
Increased Dietary Protein to Energy Ratio in Pre-Weaning Lambs Increases Average Daily Gain and Cortical Bone Thickness in the Tibia

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animal Management
2.2. Slaughter
2.3. Peripheral Quantitative Computed Tomography (pQCT)
2.4. Statistical Analysis
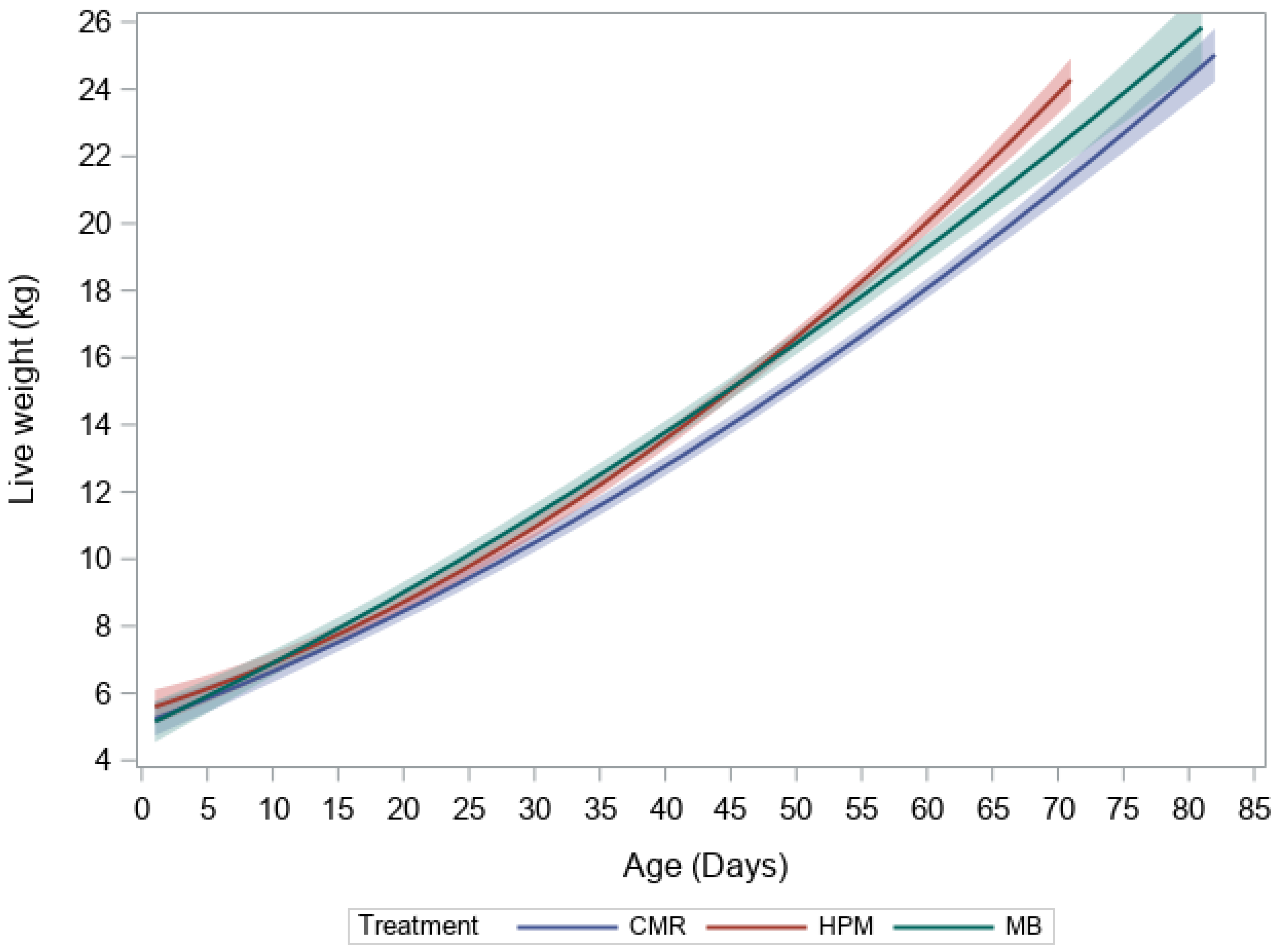
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wallace, T.C. Optimizing dietary protein for lifelong bone health: A paradox unraveled. Nutr. Today 2019, 54, 107–115. [Google Scholar] [CrossRef]
- Schüler, R.; Markova, M.; Osterhoff, M.A.; Arafat, A.; Pivovarova, O.; Machann, J.; Hierholzer, J.; Hornemann, S.; Rohn, S.; Pfeiffer, A.F. Similar dietary regulation of IGF-1-and IGF-binding proteins by animal and plant protein in subjects with type 2 diabetes. Eur. J. Nutr. 2021, 60, 3499–3504. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Bone “mass” and the “mechanostat”: A proposal. Anat. Rec. 1987, 219, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tagliaferri, C.; Wittrant, Y.; Davicco, M.-J.; Walrand, S.; Coxam, V. Muscle and bone, two interconnected tissues. Ageing Res. Rev. 2015, 21, 55–70. [Google Scholar] [PubMed]
- Turner, J.; Bulsara, M.; McDermott, B.; Byrne, G.; Prince, R.; Forbes, D. Predictors of low bone density in young adolescent females with anorexia nervosa and other dieting disorders. Int. J. Eat. Disord. 2001, 30, 245–251. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.K.; Grinspoon, S.K.; Ciampa, J.; Hier, J.; Herzog, D.; Klibanski, A. Medical Findings in Outpatients With Anorexia Nervosa. Arch. Intern. Med. 2005, 165, 561–566. [Google Scholar] [CrossRef]
- Gibson, M.; Dittmer, K.; Hickson, R.; Back, P.; Wehrle-Martinez, A.; Rogers, C. The Mid-Diaphysis Is a Poor Predictor of Humeral Fracture Risk Indicating That Predisposing Factors Are Recent. Ruminants 2021, 1, 23–30. [Google Scholar] [CrossRef]
- Wehrle-Martinez, A.; Lawrence, K.; Back, P.J.; Rogers, C.W.; Gibson, M.; Dittmer, K.E. Osteoporosis is the cause of spontaneous humeral fracture in dairy cows from New Zealand. Vet. Pathol. 2022; ahead of print. [Google Scholar]
