
Effect of the Ozone Application in the Nutrient Solution and the Yield and Oxidative Stress of Hydroponic Baby Red Chard


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Raw Material
2.2. Plant Tissue Evaluations
- Ca: Chlorophyll a content (mg·g−1 fresh weight, fw)
- Cb: Chlorophyll b content (mg·g−1 fresh weight, fw)
- A663: sample absorbance measured at 663 nm
- A646: sample absorbance measured at 646 nm
2.3. Determinations of the Nutritive Solution
2.4. Determinations Associated with Oxidative Stress in the Leaves
2.4.1. Antioxidant Capacity
2.4.2. Oxidative Stress Enzymes
2.5. Experimental Design and Statistical Analysis
3. Results and Discussion
3.1. Evaluation of Plant Growth
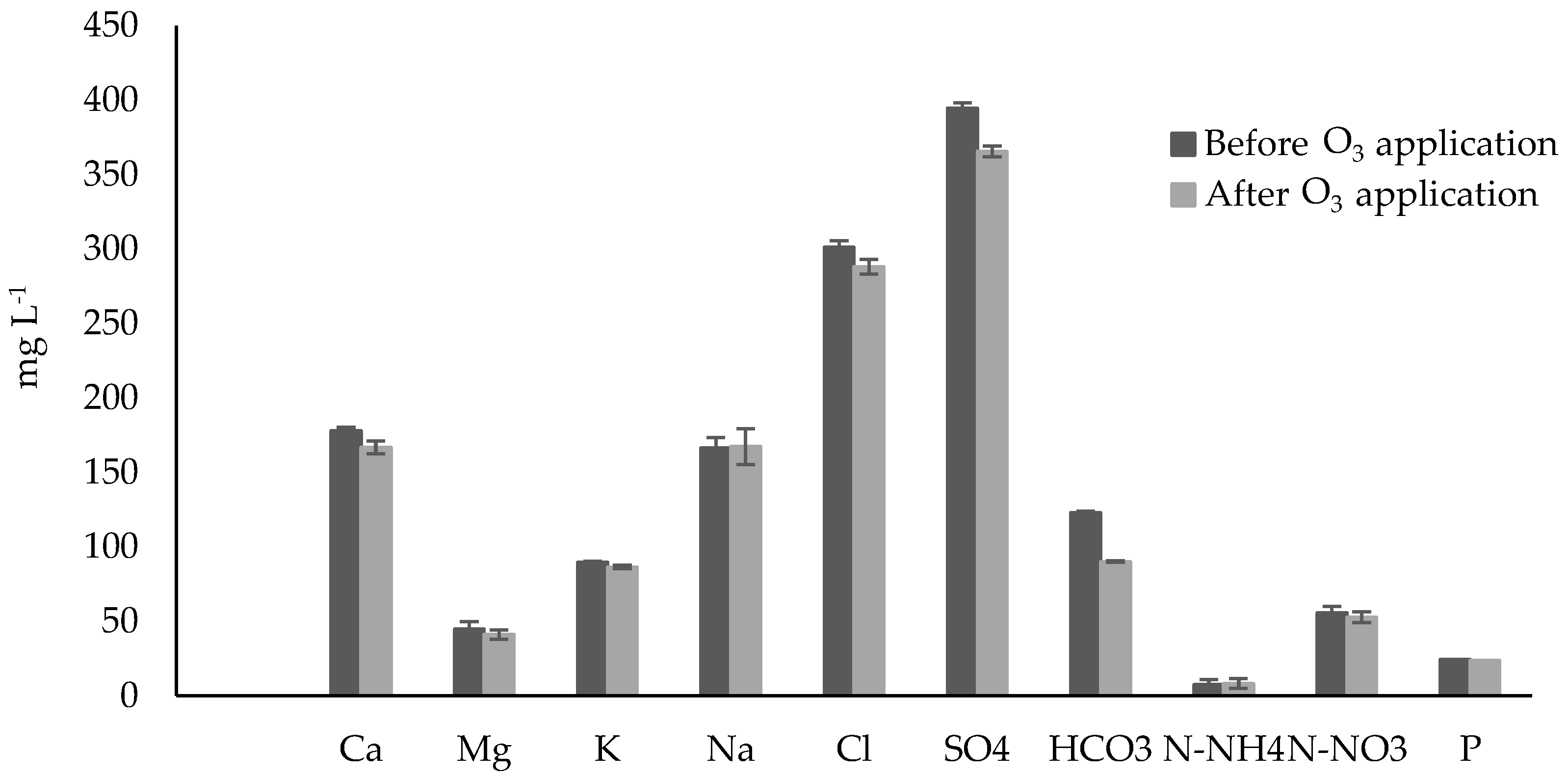
3.2. Chemical Analysis in the Nutrient Solution
3.3. Oxidative Stress in the Leaves
3.3.1. Total Phenol Content
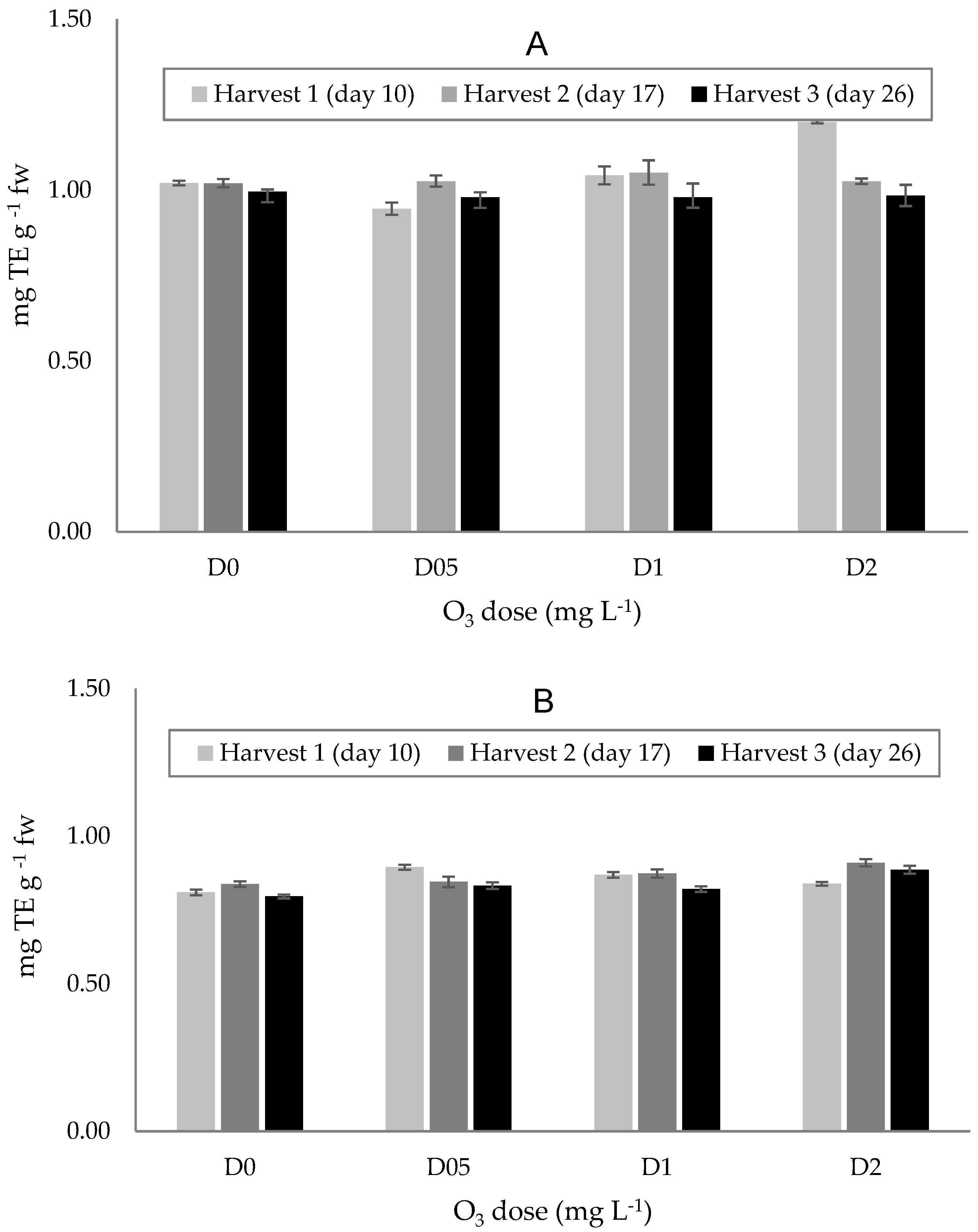
3.3.2. Antioxidant Capacity
3.4. Oxidative Stress Enzymes
3.4.1. Superoxide Dismutase
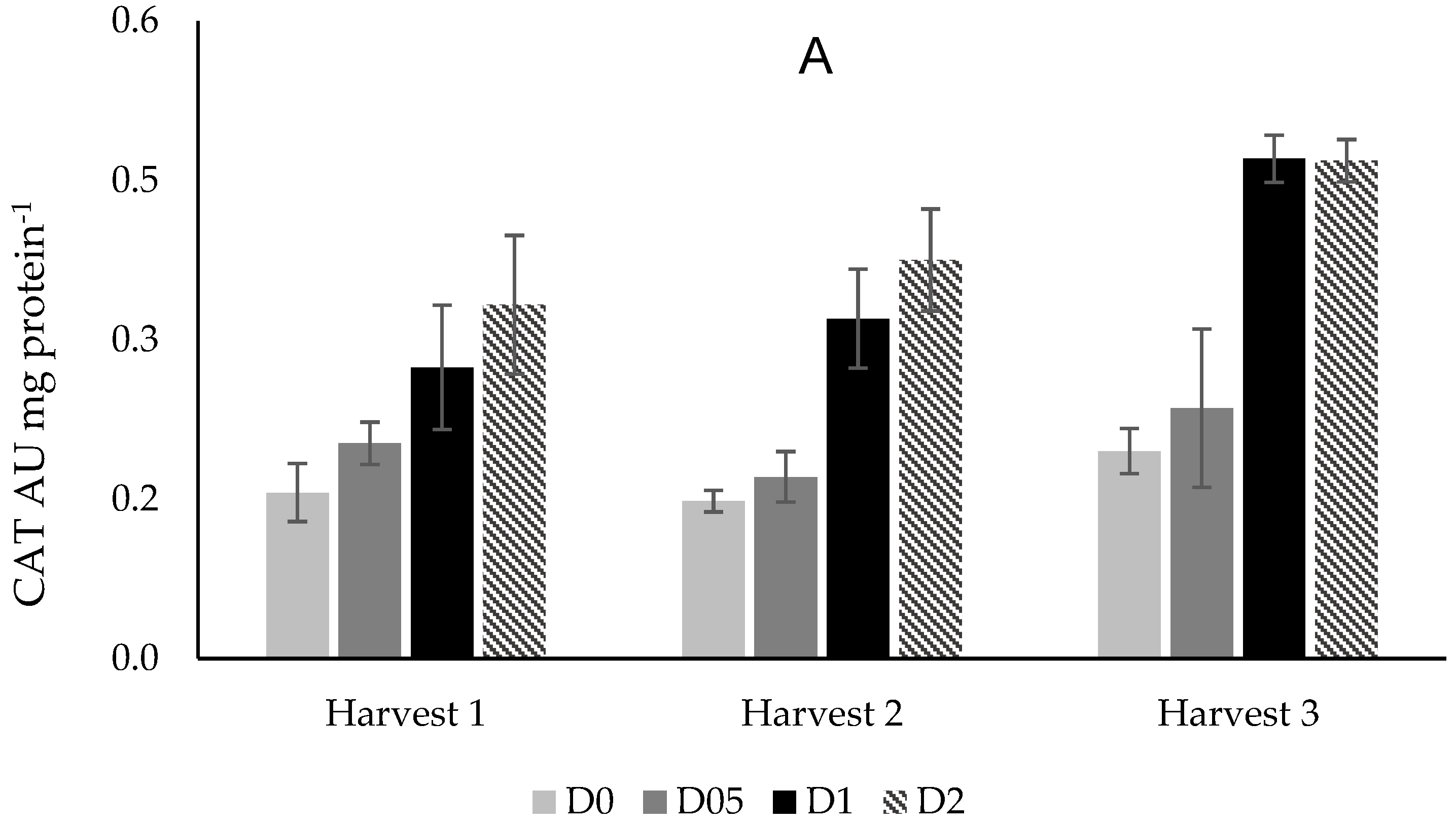
3.4.2. Catalase
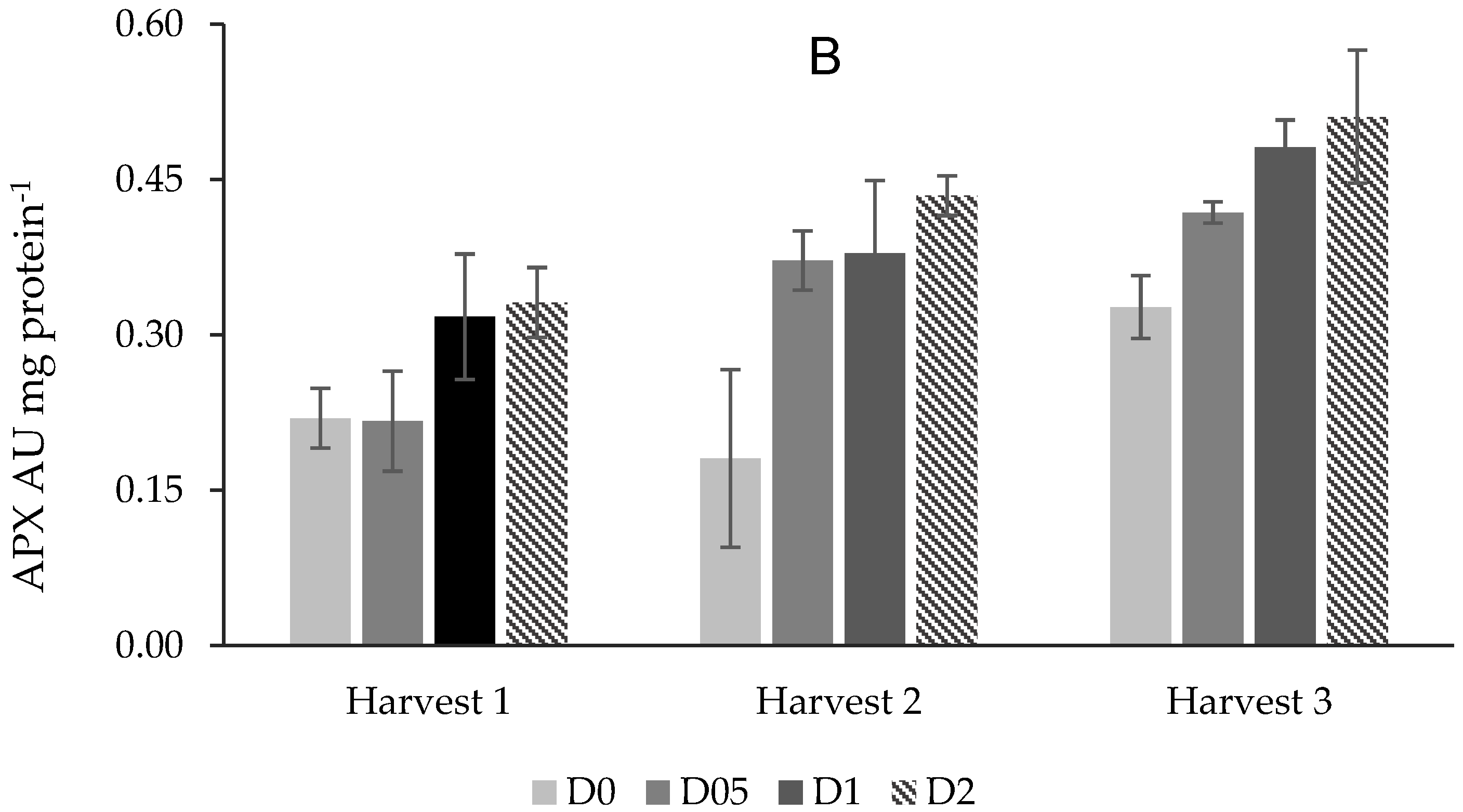
3.4.3. Ascorbate Peroxidase
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pfister, S.P.; Bayer, P.; Koehler, A.; Hellweg, S. Projected water consumption in future global agriculture: Scenarios and related impacts. Sci. Total Environ. 2011, 409, 4206–4216. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cai, H.; Jiang, T.; Sun, S.; Wang, Y.; Zhao, J.; Yu, X.; Sun, J. Assessment of global drought propensity and its impacts on agricultural water use in future climate scenarios. Agric. For. Meteorol. 2019, 278, 107623. [Google Scholar] [CrossRef]
- Schmidt, L.; Galvão, A.; Santos, R.; Monteiro, S. Impact of water reuse on agricultural practices and human health. Environ. Res. 2023, 216, 114762. [Google Scholar]
- Graham, T.; Zhang, P.; Woyzbun, E.; Dixon, M. Response of hydroponic tomato to daily applications of aqueous ozone via drip irrigation. Sci. Hort. 2011, 129, 464–471. [Google Scholar] [CrossRef]
- Hosseinzadeh, S.; Bonarrigo, G.; Verheust, Y.; Roccaro, P.; Van Hulle, S. Water reuse in closed hydroponic systems: Comparison of GAC adsorption, ion exchange and ozonation processes to treat recycled nutrient solution. Aquac. Eng. 2017, 78, 190–195. [Google Scholar] [CrossRef]
- Ikeda, H.; Koohakan, P.; Jaenaksorn, T. Problems and countermeasures in the re-use of the nutrient solution in soilless production. Acta Hort. 2002, 578, 213–219. [Google Scholar] [CrossRef]
