
Physiological and Productive Responses of Two Vitis vinifera L. Cultivars across Three Sites in Central-South Italy

 ,
,  ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Material and Methods
2.1. Site Description, Plant Material, and Experimental Design
2.2. Climate and Soil
2.3. Treatments
2.4. Physiological Behavior and Vine Water Status Measurements
2.5. Crop Yield and Berry Characteristics
2.6. Statistical Analyses
3. Results
3.1. Environmental Variables
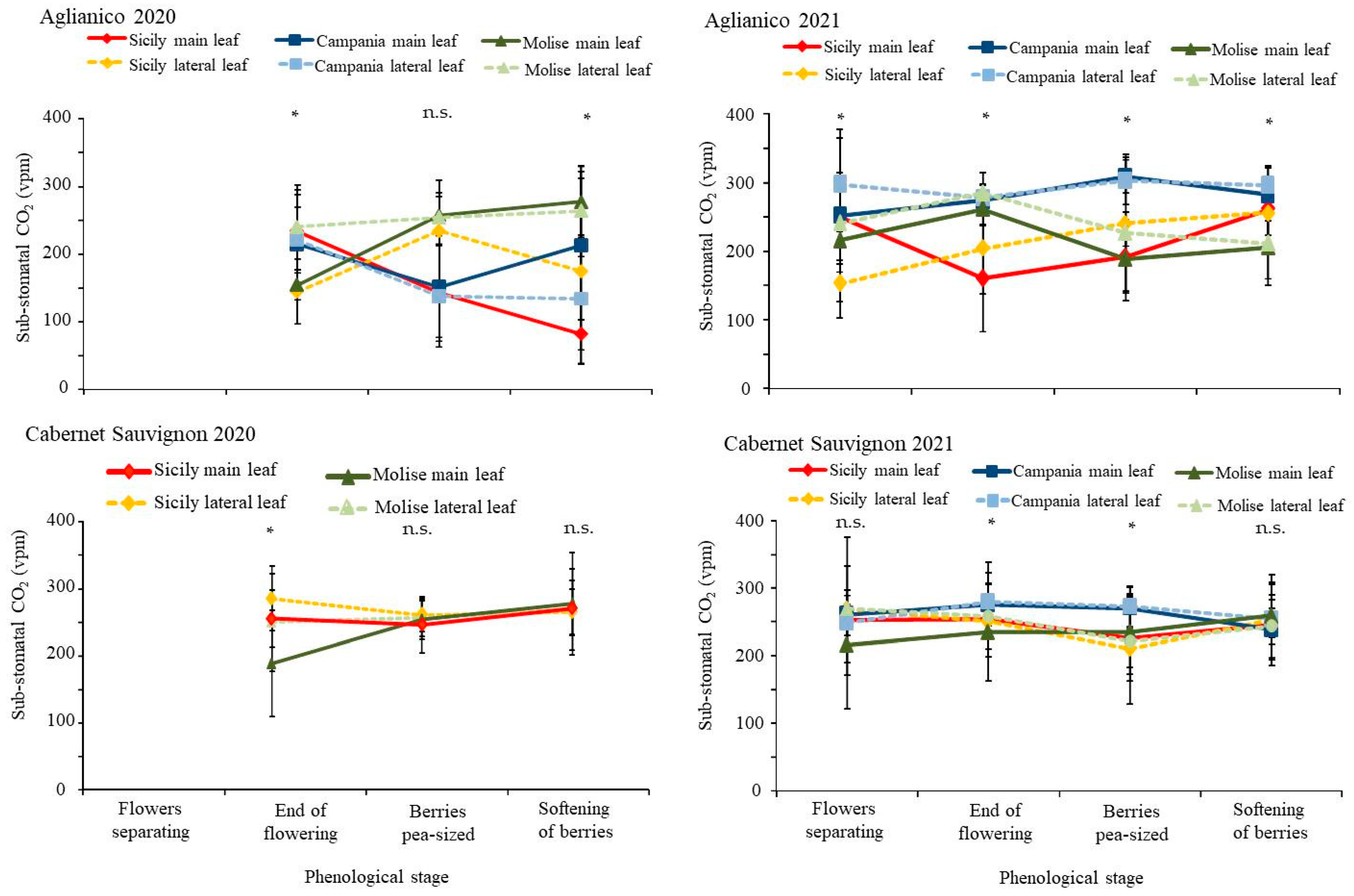
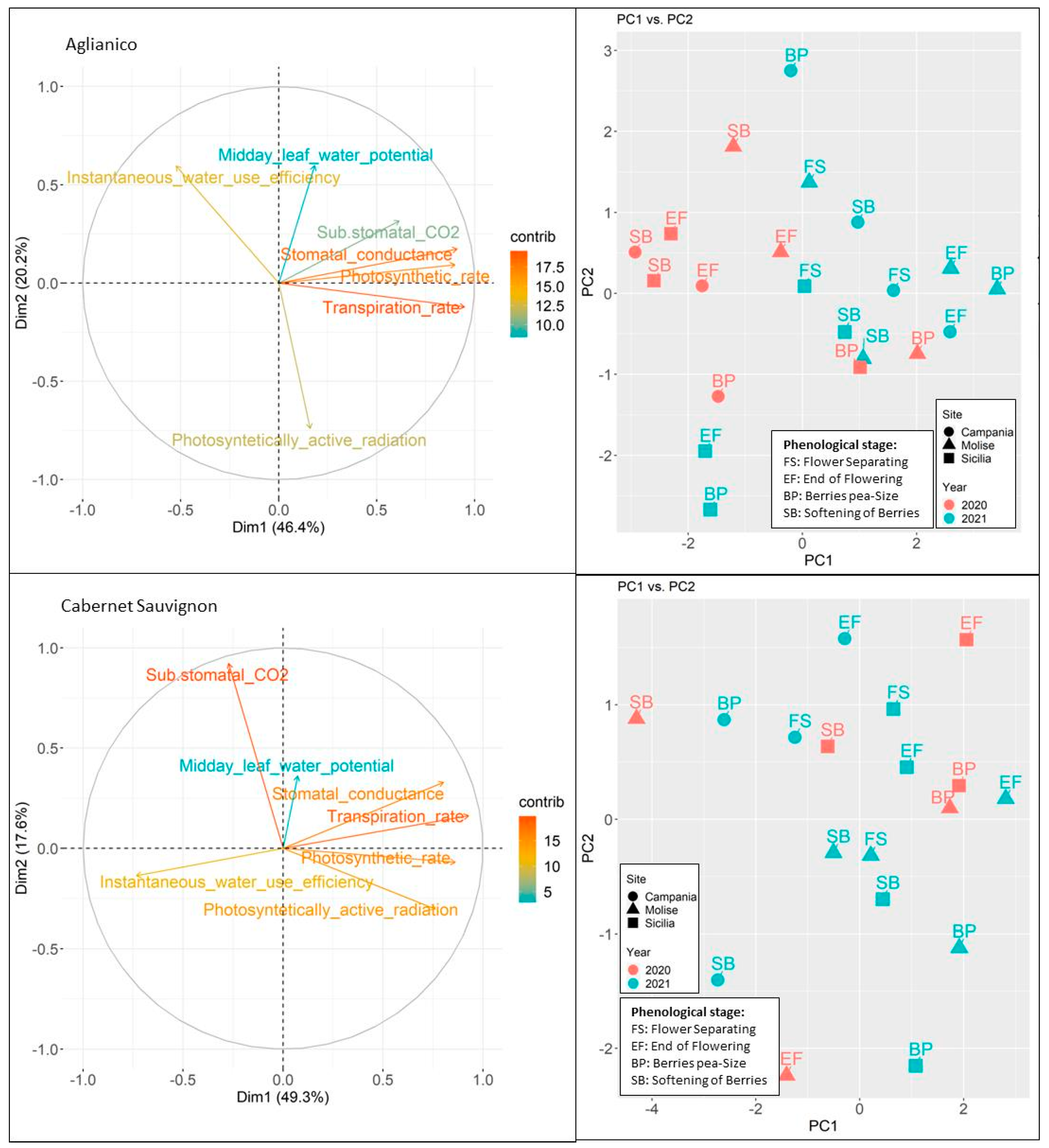
3.2. Vines Ecophysiological Response
3.3. Yield and Its Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAOSTAT). Available online: http://faostat3.fao.org/home/E (accessed on 25 July 2023).
