
Temperature and Light Spectrum Differently Affect Growth, Morphology, and Leaf Mineral Content of Two Indoor-Grown Leafy Vegetables




Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture
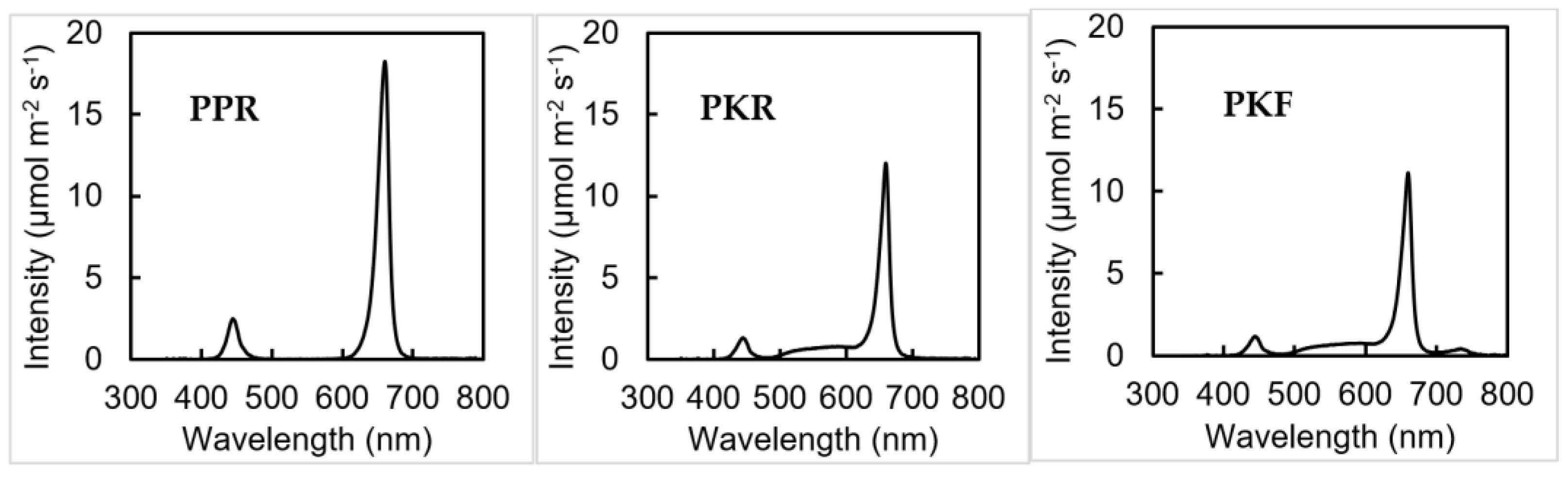
2.2. Experiment Design and Treatments
2.3. Data Collection
2.4. Statistical Analysis
3. Results
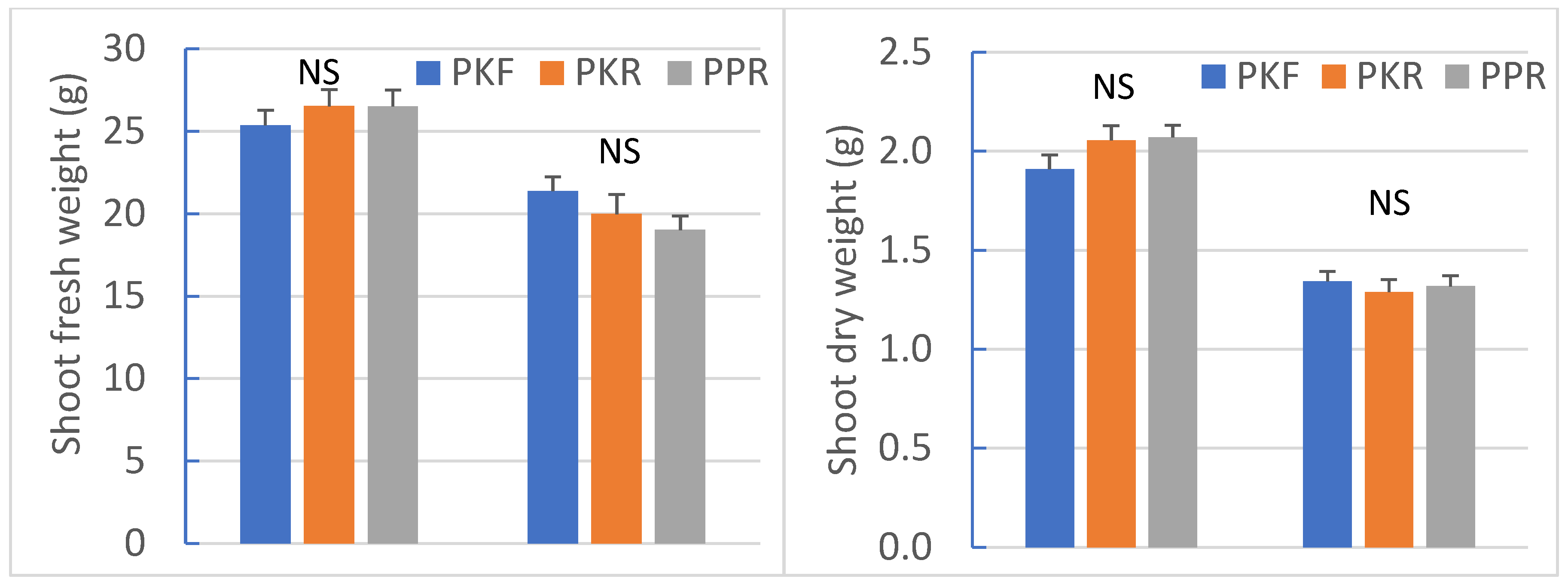
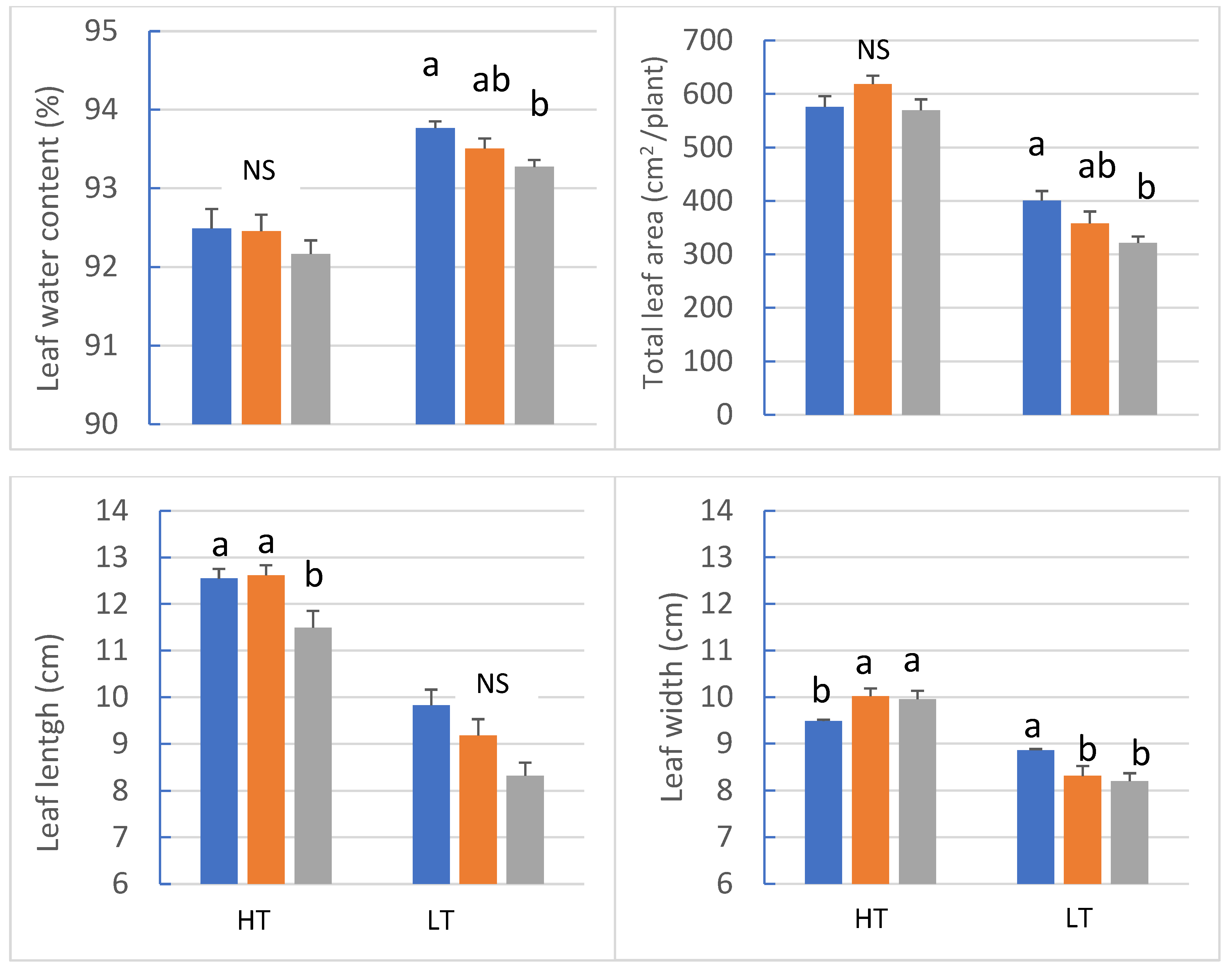
3.1. Growth and Morphology
3.2. Mineral Nutrition
4. Discussion
4.1. Light Spectrum Does Not Affect Plant Biomass Regardless of Crop Species and Temperature
4.2. Light Spectrum Effects on Plant Morphology and Mineral Content Are Species- and Temperature-Dependent
4.3. Temperature Influences Biomass, Morphology, and Mineral Nutrition
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kozai, T.; Niu, G. Role of the Plant Factory with Artificial Lighting (PFAL) in Urban Areas. In Plant Factory: An Indoor Vertical Farming System for Efficient Quality Food Production, 2nd ed.; Kozai, T., Niu, G., Takagaki, M., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 7–34. ISBN 9780128166918. [Google Scholar]
- Wong, C.E.; Teo, Z.W.N.; Shen, L.; Yu, H. Seeing the Lights for Leafy Greens in Indoor Vertical Farming. Trends Food Sci. Technol. 2020, 106, 48–63. [Google Scholar] [CrossRef]
- Zhuang, Y.; Lu, N.; Shimamura, S.; Maruyama, A.; Kikuchi, M.; Takagaki, M. Economies of Scale in Constructing Plant Factories with Artificial Lighting and the Economic Viability of Crop Production. Front. Plant Sci. 2022, 13, 992194. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Bugbee, B. Far-Red Photons Have Equivalent Efficiency to Traditional Photosynthetic Photons: Implications for Redefining Photosynthetically Active Radiation. Plant Cell Environ. 2020, 43, 1259–1272. [Google Scholar] [CrossRef] [PubMed]
