Operational Protocols for the Use of Drones in Marine Animal Research

 , ,
, ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
Abstract
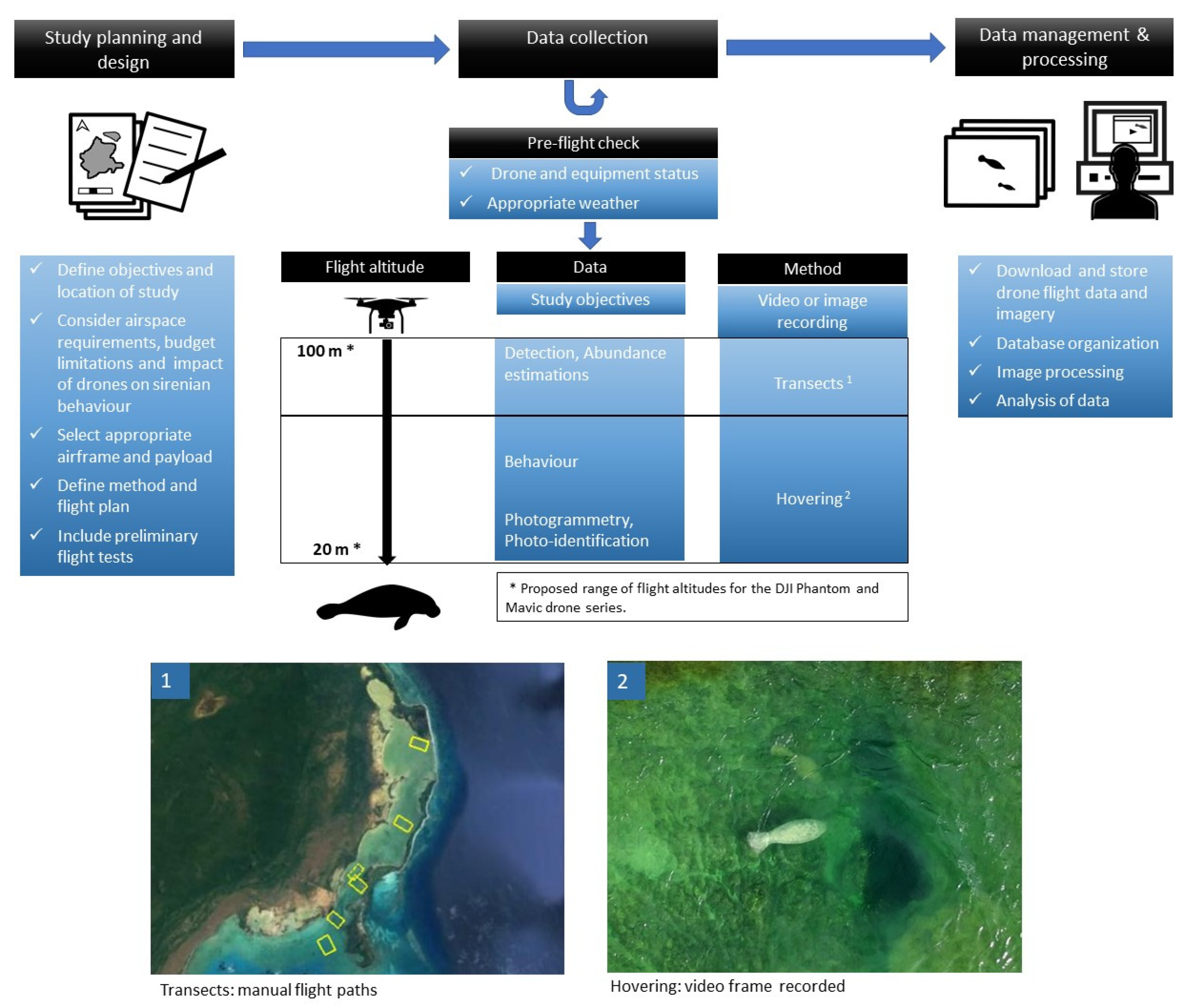
:1. Overview
1.1. Different Drone Platforms Used in Marine Research
1.2. Payloads Available to Drones for Marine Research
1.3. Photogrammetry and Structure-from-Motion from Drones
1.4. Current Limitations of Drone Technology
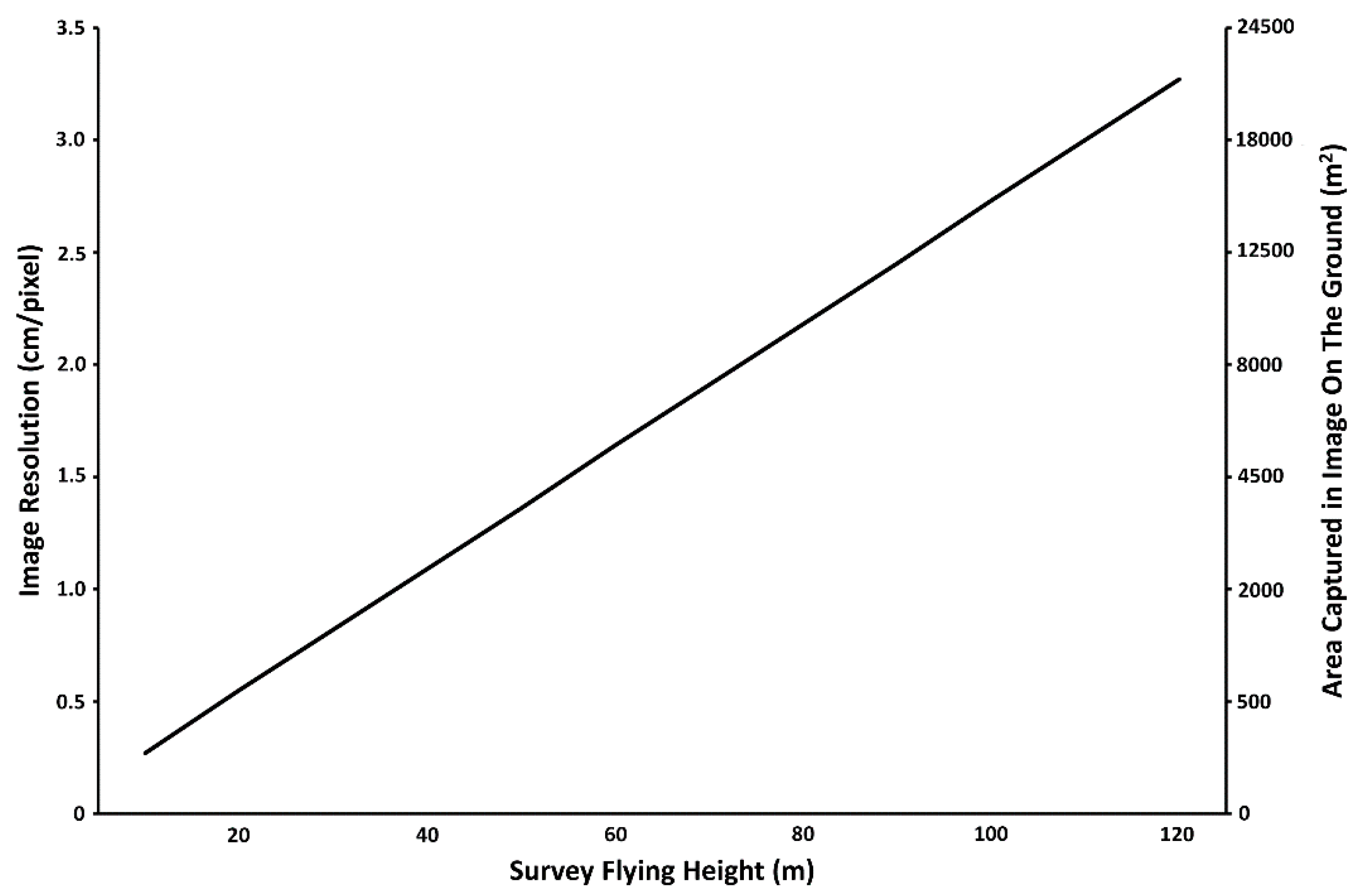
1.5. Considerations for Drone Operations in Marine Environments
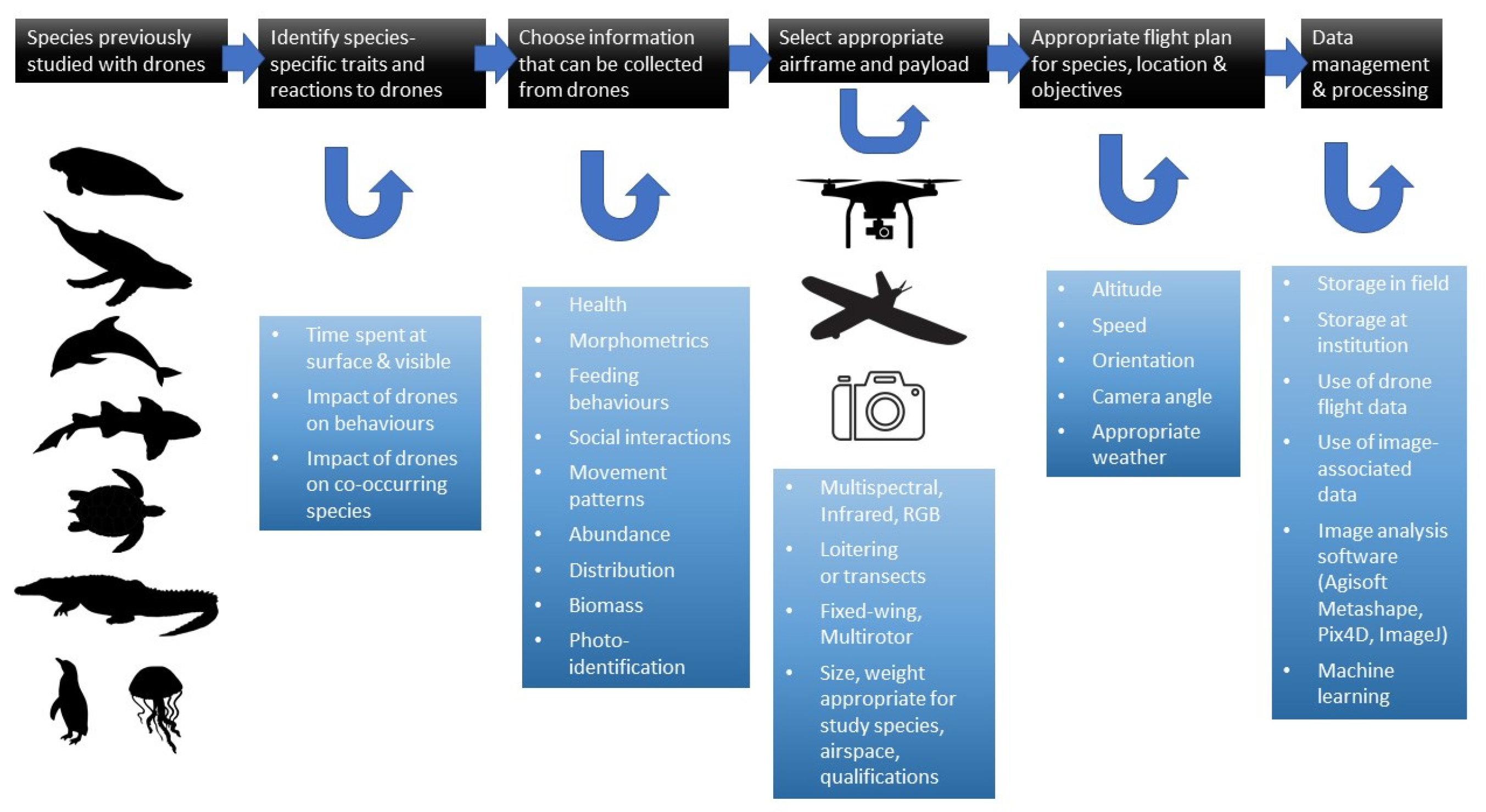
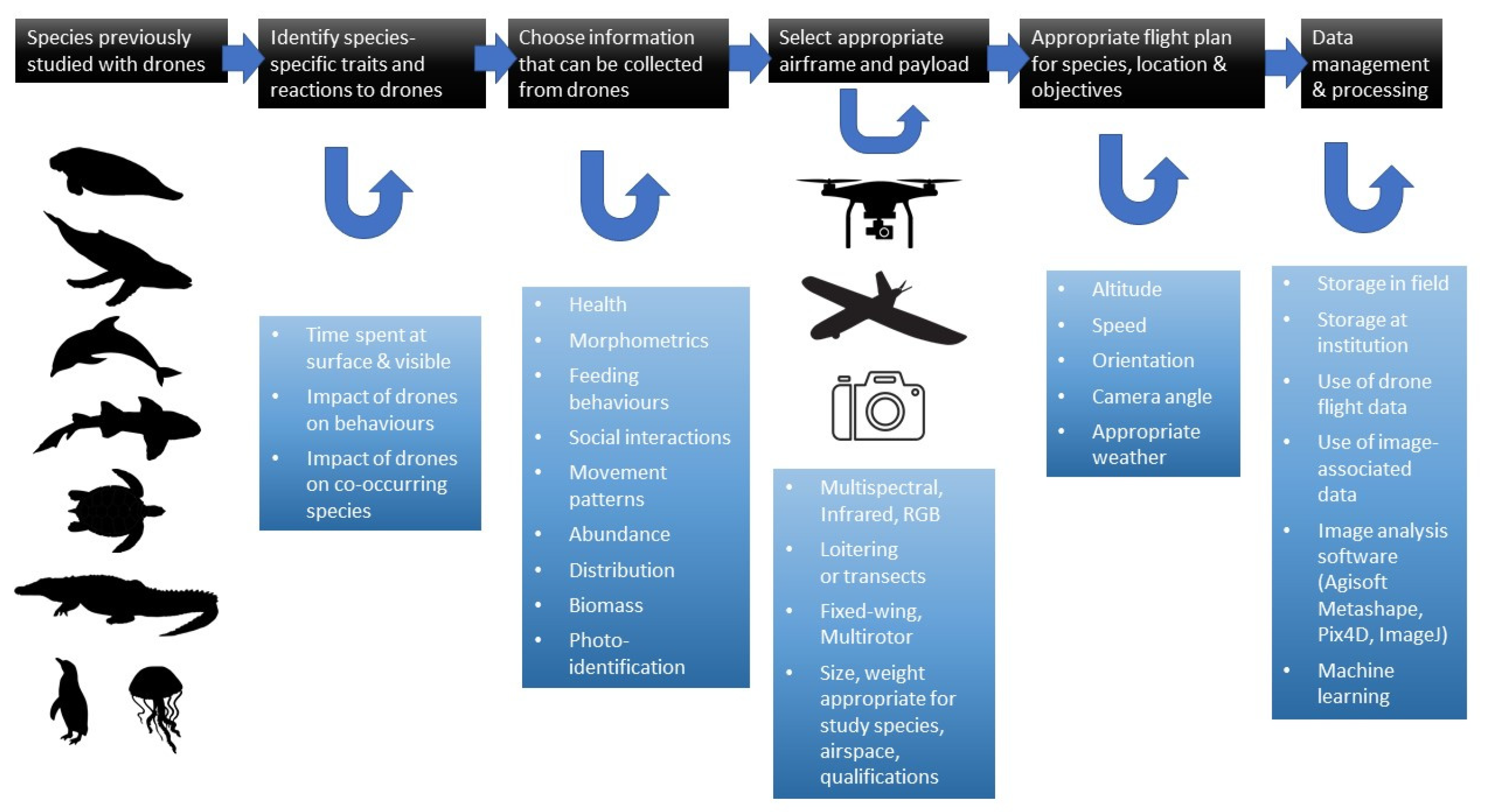
2. Protocols for the Use of Drones with Specific Taxa
2.1. Protocols for the Study of Jellyfish
2.1.1. Unique Aspects of Jellyfish Protocols
2.1.2. Drone Selection for Jellyfish Research
2.1.3. Unmanned Aerial Vehicle (UAV) Flight Plan
2.1.4. Field Validation
2.1.5. Image Processing
2.1.6. Example Protocol for Studying Jellyfish with Drones
2.2. Protocols for the Use of Drones for Shark Research
2.2.1. Shark Traits
2.2.2. Impact of Drones on Sharks
2.2.3. Drone Selection for Shark Research
2.2.4. Example Protocol for Studying Sharks with Drones
2.3. Protocols for the Study of Marine Reptiles
2.3.1. Traits of Marine Reptiles
2.3.2. Drone Selection for Marine Reptiles
2.3.3. Methodological Difficulties
2.3.4. Example Protocol for Studying Marine Reptiles with Drones
2.4. Protocols for the Study of Marine Birds
2.4.1. Traits of Marine Birds
2.4.2. Impacts of Drones on Marine Birds
2.4.3. Selection of Drone for Marine Bird Research
2.4.4. Potential Hazards and Methodological Difficulties
2.4.5. Example Protocols for Studying Marine Birds with Drones
2.5. Protocols for the Study of Pinnipeds
2.5.1. Pinniped Traits
2.5.2. Impact of Drones on Pinnipeds
2.5.3. Drone Selection for Pinniped Research
2.5.4. Example Protocol for Studying Pinnipeds with Drones
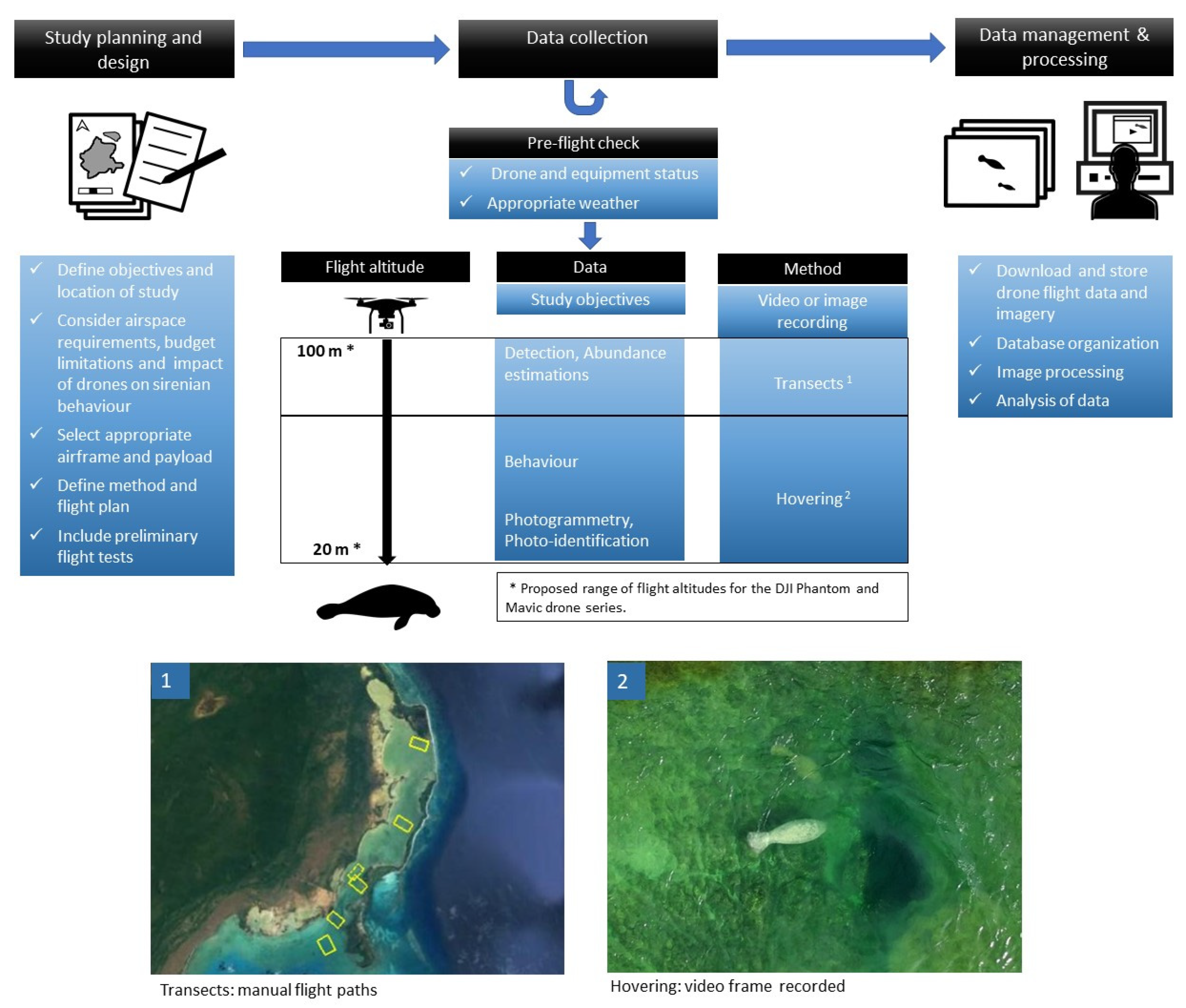
2.6. Protocols for the Study of Sirenians
2.6.1. Traits of Sirenians
2.6.2. Impact of Drones on Sirenians
2.6.3. Drone Selection for Sirenian Research
2.6.4. Detection and Density Estimations
2.6.5. Photoidentification and Photogrammetry
2.6.6. Behavioral Studies
2.6.7. Capturing and Health Monitoring
2.6.8. Methodological Difficulties
2.7. Protocols for the Study of Odontocetes
2.7.1. Traits of Dolphins
2.7.2. Drone Selection for Dolphin Research
2.7.3. Behavioural Impacts of Drones on Dolphins
2.8. Protocols for the Study of Baleen Whales
2.8.1. Traits of Baleen Whales
2.8.2. Reducing Disturbance
2.8.3. Drone Selection for Baleen Whales
2.8.4. Individual Sampling
2.8.5. Flight Planning, Control and Other Operational Considerations
2.8.6. Example Protocol for Studying Whales with Drones
3. Synopsis
3.1. Impacts of Drones on Marine Animals
3.2. Use of Drones to Study Fishes
3.3. Lack of Use of Vertical Take Off and Landing (VTOL) Aircraft for Marine Research
3.4. Legislation
3.5. Safety and Personal Protective Equipment for Use of Drones in Marine Research
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Ivošević, B.; Han, Y.-G.; Cho, Y.; Kwon, O. The use of conservation drones in ecology and wildlife research. J. Ecol. Environ. 2015, 38, 113–118. [Google Scholar] [CrossRef] [Green Version]
- Schofield, G.; Esteban, N.; Katselidis, K.A.; Hays, G.C. Drones for research on sea turtles and other marine vertebrates–A review. Biol. Conserv. 2019, 238, 108214. [Google Scholar] [CrossRef]
- Mulero-Pázmány, M.; Jenni-Eiermann, S.; Strebel, N.; Sattler, T.; Negro, J.J.; Tablado, Z. Unmanned aircraft systems as a new source of disturbance for wildlife: A systematic review. PLoS ONE 2017, 12, e0178448. [Google Scholar] [CrossRef] [Green Version]
- Acevedo-Whitehouse, K.; Rocha-Gosselin, A.; Gendron, D. A novel non-invasive tool for disease surveillance of free-ranging whales and its relevance to conservation programs. Anim. Conserv. 2010, 13, 217–225. [Google Scholar] [CrossRef]
- Merheb, A.-R.; Noura, H.; Bateman, F. Emergency control of AR drone quadrotor UAV suffering a total loss of one rotor. IEEE/ASME Trans. Mechatron. 2017, 22, 961–971. [Google Scholar] [CrossRef]
- Harvey, M.C.; Rowland, J.V.; Luketina, K.M. Drone with thermal infrared camera provides high resolution georeferenced imagery of the Waikite geothermal area, New Zealand. J. Volcanol. Geotherm. Res. 2016, 325, 61–69. [Google Scholar] [CrossRef]
- Burke, C.; Rashman, M.; Wich, S.; Symons, A.; Theron, C.; Longmore, S. Optimizing observing strategies for monitoring animals using drone-mounted thermal infrared cameras. Int. J. Remote Sens. 2019, 40, 439–467. [Google Scholar] [CrossRef] [Green Version]
- Wosnick, N.; Navas, C.A.; Niella, Y.V.; Monteiro-Filho, E.L.A.; Freire, C.A.; Hammerschlag, N. Thermal imaging reveals changes in body surface temperatures of blacktip sharks (Carcharhinus limbatus) during air exposure. Physiol. Biochem. Zool. 2018, 91, 1005–1012. [Google Scholar] [CrossRef] [Green Version]