- Wehrle-Martinez, A.; Naffa, R.; Back, P.; Rogers, C.W.; Lawrence, K.; Loo, T.; Sutherland-Smith, A.; Dittmer, K. Novel Assessment of Collagen and Its Crosslink Content in the Humerus from Primiparous Dairy Cows with Spontaneous Humeral Fractures Due to Osteoporosis from New Zealand. Biology 2022, 11, 1387. [Google Scholar] [CrossRef]
- Rickets in heifers grazing fodder beet. Vet. Rec. 2022, 190, 407–410. [CrossRef]
- Bones in Protein Deficiency. Nutr. Rev. 1963, 21, 242–244. [CrossRef]
- Stewart, R.J.C.; Platt, B.S. Dietary protein deficiency and growth and mineralisation of bones of pig. In Proceedings of the Nutrition Society; CAB International: Wallingford, UK, 1961; Volume 20. [Google Scholar]
- Cabrera, D.; Wolber, F.M.; Dittmer, K.; Rogers, C.; Ridler, A.; Aberdein, D.; Parkinson, T.; Chambers, P.; Fraser, K.; Roy, N.C. Glucocorticoids affect bone mineral density and bone remodelling in OVX sheep: A pilot study. Bone Rep. 2018, 9, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Nicholson, P.; Wang, Q.; Alén, M.; Cheng, S. Bone and muscle development during puberty in girls: A seven-year longitudinal study. J. Bone Miner. Res. 2009, 24, 1693–1698. [Google Scholar] [CrossRef] [PubMed]
- Frølich, J.; Hansen, S.; Winkler, L.A.-D.; Andresen, A.K.; Hermann, A.P.; Støving, R.K. The role of body weight on bone in anorexia nervosa: A HR-pQCT study. Calcif. Tissue Int. 2017, 101, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Gibson, M.J.; Rogers, C.W.; Pettigrew, E.J.; Pain, S.J.; Dittmer, K.E.; Herath, H.M.G.P.; Back, P.J. The Effect of Artificial Rearing on Live Weight Gain and Bone Morphology of the Tibia in Lambs Prior to Weaning. Ruminants 2022, 2, 101–111. [Google Scholar] [CrossRef]