- Bugbee, B.; Monje, O. The Limits of Crop Productivity. Bioscience 1992, 42, 494–502. [Google Scholar] [CrossRef]
- Emerson, R.; Chalmers, R.; Cederstrand, C. Some factors influencing the long-wave limit of photosynthesis. Proc. Natl. Acad. Sci. USA 1957, 43, 133–143. [Google Scholar] [CrossRef]
- Zhen, S.; van Iersel, M.W. Far-Red Light Is Needed for Efficient Photochemistry and Photosynthesis. J. Plant Physiol. 2017, 209, 115–122. [Google Scholar] [CrossRef]
- Zou, J.; Zhang, Y.; Zhang, Y.; Bian, Z.; Fanourakis, D.; Yang, Q.; Li, T. Morphological and Physiological Properties of Indoor Cultivated Lettuce in Response to Additional Far-Red Light. Sci. Hortic. 2019, 257, 108725. [Google Scholar] [CrossRef]
- Liu, J.; van Iersel, M.W. Far-Red Light Effects on Lettuce Growth and Morphology in Indoor Production Are Cultivar Specific. Plants 2022, 11, 2714. [Google Scholar] [CrossRef]
- Meng, Q.; Runkle, E.S. Far-Red Radiation Interacts with Relative and Absolute Blue and Red Photon Flux Densities to Regulate Growth, Morphology, and Pigmentation of Lettuce and Basil Seedlings. Sci. Hortic. 2019, 255, 269–280. [Google Scholar] [CrossRef]
- Feezell, D.; Nakamura, S. Invention, Development, and Status of the Blue Light-Emitting Diode, the Enabler of Solid-State Lighting. Comptes Rendus Phys. 2018, 19, 113–133. [Google Scholar] [CrossRef]
- Tsao, J.Y.; Han, J.; Haitz, R.H.; Pattison, P.M. The Blue LED Nobel Prize: Historical Context, Current Scientific Understanding, Human Benefit. Ann. Phys. 2015, 527, A53–A61. [Google Scholar] [CrossRef]
- Kusuma, P.; Pattison, P.M.; Bugbee, B. Photon Efficacy in Horticulture: Turning LED Packages into LED Luminaires; Kozai, N.M., Ed.; Academic Press: Cambridge, MA, USA, 2022; Chapter 7; ISBN 978-0-323-85152-7. [Google Scholar]