- Dawson, S.M.; Bowman, M.H.; Leunissen, E.; Sirguey, P. Inexpensive aerial photogrammetry for studies of whales and large marine animals. Front. Mar. Sci. 2017, 4, 366. [Google Scholar] [CrossRef] [Green Version]
- Manfreda, S.; Dal Sasso, S.F.; Pizarro, A.; Tauro, F. New Insights Offered by UAS for River Monitoring. In Applications of Small Unmanned Aircraft Systems: Best Practices and Case Studies; CRC Press-Taylor Francis Group: Boca Raton, FL, USA, 2019; pp. 211–234. [Google Scholar]
- Casella, E.; Collin, A.; Harris, D.; Ferse, S.; Bejarano, S.; Parravicini, V.; Hench, J.L.; Rovere, A. Mapping coral reefs using consumer-grade drones and structure from motion photogrammetry techniques. Coral Reefs 2017, 36, 269–275. [Google Scholar] [CrossRef]
- Geraeds, M.; Van Emmerik, T.; De Vries, R.; Bin Ab Razak, M.S. Riverine plastic litter monitoring using unmanned aerial vehicles (UAVs). Remote Sens. 2019, 11, 2045. [Google Scholar] [CrossRef] [Green Version]
- Pirotta, V.; Smith, A.; Ostrowski, M.; Russell, D.; Jonsen, I.D.; Grech, A.; Harcourt, R. An economical custom-built drone for assessing whale health. Front. Mar. Sci. 2017, 4, 425. [Google Scholar] [CrossRef]
- Hensel, E.; Wenclawski, S.; Layman, C.A. Using a small, consumer-grade drone to identify and count marine megafauna in shallow habitats. Lat. Am. J. Aquat. Res. 2018, 46, 1025–1033. [Google Scholar] [CrossRef]
- Hodgson, A.; Kelly, N.; Peel, D. Unmanned aerial vehicles (UAVs) for surveying marine fauna: A dugong case study. PLoS ONE 2013, 8, e79556. [Google Scholar] [CrossRef] [Green Version]
- Schofield, G.; Katselidis, K.A.; Lilley, M.K.; Reina, R.D.; Hays, G.C. Detecting elusive aspects of wildlife ecology using drones: New insights on the mating dynamics and operational sex ratios of sea turtles. Funct. Ecol. 2017, 31, 2310–2319. [Google Scholar] [CrossRef]
- Bevan, E.; Wibbels, T.; Navarro, E.; Rosas, M.; Najera, B.M.Z.; Sarti, L.; Illescas, F.; Montano, J.; Peña, L.J.; Burchfield, P. Using unmanned aerial vehicle (UAV) technology for locating, identifying, and monitoring courtship and mating behaviour in the green turtle (Chelonia mydas). Herpetol. Rev. 2016, 47, 27–32. [Google Scholar]
- Joyce, K.E.; Duce, S.; Leahy, S.M.; Leon, J.X.; Maier, S.W. Principles and practice of acquiring drone–based image data in marine environments. Mar. Freshwat. Res. 2019, 70, 952–963. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Colefax, A.P.; Tagliafico, A.; Bishop, M.J.; Giles, A.; Butcher, P.A. Assessing variation in assemblages of large marine fauna off ocean beaches using drones. Mar. Freshw. Res. 2019, 71, 68–77. [Google Scholar] [CrossRef] [Green Version]
- Bevan, E.; Wibbels, T.; Najera, B.M.; Martinez, M.A.; Martinez, L.A.; Martinez, F.I.; Cuevas, J.M.; Anderson, T.; Bonka, A.; Hernandez, M.H.; et al. Unmanned aerial vehicles (UAVs) for monitoring sea turtles in near-shore waters. Mar. Turtle Newsl. 2015, 145, 19–22. [Google Scholar]
- Brooke, S.; Graham, D.; Jacobs, T.; Littnan, C.; Manuel, M.; O’Conner, R. Testing marine conservation applications of unmanned aerial systems (UAS) in a remote marine protected area. J. Unmanned Veh. Syst. 2015, 3, 237–251. [Google Scholar] [CrossRef]
- Pitt, K.A.; Budarf, A.C.; Browne, J.G.; Condon, R.H. Bloom and bust: Why do blooms of jellyfish collapse? In Jellyfish Blooms; Springer: Dordrecht, The Netherlands, 2014; pp. 79–103. [Google Scholar]
- Fuentes, V.L.; Purcell, J.E.; Condon, R.H.; Lombard, F.; Lucas, C.H. Jellyfish blooms: Advances and challenges. Mar. Ecol. Prog. Ser. 2018, 591, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Doyle, T.K.; Houghton, J.D. A paradigm shift in the trophic importance of jellyfish? Trends Ecol. Evol. 2018, 33, 874–884. [Google Scholar] [CrossRef] [Green Version]
- Raoult, V.; Gaston, T.F. Rapid biomass and size-frequency estimates of edible jellyfish populations using drones. Fish. Res. 2018, 207, 160–164. [Google Scholar] [CrossRef]
- Schaub, J.; Hunt, B.P.V.; Pakhomov, E.A.; Holmes, K.; Lu, Y.; Quayle, L. Using unmanned aerial vehicles (UAVs) to measure jellyfish aggregations. Mar. Ecol. Prog. Ser. 2018, 591, 29–36. [Google Scholar] [CrossRef]
- Vautherin, J.; Rutishauser, S.; Schneider-Zapp, K.; Choi, H.F.; Chovancova, V.; Glass, A.; Strecha, C. Photogrammetric accuracy and modeling of rolling shutter cameras. In Proceedings of the ISPRS Annals of the Photogrammetry, Remote Sensing and Spatial Information Sciences, Prague, Czech Republic, 12–19 July 2016; Volume 3. [Google Scholar]
- Pitt, K.A.; Kingsford, M.J. Temporal variation in the virgin biomass of the edible jellyfish, Catostylus mosaicus (Scyphozoa, Rhizostomeae). Fish. Res. 2003, 63, 303–313. [Google Scholar] [CrossRef]