- Danso, A.; Morel, P.; Kenyon, P.; Blair, H. Effect of different feeding regimens on energy and protein utilization and partitioning for maintenance and growth in pre-weaned lambs reared artificially. J. Anim. Sci. 2016, 94, 5359–5371. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Khan, M.; Lee, W.; Yang, S.; Kim, S.; Ki, K.; Kim, H.; Ha, J.; Choi, Y. Influence of equalizing the gross composition of milk replacer to that of whole milk on the performance of Holstein calves. J. Anim. Sci. 2009, 87, 1129–1137. [Google Scholar] [CrossRef]
- Herath, H.; Pain, S.; Kenyon, P.; Blair, H.; Morel, P. Effect of dietary protein to energy ratio of milk replacer on growth and body composition of pre-weaned lambs reared artificially. Anim. Feed Sci. Technol. 2020, 264, 114478. [Google Scholar] [CrossRef]
- Gibson, M.; Dittmer, K.; Hickson, R.; Back, P.; Rogers, C. Bone Morphology and Strength in the Mid-Diaphysis of the Humerus and Metacarpus in Dairy Calves Prior to Weaning. Animals 2020, 10, 1422. [Google Scholar] [CrossRef]
- Moallem, U.; Werner, D.; Lehrer, H.; Zachut, M.; Livshitz, L.; Yakoby, S.; Shamay, A. Long-term effects of ad libitum whole milk prior to weaning and prepubertal protein supplementation on skeletal growth rate and first-lactation milk production. J. Dairy Sci. 2010, 93, 2639–2650. [Google Scholar] [CrossRef]
- Vatanparast, H.; Bailey, D.A.; Baxter-Jones, A.D.; Whiting, S.J. The effects of dietary protein on bone mineral mass in young adults may be modulated by adolescent calcium intake. J. Nutr. 2007, 137, 2674–2679. [Google Scholar] [CrossRef]
- Yayha, Z.; Millward, D.J. Dietary protein and the regulation of long-bone and muscle growth in the rat. Clin. Sci. 1994, 87, 213–224. [Google Scholar] [CrossRef] [PubMed]
- Mays, S.; Ives, R.; Brickley, M. The effects of socioeconomic status on endochondral and appositional bone growth, and acquisition of cortical bone in children from 19th century Birmingham, England. Am. J. Phys. Anthropol. Off. Publ. Am. Assoc. Phys. Anthropol. 2009, 140, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Cooper, C.; Cawley, M.; Bhalla, A.; Egger, P.; Ring, F.; Morton, L.; Barker, D. Childhood growth, physical activity, and peak bone mass in women. J. Bone Miner. Res. 1995, 10, 940–947. [Google Scholar] [CrossRef] [PubMed]