- Jeong, S.J.; Zhen, S.; Niu, G. The Interactive Effects between Far-Red and Temperature on Plant Growth and Morphology: Dependency of the Predictive Power of Phytochrome Photoequilbrium on Temperature; HortScience, 2022; pp. S152–S153. [Google Scholar]
- Soltanpour, J.L.H.; Soltanpour, P.N. A Nitric Acid Plant Tissue Digest Method for Use with Inductively Coupled Plasma Spectrometry. Commun. Soil Sci. Plant Anal. 1980, 11, 969–980. [Google Scholar]
- Robert, A.; Isaac, W.C.J. Collaborative Study of Wet and Dry Ashing Techniques for the Elemental Analysis of Plant Tissue by Atomic Absorption Spectrophotometry. J. Assoc. Off. Anal. Chem. 1975, 58, 436–440. [Google Scholar]
- Zhen, S.; Haidekker, M.; van Iersel, M.W. Far-Red Light Enhances Photochemical Efficiency in a Wavelength-Dependent Manner. Physiol. Plant 2019, 167, 21–33. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Far-Red Radiation Promotes Growth of Seedlings by Increasing Leaf Expansion and Whole-Plant Net Assimilation. Environ. Exp. Bot. 2017, 136, 41–49. [Google Scholar] [CrossRef]
- Kong, Y.; Nemali, K. Blue and Far-Red Light Affect Area and Number of Individual Leaves to Influence Vegetative Growth and Pigment Synthesis in Lettuce. Front. Plant Sci. 2021, 12, 667407. [Google Scholar] [CrossRef]
- Hooks, T.; Sun, L.; Kong, Y.; Masabni, J.; Niu, G. Adding UVA and Far-Red Light to White LED Affects Growth, Morphology, and Phytochemicals of Indoor-Grown Microgreens. Sustainability 2022, 14, 8552. [Google Scholar] [CrossRef]
- Li, Q.; Kubota, C. Effects of Supplemental Light Quality on Growth and Phytochemicals of Baby Leaf Lettuce. Environ. Exp. Bot. 2009, 67, 59–64. [Google Scholar] [CrossRef]
- Klose, C.; Nagy, F.; Schäfer, E. Thermal Reversion of Plant Phytochromes. Mol. Plant 2020, 13, 386–397. [Google Scholar] [CrossRef]
- Yan, W.; Hunt, L.A. An Equation for Modelling the Temperature Response of Plants Using Only the Cardinal Temperatures. Ann. Bot. 1999, 84, 607–614. [Google Scholar] [CrossRef]