- Kiszka, J.J.; Mourier, J.; Gastrich, K.; Heithaus, M.R. Using unmanned aerial vehicles (UAVs) to investigate shark and ray densities in a shallow coral lagoon. Mar. Ecol. Prog. Ser. 2016, 560, 237–242. [Google Scholar] [CrossRef]
- Colefax, A.P.; Kelaher, B.P.; Pagendam, D.E.; Butcher, P.A. Assessing White Shark (Carcharodon carcharias) Behavior Along Coastal Beaches for Conservation-Focused Shark Mitigation. Front. Mar. Sci. 2020, 7, 268. [Google Scholar] [CrossRef]
- Rieucau, G.; Kiszka, J.J.; Castillo, J.C.; Mourier, J.; Boswell, K.M.; Heithaus, M.R. Using unmanned aerial vehicle (UAV) surveys and image analysis in the study of large surface-associated marine species: A case study on reef sharks Carcharhinus melanopterus shoaling behaviour. J. Fish Biol. 2018, 93, 119–127. [Google Scholar] [CrossRef]
- Doan, M.D.; Kajiura, S.M. Adult blacktip sharks (Carcharhinus limbatus) use shallow water as a refuge from great hammerheads (Sphyrna mokarran). J. Fish. Biol. 2020, 96. [Google Scholar] [CrossRef]
- Gore, M.; Abels, L.; Wasik, S.; Saddler, L.; Ormond, R. Are close-following and breaching behaviours by basking sharks at aggregation sites related to courtship? J. Mar. Biol. Assoc. UK 2018, 99, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lea, J.S.E.; Daly, R.; Leon, C.; Daly, C.A.K.; Clarke, C.R. Life after death: Behaviour of multiple shark species scavenging a whale carcass. Mar. Freshw. Res. 2019, 70, 302–306. [Google Scholar] [CrossRef]
- Ho, C.; Joly, K.; Nosal, A.P.; Lowe, C.G.; Clark, C.M. Predicting coordinated group movements of sharks with limited observations using AUVs. In Proceedings of the Symposium on Applied Computing, Marrakech, Morocco, 3 April 2017; pp. 289–296. [Google Scholar]
- Colefax, A.P.; Butcher, P.A.; Kelaher, B.P. The potential for unmanned aerial vehicles (UAVs) to conduct marine fauna surveys in place of manned aircraft. ICES J. Mar. Sci. 2018, 75, 1–8. [Google Scholar] [CrossRef]
- Christie, K.S.; Gilbert, S.L.; Brown, C.L.; Hatfield, M.; Hanson, L. Unmanned aircraft systems in wildlife research: Current and future applications of a transformative technology. Front. Ecol. Environ. 2016, 14, 241–251. [Google Scholar] [CrossRef]
- Kelaher, B.P.; Peddemors, V.M.; Hoade, B.; Colefax, A.P.; Butcher, P.A. Comparison of sampling precision for nearshore marine wildlife using unmanned and manned aerial surveys. J. Unmanned Veh. Syst. 2019, 8, 30–43. [Google Scholar] [CrossRef]
- Butcher, P.A.; Piddocke, T.P.; Colefax, A.P.; Hoade, B.; Peddemors, V.M.; Borg, L.; Cullis, B.R. Beach safety: Can drones provide a platform for sighting sharks? Wildl. Res. 2020, 46, 701–712. [Google Scholar] [CrossRef] [Green Version]
- Colefax, A.P.; Butcher, P.A.; Pagendam, D.E.; Kelaher, B.P. Reliability of marine faunal detections in drone-based monitoring. Ocean Coast. Manag. 2019, 174, 108–115. [Google Scholar] [CrossRef]
- Rowat, D.; Gore, M.; Meekan, M.G.; Lawler, I.R.; Bradshaw, C.J.A. Aerial survey as a tool to estimate whale shark abundance trends. J. Exp. Mar. Biol. Ecol. 2009, 368, 1–8. [Google Scholar] [CrossRef]
- Benavides, M.T.; Fodrie, F.J.; Johnston, D.W. Shark detection probability from aerial drone surveys within a temperate estuary. J. Unmanned Veh. Syst. 2019, 8, 44–56. [Google Scholar] [CrossRef]
- Spaet, J.L.Y.; Patterson, T.A.; Bradford, R.W.; Butcher, P.A. Spatiotemporal distribution patterns of immature Australasian white sharks (Carcharodon carcharias). Sci. Rep. 2020, 10, 1–13. [Google Scholar]
- Westgate, A.J.; Koopman, H.N.; Siders, Z.A.; Wong, S.N.; Ronconi, R.A. Population density and abundance of basking sharks Cetorhinus maximus in the lower Bay of Fundy, Canada. Endanger. Spec. Res. 2014, 23, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Saqib, M.; Khan, S.D.; Sharma, N.; Scully-Power, P.; Butcher, P.; Colefax, A.; Blumenstein, M. Real-time drone surveillance and population estimation of marine animals from aerial imagery. In Proceedings of the 2018 International Conference on Image and Vision Computing New Zealand (IVCNZ), Auckland, New Zealand, 19–21 November 2018; pp. 1–6. [Google Scholar]
- Christiansen, F.; Rojano-Doñate, L.; Madsen, P.T.; Bejder, L. Noise levels of multi-rotor unmanned aerial vehicles with implications for potential underwater impacts on marine mammals. Front. Mar. Sci. 2016, 3, 277. [Google Scholar] [CrossRef] [Green Version]
- Raoult, V.; Tosetto, L.; Williamson, J.E. Drone-Based High-Resolution Tracking of Aquatic Vertebrates. Drones 2018, 2, 37. [Google Scholar] [CrossRef] [Green Version]
- Tucker, J.P.; Vercoe, B.; Santos, I.R.; Dujmovic, M.; Butcher, P.A. Whale carcass scavenging by sharks. Glob. Ecol. Conserv. 2019, 19, e00655. [Google Scholar] [CrossRef]
- Martin, R.A.; Rossmo, D.K.; Hammerschlag, N. Hunting patterns and geographic profiling of white shark predation. J. Zool. 2009, 279, 111–118. [Google Scholar] [CrossRef]
