

The Effects of Different Carbon Sources on Water Quality, Growth Performance, Hematology, Immune, and Antioxidant Status in Cultured Nile Tilapia with Biofloc Technology

 ,
,  ,
,  ,
,  ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Analysis of Water Quality
2.3. Growth Performance and Survival
2.4. Blood Sampling
2.5. Hematological Parameters
2.6. Humoral Non-Specific Immune Parameters
2.7. Antioxidant Parameters
2.8. Statistical Analysis
3. Results
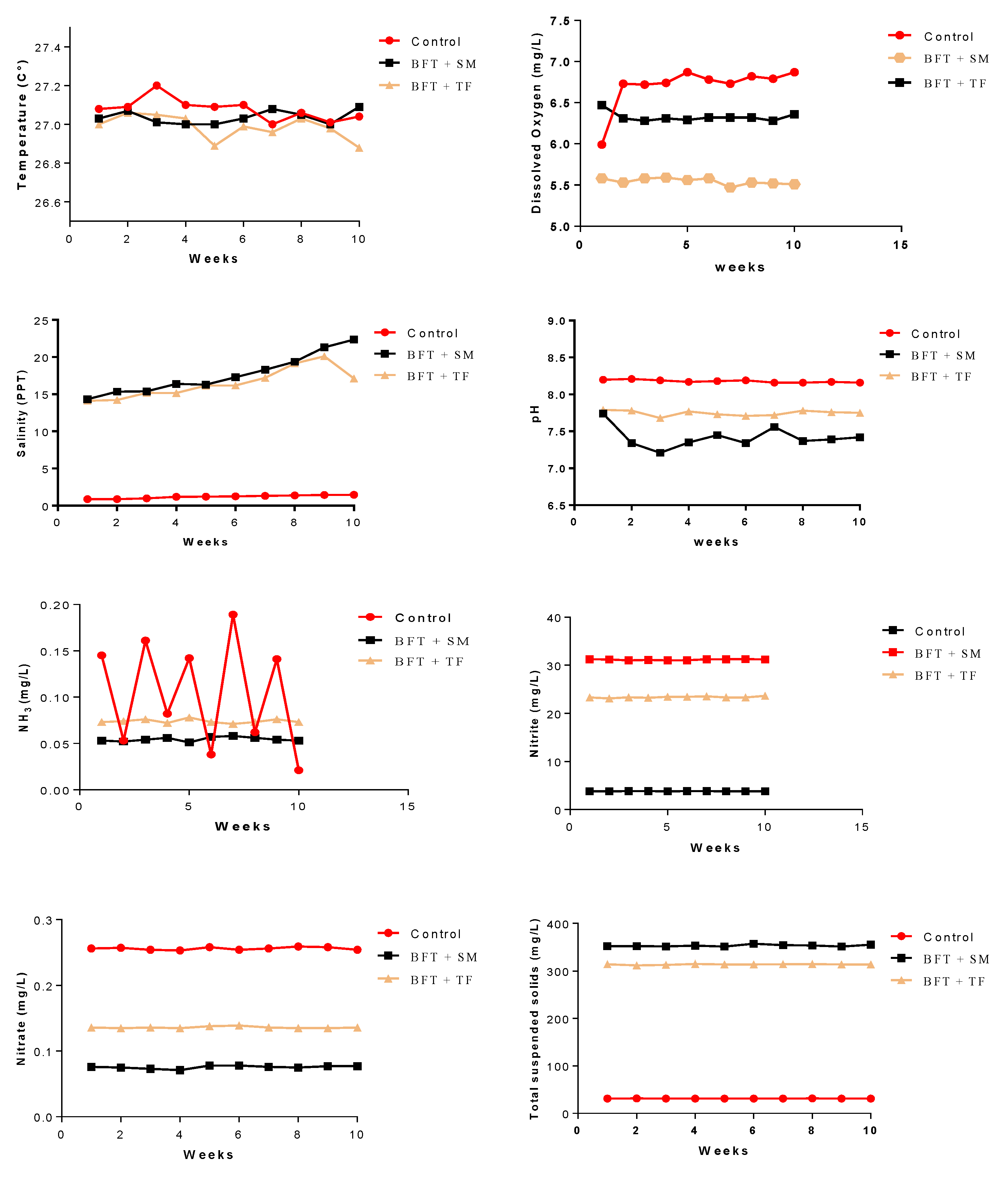
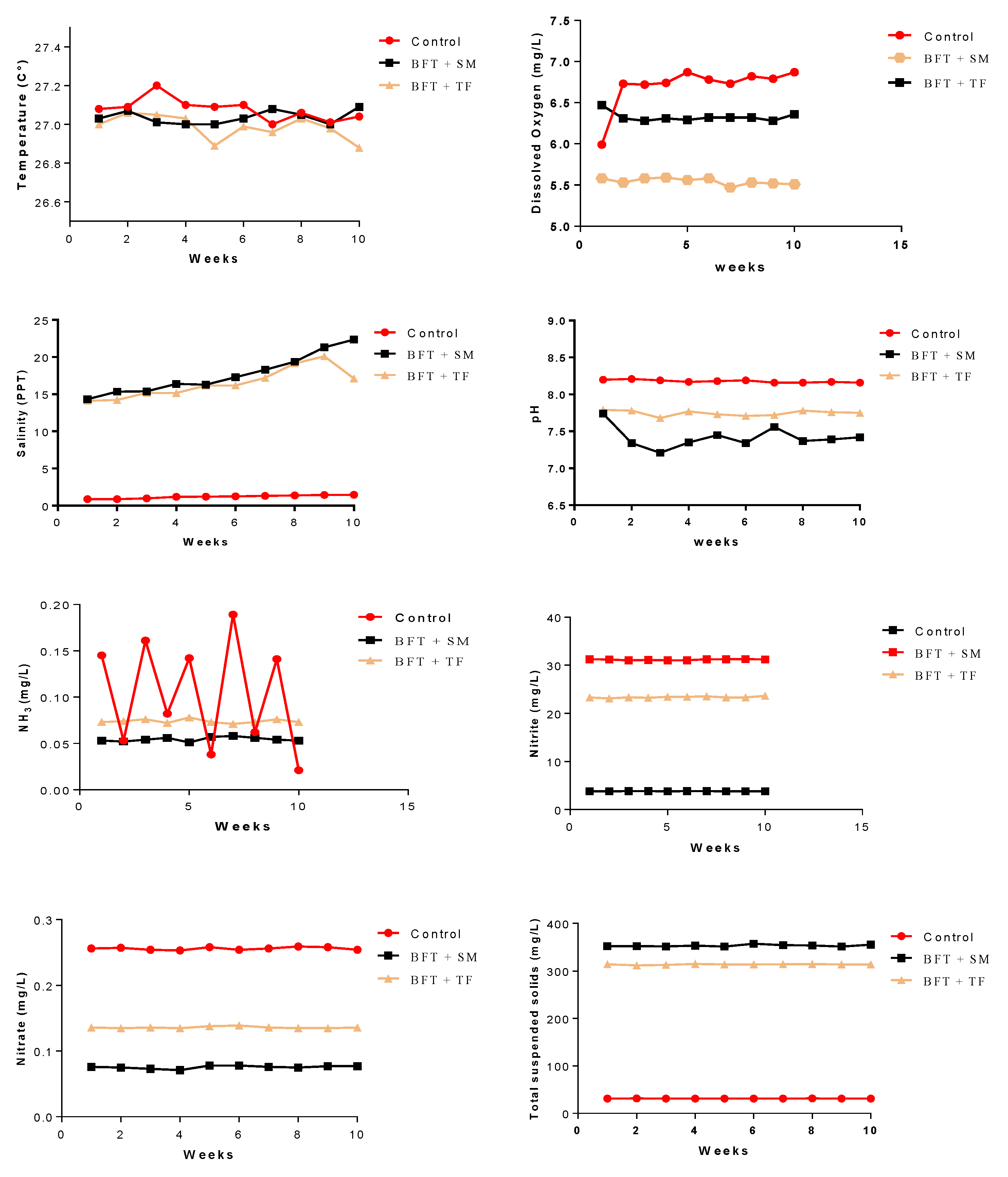
3.1. Water Quality Characteristics
3.2. Fish Growth Performance
3.3. Hematological Parameters
3.4. Humoral Non-Specific Immune Parameters
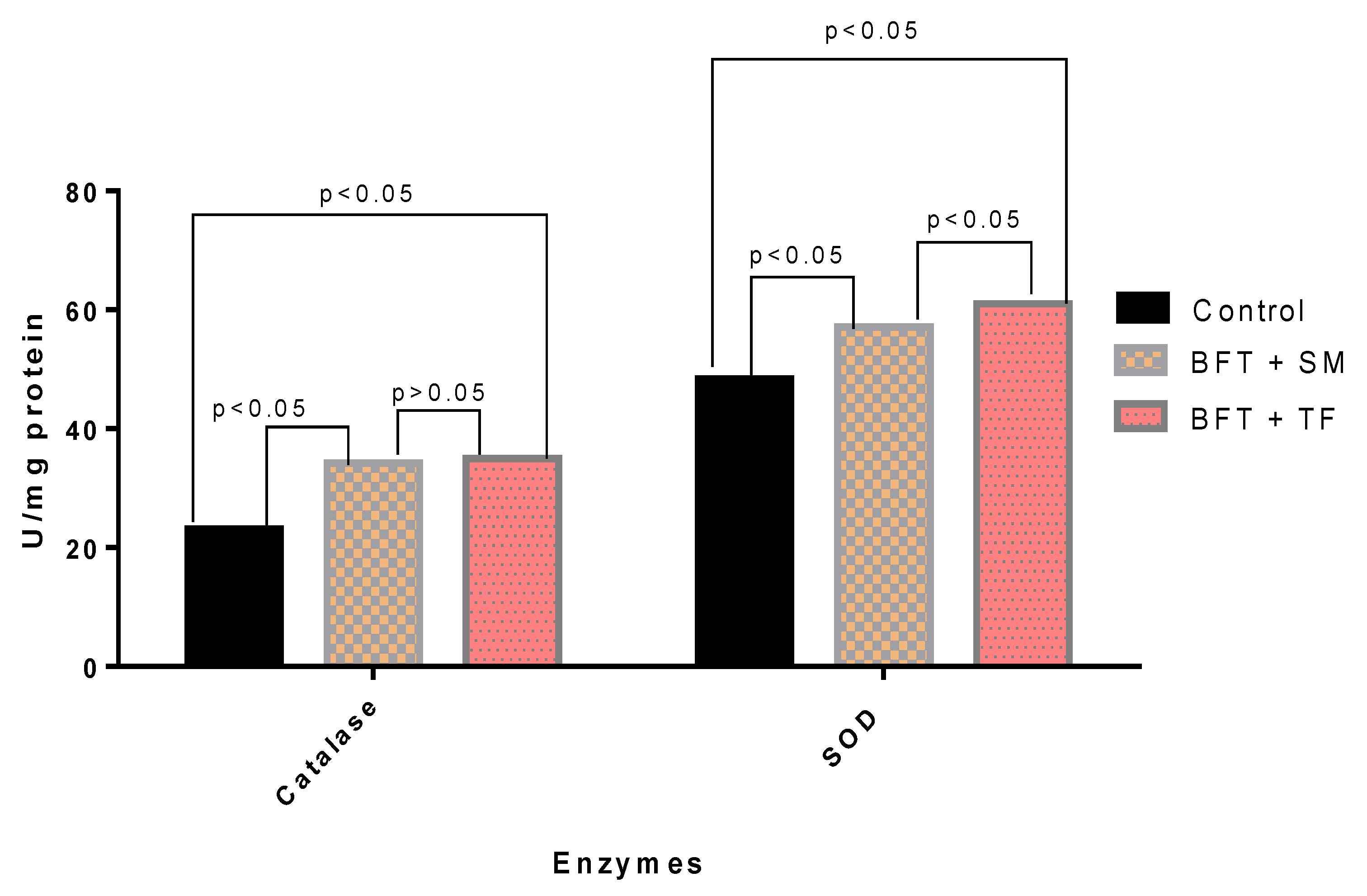
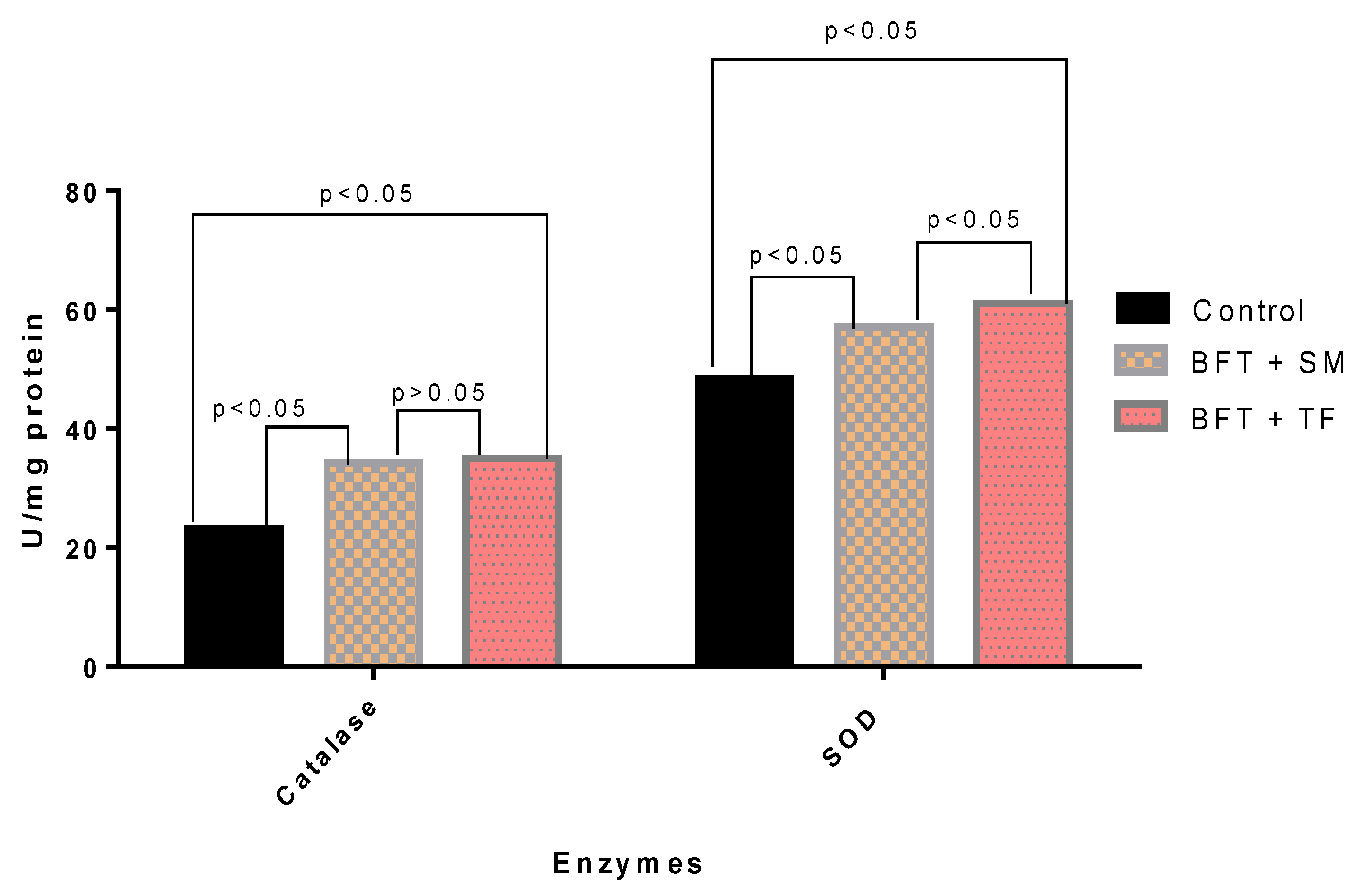
3.5. Antioxidant Enzymatic Activities
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tahergorabi, R.; Matak, K.E.; Jaczynski, J. Fish protein isolate: Development of functional foods with nutraceutical ingredients. J. Funct. Foods 2015, 18, 746–756. [Google Scholar] [CrossRef]
- Boyd, C.E.; D’Abramo, L.R.; Glencross, B.D.; Huyben, D.C.; Juarez, L.M.; Lockwood, G.S.; McNevin, A.A.; Tacon, A.G.; Teletchea, F.; Tomasso, J.R., Jr.; et al. Achieving sustainable aquaculture: Historical and current perspectives and future needs and challenges. J. World Aquac. Soc. 2020, 51, 578–633. [Google Scholar] [CrossRef]
- Ching, J.J.; Shuib, A.S.; Abdullah, N.; Majid, N.A.; Taufek, N.M.; Sutra, J.; Azmai, M.N.A. Hot water extract of Pleurotus pulmonarius stalk waste enhances innate immune response and immune-related gene expression in red hybrid tilapia Oreochromis sp. following challenge with pathogen-associated molecular patterns. Fish Shellfish Immunol. 2021, 116, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Habib, S.S.; Batool, A.I.; Rehman, M.F.U.; Naz, S. Comparative analysis of the haemato-biochemical parameters and growth characteristics of Oreochromis niloticus (Nile tilapia) cultured under different feed and habitats (biofloc technology and earthen pond system). Aquac. Res. 2022, 53, 6184–6192. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Sharifinia, M. Biofloc technology as a promising tool to improve aquaculture production. Rev. Aquac. 2020, 12, 1836–1850. [Google Scholar] [CrossRef]
- Khanjani, M.H.; Sharifinia, M.; Hajirezaee, S. Recent progress towards the application of biofloc technology for tilapia farming. Aquaculture 2022, 552, 738021. [Google Scholar] [CrossRef]
- Vethathirri, R.S.; Santillan, E.; Wuertz, S. Microbial community-based protein production from wastewater for animal feed applications. Bioresou. Technol. 2021, 341, 125723. [Google Scholar] [CrossRef]