- Inthichack, P.; Nishimura, Y.; Fukumoto, Y. Diurnal Temperature Alternations on Plant Growth and Mineral Absorption in Eggplant, Sweet Pepper, and Tomato. Hortic. Environ. Biotechnol. 2013, 54, 37–43. [Google Scholar] [CrossRef]
- Wang, H.; Wu, L.; Zhu, Y.; Tao, Q. Growth, Nitrate Accumulation, and Macronutrient Concentration of Pakchoi as Affected by External Nitrate-N: Amino Acid-N Ratio. J. Plant Nutr. 2008, 31, 1789–1799. [Google Scholar] [CrossRef]
- Khader, V.; Rama, S. Selected Mineral Content of Common Leafy Vegetables Consumed in India at Different Stages of Maturity. Plant Foods Hum. Nutr. 1998, 53, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Waterland, N.L.; Moon, Y.; Tou, J.C.; Kim, M.J.; Pena-Yewtukhiw, E.M.; Park, S. Mineral Content Differs among Microgreen, Baby Leaf, and Adult Stages in Three Cultivars of Kale. HortScience 2017, 52, 566–571. [Google Scholar] [CrossRef]
- Kroggel, M.; Lovichit, W.; Kubota, C.; Thomson, C. Greenhouse Baby Leaf Production of Lettuce and Komatsuna in Semi-Arid Climate: Seasonal Effects on Yield and Quality. Acta Hortic. 2012, 952, 827–834. [Google Scholar] [CrossRef]
- Lee, S.G.; Choi, C.S.; Lee, J.G.; Jang, Y.A.; Lee, H.J.; Lee, H.J.; Chae, W.B.; Um, Y.C. Influence of Air Temperature on Yield and Phytochemical Content of Red Chicory and Garland Chrysanthemum Grown in Plant Factory. Hortic. Environ. Biotechnol. 2013, 54, 399–404. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | SPAD | Shoot FW | Leaf FW | TLA | Leaf DW | Shoot DW | Leaf WC | Shoot WC | LL | LW |
|---|---|---|---|---|---|---|---|---|---|---|
| Lettuce | ||||||||||
| Temp. (T) | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| Light (L) | * | NS | NS | * | NS | NS | * | * | *** | NS |