- Green, H.; Kidd, J.; Jackson, L. Novel and Emerging Cleaning and Sanitization Technologies, Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2023; pp. 1–7. [Google Scholar]
- Han, Y.J.; Gharibeshghi, A.; Mewis, I.; Förster, N.; Beck, W.; Ulrichs, C. Plant responses to ozone. Effects of different ozone exposure durations on plant growth and biochemical quality of Brassica campestris L. ssp. Chinensis. Sci. Hortic. 2020, 262, 108921. [Google Scholar] [CrossRef]
- Glaze, W.H. Drinking-water treatment with ozone. Environ. Sci. Technol. 1987, 21, 224–230. [Google Scholar] [CrossRef]
- Singh, D.; Rai, K.; Gupta, A.; Bhushan, S.; Agrawal, M. Chapter Two—Cross-talk between ROS and phytohormones signaling determines crop sensitivity against ozone. In Advances in Botanical Research; Pui, F., Ed.; Academic Press: Cambridge, MA, USA, 2023; Volume 108, pp. 47–81. [Google Scholar]
- Chhem-Kieth, S.; Holm, L.; Rosenfjeld, M.; Larsen, M. Effects of vegetables and fruit with varying physical damage, fungal infection, and soil contamination on stability of aqueous ozone. Food Biosci. 2022, 50, 102157. [Google Scholar] [CrossRef]
- Ueda, Y.; Uehara, N.; Sasaki, H.; Kobayashi, K.; Yamakawa, T. Impacts of acute ozone stress on superoxide dismutase (SOD) expression and reactive oxygen species (ROS) formation in rice leaves. Plant Physiol. Biochem. 2013, 70, 396–402. [Google Scholar] [CrossRef]
- Tiwari, B.K.; O’Donnell, C.P.; Patras, A.; Brunton, N.; Cullen, P.J. Effect of ozone processing on anthocyanins and ascorbic acid degradation of strawberry juice. Food Chem. 2009, 113, 1119–1126. [Google Scholar] [CrossRef]
- Risoli, S.; Lauria, G. Ozonated water application as an innovative tool for elicitation of plant defense response: A minireview. Curr. Opin. Environ. Sci. Health 2022, 28, 100375. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, L.; Dixon, M. An upper limit for elevated root zone dissolved oxygen concentration for tomato. Sci. Hort. 2007, 113, 162–165. [Google Scholar] [CrossRef]
- Martínez-Sánchez, A.; Aguayo, E. Effect of irrigation with ozonated water on the quality of capsicum seedlings grown in the nursery. Agric. Water Manag. 2019, 221, 547–555. [Google Scholar] [CrossRef]
- Díaz-López, M.; Siles, J.A.; Ros, C.; Bastida, F.; Nicolás, E. The effects of ozone treatments on the agro-physiological parameters of tomato plants and the soil microbial community. Sci. Total Environ. 2022, 812, 151429. [Google Scholar] [CrossRef]
