
Physiological Effects of Recapture and Transport from Net-Cages in Lumpfish


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish
2.2. Experimental Set-Up
2.3. Blood Plasma Analysis
2.4. Statistical Analyses
3. Results
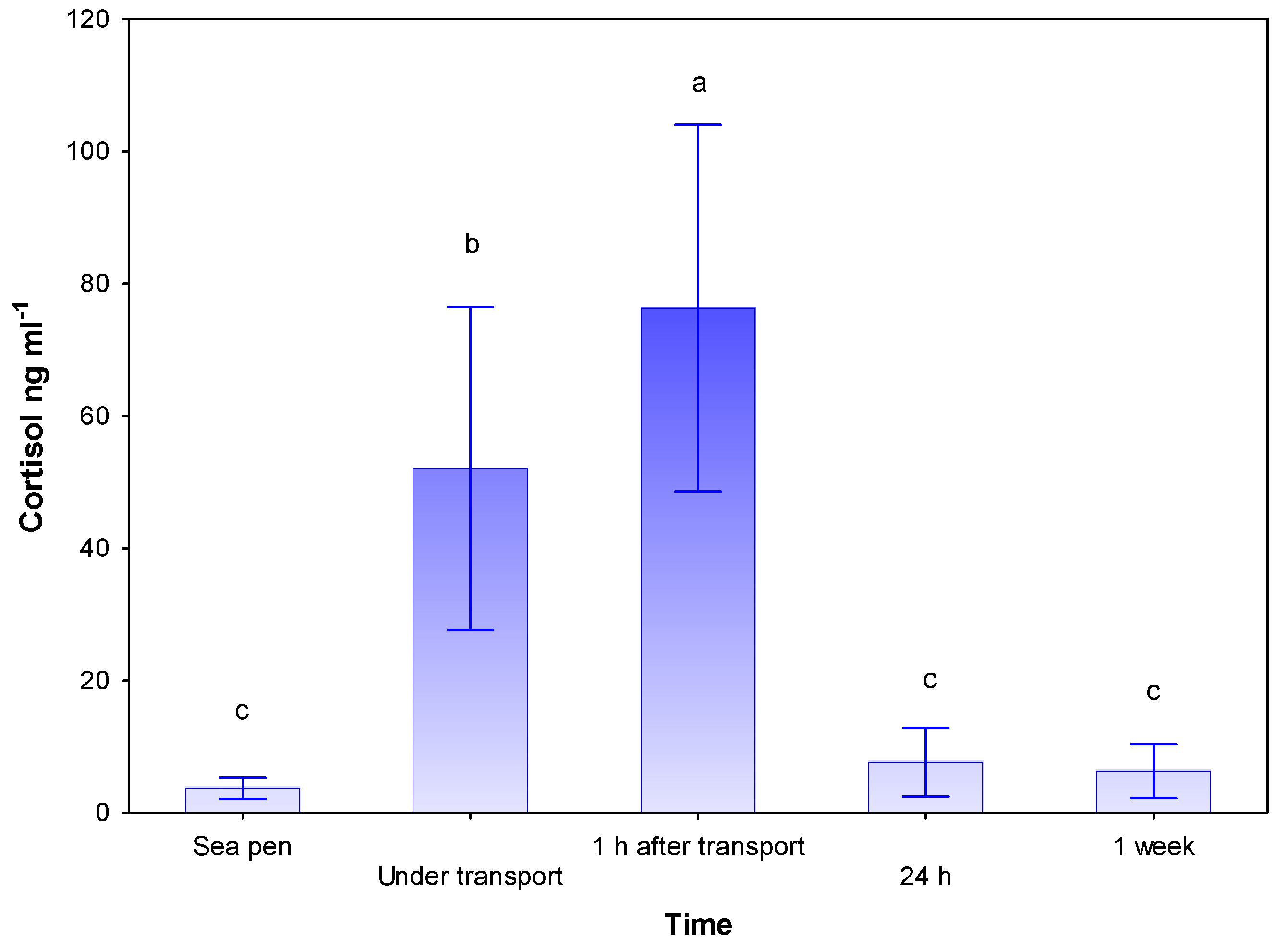
3.1. Blood Plasma Cortisol
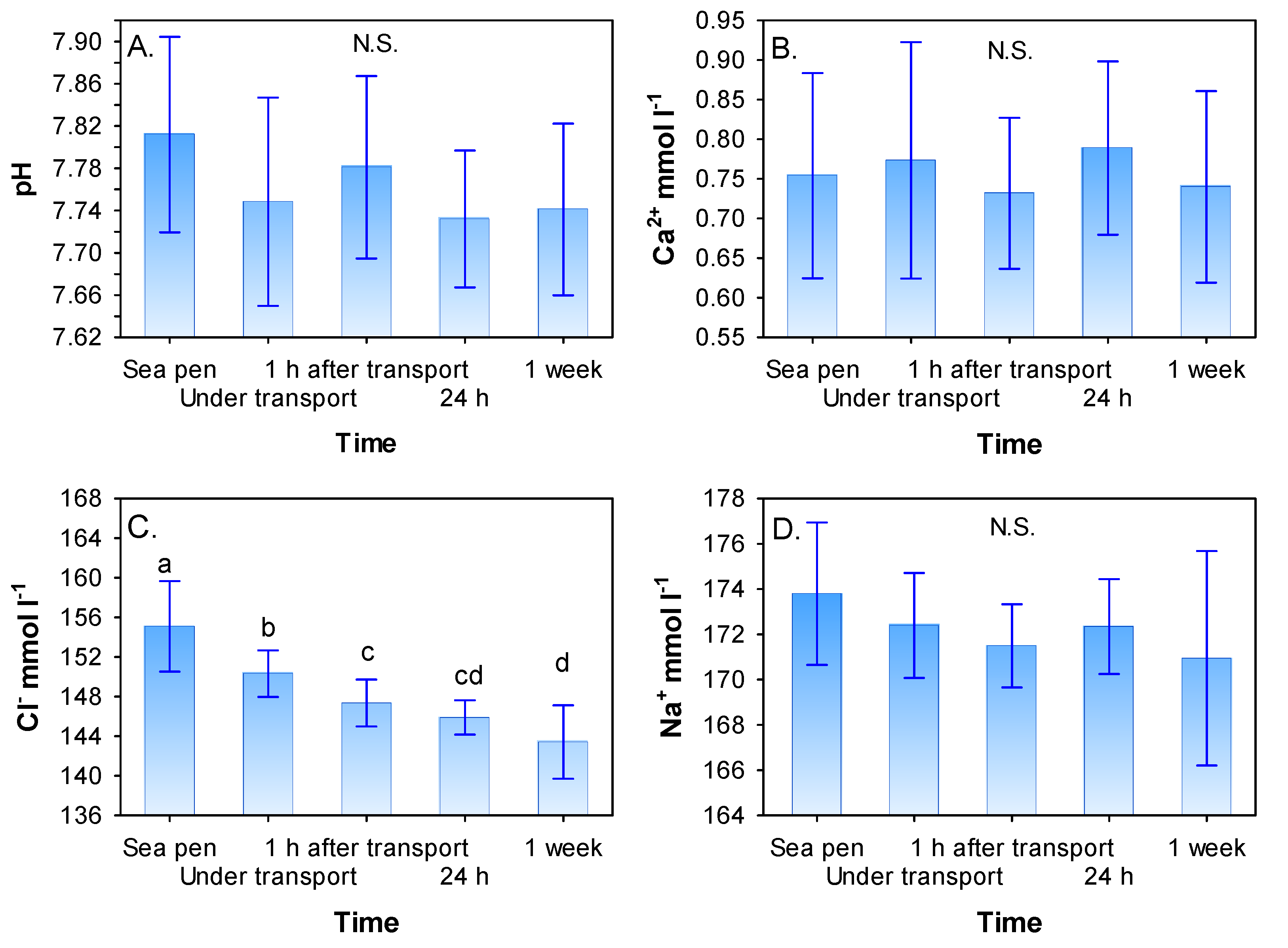
3.2. Blood Plasma pH, Ca2+, Cl−, Na+
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grutter, A.S. Cleaner fish really do clean. Nature 1999, 398, 672–673. [Google Scholar] [CrossRef]
- Vaughan, D.B.; Grutter, A.S.; Costello, M.J.; Hutson, K.S. Cleaner fishes and shrimp diversity and re-evaluation of cleaning symbioses. Fish Fish. 2017, 18, 698–716. [Google Scholar] [CrossRef]
- Skiftesvik, A.B.; Bjelland, R.M.; Durif, C.M.F.; Johansen, I.S.; Browman, H.I. Delousing of Atlantic salmon (Salmo salar) by cultured vs. wild ballan wrasse (Labrus bergylta). Aquaculture 2013, 402–403, 113–118. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Foss, A.; Vikingstad, E.; Elvegård, T.A. The use of lumpfish (Cyclopterus lumpus L.) to control sea lice (Lepeophtheirus salmonis Krøyer) infestations in intensively farmed Atlantic salmon (Salmo salar L.). Aquaculture 2014, 424–425, 18–23. [Google Scholar] [CrossRef]
- Powell, A.; Treasurer, J.W.; Pooley, C.L.; Keay, A.J.; Lloyd, R.; Imsland, A.K.; Garcia de Leaniz, C. Cleaner fish for sea-lice control in salmon farming: Challenges and opportunities using lumpfish. Rev. Aquac. 2018, 10, 683–702. [Google Scholar] [CrossRef]