- Deng, M.; Chen, J.; Gou, J.; Hou, J.; Li, D.; He, X. The effect of different carbon sources on water quality, microbial community and structure of biofloc systems. Aquaculture 2018, 482, 103–110. [Google Scholar] [CrossRef]
- Li, J.; Liu, G.; Li, C.; Deng, Y.; Tadda, M.A.; Lan, L.; Zhu, S.; Liu, D. Effects of different solid carbon sources on water quality, biofloc quality and gut microbiota of Nile tilapia (Oreochromis niloticus) larvae. Aquaculture 2018, 495, 919–931. [Google Scholar] [CrossRef]
- Wei, Y.; Liao, S.A.; Wang, A.L. The effect of different carbon sources on the nutritional composition, microbial community and structure of bioflocs. Aquaculture 2016, 465, 88–93. [Google Scholar] [CrossRef]
- Rohani, M.F.; Islam, S.M.; Hossain, M.K.; Ferdous, Z.; Siddik, M.A.; Nuruzzaman, M.; Padeniya, U.; Brown, C.; Shahjahan, M. Probiotics, prebiotics and synbiotics improved the functionality of aquafeed: Upgrading growth, reproduction, immunity and disease resistance in fish. Fish Immunol. 2022, 120, 569–589. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Pan, L.; Huang, F.; Wang, C.; Xu, W. Effects of different carbon sources on bioactive compound production of biofloc, immune response, antioxidant level, and growth performance of Litopenaeus vannamei in zero-water exchange culture tanks. J. World Aquac. Soc. 2016, 47, 566–576. [Google Scholar] [CrossRef]
- Elayaraja, S.; Mabrok, M.; Algammal, A.; Sabitha, E.; Rajeswari, M.V.; Zágoršek, K.; Ye, Z.; Zhu, S.; Rodkhum, C. Potential influence of jaggery-based biofloc technology at different C: N ratios on water quality, growth performance, innate immunity, immune-related genes expression profiles, and disease resistance against Aeromonas hydrophila in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 107, 118–128. [Google Scholar] [PubMed]
- Khanjani, M.H.; Sharifinia, M.; Emerenciano, M.G.C. A detailed look at the impacts of biofloc on immunological and hematological parameters and improving resistance to diseases. Fish Shellfish Immunol. 2023, 137, 108796. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.T.; Esteban, M.Á. Effects of carbon sources and plant protein levels in a biofloc system on growth performance, and the immune and antioxidant status of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2017, 64, 202–209. [Google Scholar] [CrossRef]
- Bakhshi, F.; Najdegerami, E.H.; Manaffar, R.; Tokmechi, A.; Rahmani Farah, K.; Shalizar Jalali, A. Growth performance, haematology, antioxidant status, immune response and histology of common carp (Cyprinus carpio L.) fed biofloc grown on different carbon sources. Aquac. Res. 2018, 49, 393–403. [Google Scholar] [CrossRef]
- Avnimelech, Y. Biofloc Technology: A Practical Guide Book, 3rd ed.; World Aquaculture Society: San Diego, CA, USA, 2009. [Google Scholar]