- Harvey, N.C.; Robinson, S.M.; Crozier, S.R.; Marriott, L.D.; Gale, C.R.; Cole, Z.A.; Inskip, H.M.; Godfrey, K.M.; Cooper, C. Breast-feeding and adherence to infant feeding guidelines do not influence bone mass at age 4 years. Br. J. Nutr. 2009, 102, 915–920. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Pooled (Mean ± SE) | Treatment (Mean ± SE) | ||||
|---|---|---|---|---|---|
| CMR | HPM | MB | p-Value | ||
| n= | 27 | 10 | 9 | 8 | |
| Birth weight (kg) | 5.9 ± 0.2 | 5.9 ± 0.3 | 5.9 ± 0.3 | 5.9 ± 0.3 | 0.99 |
| Age (days) at slaughter | 66.6 ± 1.4 | 70.2 ± 2.2 | 65.4 ± 1.4 | 63.4 ± 3.1 | 0.11 |
| Hot carcass weight (kg) | 10.7 ± 0.1 | 10.6 ± 0.1 | 10.6 ± 0.1 | 10.9 ± 0.2 | 0.28 |
| Average daily gain (g/day) | 253.2 ± 4.6 | 236.0 ± 6.9 a | 261.0 ± 5.4 b | 267.0 ± 7.4 b | 0.01 |
| Tibia length (mm) | 162.0 ± 0.8 | 164.0 ± 0.9 | 161.0 ± 1.7 | 162.0 ± 1.3 | 0.37 |
| Pooled Values (Mean ± SE) | Treatment (Mean ± SE) | |||||
|---|---|---|---|---|---|---|
| CMR | HPM | MB | p-Value | |||
| pQCT parameter | 27 | 10 | 9 | 8 | ||
| Bone | Periosteal Circumference (mm) | 46.3 ± 0.3 | 46.2 ± 0.7 | 46.9 ± 0.5 | 45.8 ± 0.5 | 0.47 |
| Endosteal Circumference (mm) | 24.6 ± 0.5 | 25.4 ± 0.8 | 24.8 ± 0.7 | 23.4 ± 1.1 | 0.24 | |
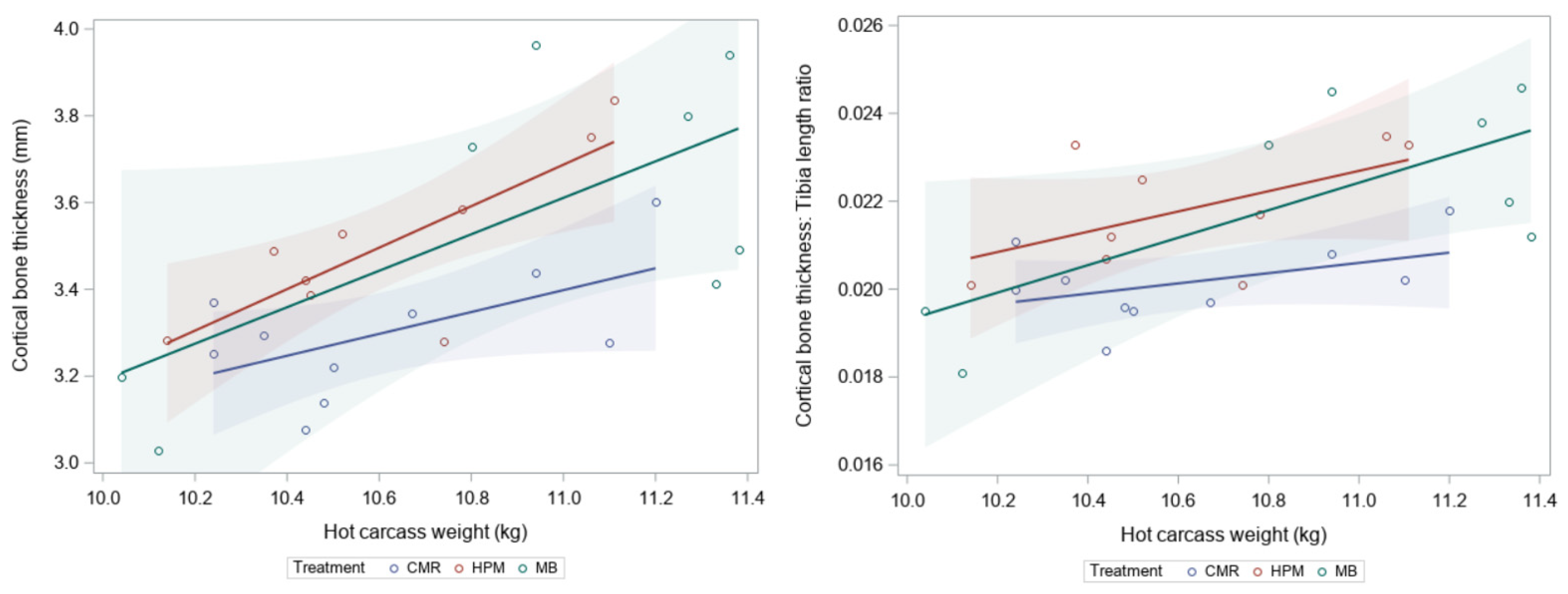
| Cortical bone thickness (mm) | 3.45 ± 0.05 | 3.3 ± 0.05 a | 3.5 ± 0.06 ab | 3.6 ± 0.1 b | 0.05 | |
| Total bone area (mm2) | 170.7 ± 2.6 | 169.9 ± 5.3 | 174.9 ± 4.0 | 167.0 ± 3.4 | 0.48 | |
| Total bone content (mg/mm) | 134.2 ± 1.8 | 130.3 ± 3.0 | 138.1 ± 3.0 | 134.9 ± 3.2 | 0.07 | |
| Total bone mineral density (mg/cm3) | 788.8 ± 10.8 | 769.5 ± 14.8 | 791.0 ± 16.8 | 810.5 ± 24.7 | 0.32 | |
| Cortical bone area (mm2) | 122.0 ± 1.5 | 118.1 ± 2.6 | 125.6 ± 2.4 | 122.9 ± 2.5 | 0.11 | |
| Cortical bone mineral density (mg/cm3) | 1087.4 ± 4.5 | 1089.8 ± 8.8 | 1086.4 ± 8.9 | 1085.4 ± 4.9 | 0.92 | |
| Stress–strain index (SSI) (mm3) | 435.2 ± 9.6 | 431.6 ± 13.1 | 457.4 ± 18.7 | 415.0 ± 17.0 | 0.21 | |