- Brooker, A.J.; Papadopoulou, A.; Gutierrez, C.; Rey, S.; Davie, A.; Migaud, H. Sustainable production and use of cleaner fish for the biological control of sea lice: Recent advances and current challenges. Vet. Rec. 2018, 183, 383. [Google Scholar] [CrossRef]
- Treasurer, J.W. A review of potential pathogens of sea lice and the application of cleaner fish in biological control. Pest. Manag. Sci. 2002, 58, 546–558. [Google Scholar] [CrossRef] [PubMed]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Notes on the behaviour of lumfish with and without Atlantic salmon present. J. Ethol. 2014, 32, 117–122. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Nytrø, A.V.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Assessment of growth and sea lice infection levels in Atlantic salmon stocked in small-scale cages with lumpfish. Aquaculture 2014, 433, 137–142. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Nytrø, A.V.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Feeding preferences of lumpfish (Cyclopterus lumpus L.) maintained in open net-pens with Atlantic salmon (Salmo salar L.). Aquaculture 2015, 436, 47–51. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Eliassen, G.; Hangstad, T.A.; Nytrø, A.V.; Foss, A.; Vikingstad, E.; Elvegård, T.A. Assessment of suitable substrates for lumpfish in sea pens. Aquac. Int. 2015, 23, 639–645. [Google Scholar] [CrossRef]