- Avnimelech, Y. Carbon/nitrogen ratio as a control element in aquaculture systems. Aquaculture 1999, 176, 227–235. [Google Scholar] [CrossRef]
- Habib, S.S.; Naz, S.; Batool, A.I.; Rehman, F.U.; Ullah, M.; Kesbiç, O.S.; Maricchiolo, G.; Fazio, F. Effect of Different Anaesthetics on Hematology and Blood Biochemistry of Labeo rohita. Aquac. Stud. 2023, 24, AQUAST1195. [Google Scholar] [CrossRef]
- Goldenfarb, P.B.; Bowyer, F.P.; Hall, E.; Brosious, E. Reproducibility in the hematology laboratory: The microhematocrit determination. Am. J. Clin. Pathol. 1971, 56, 35–39. [Google Scholar] [CrossRef]
- Schütt, D.A.; Lehmann, J.; Goerlich, R.; Hamers, R. Haematology of swordtail, Xiphophorus helleri. I: Blood parameters and light microscopy of blood cells. J. Appl. Ichthyol. 1997, 13, 83–89. [Google Scholar] [CrossRef]
- Gornall, A.G.; Bardawill, C.J.; David, M.M. Determination of serum proteins by means of the biuret reaction. J. Biol. Chem. 1949, 177, 751–766. [Google Scholar] [CrossRef]
- Doumas, B.T.; Watson, W.A.; Biggs, H.G. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta 1971, 31, 87–96. [Google Scholar] [CrossRef]
- Siwicki, A.K.; Anderson, D.P. Nonspecific Defence Mechanisms Assay in Fish: II. Potential Killing Activity of Neutrophils and Macrophages, Lysozyme Activity in Serum and Organs. In Disease Diagnosis and Prevention Methods; Siwicki, A.K., Anderson, D.P., Waluga, J., Eds.; Wydawnictwo Instytutu Rybactwa Strodladowego: Olsztyn, Poland, 1993; pp. 105–111. [Google Scholar]
- Sun, S.; Ye, J.; Chen, J.; Wang, Y.; Chen, L. Effect of dietary fish oil replacement by rapeseed oil on the growth, fatty acid composition and serum non-specific immunity response of fingerling black carp, Mylopharyngodon piceus. Aquac. Nutr. 2011, 17, 441–450. [Google Scholar]
- Luck, H. Catalase. In Method of Enzymatic Analysis; Bergmayer, M.V., Ed.; Verlag Chemic/Academic Press: New York, NY, USA, 1974; p. 885. [Google Scholar]
- Pedrajas, J.R.; Peinado, J.; Lopez-Barea, J. Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of Cu, Zn-superoxide dismutase as potential biomarkers. Chem. Biol. Interact. 1995, 98, 267–282. [Google Scholar] [PubMed]
- El-Sayed, A.F.M. Use of biofloc technology in shrimp aquaculture: A comprehensive review, with emphasis on the last decade. Rev. Aquac. 2021, 13, 676–705. [Google Scholar]
- Soliman, A.M.; Abdel-Tawwab, M. Effects of different carbon sources on water quality, biofloc quality, and the productivity of Nile tilapia reared in biofloc-based ponds. Ann. Anim. Sci. 2022, 22, 1281–1289. [Google Scholar]