- Peng, Y.; Yan-zhi, G.; Ling, Q. Effects of ozone-treated domestic sludge on hydroponic lettuce growth and nutrition. J. Integr. Agric. 2018, 17, 593–602. [Google Scholar]
- Nagatomo, T.; Abiru, T.; Mitsugi, F.; Ebihara, K.; Nagahama, K. Study on ozone treatment of soil for agricultural application of surface dielectric barrier discharge. Jpn. J. Appl. Phys. 2015, 55, 01AB06. [Google Scholar] [CrossRef]
- Ikeura, H.; Goto, T.; Tamaki, M. Effects of adding a chelator after ozone microbubble generation on nutrient composition, medium sterility, and plant growth. Water Air Soil Pollut. 2018, 229, 1. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil, Circular-347; California Agricultural Experiment Station, The College of Agriculture University of California Berkeley: Berkeley, CA, USA, 1950; pp. 1–32. [Google Scholar]
- Kobayashi, F.; Ikeura, H.; Ohsato, S.; Goto, T.; Tamaki, M. Disinfection using ozone microbubbles to inactivate Fusarium oxysporum f. sp. melonis and Pectobacterium carotovorum subsp. carotovorum. Crop Prot. 2011, 30, 1514–1518. [Google Scholar] [CrossRef]
- MINSAL. Reglamento Sanitario de Alimentos, 1997 (Actualized 2023). Available online: https://www.dinta.cl/wp-content/uploads/2023/03/RSA-decreto-977-96-act-al-26-01-23.pdf (accessed on 22 May 2023).
- Lichtenthaler, K.; Welburn, A.R. Determination of total carotenoids and chlorophylls A and B of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Sadzawka, A.; Carrasco, M.A.; Demanet, R.; Flores, H.; Grez, R.; Mora, M.L.; Neaman, A. Métodos de Análisis de Tejidos Vegetales, 2nd ed.; Serie Actas INIA N°40; Instituto de Investigaciones Agropecuarias (INIA): Santiago, Chile, 2007; 140p. [Google Scholar]
- Rodrigo, J.G.K.; Organo, V.G. Universal pH indicator as a colorimetric reagent for differentiating inorganic anions. Orient. J. Chem. 2020, 36, 442–445. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagent. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. LWT—Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Benzie, I.; Strain, J. Ferric reducing ability of plasma (FRAP) as a measure of antioxidant power: The FRAP assay. Anal Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Lemoine, M.L.; Civello, P.M.; Chaves, A.R.; Martıinez, G.A. Influence of a combined hot air and UV-C treatment on quality parameters of fresh-cut broccoli florets at 0 °C. Food Sci. Technol. 2010, 45, 1212–1218. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplast. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar]