| T ×L | NS | NS | NS | * | NS | NS | NS | NS | NS | ** |
| Pak choy | ||||||||||
| Temp. (T) | NS | *** | *** | *** | ** | ** | NS | NS | ** | ** |
| Light (L) | NS | NS | NS | NS | NS | NS | *** | *** | NS | NS |
| T × L | NS | NS | NS | NS | NS | NS | NS | NS | NS | NS |
| Factor | N | P | K | Ca | Mg | S | Na | Zn | Fe | Cu | Mn | B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (%) | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (mg/kg) | (mg/kg) | (mg/kg) | (mg/kg) | (mg/kg) | |
| High temperature (HT) | ||||||||||||
| PKF | 2.85 | 3.72 | 21.68 | 14.64 | 3.18 | 5.27 | 3.87 | 30.22 | 30.21 | 2.37 | 15.29 | 18.59 |
| PKR | 2.71 | 3.65 | 20.48 | 15.10 | 3.06 | 5.01 | 3.67 | 27.52 | 29.58 | 2.02 | 17.85 | 18.28 |
| PPR | 2.58 | 3.52 | 19.84 | 14.64 | 3.03 | 4.71 | 2.84 | 25.91 | 28.29 | 1.92 | 18.35 | 15.34 |
| Low temperature (LT) | ||||||||||||
| PKF | 3.59 a z | 4.71 | 24.93 | 17.48 | 4.37 a | 4.53 a | 4.38 a | 37.24 | 34.02 | 2.61 a | 20.82 a | 25.72 b |
| PKR | 3.24 a | 4.27 | 21.16 | 15.51 | 4.13 a | 4.23 b | 2.85 b | 35.83 | 33.02 | 2.39 b | 18.32 b | 23.30 b |
| PPR | 3.05 b | 4.59 | 22.19 | 15.04 | 3.75 b | 4.31 ab | 2.97 ab | 37.21 | 33.87 | 2.94 a | 18.61 ab | 30.36 a |
| ANOVA Summary | ||||||||||||
| T y | *** | *** | ** | NS | *** | *** | *** | *** | *** | *** | *** | *** |
| L | * | NS | NS | NS | ** | NS | NS | NS | NS | * | NS | NS |
| T x L | NS | NS | NS | NS | ** | NS | NS | NS | NS | NS | ** | NS |
| Factor | N | P | K | Ca | Mg | S | Na | Zn | Fe | Cu | Mn | B |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (%) | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (g/kg) | (mg/kg) | (mg/kg) | (mg/kg) | (mg/kg) | (mg/kg) | |
| High temperature (HT) | ||||||||||||
| PKF | 2.93 | 3.90 | 21.23 | 17.20 | 2.81 | 8.87 | 2.66 a | 33.61 | 32.72 | 2.59 a | 17.99 b | 27.56 |