- Khanjani, M.H.; Alizadeh, M.; Sharifinia, M. Effects of different carbon sources on water quality, biofloc quality, and growth performance of Nile tilapia (Oreochromis niloticus) fingerlings in a heterotrophic culture system. Aquac. Int. 2021, 29, 307–321. [Google Scholar]
- Hostins, B.; Braga, A.; Lopes, D.L.; Wasielesky, W.; Poersch, L.H. Effect of temperature on nursery and compensatory growth of pink shrimp Farfantepenaeus brasiliensis reared in a super-intensive biofloc system. Aquac. Eng. 2015, 66, 62–67. [Google Scholar]
- Wilen, B.M.; Nielsen, J.L.; Keiding, K.; Nielsen, P.H. Influence of microbial activity on the stability of activated sludge flocs. Colloids Surf. B Biointerfaces 2000, 18, 145–156. [Google Scholar]
- Krishna, C.; Van Loosdrecht, M.C.M. Effect of temperature on storage polymers and settleability of activated sludge. Water Res. 1999, 33, 2374–2382. [Google Scholar]
- De Schryver, P.; Crab, R.; Defoirdt, T.; Boon, N.; Verstraete, W. The basics of bio-flocs technology: The added value for aquaculture. Aquaculture 2008, 277, 125–137. [Google Scholar]
- Martins, A.M.P.; Heijnen, J.J.; van Loosdrecht, M.C.M. Effect of dissolved oxygen concentration on sludge settleability. Appl. Microbiol. Biotechnol. 2003, 62, 586–593. [Google Scholar]
- Shamsuddin, M.; Hossain, M.B.; Rahman, M.; Kawla, M.S.; Shufol, M.B.A.; Rashid, M.M.; Asadujjaman, M.; Rakib, M.R.J. Application of Biofloc Technology for the culture of Heteropneustes fossilis (Bloch) in Bangladesh: Stocking density, floc volume, growth performance, and profitability. Aquac. Int. 2022, 30, 1047–1070. [Google Scholar]
- Luo, G.; Gao, Q.; Wang, C.; Liu, W.; Sun, D.; Li, L.; Tan, H. Growth, digestive activity, welfare, and partial cost-effectiveness of genetically improved farmed tilapia (Oreochromis niloticus) cultured in a recirculating aquaculture system and an indoor biofloc system. Aquaculture 2014, 422, 1–7. [Google Scholar]
- Mustapha, M.K.; Atolagbe, S.D. Tolerance level of different life stages of Nile tilapia Oreochromis niloticus (Linnaeus, 1758) to low pH and acidified waters. J. Basic Appl. Zool. 2018, 79, 1–6. [Google Scholar]
- Khanjani, M.H.; Alizadeh, M.; Mohammadi, M.; Sarsangi Aliabad, H. Biofloc system applied to Nile tilapia (Oreochromis niloticus) farming using different carbon sources: Growth performance, carcass analysis, digestive and hepatic enzyme activity. Iran. J. Fish. Sci. 2021, 20, 490–513. [Google Scholar]
- Xu, W.J.; Morris, T.C.; Samocha, T.M. Effects of C/N ratio on biofloc development, water quality, and performance of Litopenaeus vannamei juveniles in a biofloc-based, high-density, zero-exchange, outdoor tank system. Aquaculture 2016, 453, 169–175. [Google Scholar]