- Dhindsa, R.S.; Plumb-Dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of micro-gram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sarkar, A.; Agrawal, S.B. Elevated ozone and two modern wheat cultivars: An assessment of dose dependent sensitivity with respect to growth, reproductive and yield parameters. Environ. Exp. Bot. 2010, 69, 328–337. [Google Scholar] [CrossRef]
- Kleiber, T.; Borowiak, K.; Schroeter-Zakrzewska, A.; Budka, A.; Osiecki, S. Effect of ozone treatment and light colour on photosynthesis and yield of lettuce. Sci. Hortic. 2017, 217, 130–136. [Google Scholar] [CrossRef]
- Malaiyandi, M.; Natarajan, M. Impact of ozone on morphological, physiological, and biochemical changes in cowpea (Vigna unguiculata (L.) Walp.). Ozone Sci. Eng. 2014, 36, 36–42. [Google Scholar] [CrossRef]
- Knudson, L.L.; Tibbitts, T.W.; Edwards, G.E. Measurement of ozone injury by determination of leaf chlorophyll concentration. Plant Physiol. 1977, 60, 606–608. [Google Scholar] [CrossRef] [PubMed]
- Heagle, A.S.; Miller, J.E.; Booker, F.L. Influence of ozone stress on soybean response to carbon dioxide enrichment: I. Foliar properties. Crop Sci. 1998, 38, 113–121. [Google Scholar] [CrossRef]
- Bindi, M.; Hacour, A.; Vandermeiren, K.; Craigon, J.; Ojanperä, K.; Selldén, G.; Högy, P.; Finnan, J.; Fibbi, L. Chlorophyll concentration of potatoes grown under elevated carbon dioxide and/or ozone concentrations. Eur. J. Agron. 2002, 17, 319–335. [Google Scholar] [CrossRef]
- Ismail, M.; Basahi, J.M.; Hassan, I.A. Gas exchange and chlorophyll fluorescence of pea (Pisum sativum L.) plants in response to ambient ozone at a rural site in Egypt. Sci. Total Environ. 2014, 497–498, 585–593. [Google Scholar] [CrossRef]
- Salvatori, E.; Fusaro, L.; Mereu, S.; Bernardini, A.; Puppi, G.; Manes, F. Different O3 response of sensitive and resistant snap bean genotypes (Phaseolus vulgaris L.): The key role of growth stage, stomatal conductance, and PSI activity. Environ. Exp. Bot. 2012, 87, 79–91. [Google Scholar] [CrossRef]
- Gottardini, E.; Cristofori, A.; Cristofolini, F.; Nali, C.; Pellegrini, E.; Bussotti, F.; Ferretti, M. Chlorophyll-related indicators are linked to visible ozone symptoms: Evidence from a field study on native Viburnum lantana L. plants in northern Italy. Ecol. Indic. 2014, 39, 65–74. [Google Scholar] [CrossRef]
- Harrison, J.P. Ozone for Point-of-Use, Point-of-Entry, and Small Water System Water Treatment Application; Water Quality Association, Ed.; Water Quality Association: Lisle, IL, USA, 2004. [Google Scholar]