- Harasti, D.; Lee, K.; Laird, R.; Bradford, R.; Bruce, B. Use of stereo baited remote underwater video systems to estimate the presence and size of white sharks (Carcharodon carcharias). Mar. Freshw. Res. 2017, 68, 1391–1396. [Google Scholar] [CrossRef]
- Queiroz, N.; Humphries, N.E.; Couto, A.; Vedor, M.; da Costa, I.; Sequeira, A.M.; Mucientes, G.; Santos, A.M.; Abascal, F.J.; Abercrombie, D.L.; et al. Global spatial risk assessment of sharks under the footprint of fisheries. Nature 2019, 572, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, N.; Humphries, N.E.; Mucientes, G.; Hammerschlag, N.; Lima, F.P.; Scales, K.L.; Miller, P.I.; Sousa, L.L.; Seabra, R.; Sims, D.W. Ocean-wide tracking of pelagic sharks reveals extent of overlap with longline fishing hotspots. Proc. Natl. Acad. Sci. USA 2016, 113, 1582–1587. [Google Scholar] [CrossRef] [Green Version]
- Chirayath, V.; Earle, S.A. Drones that see through waves–preliminary results from airborne fluid lensing for centimetre-scale aquatic conservation. Aquat. Conserv. Mar. Freshwat. Ecosyst. 2016, 26, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Rees, A.; Alfaro-Shigueto, J.; Barata, P.; Bjorndal, K.A.; Bolten, A.B.; Bourjea, J.; Broderick, A.; Campbell, L.; Cardona, L.; Carreras, C.; et al. Are we working towards global research priorities for management and conservation of sea turtles? Endanger. Spec. Res. 2016, 31, 337–382. [Google Scholar] [CrossRef] [Green Version]
- Hays, G.C.; Bailey, H.; Bograd, S.J.; Bowen, W.D.; Campagna, C.; Carmichael, R.H.; Casale, P.; Chiaradia, A.; Costa, D.P.; Cuevas, E.; et al. Translating marine animal tracking data into conservation policy and management. Trends Ecol. Evol. 2019, 34, 459–473. [Google Scholar] [CrossRef] [Green Version]
- Pfaller, J.B.; Bjorndal, K.A.; Chaloupka, M.; Williams, K.L.; Frick, M.G.; Bolten, A.B. Accounting for imperfect detection is critical for inferring marine turtle nesting population trends. PLoS ONE 2013, 8, e62326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunstan, A.; Robertson, K.; Fitzpatrick, R.; Pickford, J.; Meager, J. Use of unmanned aerial vehicles (UAVs) for mark-resight nesting population estimation of adult female green sea turtles at Raine Island. PLoS ONE 2020, 15, e0228524. [Google Scholar] [CrossRef] [PubMed]
- Schofield, G.; Papafitsoros, K.; Haughey, R.; Katselidis, K. Aerial and underwater surveys reveal temporal variation in cleaning-station use by sea turtles at a temperate breeding area. Mar. Ecol. Prog. Ser. 2017, 575, 153–164. [Google Scholar] [CrossRef]
- Fuentes, M.M.; Pike, D.A.; Dimatteo, A.; Wallace, B.P. Resilience of marine turtle regional management units to climate change. Glob. Change Biol. 2013, 19, 1399–1406. [Google Scholar] [CrossRef]
- Campbell, H.A.; Watts, M.E.; Sullivan, S.; Read, M.A.; Choukroun, S.; Irwin, S.R.; Franklin, C.E. Estuarine crocodiles ride surface currents to facilitate long-distance travel. J. Anim. Ecol. 2010, 79, 955–964. [Google Scholar] [CrossRef]
- Bevan, E.; Whiting, S.; Tucker, T.; Guinea, M.; Raith, A.; Douglas, R. Measuring behavioral responses of sea turtles, saltwater crocodiles, and crested terns to drone disturbance to define ethical operating thresholds. PLoS ONE 2018, 13, e0194460. [Google Scholar] [CrossRef]
- Gallagher, A.J.; Papastamatiou, Y.P.; Barnett, A. Apex predatory sharks and crocodiles simultaneously scavenge a whale carcass. J. Ethol. 2018, 36, 205–209. [Google Scholar] [CrossRef]
- Sykora-Bodie, S.T.; Bezy, V.; Johnston, D.W.; Newton, E.; Lohmann, K.J. Quantifying nearshore sea turtle densities: Applications of unmanned aerial systems for population assessments. Sci. Rep. 2017, 7, 1–7. [Google Scholar] [CrossRef]
- Tapilatu, R.F.; Bonka, A.; Iwanggin, W.G.; Wona, H.; Ampnir, T.; Rumbiak, R.; Bawole, R.; Wibbels, T. Unmanned Aerial Vehicle (UAV) Use as a Tool to Assess Crawling and Swimming Speeds in Hatchling Sea Turtles. Soc. Stud. Amphib. Reptiles 2019, 50, 722–726. [Google Scholar]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Raja Segaran, R.; Reid, I.; Terauds, A.; Koh, L.P. Drones count wildlife more accurately and precisely than humans. Methods Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef] [Green Version]
- Varela, M.R.; Patrício, A.R.; Anderson, K.; Broderick, A.C.; DeBell, L.; Hawkes, L.A.; Tilley, D.; Snape, R.T.; Westoby, M.J.; Godley, B.J. Assessing climate change associated sea-level rise impacts on sea turtle nesting beaches using drones, photogrammetry and a novel GPS system. Glob. Change Biol. 2019, 25, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Watts, A.C.; Perry, J.H.; Smith, S.E.; Burgess, M.A.; Wilkinson, B.E.; Szantoi, Z.; Ifju, P.G.; Percival, H.F. Small unmanned aircraft systems for low-altitude aerial surveys. J. Wildl. Manag. 2010, 74, 1614–1619. [Google Scholar] [CrossRef]