- Kamilya, D.; Debbarma, M.; Pal, P.; Kheti, B.; Sarkar, S.; Singh, S.T. Biofloc technology application in indoor culture of Labeo rohita (Hamilton, 1822) fingerlings: The effects on inorganic nitrogen control, growth and immunity. Chemosphere 2017, 182, 8–14. [Google Scholar]
- Azim, M.E.; Little, D.C. The biofloc technology (BFT) in indoor tanks: Water quality, biofloc composition, and growth and welfare of Nile tilapia (Oreochromis niloticus). Aquaculture 2008, 283, 29–35. [Google Scholar]
- Wang, G.; Yu, E.; Xie, J.; Yu, D.; Li, Z.; Luo, W.; Qiu, L.; Zheng, Z. Effect of C/N ratio on water quality in zero-water exchange tanks and the biofloc supplementation in feed on the growth performance of crucian carp, Carassius auratus. Aquaculture 2015, 443, 98–104. [Google Scholar]
- Adineh, H.; Naderi, M.; Hamidi, M.K.; Harsij, M. Biofloc technology improves growth, innate immune responses, oxidative status, and resistance to acute stress in common carp (Cyprinus carpio) under high stocking density. Fish Shellfish Immunol. 2019, 95, 440–448. [Google Scholar] [CrossRef]
- Long, L.; Yang, J.; Li, Y.; Guan, C.; Wu, F. Effect of biofloc technology on growth, digestive enzyme activity, hematology, and immune response of genetically improved farmed tilapia (Oreochromis niloticus). Aquaculture 2015, 448, 135–141. [Google Scholar] [CrossRef]
- García-Ríos, L.; Miranda-Baeza, A.; Coelho-Emerenciano, M.G.; Huerta-Rábago, J.A.; Osuna-Amarillas, P. Biofloc technology (BFT) applied to tilapia fingerlings production using different carbon sources: Emphasis on commercial applicatons. Aquaculture 2019, 502, 26–31. [Google Scholar] [CrossRef]
- Arnold, S.J.; Coman, F.E.; Jackson, C.J.; Groves, S.A. High-intensity, zero water-exchange production of juvenile tiger shrimp, Penaeus monodon: An evaluation of artificial substrates and stocking density. Aquaculture 2009, 293, 42–48. [Google Scholar] [CrossRef]
- Kishawy, A.T.; Sewid, A.H.; Nada, H.S.; Kamel, M.A.; El-Mandrawy, S.A.; Abdelhakim, T.M.; El-Murr, A.E.I.; Nahhas, N.E.; Hozzein, W.N.; Ibrahim, D. Mannanoligosaccharides as a carbon source in Biofloc boost dietary plant protein and water quality, growth, immunity and Aeromonas hydrophila resistance in Nile tilapia (Oreochromis niloticus). Animals 2020, 10, 1724. [Google Scholar] [CrossRef]
- Mahanand, S.S.; Moulick, S.; Rao, P.S. Water quality and growth of Rohu, Labeo rohita, in a biofloc system. J. Appl. Aquac. 2013, 25, 121–131. [Google Scholar] [CrossRef]
- Crab, R.; Kochva, M.; Verstraete, W.; Avnimelech, Y. Bio-flocs technology application in over-wintering of tilapia. Aquac. Eng. 2009, 40, 105–112. [Google Scholar] [CrossRef]
- Habib, S.S.; Naz, S.; Khalid, S.; Kanwal, R.; Ameer, I.; Khan, S.N.A.; Rehman, A.U.; Kousar, M.; Khan, S.U.; Nazir, N. Effect of white button mushroom (Agaricus bisporus) on immunity and haematological parameters of Oreochromis niloticus. Pak. J. Zool. 2022, 54, 785–790. [Google Scholar] [CrossRef]