- Alleweldt, G.; Possingham, J.V. Progress in grapevine breeding. Theor. Appl. Genet. 1988, 75, 669–673. [Google Scholar] [CrossRef]
- Galet, P. Dictionnaire Encylcopédique des Cépages; Hachette: Paris, France, 2000. [Google Scholar]
- Levadoux, L. Les populations sauvages et cultivées de Vitis vinifera L. Ann. Amélioration Plantes 1956, 1, 59–118. [Google Scholar]
- Olmo, H.P. The origin and domestication of the vinifera grape. In The Origins and Ancient History of Wine; McGovern, P.E., Fleming, S.J., Katz, S.H., Eds.; Gordon and Breach: New York, NY, USA, 1996; pp. 31–43. [Google Scholar]
- Scozzafava, G.; Casini, L.; Contini, C. Analysis of Italian Consumer preferences for beef. New Medit. 2014, 13, 66–72. [Google Scholar]
- Ferlito, F.; Nicolosi, E.; La Malfa, S.; Cicala, A.; Gentile, A. First characterisation of minor and neglected Vitis vinifera L. cultivars from mount etna. Hortic. Sci. 2018, 45, 37–46. [Google Scholar] [CrossRef]
- Iorizzo, M.; Sicilia, A.; Nicolosi, E.; Forino, M.; Picariello, L.; Lo Piero, A.R.; Vitale, A.; Monaco, E.; Ferlito, F.; Succi, M.; et al. Investigating the impact of pedoclimatic conditions on the oenological performance of two red cultivars grown throughout southern Italy. Front. Plant Sci. 2023, 14, 1250208. [Google Scholar] [CrossRef]
- Lai, M.; Del Giudice, T.; Pomarici, E. Unobserved Heterogeneity in the Wine Market: An Analysis on Sardinia Wine via Mixed Logit; Colloque International de la Vineyard Data Quantification Society: Bordeaux, France, 2006. [Google Scholar]
- Boncinelli, F.; Casinia, L.; Continia, C.; Gerinia, F.; Scozza Fava, G. The consumer loves typicality but prefers the international wine. Agric. Agric. Sci. Procedia 2016, 8, 236–242. [Google Scholar] [CrossRef]
- Nicolosi, E.; Continella, A.; Gentile, A.; Cicala, A.; Ferlito, F. Influence of early leaf removal on autochthonous and international grapevines in Sicily. Sci. Hortic. 2012, 146, 1–6. [Google Scholar] [CrossRef]
- Ferlito, F.; Nicolosi, E.; Gentile, A.; Lo Piero, A.R.; Squadrito, M.; Continella, A. Response of four grapevines to water stress and canopy management in an arid environment. Vitis 2014, 53, 73–80. [Google Scholar]
- Lo Cicero, L.; Puglisi, I.; Nicolosi, E.; Gentile, A.; Ferlito, F.; Continella, A.; Lo Piero, A.R. Anthocyanin levels and expression analysis of biosynthesis-related genes during ripening of sicilian and international grape berries subjected to leaf removal and water deficit. J. Agric. Sci. 2016, 18, 1333–1344. [Google Scholar]
- Schultz, H.R.; Stoll, M. Some critical issues in environmental physiology of grapevines: Future challenges and current limitations. Aust. J. Grape Wine Res. 2000, 16, 4–24. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change. Climate Change, 2007: The Physical Science Basis; Intergovernmental Panel on Climate Change: Paris, France; World Meterological Organization: Geneva, Switzerland, 2007. [Google Scholar]
- Schultz, H.R. Differences in hydraulic architecture account for near-isohydric and anisohydric behaviour of two field-grown Vitis vinifera L. cultivars during drought. Plant Cell Environ. 2003, 26, 1393–1405. [Google Scholar] [CrossRef]
- Etuk, S.E.; Nwokolo, S.C.; Okechukwu, E.A.; John-Jaja, S.A. Analysis of photosynthetically active radiation over six tropical ecological zones in Nigeria. J. Geogr. Environ. Earth Sci. Int. 2016, 7, 1–15. [Google Scholar]
- Poni, S.; Intrieri, C. Grapevine photosynthesis: Effects linked to light radiation and leaf age. Adv. Hortic. Sci. 2001, 15, 5–15. [Google Scholar]