- Vanachter, A.; Thys, L.; Van Wambeke, E.; Van Assche, C. Possible use of ozone [sic] for disinfestation of plant nutrient solutions. Symp. Hort. Substrates Anal. 1987, 221, 295–302. [Google Scholar]
- Alzate, M.Y.; Cardarelli, M.; Rouphael, Y.; Cesco, S.; Pii, Y.; Colla, G. Iron nutrition in agriculture: From synthetic chelates to biochelates. Sci. Hortic. 2023, 312, 111833. [Google Scholar]
- Bughio, N.; Takahashi, M.; Yoshimura, E.; Nishizawa, N.K.; Mori, S. Light- dependent iron transport into isolated barley chloroplasts. Plant Cell Physiol. 1997, 38, 101–105. [Google Scholar] [CrossRef]
- Kosegarten, H.; Wilson, G.H.; Esch, A. The effect of nitrate nutrition on iron chlorosis and leaf growth in sunflower (Helianthus annuus L.). Eur. J. Agron. 1998, 8, 283–292. [Google Scholar] [CrossRef]
- Donnini, S.; Dell’Orto, M.; Zocchi, G.; Tognetti, R. Oxidative stress responses and root lignification induced by Fe deficiency conditions in pear and quince genotypes. Tree Physiol. 2011, 31, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Yadavalli, V.; Neelam, S.; Rao, A.S.V.C.; Reddy, A.R.; Subramanyam, R. Differential degradation of photosystem I subunits under iron deficiency in rice. J. Plant Physiol. 2012, 169, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gopal, R.; Dube, B.K. Impact of iron stress on biomass, yield, metabolism and quality of potato (Solanum tuberosum L.). Sci. Hortic. 2006, 108, 1–6. [Google Scholar] [CrossRef]
- Jin, C.W.; Du, S.T.; Chen, W.W.; Li, G.X.; Zhang, Y.S.; Zheng, S.J. Elevated carbon dioxide improves plant Fe nutrition through enhancing the Fe- deficiency-induced responses under Fe-limited conditions in tomato. Plant Physiol. 2009, 150, 272–280. [Google Scholar] [CrossRef]
- Briat, J.F.; Dubos, C.; Gaymard, F. Iron nutrition, biomass production, and plant product quality. Trends Plant Sci. 2015, 20, 33–40. [Google Scholar] [CrossRef]
- Jin, C.W.; Liu, Y.; Mao, Q.Q.; Wang, Q.; Du, S.T. Mild Fe-deficiency improves biomass production and quality of hydroponic-cultivated spinach plants (Spinacia oleracea L.). Food Chem. 2013, 138, 2188–2194. [Google Scholar] [CrossRef]
- Wang, H.Y.; Garvin, D.F.; Kochian, L.V. Rapid induction of regulatory and transporter genes in response to phosphorus, potassium, and iron deficiencies in tomato roots. Evidence for cross talk and root/rhizosphere-mediated signals. Plant Physiol. 2002, 130, 1361–1370. [Google Scholar] [CrossRef]