- Amenyogbe, E.; Yang, E.J.; Xie, R.T.; Huang, J.S.; Chen, G. Influences of indigenous isolates Pantoea agglomerans RCS2 on growth, proximate analysis, haematological parameters, digestive enzyme activities, serum biochemical parameters, antioxidants activities, intestinal morphology, disease resistance, and molecular immune response in juvenile’s cobia fish (Rachycentron canadum). Aquaculture 2022, 551, 737942. [Google Scholar]
- Rebl, A.; Goldammer, T. Under control: The innate immunity of fish from the inhibitors’ perspective. Fish Shellfish Immunol. 2018, 77, 328–349. [Google Scholar] [CrossRef]
- Habib, S.S.; Naz, S.; Nawaz, S.; Ameer, I.; Khatoon, A.; Rehman, H.U.; Jawad, S.M.; Ali, H. Comparative analysis of hematological parameters of some farmed and wild fish species. Pak. J. Zool. 2021, 54, 591–598. [Google Scholar] [CrossRef]
- Habib, S.S.; Fazio, F.; Naz, S.; Arfuso, F.; Piccione, G.; Rehman, H.U.; Achakzai, W.M.; Uddin, M.N.; Rind, K.H.; Rind, N.A. Seasonal variations in haematological parameters and body composition of Labeo rohita (Rohu) and Cirrhinus mrigala (Mrigal carp) in River Indus, District Dera Ismail Khan, Pakistan. Turk. J. Fish. Aquat. Sci. 2021, 21, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Burgos-Aceves, M.A.; Lionetti, L.; Faggio, C. Multidisciplinary haematology as prognostic device in environmental and xenobiotic stress-induced response in fish. Sci. Total Environ. 2019, 670, 1170–1183. [Google Scholar] [CrossRef] [PubMed]
- Akrami, R.; Gharaei, A.; Mansour, M.R.; Galeshi, A. Effects of dietary onion (Allium cepa) powder on growth, innate immune response and hemato–biochemical parameters of beluga (Huso huso Linnaeus, 1754) juvenile. Fish Shellfish Immunol. 2015, 45, 828–834. [Google Scholar] [CrossRef]
- Chinnadurai, V.; Viswanathan, P.; Kalimuthu, K.; Vanitha, A.; Ranjitha, V.; Pugazhendhi, A. Comparative studies of phytochemical analysis and pharmacological activities of wild and micropropagated plant ethanol extracts of Manihot esculenta. Biocatal. Agric. Biotechnol. 2019, 19, 101166. [Google Scholar] [CrossRef]
- Chirra, M.; Newton, H.S.; Gawali, V.S.; Wise-Draper, T.M.; Chimote, A.A.; Conforti, L. How the potassium channel response of T lymphocytes to the tumor microenvironment shapes antitumor immunity. Cancers 2022, 14, 3564. [Google Scholar] [CrossRef]
- Verma, A.K.; Rani, A.B.; Rathore, G.; Saharan, N.; Gora, A.H. Growth, non-specific immunity and disease resistance of Labeo rohita against Aeromonas hydrophila in biofloc systems using different carbon sources. Aquaculture 2016, 457, 61–67. [Google Scholar]