- Elsey, R.M.; Trosclair, P.L. The use of an unmanned aerial vehicle to locate alligator nests. Southeast. Nat. 2016, 15, 76–82. [Google Scholar] [CrossRef]
- Thapa, G.J.; Thapa, K.; Thapa, R.; Jnawali, S.R.; Wich, S.A.; Poudyal, L.P.; Karki, S. Counting crocodiles from the sky: Monitoring the critically endangered gharial (Gavialis gangeticus) population with an unmanned aerial vehicle (UAV). J. Unmanned Veh. Syst. 2018, 6, 71–82. [Google Scholar] [CrossRef] [Green Version]
- Buckland, S.T.; Anderson, D.R.; Burnham, K.P.; Laake, J.L.; Borchers, D.L.; Thomas, L. Introduction to Distance Sampling: Estimating Abundance of Biological Populations; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Gray, P.C.; Fleishman, A.B.; Klein, D.J.; McKown, M.W.; Bézy, V.S.; Lohmann, K.J.; Johnston, D.W. A convolutional neural network for detecting sea turtles in drone imagery. Methods Ecol. Evol. 2019, 10, 345–355. [Google Scholar] [CrossRef]
- Dujon, A.M.; Schofield, G. Importance of machine learning for enhancing ecological studies using information-rich imagery. Endanger. Spec. Res. 2019, 39, 91–104. [Google Scholar] [CrossRef] [Green Version]
- Blackmer, A.L.; Ackerman, J.T.; Nevitt, G.A. Effects of investigator disturbance on hatching success and nest-site fidelity in a long-lived seabird, Leach’s storm-petrel. Biol. Conserv. 2004, 116, 141–148. [Google Scholar] [CrossRef]
- Fay, R.; Barbraud, C.; Delord, K.; Weimerskirch, H. From early life to senescence: Individual heterogeneity in a long-lived seabird. Ecol. Monogr. 2018, 88, 60–73. [Google Scholar] [CrossRef] [Green Version]
- Brisson-Curadeau, É.; Bird, D.; Burke, C.; Fifield, D.A.; Pace, P.; Sherley, R.B.; Elliott, K.H. Seabird species vary in behavioural response to drone census. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Weimerskirch, H.; Prudor, A.; Schull, Q. Flights of drones over sub-Antarctic seabirds show species-and status-specific behavioural and physiological responses. Polar Biol. 2018, 41, 259–266. [Google Scholar] [CrossRef]
- Rush, G.P.; Clarke, L.E.; Stone, M.; Wood, M.J. Can drones count gulls? Minimal disturbance and semiautomated image processing with an unmanned aerial vehicle for colony-nesting seabirds. Ecol. Evol. 2018, 8, 12322–12334. [Google Scholar] [CrossRef] [PubMed]
- Sardà-Palomera, F.; Bota, G.; Padilla, N.; Brotons, L.; Sardà, F. Unmanned aircraft systems to unravel spatial and temporal factors affecting dynamics of colony formation and nesting success in birds. J. Avian Biol. 2017, 48, 1273–1280. [Google Scholar] [CrossRef]
- Ratcliffe, N.; Guihen, D.; Robst, J.; Crofts, S.; Stanworth, A.; Enderlein, P. A protocol for the aerial survey of penguin colonies using UAVs. J. Unmanned Veh. Syst. 2015, 3, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Rümmler, M.-C.; Mustafa, O.; Maercker, J.; Peter, H.-U.; Esefeld, J. Sensitivity of Adélie and Gentoo penguins to various flight activities of a micro UAV. Polar Biol. 2018, 41, 2481–2493. [Google Scholar] [CrossRef]
- Lyons, M.B.; Brandis, K.J.; Murray, N.J.; Wilshire, J.H.; McCann, J.A.; Kingsford, R.T.; Callaghan, C.T. Monitoring large and complex wildlife aggregations with drones. Methods Ecol. Evol. 2019, 10, 1024–1035. [Google Scholar] [CrossRef] [Green Version]
- Valle, R.G.; Scarton, F. Drones improve effectiveness and reduce disturbance of censusing Common Redshanks Tringa totanus breeding on salt marshes. Ardea 2020, 107, 275–282. [Google Scholar] [CrossRef]
- Albores-Barajas, Y.V.; Soldatini, C.; Ramos-Rodríguez, A.; Alcala-Santoyo, J.E.; Carmona, R.; Dell’Omo, G. A new use of technology to solve an old problem: Estimating the population size of a burrow nesting seabird. PLoS ONE 2018, 13, e0202094. [Google Scholar] [CrossRef] [Green Version]
- Chabot, D.; Craik, S.R.; Bird, D.M. Population census of a large common tern colony with a small unmanned aircraft. PLoS ONE 2015, 10, e0122588. [Google Scholar] [CrossRef] [Green Version]
- Groom, G.; Stjernholm, M.; Nielsen, R.D.; Fleetwood, A.; Petersen, I.K. Remote sensing image data and automated analysis to describe marine bird distributions and abundances. Ecol. Inform. 2013, 14, 2–8. [Google Scholar] [CrossRef]
- Lyons, M.; Brandis, K.; Callaghan, C.; McCann, J.; Mills, C.; Ryall, S.; Kingsford, R. Bird interactions with drones, from individuals to large colonies. bioRxiv 2017. bioRxiv:109926. [Google Scholar] [CrossRef] [Green Version]
- Kirkwood, R.; Gales, R.; Terauds, A.; Arnould, J.P.; Pemberton, D.; Shaughnessy, P.D.; Mitchell, A.T.; Gibbens, J. Pup production and population trends of the Australian fur seal (Arctocephalus pusillus doriferus). Mar. Mamm. Sci. 2005, 21, 260–282. [Google Scholar] [CrossRef]
- DeMaster, D.P. Pinniped Monitoring Handbook: Channel Islands National Park, California; Channel Islands National Park, National Park Service: Ventura, CA, USA, 1988.