- Imsland, A.K.; Hanssen, A.; Reynolds, P.; Nytrø, A.V.; Jonassen, T.M.; Hangstad, T.A.; Elvegård, T.A.; Urskog, T.C.; Mikalsen, B. It works! Lumpfish can significantly lower sea lice infections in large scale salmon farming. Biol. Open 2018, 7, bio036301. [Google Scholar] [CrossRef] [PubMed]
- Eliasen, K.; Danielsen, E.; Johannesen, Á.; Joensen, L.L.; Patursson, E.J. The cleaning efficacy of lumpfish (Cyclopterus lumpus L.) in Faroese salmon (Salmo salar L.) farming pens in relation to lumpfish size and season. Aquaculture 2018, 488, 61–65. [Google Scholar] [CrossRef]
- Boissonnot, L.; Kharlova, I.; Iversen, N.S.; Staven, F.R.; Austad, M. Characteristics of lumpfish (Cyclopterus lumpus) with high cleaning efficacy in commercial Atlantic salmon (Salmo salar) production. Aquaculture 2022, 560, 738544. [Google Scholar] [CrossRef]
- Imsland, A.K.D.; Reynolds, P. In lumpfish we trust? The efficacy of lumpfish to control Lepeophtheirus salmonis infestations on farmed Atlantic salmon: A review. Fishes 2022, 7, 220. [Google Scholar] [CrossRef]
- Imsland, A.K.; Reynolds, P.; Nytrø, A.V.; Eliassen, G.; Hangstad, T.A.; Jónsdóttir, Ó.D.B.; Emaus, P.A.; Elvegård, T.A.; Lemmens, S.C.A.; Rydland, R.; et al. Effects of lumpfish size on foraging behaviour and co-existence with sea lice infected Atlantic salmon in sea cages. Aquaculture 2016, 465, 19–27. [Google Scholar] [CrossRef]
- Roth, B. Avlusing av Laksefisk med Optilice: Effekt på Avlusing og Fiskevelferd. Nofima AS, Rapport 59/2016. 2016. In Norwegian with Abstract in English. Available online: https://nofima.no/publikasjon/1408716/ (accessed on 2 August 2022).