- Wang, H.Y.; Garvin, D.F.; Kochian, L.V. Nitrate-induced genes in tomato roots. Array analysis reveals novel genes that may play a role in nitrogen nutrition. Plant Physiol. 2001, 127, 345–359. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2010; 782p. [Google Scholar]
- Virgili, A. Introducción a la Fertilización con Microelementos, 2nd ed.; Welgro Micromix: Barcelona, Spain, 1996; 25p. [Google Scholar]
- Vernon, G. Ozone Efficacy on Reduction of Phytophthora capsici in Recirculated Horticultural Irrigation Water. Ph.D. Thesis, Texas A&M University, College Station, TX, USA, 2007; 131p. [Google Scholar]
- Saviranta, N.; Julkunen-Tiitto, R.; Oksanen, E.; Karjalainen, R.O. Leaf phenolic compounds in red clover (Trifolium pratense L.) induced by exposure to moderately elevated ozone. Environ. Poll. 2010, 158, 440–446. [Google Scholar] [CrossRef]
- Li, G.; Shi, Y.; Chen, X. Effects of elevated CO2 and O3 on phenolic compounds in spring wheat and maize leaves. Bull Environ. Contam. Toxicol. 2008, 81, 436–439. [Google Scholar] [CrossRef]
- Lee, J.H.; Goto, E. Ozone control as a novel method to improve health-promoting bioactive compounds in red leaf lettuce (Lactuca sativa L.). Front. Plant Sci. 2022, 13, 1045239. [Google Scholar] [CrossRef] [PubMed]
- Caregnato, F.F.; Bortoli, R.C.; Molina, A.; Junior, D.; Fonseca Moreira, J.C. Exposure to elevated ozone levels differentially affects the antioxidant capacity and the redox homeostasis of two subtropical Phaseolus vulgaris L. varieties. Chemosphere. 2013, 93, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Severino, J.F.; Stich, K.; Soja, G. Ozone stress and antioxidant substances in Trifolium repens and Centaurea jacea leaves. Environ. Poll. 2007, 146, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Alscher, R.G.; Erturk, N.; Heath, L.S. Role of superoxide dismutases (SODs) in controlling oxidative stress in plants. J. Exp. Bot. 2010, 53, 1331–1341. [Google Scholar] [CrossRef]
- Alonso, R.; Elvira, S.; Castillo, F.J.; Gimeno, B.S. Interactive effects of ozone and drought stress on pigments and activities of antioxidative enzymes in Pinus halepensis. Plant Cell Environ. 2001, 24, 905–916. [Google Scholar] [CrossRef]
- Rao, M.V.; Paliyath, G.; Ormrod, D.P. Ultraviolet-B-and ozone-induced biochemical changes in antioxidant enzymes of Arabidopsis thaliana. Plant Physiol. 1996, 110, 125136. [Google Scholar] [CrossRef]
- Calatayud, A.; Iglesias, D.J.; Talón, M.; Barreno, E. Effects of 2-month ozone exposure in spinach leaves on photosynthesis, antioxidant systems and lipid peroxidation. Plant Physiol. Biochem. 2003, 41, 839–845. [Google Scholar] [CrossRef]