- McIntosh, R.R.; Kirkwood, R.; Sutherland, D.R.; Dann, P. Drivers and annual estimates of marine wildlife entanglement rates: A long-term case study with Australian fur seals. Mar. Pollut. Bull. 2015, 101, 716–725. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, K.M.; Aguilar, A.; Aurioles, D.; Burkanov, V.; Campagna, C.; Gales, N.; Gelatt, T.; Goldsworthy, S.D.; Goodman, S.J.; Hofmeyr, G.J.; et al. Global threats to pinnipeds. Mar. Mamm. Sci. 2012, 28, 414–436. [Google Scholar] [CrossRef]
- Ross, P.S. Marine Mammals as Sentinels in Ecological Risk Assessment. Hum. Ecol. Risk Assess. Int. J. 2000, 6, 29–46. [Google Scholar] [CrossRef]
- Udevitz, M.S.; Jay, C.; Cody, M. Observer variability in pinniped counts: Ground-based enumeration of walruses at haul-out sites. Mar. Mamm. Sci. 2005, 21, 108–120. [Google Scholar] [CrossRef]
- Bradshaw, C.J.; Barker, R.J.; Harcourt, R.G.; Davis, L.S. Estimating survival and capture probability of fur seal pups using multistate mark-recapture models. J. Mammal. 2003, 84, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, K.L.; Shertzer, K.W.; Fritz, L.W.; Read, A.J. A novel approach to compare pinniped populations across a broad geographic range. Can. J. Aquat. Sci. 2015, 72, 175–185. [Google Scholar] [CrossRef]
- Wade, P.R. Calculating limits to the allowable human-caused mortality of cetaceans and pinnipeds. Mar. Mamm. Sci. 1998, 14, 1–37. [Google Scholar] [CrossRef]
- Van der Hoop, J.; Barco, S.G.; Costidis, A.M.; Gulland, F.M.; Jepson, P.D.; Moore, K.T.; Raverty, S.; McLellan, W.A. Criteria and case definitions for serious injury and death of pinnipeds and cetaceans caused by anthropogenic trauma. Dis. Aquat. Org. 2013, 103, 229–264. [Google Scholar]
- Sorrell, K.J.; Clarke, R.H.; Holmberg, R.; McIntosh, R.R. Remotely piloted aircraft improve precision of capture-mark-resight population estimates of Australian fur seals. Ecosphere 2019, 10, 14. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, J.C.; Holman, D.; Terauds, A.; Koh, L.P.; Goldsworthy, S.D. Rapid condition monitoring of an endangered marine vertebrate using precise, non-invasive morphometrics. Biol. Conserv. 2020, 242, 11. [Google Scholar] [CrossRef]
- Allan, B.M.; Ierodiaconou, D.; Hoskins, A.J.; Arnould, J.P. A rapid UAV method for assessing body condition in fur seals. Drones 2019, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, R.R.; Holmberg, R.; Dann, P. Looking Without Landing-Using Remote Piloted Aircraft to Monitor Fur Seal Populations Without Disturbance. Front. Mar. Sci. 2018, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Wieting, D. Environmental Assessment for Issuance of Permits to take Steller Sea Lions by Harassment During Surveys Using Unmanned Aerial Systems; NOAA: Silver Spring, MD, USA, 2014.
- Bejder, M.; Johnston, D.W.; Smith, J.; Friedlaender, A.; Bejder, L. Embracing conservation success of recovering humpback whale populations: Evaluating the case for downlisting their conservation status in Australia. Mar. Policy 2016, 66, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Goebel, M.E.; Perryman, W.L.; Hinke, J.T.; Krause, D.J.; Hann, N.A.; Gardner, S.; LeRoi, D.J. A small unmanned aerial system for estimating abundance and size of Antarctic predators. Polar Biol. 2015, 38, 619–630. [Google Scholar] [CrossRef] [Green Version]
- Pomeroy, P.; O’Connor, L.; Davies, P. Assessing use of and reaction to unmanned aerial systems in gray and harbor seals during breeding and molt in the UK. J. Unmanned Veh. Syst. 2015, 3, 102–113. [Google Scholar] [CrossRef] [Green Version]
- Moreland, E.E.; Cameron, M.F.; Angliss, R.P.; Boveng, P.L. Evaluation of a ship-based unoccupied aircraft system (UAS) for surveys of spotted and ribbon seals in the Bering Sea pack ice. J. Unmanned Veh. Syst. 2015, 3, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Arona, L.; Dale, J.; Heaslip, S.G.; Hammill, M.O.; Johnston, D.W. Assessing the disturbance potential of small unoccupied aircraft systems (UAS) on gray seals (Halichoerus grypus) at breeding colonies in Nova Scotia, Canada. PeerJ 2018, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Mustafa, O.; Barbosa, A.; Krause, D.J.; Peter, H.-U.; Vieira, G.; Rümmler, M.-C. State of knowledge: Antarctic wildlife response to unmanned aerial systems. Polar Biol. 2018, 41, 2387–2398. [Google Scholar] [CrossRef]
- Mamaev, E.G. A new method of counting phoca vitulina ssp. stejnegeri (phocidae, carnivora) on the commander islands (Russia). Nat. Conserv. Res. 2018, 3, 44–58. [Google Scholar] [CrossRef]
- Hartman, D.S. Ecology and Behavior of the Manatee (Trichechus Manatus) in Florida; Special Publication No 5; The American Society of Mammologists: Lawrence, KS, USA, 1979; p. 153. [Google Scholar]
- Anderson, P.K. Habitat, niche, and evolution of Sirenian mating systems. J. Mamm. Evol. 2002, 9, 55–98. [Google Scholar] [CrossRef]
- Lanyon, J.M. Distribution and abundance of dugongs in Moreton Bay, Queensland, Australia. Wildl. Res. 2003, 30, 397–409. [Google Scholar] [CrossRef]
- Ackerman, B.B. Aerial Surveys of Manatees: A Summary and Progress Report. In Population Biology of the Florida Manatee; O’Shea, T.J., Ackerman, B.B., Percival, H.F., Eds.; US Department of the Interior, National Biological Service: Washington, DC, USA, 1995; Volume 1, pp. 13–33. [Google Scholar]
- Alves, M.D.; Kinas, P.G.; Marmontel, M.; Borges, J.C.G.; Costa, A.F.; Schiel, N.; Araújo, M.E. First abundance estimate of the Antillean manatee (Trichechus manatus manatus) in Brazil by aerial survey. J. Mar. Biol. Assoc. UK 2016, 96, 955–966. [Google Scholar] [CrossRef]
- Gonzalez-Socoloske, D.; Olivera-Gomez, L.D.; Ford, R.E. Detection of free-ranging West Indian manatees Trichechus manatus using side-scan sonar. Endanger. Spec. Res. 2009, 8, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Brice, C.E. The detection of Amazonian Manatees (Trichechus inunguis) using side-scan sonar and the effect of oil activities on their habitats in Eastern Ecuador. Master’s Thesis, Nova Southeastern University Oceanographic Center, Davie, FL, USA, 2014. [Google Scholar]
- Ramos, E.A.; Maloney, B.M.; Magnasco, M.O.; Reiss, D. Bottlenose dolphins and Antillean manatees respond to small multi-rotor unmanned aerial systems. Front. Mar. Sci. 2018, 5, 316. [Google Scholar] [CrossRef]
- Landeo-Yauri, S.S.; Ramos, E.A.; Castelblanco-Martínez, D.N.; Niño-Torres, C.A.; Searle, L. Using small drones to photo-identify Antillean manatees: A novel method for monitoring an endangered marine mammal in the Caribbean Sea. Endanger. Spec. Res. 2020, 41, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Cleguer, C.; Tyne, J.; Wieser, M.; Kelly, N.; Peel, D.; Hodgson, A. Development of a novel drone-based method to survey marine megafauna at local spatial scales. Lessons learnt from a dugong drone survey in the Pilbara, Western Australia. In Proceedings of the World Marine Mammal Conference, Barcelona, Spain, 9–12 December 2019. [Google Scholar]
- Landeo-Yauri, S.S.; Castelblanco-Martínez, N.; Niño-Torres, C.A.; Ramos, E.; Hénaut, Y.; Cuevas-Domínguez, J.D. Protocol proposal: Using small drones for detection of Antillean manatees on the Mexican Caribbean. In Proceedings of the XII Congress of the Latin American Society of Specialists in Aquatic Mammals, Lima, Perú, 5–8 November 2018. [Google Scholar]