- Oliveira, V.H.S.; Dean, K.R.; Qviller, L.; Kirkeby, C.; Jensen, B.B. Factors associated with baseline mortality in Norwegian Atlantic salmon farming. Sci. Rep. 2021, 11, 14702. [Google Scholar] [CrossRef]
- Sommerset, I.; Walde, C.S.; Bang Jensen, B.; Bornø, B.; Haukaas, A.; Brun, E. The Health Situation in Norwegian Aquaculture 2019. Available online: https://www.vetinst.no/rapporter-og-publikasjoner/rapporter/2020/fiskehelserapporten-2019 (accessed on 2 August 2022).
- Jonassen, T.M.; Remen, M. Utvikling av Transport- og Mottaksprosedyrer for Rognkjeks Basert på Kartlegging av Miljø og Stress. Akvaplan-Niva Rapport nr. 7707-1. 2017. Available online: https://www.fhf.no/prosjekter/prosjektbasen/901158/ (accessed on 8 August 2022).
- Zar, J.H. Biostatistical Analysis, 2nd ed.; Prentice-Hall, Inc.: Englewood Cliffs, NJ, USA, 1984; 718p. [Google Scholar]
- Brown, M.B.; Forsythe, A.B. Robust tests for the equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Iversen, M.H. Stress and Its Impact on Animal Welfare during Commercial Production of Atlantic Salmon (Salmo salar L.). Ph.D. Thesis, Nord University, Bodø, Norway, April 2013. [Google Scholar]
- Iversen, M.H.; Jakobsen, R.; Eliassen, R.; Ottesen, O. Sedasjon av berggylt og rognkjeks for å redusere stress og dødelighet. NFexpert 2015, 39, 42–46. [Google Scholar]
- Iversen, M.H.; Eliassen, R.A. The effect of allostatic load on hypothalamic-pituitary-interrenal (HPI) axis before and after secondary vaccination in Atlantic salmon postsmolts (Salmo salar L.). Fish Physiol. Biochem. 2014, 40, 527–538. [Google Scholar] [CrossRef]
- Hanssen, H.J. The Effect of Long-Term Stress on Basal Levels of Plasma Cortisol and Hypothalamic–Pituitary–Interrenal (hpi) Axis in Lumpsucker (Cyclopterus lumpus). Master’s Thesis, Nord University, Bodø, Norway, June 2016. [Google Scholar]
- Hvas, M.; Folkedal, O.; Imsland, A.K.; Oppedal, F. Metabolic rates, swimming capabilities, thermal niche and stress response of the lumpfish, Cyclopterus lumpus. Biol. Open 2018, 7, bio036079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finstad, B.; Iversen, M.; Sandodden, R. Stress reducing methods for releases of Atlantic salmon (Salmo salar) smolts in Norway. Aquaculture 2003, 222, 203–214. [Google Scholar] [CrossRef]
- Iversen, M.; Eliassen, R.A. The effect of AQUI-S® sedation on primary, secondary, and tertiary stress responses during salmon smolt, Salmo salar L., transport and transfer to sea. J. World Aquac. Soc. 2009, 40, 216–225. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. The stress response in fish. Physiol. Rev. 1997, 77, 591–625. [Google Scholar] [CrossRef]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Frantzen, M.; Hansen, B.H.; Geraudie, P.; Palerud, J.; Falk-Petersen, I.B.; Olsen, G.H.; Camus, L. Acute and long-term biological effects of mechanically and chemically dispersed oil on lumpsucker (Cyclopterus lumpus). Mar. Environ. Res. 2015, 105, 8–19. [Google Scholar] [CrossRef]
- Davenport, J.; Kjørsvik, E. Buoyancy in the lumpsucker Cyclopterus lumpus. J. Mar. Biol. Assoc. U. K. 1986, 66, 159–174. [Google Scholar] [CrossRef]
- Berg, M. Comparing Density of Lumpfish (Cyclopterus lumpus) Used in Aquaculture to Different Welfare Indicators. Master’s Thesis, University of Bergen, Bergen, Norway, June 2021. [Google Scholar]
