genetics, ontogenesis and the environment. Sci. Hortic. 2006, 109, 130–137. [Google Scholar] [CrossRef]
- Jakopic, J.; Stampar, F.; Veberic, R. Daily dynamics of sugar and phenol contents in apple fruitlets during June drop. Notulae Botanicae Horti Agrobotanici Cluj-Napoca 2018, 46, 75–81. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Plant aging increases oxidative stress in chloroplasts. Planta 2002, 214, 608–615. [Google Scholar] [PubMed]
- Blum-Silva, C.H.; Chaves, V.C.; Schenkel, E.P.; Coelho, G.C.; Reginatto, F.H. The influence of leaf age on methylxanthines, total phenolic content, and free radical scavenging capacity of Ilex paraguariensis aqueous extracts. Rev. Bras. Farmacogn. 2015, 25, 1–6. [Google Scholar] [CrossRef]
- Bowers, M.D.; Stamp, N.E. Effects of plant age, genotype, and herbivory on Plantago performance and chemistry. Ecology 1993, 74, 1778–1791. [Google Scholar] [CrossRef]
- Doan, A.T.; Ervin, G.; Felton, G. Temporal effects on jasmonate induction of anti-herbivore defense in Physalis angulata: Seasonal and ontogenetic gradients. Biochem. Syst. Ecol. 2004, 32, 117–126. [Google Scholar] [CrossRef]
- Höft, M.; Verpoorte, R.; Beck, E. Leaf alkaloid contents of Tabernaemontana pachysiphon as influenced by endogenous and environmental factors in the natural habitat. Planta Med. 1998, 64, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, T. Diversity and variability of plant secondary metabolism: A mechanistic view. In Proceedings of the 9th International Symposium on Insect-Plant Relationships; Springer: Dordrecht, The Netherlands, 1996; pp. 177–188. [Google Scholar]
- Wilkinson, T.L.; Douglas, A.E. Phloem amino acids and the host plant range of the polyphagous aphid, Aphis fabae. Entomol. Exp. Appl. 2003, 106, 103–113. [Google Scholar] [CrossRef]
- Weibull, J.; Ronquist, F.; Brishammar, S. Free amino acid composition of leaf exudates and phloem sap: A comparative study in oats and barley. Plant Physiol. 1990, 92, 222–226. [Google Scholar] [CrossRef]
- Kretz, A. Zucker, Cyclite und organische Säuren des Cambialsaftes von Pinus silvestris L., Picea abies Karst. und Abies alba Mill. Planta 1973, 110, 1–14. [Google Scholar] [CrossRef]
- De Micco, V.; Aronne, G. Root structure of Rumex scutatus growing on slopes. IAWA J. 2010, 31, 13–28. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Factor Level | TCC | PAs | TPC | AOA (DPPH) | AOA (ABTS) | AOA (FRAP) |
|---|---|---|---|---|---|---|---|
| Main effects | |||||||
| Species (Sp) | R. crispus | 3.12 b | 7.5 b | 48.4 b | 37.2 b | 64.3 b | 42.1 a |
| R. obtusifolius | 3.43 a | 10.7 a | 62.7 a | 46.4 a | 86.0 a | 40.7 a | |
| Plant part (P) | Roots | 1.93 c | 5.5 c | 15.9 c | 34.3 c | 58.0 c | 12.1 d |
| Stem | 1.09 d | 3.9 c | 19.8 c | 22.9 d | 30.8 d | 29.2 c | |
| Leaves | 4.00 b | 10.4 b | 82.1 b | 47.5 b | 99.6 b | 71.1 a | |
| Flowers/Seeds | 5.95 a | 16.6 a | 104.6 a | 62.5 a | 112.2 a | 53.2 b | |
| Growth stage (GS) | Flowering | 3.03 b | 8.6 a | 63.4 a | 46.6 a | 75.4 a | 40.6 a |
| Ripening | 3.45 a | 9.6 a | 47.7 b | 37.0 b | 74.9 a | 42.2 a | |
| Significance | |||||||
| Sp | 0.046 * | <0.001 * | <0.001 * | 0.024 * | <0.001 * | 0.086 ns | |
| P | <0.001 * | 0.028 * | 0.035 * | <0.001 * | <0.001 * | <0.001 * | |
| GS | <0.001 * | 0.112 ns | <0.001 * | <0.001 * | 0.142 ns | 0.054 ns | |
| Sp*P | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | <0.001 * | |
| Sp*GS | <0.001 * | <0.001 * | 0.008 * | <0.001 * | <0.001 * | 0.032 * | |
| P*GS | <0.001 * | 0.263 ns | <0.001 * | <0.001 * | <0.001 * | <0.001 * | |
| Sp*P*GS | <0.001 * | <0.001 * | 0.018 * | 0.020 * | <0.001 * | 0.046 * |
| Parameters | TCC | PAs | TPC | AOA (DPPH) | AOA (ABTS) | AOA (FRAP) |
|---|---|---|---|---|---|---|
| TCC | 1.0000 | |||||
| PAs | 0.6020 (p < 0.0001) | 1.0000 | ||||
| TPC | 0.6343 (p < 0.0001) | 0.4894 (p < 0.0001) | 1.0000 | |||
| AOA (DPPH) | 0.6255 (p < 0.0001) | 0.4396 (p < 0.0001) | 0.8789 (p < 0.0001) | 1.0000 | ||
| AOA (ABTS) | 0.6859 (p < 0.0001) | 0.3867 (p < 0.0001) | 0.8741 (p < 0.0001) | 0.8125 (p < 0.0001) | 1.0000 | |
| AOA (FRAP) | 0.6803 (p < 0.0001) | 0.6701 (p < 0.0001) | 0.7032 (p < 0.0001) | 0.5523 (p < 0.0001) | 0.6381 (p < 0.0001) | 1.0000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Feduraev, P.; Chupakhina, G.; Maslennikov, P.; Tacenko, N.; Skrypnik, L. Variation in Phenolic Compounds Content and Antioxidant Activity of Different Plant Organs from Rumex crispus L. and Rumex obtusifolius L. at Different Growth Stages. Antioxidants 2019, 8, 237. https://doi.org/10.3390/antiox8070237
Feduraev P, Chupakhina G, Maslennikov P, Tacenko N, Skrypnik L. Variation in Phenolic Compounds Content and Antioxidant Activity of Different Plant Organs from Rumex crispus L. and Rumex obtusifolius L. at Different Growth Stages. Antioxidants. 2019; 8(7):237. https://doi.org/10.3390/antiox8070237
Chicago/Turabian StyleFeduraev, Pavel, Galina Chupakhina, Pavel Maslennikov, Natalia Tacenko, and Liubov Skrypnik. 2019. "Variation in Phenolic Compounds Content and Antioxidant Activity of Different Plant Organs from Rumex crispus L. and Rumex obtusifolius L. at Different Growth Stages" Antioxidants 8, no. 7: 237. https://doi.org/10.3390/antiox8070237
APA StyleFeduraev, P., Chupakhina, G., Maslennikov, P., Tacenko, N., & Skrypnik, L. (2019). Variation in Phenolic Compounds Content and Antioxidant Activity of Different Plant Organs from Rumex crispus L. and Rumex obtusifolius L. at Different Growth Stages. Antioxidants, 8(7), 237. https://doi.org/10.3390/antiox8070237