- Edwards, H.H.; Hostetler, J.A.; Stith, B.M.; Martin, J. Using an Unmanned Aerial System (UAS) to monitor abundance and distribution of Florida manatees (Trichechus manatus latirostris) at warmwater aggregation sites. In Proceedings of the Ninth International Sirenian Symposium, Barcelona, Spain, 8 December 2018. [Google Scholar]
- Landeo-Yauri, S. Using light UAVs as Complementary Tools to Study and Conserve Antillean Manatees in the Mexican Caribbean. Master’s Thesis, National Autonomous University of Mexico, Mexico, Mexico, 2019. [Google Scholar]
- Maire, F.; Mejias, L.; Hodgson, A.; Duclos, G. Detection of dugongs from unmanned aerial vehicles. In Proceedings of the 2013 IEEE/RSJ International Conference on Intelligent Robots and Systems, Tokyo, Japan, 3–7 November 2013; pp. 2750–2756. [Google Scholar]
- Hodgson, A.; Cleguer, C.; Maire, F.; Wieser, M.; Tyne, J.; Matthews, S.; Kwan, D. Using drones, artificial intelligence and geospatial techniques to revolutionise dugong and seagrass protection globally. In Proceedings of the World Marine Mammal Conference, Barcelona, Spain, 9–12 December 2019. [Google Scholar]
- Beck, C.; Clark, A. Individual identification of sirenians. In Sirenian Conservation: Issues and Strategies in Developing Countries; Hines, E.M., III, Aragones, L.V., Mignucci-Giannoni, A.A., Marmontel, M., Eds.; University Press of Florida: Gainesville, FL, USA, 2012; pp. 133–138. [Google Scholar]
- Beck, C.A.; Bonde, R.K.; Rathbun, G.B. Analyses of propeller wounds on manatees in Florida. J. Wildl. Manag. 1982, 46, 531–535. [Google Scholar] [CrossRef]
- Christiansen, F.; Sprogis, K.R.; Gross, J.; Castrillon, J.; Warick, H.A.; Leunissen, E.; Nash, S.B. Variation in outer blubber lipid concentration does not reflect morphological body condition in humpback whales. J. Exp. Biol. 2020, 223. [Google Scholar] [CrossRef] [PubMed]
- Landeo-Yauri, S.S.; Castelblanco-Martínez, N.; Niño-Torres, C.A.; Hénaut, Y.; Ramos, E. First assessment of Antillean manatee watching tourism in Mexico: A case study in the Sian Ka’an Biosphere Reserve, Quintana Roo. In Proceedings of the XII Congress of the Latin American Society of Specialists in Aquatic Mammals, Lima, Perú, 5–8 November 2018. [Google Scholar]
- Bonde, R.K.; Garrett, A.; Belanger, M.; Askin, N.; Tan, L.; Wittnich, C. Biomedical health assessments of the Florida manatee in Crystal River—Providing opportunities for training during the capture, handling, and processing of this endangered aquatic mammal. J. Mar. Anim. Ecol. 2012, 5, 17–28. [Google Scholar]
- Lanyon, J.M.; Slade, R.W.; Sneath, H.L.; Broderick, D.; Kirkwood, J.M.; Limpus, D.; Limpus, C.J.; Jessop, T. A method for capturing dugongs (Dugong dugon) in open water. Aquat. Mamm. 2006, 32, 196. [Google Scholar] [CrossRef]
- Lanyon, J.M.; Sneath, H.L.; Long, T.; Bonde, R.K. Physiological Response of Wild Dugongs (Dugong dugon) to Out-of-Water Sampling for Health Assessment. Aquat. Mamm. 2010, 36, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Alencar, A.E.B.; Attademo, F.L.N.; Normande, I.C.; Luna, F.d.O. Uso de Aeronaves não Tripuladas (DRONES) para Pesquisa e Monitoramento de Peixe-Boi-Marinho e seu Habitat; ICMBio: Brasilia, Brazil, 2020; p. 45.
- Cleguer, C.; Garrigue, C. The dugongs’ use of the Vavouto mining port in the lagoonal reefs of New Caledonia—Preliminary results. In Proceedings of the Ninth International Sirenian Symposium, Barcelona, Spain, 8 December 2019. [Google Scholar]
- Christiansen, F.; Sironi, M.; Moore, M.J.; Di Martino, M.; Ricciardi, M.; Warick, H.A.; Irschick, D.J.; Gutierrez, R.; Uhart, M.M. Estimating body mass of free-living whales using aerial photogrammetry and 3D volumetrics. Methods Ecol. Evol. 2019, 10, 2034–2044. [Google Scholar] [CrossRef]
- Scharffenberg, K.; Whalen, D.; Marcoux, M.; Iacozza, J.; Davoren, G.; Loseto, L. Environmental drivers of beluga whale Delphinapterus leucas habitat use in the Mackenzie Estuary, Northwest Territories, Canada. Mar. Ecol. Prog. Ser. 2019, 626, 209–226. [Google Scholar] [CrossRef]
- Christiansen, F.; Dujon, A.M.; Sprogis, K.R.; Arnould, J.P.; Bejder, L. Noninvasive unmanned aerial vehicle provides estimates of the energetic cost of reproduction in humpback whales. Ecosphere 2016, 7, e01468. [Google Scholar] [CrossRef]
- Geoghegan, J.L.; Pirotta, V.; Harvey, E.; Smith, A.; Buchmann, J.P.; Ostrowski, M.; Eden, J.-S.; Harcourt, R.; Holmes, E.C. Virological sampling of inaccessible wildlife with drones. Viruses 2018, 10, 300. [Google Scholar] [CrossRef] [Green Version]
- Oliveira-da-Costa, M.; Marmontel, M.; da-Rosa, D.S.; Coelho, A.; Wich, S.; Mosquera-Guerra, F.; Trujillo, F. Effectiveness of unmanned aerial vehicles to detect Amazon dolphins. Oryx 2020, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Fettermann, T.; Fiori, L.; Bader, M.; Doshi, A.; Breen, D.; Stockin, K.A.; Bollard, B. Behaviour reactions of bottlenose dolphins (Tursiops truncatus) to multirotor Unmanned Aerial Vehicles (UAVs). Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Orbach, D.N.; Eaton, J.; Fiori, L.; Piwetz, S.; Weir, J.S.; Würsig, M.; Würsig, B. Mating patterns of dusky dolphins (Lagenorhynchus obscurus) explored using an unmanned aerial vehicle. Mar. Mamm. Sci. 2020. [Google Scholar] [CrossRef]
- Giles, A.; Butcher, P.; Colefax, A.P.; Pagendam, D.E.; Mayior, M.; Kelaher, B.P. Responses of bottlenose dolphins (Tursiops spp.) to small drones. Aquat. Conserv. 2020, in press. [Google Scholar]
- Centelleghe, C.; Carraro, L.; Gonzalvo, J.; Rosso, M.; Esposti, E.; Gili, C.; Bonato, M.; Pedrotti, D.; Cardazzo, B.; Povinelli, M.; et al. The use of Unmanned Aerial Vehicles (UAVs) to sample the blow microbiome of small cetaceans. PLoS ONE 2020, 15, e0235537. [Google Scholar] [CrossRef]
- Raudino, H.C.; Tyne, J.A.; Smith, A.; Ottewell, K.; McArthur, S.; Kopps, A.M.; Chabanne, D.; Harcourt, R.G.; Pirotta, V.; Waples, K. Challenges of collecting blow from small cetaceans. Ecosphere 2019, 10, e02901. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.E.; Sykora-Bodie, S.T.; Bloodworth, B.; Pack, S.M.; Spradlin, T.R.; LeBoeuf, N.R. Assessment of known impacts of unmanned aerial systems (UAS) on marine mammals: Data gaps and recommendations for researchers in the United States. J. Unmanned Veh. Syst. 2016, 4, 31–44. [Google Scholar] [CrossRef] [Green Version]
- Goldbogen, J.A.; Cade, D.E.; Wisniewska, D.M.; Potvin, J.; Segre, P.S.; Savoca, M.S.; Hazen, E.L.; Czapanskiy, M.; Kahane-Rapport, S.R.; DeRuiter, S.L.; et al. Why whales are big but not bigger: Physiological drivers and ecological limits in the age of ocean giants. Science 2019, 366, 1367–1372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popper, A.N.; Fay, R.R.; Au, W. Hearing by Whales and Dolphins; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2000; Volume 12. [Google Scholar]
- Robbins, J.; Dalla Rosa, L.; Allen, J.M.; Mattila, D.K.; Secchi, E.R.; Friedlaender, A.S.; Stevick, P.T.; Nowacek, D.P.; Steel, D. Return movement of a humpback whale between the Antarctic Peninsula and American Samoa: A seasonal migration record. Endanger. Spec. Res. 2011, 13, 117–121. [Google Scholar] [CrossRef]
- Johnston, D.; Friedlaender, A.; Read, A.; Nowacek, D. Initial density estimates of humpback whales Megaptera novaeangliae in the inshore waters of the western Antarctic Peninsula during the late autumn. Endanger. Spec. Res. 2012, 18, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Nowacek, D.P.; Bröker, K.; Donovan, G.; Gailey, G.; Racca, R.; Reeves, R.R.; Vedenev, A.I.; Weller, D.W.; Southall, B.L. Responsible practices for minimizing and monitoring environmental impacts of marine seismic surveys with an emphasis on marine mammals. Aquat. Mamm. 2013, 39, 356. [Google Scholar] [CrossRef] [Green Version]