- Ainsworth, E.A.; Rogers, A. The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions. Plant Cell Environ. 2007, 30, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Von Caemmerer, S.; Setchell, B.A.; Hudson, G.S. The relationship between CO2 transfer and leaf anatomy in transgenic tobacco with a reduced content of rubisco. Aust. J. Plant Physiol. 1994, 21, 475–495. [Google Scholar] [CrossRef]
- Chaves, M.M.; Zarrouk, O.; Francisco, R.; Costa, J.M.; Santos, T.; Regalado, A.P.; Rodrigues, M.L.; Lopes, C.M. Grapevine under deficit irrigation: Hints from physiological and molecular data. Ann. Bot. 2010, 105, 661–676. [Google Scholar] [CrossRef]
- Costa, J.M.; Ortuño, M.F.; Lopes, C.M.; Chaves, M.M. Grapevine varieties exhibiting differences in stomatal response to water deficit. Funct. Plant Biol. 2012, 39, 179–189. [Google Scholar] [CrossRef]
- Nicolosi, E.; Sicilia, A.; Ferlito, F.; Bonfante, A.; Monaco, E.; Lo Piero, A.R. Phenotypic Plasticity in Bud Fruitfulness Expressed in Two Distinct Wine Grape Cultivars Grown under Three Different Pedoclimatic Conditions. Agriculture 2022, 12, 1660. [Google Scholar] [CrossRef]
- Toumi, I.; M’Sehli, W.; Bourgou, S.; Jallouli, N.; Bensalem-Fnayou, A.; Ghorbel, A.; Mliki, A. Response of ungrafted and grafted grapevine cultivars and rootstocks (Vitis spp.) to water stress. J. Int. Sci. Vigne Vin 2007, 41, 85–93. [Google Scholar]
- Lovisolo, C.; Schubert, A. Effects of water stress on vessel size and xylem hydraulic conductivity in Vitis vinifera L. J. Exp. Bot. 1998, 49, 693–700. [Google Scholar]
- Bota, J.; Flexas, J.; Medrano, H. Genetic variability of photosynthesis and water use in Balearic grapevine cultivars. Ann. Appl. Biol. 2001, 138, 353–361. [Google Scholar] [CrossRef]
- Nicolosi, E.; Ferlito, F.; Allegra, M.; Cicala, A.; Trovato, F.; La Malfa, S. Influences of aspect and tillage on two winegrape cultivars on Mount Etna. N. Z. J. Crop Hortic. Sci. 2016, 44, 83–102. [Google Scholar] [CrossRef][Green Version]
- Cifre, J.; Bota, J.; Escalona, J.M.; Medrano, H.; Flexas, J. Physiological tools for irrigation scheduling in grapevine (Vitis vinifera L.). An open gate to improve water-use efficiency? Agric. Ecosyst. Environ. 2005, 106, 159–170. [Google Scholar] [CrossRef]
- Bates, L.M.; Hall, A.E. Stomatal closure with soil water depletion not associated with changes in bulk leaf water status. Oecologia 1981, 50, 62–65. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, T.R.; Holbrook, N.M.; Zwieniecki, M.A. Daily transpiration rates of woody species on drying soil. Tree Physiol. 2005, 25, 1469–1472. [Google Scholar] [CrossRef] [PubMed]
- Martin-StPaul, N.; Delzon, S.; Cochard, H. Plant resistance to drought depends on timely stomatal closure. Ecol. Lett. 2007, 20, 1437–1447. [Google Scholar] [CrossRef]
- Tardieu, F.; Simonneau, T. Variability among species of stomatal control under fluctuating soil water status and evaporative demand: Modelling isohydric and anisohydric behaviours. J. Exp. Bot. 1998, 49, 419–432. [Google Scholar] [CrossRef]
- Collins, M.J.; Fuentes, S.; Barlow, E.W. Partial rootzone drying and deficit irrigation increase stomatal sensitivity to vapour pressure deficit in anisohydric grapevines. Funct. Plant Biol. 2010, 37, 128–138. [Google Scholar] [CrossRef]
- Chaves, M.M.; Santos, T.; Souza, C.R.; Ortuño, M.F.; Rodrigues, M.L.; Lopes, C.; Maroco, J.; Pereira, J.S. Deficit irrigation in grapevine improves water-use efficiency while controlling vigour and production quality. Ann. Appl. Biol. 2007, 150, 237–252. [Google Scholar] [CrossRef]