- Caregnato, F.F.; Clebsch, C.C.; Rocha, R.F.; Feistauer, L.B.H.; Oliveira, P.L.; Divan, A.D.; Moreira, J.C.F. Ozone exposure differentially affects oxidative stress parameters in distinct Phaseolus vulgaris L. varieties. J. Plant Interact. 2010, 5, 111–115. [Google Scholar] [CrossRef]
- Sarkar, A.; Singh, A.A.; Agrawal, A.B.; Ahmad, A.; Rai, S.P. Cultivar specific variations in antioxidative defense system, genome and proteome of two tropical rice cultivars against ambient and elevated ozone. Ecotoxicol. Environ. Saf. 2015, 115, 101–111. [Google Scholar] [CrossRef]
- Almeselmani, M.; Deshmukh, P.S.; Sairam, R.K.; Kushwaha, S.R.; Singh, T.P. Protective role of antioxidant enzymes under high temperature stress. Plant Sci. 2006, 171, 382–388. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, Q.; Zhu, J.; Liu, G.; Tang, H. Dissimilarity of ascorbate–glutathione (AsA–GSH) cycle mechanism in two rice (Oryza sativa L.) cultivars under experimental free-air ozone exposure. Agric. Ecosyst. Environ. 2013, 165, 39–49. [Google Scholar] [CrossRef]
- Mehlhorn, H.; Cottam, D.A.; Lucas, P.W.; Wellburn, A.R. Induction of ascorbate peroxidase and glutathione reductase activities by interaction of mixtures of air pollutants. Free Radic. Res. Commun. 1987, 3, 193–197. [Google Scholar] [CrossRef]
- Rao, M.; Koch, J.; Davis, K. Ozone: A tool for probing programmed cell death in plants. Plant Mol. Biol. 2000, 44, 345–358. [Google Scholar] [CrossRef]
- Kangasjävi, J.; Jaspers, P.; Kollist, H. Signalling and cell death in ozone- exposed plants. Plant Cell Environ. 2005, 28, 1021–1036. [Google Scholar] [CrossRef]
- Astorino, G.; Margani, I.; Tripodo, P.; Manes, F. The response of Phaseolus vulgaris L. cv. Lit. to different dosage of the antiozonant ethylenediurea (EDU) in relation to chronic treatment with ozone. Plant Sci. 1995, 111, 237–248. [Google Scholar] [CrossRef]
- Liu, X.; Sui, L.; Huang, Y.; Geng, C.; Yin, B. Physiological and visible injury responses in different growth stages of winter wheat to ozone stress and the protection of spermidine. Atmos. Pollut. Res. 2015, 6, 596–604. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| O3 Dose (mg·L−1) | Fresh Weight (g) | ||
|---|---|---|---|
| Harvest 1 (day 10) | Harvest 2 (day 17) | Harvest 3 (day 26) | |
| D0 | 11.56 ± 0.38 1 | 10.86 ± 0.84 | 11.56 ± 0.38 |
| D05 | 14.79 ± 0.49 | 15.14 ± 0.25 | 13.12 ± 0.39 |
| D1 | 11.13 ± 0.12 | 12.06 ± 0.76 | 11.13 ± 0.12 |