- de Mello, M.M.M.; de Faria, C.D.F.P.; Zanuzzo, F.S.; Urbinati, E.C. β-glucan modulates cortisol levels in stressed pacu (Piaractus mesopotamicus) inoculated with heat-killed Aeromonas hydrophila. Fish Shellfish Immunol. 2019, 93, 1076–1083. [Google Scholar] [CrossRef]
- Uribe, C.; Folch, H.; Enríquez, R.; Moran, G.J.V.M. Innate and adaptive immunity in teleost fish: A review. Vet. Med. 2011, 56, 486. [Google Scholar] [CrossRef]
- Omar, N.F.; Hassan, S.A.; Yusoff, U.K.; Abdullah, N.A.P.; Wahab, P.E.M.; Sinniah, U.R. Phenolics, flavonoids, antioxidant activity and cyanogenic glycosides of organic and mineral-base fertilized cassava tubers. Molecules 2012, 17, 2378–2387. [Google Scholar] [CrossRef]
- Homayouni, H.; Kavoosi, G.; Nassiri, S.M. Physicochemical, antioxidant and antibacterial properties of dispersion made from tapioca and gelatinized tapioca starch incorporated with carvacrol. LWT 2017, 77, 503–509. [Google Scholar] [CrossRef]
- El-Beltagi, H.S.; Mohamed, H.I. Reactive oxygen species, lipid peroxidation and antioxidative defense mechanism. Not. Bot. Horti Agrobot. Cluj-Napoca 2013, 41, 44–57. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Composition | Fish Feed | Sugarcane Molasses | Tapioca Flour |
|---|---|---|---|
| Crude protein (% dw) | 32 ± 1.0 | 4.76 ± 0.12 | 10.78 ± 0.12 |
| Dry matter (%) | 84 ± 1.0 | 53.12 ± 0.38 | 90.13 ± 0.17 |
| Crude lipid (% dw) | 4.3 ± 1.0 | 0.2 ± 0.04 | 2.56 ± 0.02 |
| Ash (% dw) | 6.9 ± 1.0 | 7.87 ± 0.12 | 4.32 ± 0.05 |
| Fiber (% dw) | 1.8 ± 1.0 | 0.38 ± 0.06 | 6.12 ± 0.14 |
| Carbohydrate (% dw) | 26 ± 1.0 | 38.31 ± 0.21 | 65.21 ± 0.14 |
| Organic Materials | Daily Added (Grams) |
|---|---|
| Fish feed (32% protein) | 16.1 |
| Sugarcane molasses | 9.8 |
| Tapioca flour | 8.6 |
| C:N ratio | 15:1 |
| Parameters | Control | BFT + SM | BFT + TF |
|---|---|---|---|
| Water temperature (°C) | 26.99 ± 0.23 | 27.01 ± 0.18 | 27.01 ± 0.11 |
| Dissolved oxygen (mg L−1) | 6.97 ± 0.073 a | 5.58 ± 0.083 c | 6.31 ± 0.073 b |
| pH | 8.21 ± 0.057 a | 7.32 ± 0.075 c | 7.78 ± 0.054 b |
| Salinity (PPT) | 1.47 ± 0.61 b | 20.32 ± 11.21 a | 18.11 ± 13.13 a |
| Unionized ammonia (mg L−1) | 0.145 ± 0.016 a | 0.052 ± 0.026 b | 0.072 ± 0.025 ab |
| Nitrite (mg L−1) | 0.256 ± 0.021 a | 0.076 ± 0.017 b | 0.136 ± 0.023 b |
| Nitrate (mg L−1) | 3.81 ± 0.95 b | 31.24 ± 4.62 a | 23.33 ± 4.31 a |
| Total suspended solids (mg L−1) | 31.3 ± 5.27 b | 351.7 ± 70.44 a | 314.6 ± 62.67 a |
| Biofloc volume (mL) | 0.84 ± 0.13 c | 18.47 ± 2.87 b | 28.94 ± 4.12 a |
| Parameters | Control | BFT + SM | BFT + TF |
|---|---|---|---|
| Weight gain (%) | 66.03 ± 2.82 c | 87.15 ± 1.92 b | 98.61 ± 3.71 a |
| Feed intake (g kg fish−1) | 1001.21 ± 36.03 a | 867.37 ± 23.17 b | 869.31 ± 25.21 b |