- Santos, T.; Lopes, C.M.; Rodrigues, M.L.; Souza, C.R.; Maroco, J.; Pereira, J.S.; Silva, J.R.; Chaves, M.M. Partial rootzone drying irrigation affects cluster micro-climate improving fruit composition of ‘Moscatel’ field-grown grapevines. Sci. Hortic. 2007, 112, 321–330. [Google Scholar] [CrossRef]
- Intrigliolo, D.S.; Pèrez, D.; Risco, D.; Yeves, A.; Castel, J.R. Yield components and grape composition responses to seasonal water deficits in Tempranillo grapevines. Irrig. Sci. 2022, 30, 339–349. [Google Scholar] [CrossRef]
- Sadras, V.O.; Petrie, P.R.; Moran, M.A. Effects of elevated temperature in grapevine. II juice pH, titratable acidity and wine sensory attributes. Aust. J. Grape Wine Res. 2013, 19, 107–115. [Google Scholar] [CrossRef]
- Palai, G.; Caruso, G.; Gucci, R.; D’Onofrio, C. Berry flavonoids are differently modulated by timing and intensities of water deficit in Vitis vinifera L. cv. Sangiovese. Front. Plant Sci. 2022, 13, 1040899. [Google Scholar] [CrossRef]
- Price, S.F.; Breen, P.J.; Valladao, M.; Watson, B.T. Cluster sun exposure and quercetin in Pinot noir grapes and wine. Am. J. Enol. Vitic. 1995, 46, 187–194. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among wine grape microbiome, metabolome, and fermentation behavior suggest microbial contribution to regional wine characteristics. MBio 2016, 7, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Klingebiel, A.A. Land-Capability Classification (No. 210); Soil Conservation Service, US Department of Agriculture: Washington, DC, USA, 1961. [Google Scholar]
- Ferlito, F.; Torrisi, B.; Allegra, M.; Stagno, F.; Caruso, P.; Fascella, G. Evaluation of Conifer Wood Biochar as Growing Media Component for Citrus Nursery. Appl. Sci. 2020, 10, 1618. [Google Scholar] [CrossRef]
- Las Casas, G.; Ciaccia, C.; Iovino, V.; Ferlito, F.; Torrisi, B.; Lodolini, E.M.; Giuffrida, A.; Catania, R.; Nicolosi, E.; Bella, S. Effects of Different Inter-Row Soil Management and Intra-Row Living Mulch on Spontaneous Flora, Beneficial Insects, and Growth of Young Olive Trees in Southern Italy. Plants 2022, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Meier, U. BBCH-Monograph: Growth Stages of Mono-and Dicotyledonous Plants, 2nd ed.; Technical Report; Federal Biological Research Centre for Agriculture and Forestry: Bonn, Germany, 2001; p. 158. [Google Scholar]
- Schöelander, P.F.; Hammel, H.J.; Bradstreet, A.; Hemmingsen, E.A. Sap pressure in vascular plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef]
- Ferlito, F.; Allegra, M.; Torrisi, B.; Pappalardo, H.; Gentile, A.; Malfa, S.L.; Continella, A.; Stagno, F.; Nicolosi, E. Early defoliation effects on water status, fruit yield and must quality of ‘Nerello mascalese’ grapes. Sci. Agric. 2020, 77. [Google Scholar] [CrossRef]
- Nicolosi, E.; Iovino, V.; Distefano, G.; Di Guardo, M.; La Malfa, S.; Gentile, A.; Palliotti, A.; Las Casas, G.; Ferlito, F. Mid-Term Effects of Conservative Soil Management and Fruit-Zone Early Leaf Removal Treatments on the Performance of Nerello Mascalese (Vitis vinifera L.) Grapes on Mount Etna (Southern Italy). Agronomy 2021, 11, 1070. [Google Scholar] [CrossRef]
- Esmaeilpour, A.; Van Labeke, M.C.; Samson, R.; Boeckx, P.; Van Damme, P. Variation in biochemical characteristics, water status, stomata features, leaf carbon isotope composition and its relationship to water use efficiency in pistachio (Pistacia vera L.) cultivars under drought stress condition. Sci. Hortic. 2016, 211, 158–166. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.r-project.org/ (accessed on 30 October 2023).