| Cortical thickness: Tibia length | 0.021 ± 0.000 | 0.020 ± 0.000 a | 0.022 ± 0.000 b | 0.022 ± 0.001 b | 0.03 | |
| Periosteal circumference: Tibia length | 0.29 ± 0.0 | 0.28 ± 0.01 | 0.29 ± 0.00 | 0.28 ± 0.00 | 0.25 | |
| Muscle | Total muscle area (mm2) | 824.2 ± 26.6 | 802.6 ± 49.4 | 869.6 ± 46.7 | 800.1 ± 41.0 | 0.50 |
| Muscle | Total bone content: Muscle area | 0.17 ± 0.01 | 0.17 ± 0.01 | 0.16 ± 0.01 | 0.17 ± 0.01 | 0.77 |
| bone ratios | Stress–strain index: Muscle area | 0.54 ± 0.02 | 0.55 ± 0.04 | 0.53 ± 0.01 | 0.53 ± 0.03 | 0.27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gibson, M.J.; Adams, B.R.; Back, P.J.; Dittmer, K.E.; Herath, H.M.G.P.; Pain, S.J.; Kenyon, P.R.; Morel, P.C.H.; Blair, H.T.; Rogers, C.W. Increased Dietary Protein to Energy Ratio in Pre-Weaning Lambs Increases Average Daily Gain and Cortical Bone Thickness in the Tibia. Ruminants 2022, 2, 478-486. https://doi.org/10.3390/ruminants2040034
Gibson MJ, Adams BR, Back PJ, Dittmer KE, Herath HMGP, Pain SJ, Kenyon PR, Morel PCH, Blair HT, Rogers CW. Increased Dietary Protein to Energy Ratio in Pre-Weaning Lambs Increases Average Daily Gain and Cortical Bone Thickness in the Tibia. Ruminants. 2022; 2(4):478-486. https://doi.org/10.3390/ruminants2040034
Chicago/Turabian StyleGibson, Michaela J., Brooke R. Adams, Penny J. Back, Keren E. Dittmer, Hitihamy M. G. P. Herath, Sarah J. Pain, Paul R. Kenyon, Patrick C. H. Morel, Hugh T. Blair, and Chris W. Rogers. 2022. "Increased Dietary Protein to Energy Ratio in Pre-Weaning Lambs Increases Average Daily Gain and Cortical Bone Thickness in the Tibia" Ruminants 2, no. 4: 478-486. https://doi.org/10.3390/ruminants2040034
APA StyleGibson, M. J., Adams, B. R., Back, P. J., Dittmer, K. E., Herath, H. M. G. P., Pain, S. J., Kenyon, P. R., Morel, P. C. H., Blair, H. T., & Rogers, C. W. (2022). Increased Dietary Protein to Energy Ratio in Pre-Weaning Lambs Increases Average Daily Gain and Cortical Bone Thickness in the Tibia. Ruminants, 2(4), 478-486. https://doi.org/10.3390/ruminants2040034