- Remen, M.; Nes, A.M.; Hangstad, T.A.; Geraudie, P.; Reynolds, P.; Urskog, T.C.; Hanssen, A.; Stefansson, S.O.; Imsland, A.K.D. Temperature and size-dependency of lumpfish (Cyclopterus lumpus) oxygen requirement and tolerance. Aquaculture 2022, 548, 737576. [Google Scholar] [CrossRef]
- Sällebrant, J.B. Chronic Allostatic Overload on the Hypothalamic-Pituitary-Interrenal Axis of Lumpfish (Cyclopterus lumpus). Effect of Size. Master’s Thesis, Nord University, Bodø, Norway, June 2018. [Google Scholar]
- Arends, R.J.; Mancera, J.M.; Munoz, J.L.; Wendelaar Bonga, S.E.; Flik, G. The stress response of the gilthead sea bream (Sparus aurata L.) to air exposure and confinement. J. Endocrionol. 1999, 163, 149–157. [Google Scholar] [CrossRef]
- Maxime, V.; Nonnotte, G.; Peyraud, C.; Williot, P.; Truchot, J.P. Circulatory and respiratory effects of an hypoxic stress in the Siberian sturgeon. Respir. Physiol. 1995, 100, 203–212. [Google Scholar] [CrossRef]
- Barton, B.A.; Bollig, H.; Hauskins, B.L.; Jansen, C.R. Juvenile pallid (Scaphirhynchus albus) and hybrid pallid x shovelnose (S. albus x platorynchus) sturgeons exhibit low physiological responses to acute handling and severe confinement. Comp. Biochem. Physiol. 2000, 126A, 125–134. [Google Scholar] [CrossRef]
- Belanger, J.M.; Son, J.H.; Laugero, K.D.; Moberg, G.P.; Doroshov, S.I.; Lankford, S.E.; Cech, J.J., Jr. Effects of short-term management stress and ACTH injections on plasma cortisol levels in cultured white sturgeon, Acipenser transmontanus. Aquaculture 2001, 203, 165–176. [Google Scholar] [CrossRef]
- Baker, D.W.; Wood, A.M.; Litvak, M.K.; Kieffer, J.D. Haematology of juvenile Acipenser oxyrinchus and Acipenser brevirostrum at rest and following forced activity. J. Fish Biol. 2005, 66, 208–221. [Google Scholar] [CrossRef]
- Øverli, Ø.; Sørensen, C.; Kiessling, A.; Pottinger, T.G.; Gjøen, H.M. Selection for improved stress tolerance in rainbow trout (Oncorhynchus mykiss) leads to reduced feed waste. Aquaculture 2006, 261, 776–781. [Google Scholar] [CrossRef]
- Barton, B.A. Stress in fishes: A diversity of responses with particular reference to changes in circulating corticosteroids. Int. Comp. Biol. 2002, 42, 517–525. [Google Scholar] [CrossRef]
- Barton, B.A.; Iwama, G.K. Physiological changes in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Ann. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar] [CrossRef]
- Wendelaar Bonga, S.E. Hormonal Responses to Stress. In Encyclopaedia of Fish Physiology; Farrell, A., Ed.; Academic Press: San Diego, CA, USA, 2011; pp. 1515–1523. [Google Scholar]
- Remen, M.; Oppedal, F.; Torgersen, T.; Imsland, A.K.; Olsen, R.E. Effects of cyclic environmental hypoxia on physiology and feed intake of post-smolt Atlantic salmon: Initial responses and acclimation. Aquaculture 2012, 326, 148–155. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Foss, A.; Imsland, A.K.D. Physiological Effects of Recapture and Transport from Net-Cages in Lumpfish. Fishes 2022, 7, 242. https://doi.org/10.3390/fishes7050242
Foss A, Imsland AKD. Physiological Effects of Recapture and Transport from Net-Cages in Lumpfish. Fishes. 2022; 7(5):242. https://doi.org/10.3390/fishes7050242
Chicago/Turabian StyleFoss, Atle, and Albert K. D. Imsland. 2022. "Physiological Effects of Recapture and Transport from Net-Cages in Lumpfish" Fishes 7, no. 5: 242. https://doi.org/10.3390/fishes7050242
APA StyleFoss, A., & Imsland, A. K. D. (2022). Physiological Effects of Recapture and Transport from Net-Cages in Lumpfish. Fishes, 7(5), 242. https://doi.org/10.3390/fishes7050242