- Revelle, W. Psych: Procedures for Personality and Psychological Research; R package version; Northwestern University: Evanston, IL, USA, 2021. [Google Scholar]
- Long, S.P.; Ainsworth, E.A.; Leakey, A.D.; Nosberger, J.; Ort, D.R. Food for thought: Lower-than-expected crop yield stimulation with rising CO2 concentrations. Science 2006, 312, 1918–1921. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U.; Degu, A.; Fait, A.; Rachmilevitch, S. Near isohydric grapevine cultivar displays higher photosynthetic efficiency and photorespiration rates under drought stress as compared with near anisohydric grapevine cultivar. Physiol. Plant. 2013, 147, 443–452. [Google Scholar] [CrossRef]
- Hochberg, U.; Rockwell, F.E.; Holbrook, N.M.; Cochard, H. Iso/anisohydry: A plant—Environment interaction rather than a simple hydraulic trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Tramontini, S.; van Leeuwen, C.; Domec, J.C.; Destrac-Irvine, A.; Basteau, C.; Vitali, M.; Mosbach-Schulz, O.; Lovisolo, C. Impact of soil texture and water availability on the hydraulic control of plant and grape-berry development. Plant Soil 2013, 368, 215–230. [Google Scholar] [CrossRef]
- Lovisolo, C.; Perrone, I.; Carra, A.; Ferrandino, A.; Flexas, J.; Medrano, H.; Schubert, A. Drought-induced changes in development and function of grapevine (Vitis spp.) organs and in their hydraulic and non hydraulic interactions at the whole plant level: A physiological and molecular update. Funct. Plant Biol. 2010, 37, 98–116. [Google Scholar] [CrossRef]
- de Souza, C.R.; Maroco, J.P.; dos Santos, T.P.; Rodrigues, M.L.; Lopes, C.M.; Pereira, J.S.; Chaves, M. Partial rootzone drying: Regulation of stomatal aperture and carbon assimilation in field-grown grapevines (Vitis vinifera cv. Moscatel). Funct. Plant Biol. 2003, 30, 653–662. [Google Scholar] [CrossRef]
- Soar, C.J.; Speirs, J.; Maffei, S.M.; Penrose, A.B.; McCarthy, M.G.; Loveys, B.R. Grape vine varieties Shiraz and Grenache differ in their stomatal response to VPD: Apparent links with ABA physiology and gene expression in leaf tissue. Aust. J. Grape Wine Res. 2006, 12, 2–12. [Google Scholar] [CrossRef]
- Levin, A.D.; Williams, L.E.; Matthews, M.A. A continuum of stomatal responses to water deficits among 17 wine grape cultivars (Vitis vinifera). Funct. Plant Biol. 2019, 47, 11–25. [Google Scholar] [CrossRef]
- López-Urrea, R.; Montoro, A.; Mañas, F.; López-Fuster, P.; Fereres, E. Evapotranspiration and crop coefficients from lysimeter measurements of mature ‘Tempranillo’ wine grapes. Agric. Water Manag. 2012, 112, 13–20. [Google Scholar] [CrossRef]
- Santoro, D.F.; Sicilia, A.; Testa, G.; Cosentino, S.L.; Lo Piero, A.R. Global leaf and root transcriptome in response to cadmium reveals tolerance mechanisms in Arundo donax L. BMC Genom. 2022, 23, 427. [Google Scholar] [CrossRef] [PubMed]