- Johnston, D.W. Unoccupied aircraft systems in marine science and conservation. Ann. Rev. Mar. Sci. 2019, 11, 439–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, L.G.; Barlow, D.R.; Chandler, T.E.; Burnett, J.D. Insight into the kinematics of blue whale surface foraging through drone observations and prey data. PeerJ 2020, 8, e8906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gough, W.T.; Segre, P.S.; Bierlich, K.; Cade, D.E.; Potvin, J.; Fish, F.E.; Dale, J.; di Clemente, J.; Friedlaender, A.S.; Johnston, D.W. Scaling of swimming performance in baleen whales. J. Exp. Biol. 2019, 222. [Google Scholar] [CrossRef] [PubMed]
- Bird, C.; Bierlich, K.; Dale, J.; Friedlaender, A.; Goldbogen, J.A.; Johnston, D.W. A comparison of percent dorsal scar cover between populations of humpback whales (Megaptera novaeangliae) off California and the Western Antarctic Peninsula. In Proceedings of the World Marine Mammal Conference, Barcelona, Spain, 7–12 December 2019. [Google Scholar]
- Apprill, A.; Miller, C.A.; Moore, M.J.; Durban, J.W.; Fearnbach, H.; Barrett-Lennard, L.G. Extensive core microbiome in drone-captured whale blow supports a framework for health monitoring. MSystems 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, T.W.; Hauser, N.; Cassel, S.; Klaus, K.F.; Fettermann, T.; Key, N. Doctor Drone: Non-invasive Measurement of Humpback Whale Vital Signs Using Unoccupied Aerial System Infrared Thermography. Front. Mar. Sci. 2019, 6, 466. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, F.; Nielsen, M.L.; Charlton, C.; Bejder, L.; Madsen, P.T. Southern right whales show no behavioral response to low noise levels from a nearby unmanned aerial vehicle. Mar. Mamm. Sci. 2020. [Google Scholar] [CrossRef]
- Domínguez-Sánchez, C.A.; Acevedo-Whitehouse, K.A.; Gendron, D. Effect of drone-based blow sampling on blue whale (Balaenoptera musculus) behavior. Mar. Mamm. Sci. 2018, 34, 841–850. [Google Scholar] [CrossRef]
- Hodgson, A.; Peel, D.; Kelly, N. Unmanned aerial vehicles for surveying marine fauna: Assessing detection probability. Ecol. Appl. 2017, 27, 1253–1267. [Google Scholar] [CrossRef]
- Angliss, R.; Ferguson, M.; Hall, P.; Helker, V.; Kennedy, A.; Sformo, T. Comparing manned to unmanned aerial surveys for cetacean monitoring in the Arctic: Methods and operational results. J. Unmanned Veh. Syst. 2018, 6, 109–127. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, M.; Angliss, R.; Kennedy, A.; Lynch, B.; Willoughby, A.; Helker, V.; Brower, A.; Clarke, J. Performance of manned and unmanned aerial surveys to collect visual data and imagery for estimating arctic cetacean density and associated uncertainty. J. Unmanned Veh. Syst. 2018, 6, 128–154. [Google Scholar] [CrossRef]
- Seymour, A.; Dale, J.; Hammill, M.; Halpin, P.; Johnston, D. Automated detection and enumeration of marine wildlife using unmanned aircraft systems (UAS) and thermal imagery. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Durban, J.; Fearnbach, H.; Barrett-Lennard, L.; Perryman, W.; Leroi, D. Photogrammetry of killer whales using a small hexacopter launched at sea. J. Unmanned Veh. Syst. 2015, 3, 131–135. [Google Scholar] [CrossRef]
- Aleixo, F.; O’Callaghan, S.A.; Ducla Soares, L.; Nunes, P.; Prieto, R. AragoJ: A free, open-source software to aid single camera photogrammetry studies. Methods Ecol. Evol. 2020, 11, 670–677. [Google Scholar] [CrossRef] [Green Version]
- Ventura, D.; Bruno, M.; Lasinio, G.J.; Belluscio, A.; Ardizzone, G. A low-cost drone based application for identifying and mapping of coastal fish nursery grounds. Estuar. Coast. Shelf Sci. 2016, 171, 85–98. [Google Scholar] [CrossRef]
- Ventura, D.; Bonifazi, A.; Gravina, M.F.; Belluscio, A.; Ardizzone, G. Mapping and classification of ecologically sensitive marine habitats using unmanned aerial vehicle (UAV) imagery and object-based image analysis (OBIA). Remote Sens. 2018, 10, 1331. [Google Scholar] [CrossRef] [Green Version]
- Kalacska, M.; Lucanus, O.; Sousa, L.; Vieira, T.; Arroyo-Mora, J.P. Freshwater fish habitat complexity mapping using above and underwater structure-from-motion photogrammetry. Remote Sens. 2018, 10, 1912. [Google Scholar] [CrossRef] [Green Version]
- Ahilan, T.; Adityan, V.A.; Kailash, S. Efficient Utilization of Unmanned Aerial Vehicle (UAV) for Fishing through Surveillance for Fishermen. Int. J. Aerosp. Mec. Eng. 2015, 9, 1468–1471. [Google Scholar]
- Harris, J.M.; Nelson, J.A.; Rieucau, G.; Broussard, W.P., III. Use of drones in fishery science. Trans. Am. Fish. Soc. 2019, 148, 687–697. [Google Scholar] [CrossRef]
- Nowlin, M.B.; Roady, S.E.; Newton, E.; Johnston, D.W. Applying Unoccupied Aircraft Systems to study human behavior in marine science and conservation programs. Front. Mar. Sci. 2019, 6, 567. [Google Scholar] [CrossRef] [Green Version]
- Allan, B.M.; Ierodiaconou, D.; Nimmo, D.G.; Herbert, M.; Ritchie, E.G. Free as a drone: Ecologists can add UAVs to their toolbox. Front. Ecol. Environ. 2015, 13, 354–355. [Google Scholar] [CrossRef]
- Ierodiaconou, D.; Schimel, A.C.; Kennedy, D.M. A new perspective of storm bite on sandy beaches using unmanned aerial vehicles. Z. Für Geomorphol. Suppl. Issues 2016, 60, 123–137. [Google Scholar] [CrossRef]
- Moskowitz, E.E.; Siegel-Richman, Y.M.; Hertner, G.; Schroeppel, T. Aerial drone misadventure: A novel case of trauma resulting in ocular globe rupture. Am. J. Ophthalmol. Case Rep. 2018, 10, 35–37. [Google Scholar] [CrossRef]
- Koh, C.H.; Low, K.; Li, L.; Zhao, Y.; Deng, C.; Tan, S.K.; Chen, Y.; Yeap, B.C.; Li, X. Weight threshold estimation of falling UAVs (Unmanned Aerial Vehicles) based on impact energy. Trans. Res. Part C Emerg. Technol. 2018, 93, 228–255. [Google Scholar] [CrossRef]
- Wang, Q.; Mao, B.; Stoliarov, S.I.; Sun, J. A review of lithium ion battery failure mechanisms and fire prevention strategies. Prog. Energy Combust. Sci. 2019, 73, 95–131. [Google Scholar] [CrossRef]
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raoult, V.; Colefax, A.P.; Allan, B.M.; Cagnazzi, D.; Castelblanco-Martínez, N.; Ierodiaconou, D.; Johnston, D.W.; Landeo-Yauri, S.; Lyons, M.; Pirotta, V.; et al. Operational Protocols for the Use of Drones in Marine Animal Research. Drones 2020, 4, 64. https://doi.org/10.3390/drones4040064
Raoult V, Colefax AP, Allan BM, Cagnazzi D, Castelblanco-Martínez N, Ierodiaconou D, Johnston DW, Landeo-Yauri S, Lyons M, Pirotta V, et al. Operational Protocols for the Use of Drones in Marine Animal Research. Drones. 2020; 4(4):64. https://doi.org/10.3390/drones4040064
Chicago/Turabian StyleRaoult, Vincent, Andrew P Colefax, Blake M. Allan, Daniele Cagnazzi, Nataly Castelblanco-Martínez, Daniel Ierodiaconou, David W. Johnston, Sarah Landeo-Yauri, Mitchell Lyons, Vanessa Pirotta, and et al. 2020. "Operational Protocols for the Use of Drones in Marine Animal Research" Drones 4, no. 4: 64. https://doi.org/10.3390/drones4040064
APA StyleRaoult, V., Colefax, A. P., Allan, B. M., Cagnazzi, D., Castelblanco-Martínez, N., Ierodiaconou, D., Johnston, D. W., Landeo-Yauri, S., Lyons, M., Pirotta, V., Schofield, G., & Butcher, P. A. (2020). Operational Protocols for the Use of Drones in Marine Animal Research. Drones, 4(4), 64. https://doi.org/10.3390/drones4040064