| PKR | 2.81 | 3.94 | 20.01 | 18.17 | 2.70 | 8.43 | 2.32 a | 31.41 | 32.14 | 2.07 ab | 23.61 a | 27.52 |
| PPR | 2.57 | 3.65 | 18.26 | 16.79 | 2.55 | 7.80 | 1.19 b | 29.59 | 28.34 | 1.87 b | 22.43 a | 22.39 |
| Low temperature (LT) | ||||||||||||
| PKF | 3.27 a z | 4.49 | 21.55 a | 23.01 a | 3.03 | 6.90 | 4.89 a | 41.48 | 30.09 ab | 1.82 | 23.90 a | 22.97 b |
| PKR | 2.75 b | 3.93 | 17.65 b | 19.11 b | 2.77 | 6.41 | 2.08 b | 37.46 | 26.63 b | 1.98 | 17.61 b | 19.80 b |
| PPR | 2.88 ab | 4.37 | 19.25 ab | 19.27 ab | 3.10 | 6.82 | 3.20 ab | 42.58 | 32.61 a | 2.39 | 21.84 a | 33.79 a |
| ANOVA summary | ||||||||||||
| T y | NS | ** | NS | *** | *** | *** | *** | *** | NS | NS | NS | NS |
| L | NS | NS | * | NS | NS | NS | *** | NS | NS | NS | NS | NS |
| T x L | NS | NS | NS | * | * | NS | ** | NS | ** | ** | *** | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, Y.; Masabni, J.; Niu, G. Temperature and Light Spectrum Differently Affect Growth, Morphology, and Leaf Mineral Content of Two Indoor-Grown Leafy Vegetables. Horticulturae 2023, 9, 331. https://doi.org/10.3390/horticulturae9030331
Kong Y, Masabni J, Niu G. Temperature and Light Spectrum Differently Affect Growth, Morphology, and Leaf Mineral Content of Two Indoor-Grown Leafy Vegetables. Horticulturae. 2023; 9(3):331. https://doi.org/10.3390/horticulturae9030331
Chicago/Turabian StyleKong, Yun, Joseph Masabni, and Genhua Niu. 2023. "Temperature and Light Spectrum Differently Affect Growth, Morphology, and Leaf Mineral Content of Two Indoor-Grown Leafy Vegetables" Horticulturae 9, no. 3: 331. https://doi.org/10.3390/horticulturae9030331
APA StyleKong, Y., Masabni, J., & Niu, G. (2023). Temperature and Light Spectrum Differently Affect Growth, Morphology, and Leaf Mineral Content of Two Indoor-Grown Leafy Vegetables. Horticulturae, 9(3), 331. https://doi.org/10.3390/horticulturae9030331