- Williams, L.E.; Ayars, J.E. Grapevine water use and the crop coefficient are linear functions of the shaded area measured beneath the canopy. Agric. For. Meteorol. 2005, 132, 201–211. [Google Scholar] [CrossRef]
- Tramontini, S.; Vitali, M.; Centioni, L.; Schubert, A.; Lovisolo, C. Rootstock control of scion response to water stress in grapevine. Environ. Exp. Bot. 2003, 93, 20–26. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Cultivar | Site | Leaf | Instantaneous Water Use Efficiency [WUE] (μmol mol−1) | |||
|---|---|---|---|---|---|---|---|
| Flowers Separating | End of Flowering | Berries Pea-Sized | Berries Softening | ||||
| 2020 | Aglianico | Sicilia | Main | 3.68 ± 0.97 n.s. | 2.3 ± 3.11 n.s. | 4.15 ± 2.21 ab | |
| Lateral | 3.88 ± 0.77 n.s. | 2.10 ± 0.90 n.s. | 4.73 ± 2.05 ab | ||||
| Campania | Main | 3.87 ± 2.43 n.s. | 2.37 ± 0.90 n.s. | 2.54 ± 2.28 b | |||
| Lateral | 3.50 ± 1.49 n.s. | 2.80 ± 1.07 n.s. | 6.30 ± 8.03 a | ||||
| Molise | Main | 3.82 ± 2.80 n.s. | 1.85 ± 0.55 n.s. | 3.40 ± 1.54 ab | |||
| Lateral | 3.81 ± 6.15 n.s. | 1.80 ± 0.72 n.s. | 3.74 ± 1.54 ab | ||||
| Cabernet Sauvignon | Sicilia | Main | 1.76 ± 0.27 bc | 2.63 ± 0.85 a | 2.70 ± 0.81 b | ||
| Lateral | 1.29 ± 0.38 c | 2.34 ± 0.57 ab | 2.91 ± 0.65 b | ||||
| Campania | Main | ||||||
| Lateral | |||||||
| Molise | Main | 2.27 ± 1.36 b | 2.11 ± 0.71 ab | 5.25 ± 1.93 a | |||
| Lateral | 3.26 ± 1.53 a | 2.04 ± 0.49 b | 5.06 ± 2.25 a | ||||
| 2021 | Aglianico | Sicilia | Main | 2.37 ± 1.25 c | 1.99 ± 1.09 b | 2.46 ± 1.44 c | 2.19 ± 0.81 n.s. |
| Lateral | 2.22 ± 1.07 ab | 1.75 ± 0.96 ab | 2.27 2.32 ab | 2.22 0.76 n.s. | |||
| Campania | Main | 2.88 ± 1.23 ab | 1.37 ± 0.30 c | 2.89 ± 0.80 ab | 3.02 ± 0.93 n.s. | ||
| Lateral | 1.74 ± 1.11 bc | 1.36 ± 0.24 c | 3.07 ± 1.16 a | 2.71 ± 0.99 n.s. | |||
| Molise | Main | 3.62 ± 1.16 a | 2.11 ± 0.42 a | 2.47 ± 0.54 ab | 2.41 ± 0.84 n.s. | ||
| Lateral | 3.65 4.49 a | 1.68 ± 0.44 bc | 1.98 ± 0.27 b | 2.37 ± 0.80 n.s. | |||
| Cabernet Sauvignon | Sicilia | Main | 2.20 ± 0.50 b | 2.32 ± 0.45 ab | 3.26 ± 2.14 n.s. | 3.99 ± 1.20 a | |
| Lateral | 1.72 ± 0.57 b | 2.22 ± 1.07 ab | 3.45 ± 2.45 n.s. | 3.45 ± 2.55 abc | |||
| Campania | Main | 5.30 ± 1.99 a | 2.82 ± 0.94 a | 3.46 ± 1.34 n.s. | 4.55 ± 1.14 a | ||
| Lateral | 5.54 ± 2.71 a | 2.57 ± 0.86 a | 3.24 ± 1.01 n.s. | 4.19 ± 1.32 ab | |||
| Molise | Main | 4.49 ± 0.97 a | 1.68 ± 0.35 bc | 2.63 ± 0.95 n.s. | 2.42 ± 0.75 c | ||
| Lateral | 4.39 ± 1.86 a | 1.46 ± 0.50 c | 2.52 ± 0.99 n.s. | 2.87 ± 0.99 c | |||
| Parameter | Aglianico | |||||
|---|---|---|---|---|---|---|
| Molise | Campania | Sicily | ||||
| 2020 | 2021 | 2020 | 2021 | 2020 | 2021 | |