| Feed conversion ratio (FCR) | 2.35 ± 0.16 a | 1.79 ± 0.06 b | 1.69 ± 0.07 c |
| Survival (%) | 95.83 ± 1.54 | 97.65 ± 1.02 | 99.01 ± 0.32 |
| Hematological Parameters | Control | BFT + SM | BFT + TF |
|---|---|---|---|
| RBCs (106/μL) | 1.32 ± 0.12 | 1.31 ± 0.11 | 1.33 ± 0.21 |
| WBCs (103/μL) | 27.41 ± 1.15 c | 32.21 ± 1.23 b | 37.43 ± 1.51 a |
| Hematocrit (%) | 27.21 ± 0.39 c | 33.11 ± 0.42 b | 35.13 ± 0.46 a |
| Haemoglobin (%) | 7.88 ± 0.32 c | 9.03 ± 0.21 b | 11.54 ± 0.25 a |
| Lymphocytes (%) | 30.65 ± 0.84 c | 34.52 ± 0.21 b | 37.32 ± 0.24 a |
| Monocytes (%) | 4.00 ± 0.21 | 4.88 ± 0.33 | 4.68 ± 0.31 |
| Neutrophils (%) | 62.49 ± 0.82 a | 58.62 ± 0.61 b | 56.16 ± 0.32 c |
| Parameters | Control | BFT + SM | BFT + TF |
|---|---|---|---|
| Globulin (g × dL−1) | 1.45 ± 0.07 b | 1.86 ± 0.04 a | 1.93 ± 0.04 a |
| Total protein (g × dL−1) | 3.57 ± 0.05 c | 3.84 ± 0.06 b | 4.18 ± 0.08 a |
| Albumin (g × dL−1) | 2.25 ± 0.02 c | 2.31 ± 0.04 b | 2.42 ± 0.04 a |
| Lysozyme (U × mg−1 protein) | 24.00 ± 2.01 c | 28.65 ± 1.42 b | 31.43 ± 1.38 a |
| Immunoglobulin (mg × dL−1) | 2.25 ± 0.16 c | 3.02 ± 0.06 b | 3.19 ± 0.08 a |
| ACH50 (U × mL−1) | 18.85 ± 1.71 c | 30.28 ± 1.03 b | 37.12 ± 1.65 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rind, K.H.; Habib, S.S.; Ujan, J.A.; Fazio, F.; Naz, S.; Batool, A.I.; Ullah, M.; Attaullah, S.; Khayyam, K.; Khan, K. The Effects of Different Carbon Sources on Water Quality, Growth Performance, Hematology, Immune, and Antioxidant Status in Cultured Nile Tilapia with Biofloc Technology. Fishes 2023, 8, 512. https://doi.org/10.3390/fishes8100512
Rind KH, Habib SS, Ujan JA, Fazio F, Naz S, Batool AI, Ullah M, Attaullah S, Khayyam K, Khan K. The Effects of Different Carbon Sources on Water Quality, Growth Performance, Hematology, Immune, and Antioxidant Status in Cultured Nile Tilapia with Biofloc Technology. Fishes. 2023; 8(10):512. https://doi.org/10.3390/fishes8100512
Chicago/Turabian StyleRind, Khalid Hussain, Syed Sikandar Habib, Javed Ahmed Ujan, Francesco Fazio, Saira Naz, Aima Iram Batool, Mujeeb Ullah, Sobia Attaullah, Khayyam Khayyam, and Khalid Khan. 2023. "The Effects of Different Carbon Sources on Water Quality, Growth Performance, Hematology, Immune, and Antioxidant Status in Cultured Nile Tilapia with Biofloc Technology" Fishes 8, no. 10: 512. https://doi.org/10.3390/fishes8100512
APA StyleRind, K. H., Habib, S. S., Ujan, J. A., Fazio, F., Naz, S., Batool, A. I., Ullah, M., Attaullah, S., Khayyam, K., & Khan, K. (2023). The Effects of Different Carbon Sources on Water Quality, Growth Performance, Hematology, Immune, and Antioxidant Status in Cultured Nile Tilapia with Biofloc Technology. Fishes, 8(10), 512. https://doi.org/10.3390/fishes8100512