| Yield/vine (kg) | 6.80 ± 1.88 A | 9.10 ± 1.31 a | 2.11 ± 0.71 B | 6.24 ± 2.22 b | 0.24 ± 1.07 B | 0.44 ± 1.05 b |
| Bunch weight (g) | 558.60 ± 161.5 A | 345.80 ± 42.08 n.s. | 348 ± 56.35 AB | 298.20 ± 104.28 n.s. | 190 ± 15.41 B | 315.40 ± 85.23 n.s. |
| Berry weight (g) | 2.64 ± 0.22 a | 2.59 ± 0.22 b | 2.58 ± 0.17 a | 2.19 ± 0.27 a | 2.15 ± 0.03 b | 2.38 ± 0.19 ab |
| Total soluble solids (°Brix) | 21.59 ± 0.63 n.s. | 22.23 ± 0.88 A | 20.54 ± 0.55 n.s. | 23.23 ± 0.38 A | 21.06 ± 0.21 n.s. | 20.04 ± 0.73 B |
| pH | 3.48 ± 0.21 b | 3.34 ± 0.08 B | 3.85 ± 0.13 a | 3.63 ± 0.05 A | 3.5 ± 0.03 ab | 3.71 ± 0.13 A |
| Titratable acidity (g L−1) | 10.40 ± 0.03 A | 17.1 ± 0.41 A | 9.80 ± 0.02 A | 8.00 ± 0.04 B | 5.30 ± 0.04 B | 7.6 ± 0.01 B |
| Cabernet Sauvignon | ||||||
| Molise | Campania | Sicily | ||||
| 2020 | 2021 | 2020 | 2021 | 2020 | 2021 | |
| Yield/vine (kg) | 4.84 ± 0.95 A | 8.57 ± 1.56 A | - | 4.96 ± 2.31 B | 2.45 ± 0.43 B | 2.79 ± 0.66 B |
| Bunch weight (g) | 352.60 ± 74.82 a | 444.6 ± 64.92 n.s. | - | 318.80 ± 102.27 n.s. | 258.40 ± 25.38 b | 346.60 ± 56.68 n.s. |
| Berry weight (g) | 1.97 ± 0.17 n.s. | 2.75 ± 0.18 A | - | 1.89 ± 0.22 b | 1.90 ± 0.12 n.s. | 1.50 ± 0.23 c |
| Total soluble solids (°Brix) | 22.33 ± 0.89 A | 24.45 ± 0.10 A | - | 23.00 ± 0.89 AB | 19.91 ± 0.04 B | 21.67 ± 0.98 B |
| pH | 4.01 ± 0.35 A | 3.80 ± 0.20 n.s. | - | 4.23 ± 0.37 n.s. | 3.61 ± 0.14 B | 3.74 ± 0.05 n.s. |
| Titratable acidity (g L−1) | 9.8 ± 0.05 A | 7.7 ± 0.02 n.s. | - | 7.60 ± 0.01 n.s. | 5.1 ± 0.01 B | 7.5 ± 0.00 n.s. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferlito, F.; Nicolosi, E.; Sicilia, A.; Villano, C.; Aversano, R.; Lo Piero, A.R. Physiological and Productive Responses of Two Vitis vinifera L. Cultivars across Three Sites in Central-South Italy. Horticulturae 2023, 9, 1321. https://doi.org/10.3390/horticulturae9121321
Ferlito F, Nicolosi E, Sicilia A, Villano C, Aversano R, Lo Piero AR. Physiological and Productive Responses of Two Vitis vinifera L. Cultivars across Three Sites in Central-South Italy. Horticulturae. 2023; 9(12):1321. https://doi.org/10.3390/horticulturae9121321
Chicago/Turabian StyleFerlito, Filippo, Elisabetta Nicolosi, Angelo Sicilia, Clizia Villano, Riccardo Aversano, and Angela Roberta Lo Piero. 2023. "Physiological and Productive Responses of Two Vitis vinifera L. Cultivars across Three Sites in Central-South Italy" Horticulturae 9, no. 12: 1321. https://doi.org/10.3390/horticulturae9121321
APA StyleFerlito, F., Nicolosi, E., Sicilia, A., Villano, C., Aversano, R., & Lo Piero, A. R. (2023). Physiological and Productive Responses of Two Vitis vinifera L. Cultivars across Three Sites in Central-South Italy. Horticulturae, 9(12), 1321. https://doi.org/10.3390/horticulturae9121321