| D2 | 10.25 ± 0.28 | 11.73 ± 0.41 | 11.25 ± 0.13 |
| Significance level | |||
| O3 dose (D) | **** (1.49) 2,3 | ||
| Harvest time (H) | NS 3 | ||
| D × H | NS |
| O3 Dose (mg·L−1) | Fresh Weight (g) |
|---|---|
| D0 | 11.63 b 1 |
| D05 | 14.35 a |
| D1 | 11.32 b |
| D2 | 11.44 b |
| O3 Dose (mg·L−1) | Dry Weight (%) | ||
|---|---|---|---|
| Harvest 1 (day 10) | Harvest 2 (day 17) | Harvest 3 (day 26) | |
| D0 | 15.23 ± 0.71 1 | 12.87 ± 0.39 | 13.37 ± 0.27 |
| D05 | 12.87 ± 1.07 | 10.17 ± 0.23 | 11.63 ± 0.22 |
| D1 | 11.88 ± 0.63 | 11.85 ± 0.30 | 12.45 ± 0.39 |
| D2 | 14.52 ± 1.65 | 13.31 ± 0.36 | 13.10 ± 0.23 |
| Significance level | |||
| O3 dose (D) | * (1.49) 2 | ||
| Harvest time (H) | NS 3 | ||
| D × H | NS |
| O3 Dose (mg·L−1) | Dry Weight (%) |
|---|---|
| D0 | 13.82 a 1 |
| D05 | 11.56 b |
| D1 | 12.06 ab |
| D2 | 13.65 ab |
| O3 Dose (mg·L−1) | Leaf Area (cm2) | ||
|---|---|---|---|
| Harvest 1 (day 10) | Harvest 2 (day 17) | Harvest 3 (day 26) | |
| D0 | 11.44 ± 0.20 1 | 11.02 ± 0.37 | 10.83 ± 0.62 |
| D05 | 15.00 ± 0.40 | 15.21 ± 0.26 | 14.91 ± 0.35 |
| D1 | 12.78 ± 0.78 | 12.21 ± 0.25 | 12.10 ± 0.36 |
| D2 | 8.98 ± 0.38 | 11.97 ± 0.61 | 10.71 ± 0.13 |
| Significance level | |||
| O3 dose (D) | **** (1.68) 2 | ||
| Harvest time (H) | NS 3 | ||
| D x H | NS |
| O3 Dose (mg·L−1) | Leaf Area (cm2) |
|---|---|
| D0 | 10.55 a 1 |
| D05 | 15.04 c |
| D1 | 11.09 ab |
| D2 | 12.37 b |
| O3 Dose (mg·L−1) | Harvest 1 (Day 10) | Harvest 2 (Day 17) | Harvest 3 (Day 26) | |
|---|---|---|---|---|
| D0 | 1.02 Aa 1,2 | 0.80 Ba | 0.81 Aa | |
| D05 | 0.79 Aa | 0.70 Ba | 0.76 Aa | |
| Chla | D1 | 0.63 BCb | 0.86 Aba | 0.69 Bab |
| D2 | 0.49 Cc | 0.95 Aa | 0.72 Bb | |
| D0 | 1.34 Aba | 1.33 Ba | 1.34 Aa | |
| D05 | 1.33 Bb | 1.35 Aba | 1.33 Aa | |
| Chlb | D1 | 1.32 Bb | 1.36 Aa | 1.34 Aa |
| D2 | 1.36 Aa | 1.35 Aa | 1.34 Aa |
| O3 Dose (mg·L−1) | Total Phenol Content (mg GAE·g−1 fw) | ||
|---|---|---|---|
| Harvest 1 (day 10) | Harvest 2 (day 17) | Harvest 3 (day 26) | |
| D0 | 0.91 ± 0.07 1 | 0.92 ± 0.03 | 0.88 ± 0.04 |
| D05 | 0.88 ± 0.05 | 0.87 ± 0.05 | 0.90 ± 0.03 |
| D1 | 0.91 ± 0.06 | 0.89 ± 0.02 | 0.86 ± 0.04 |
| D2 | 0.94 ± 0.05 | 0.90 ± 0.07 | 0.87 ± 0.05 |
| Significance level | |||
| O3 dose (D) | NS 2 | ||
| Harvest time (H) | NS | ||
| D × H | NS |
| O3 Dose (mg·L−1) | SOD (AU mg·protein−1) |
|---|---|
| D0 | 2.42 b 1 |
| D05 | 6.32 a |
| D1 | 5.11 a |
| D2 | 6.20 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machuca Vargas, A.; Silveira Gómez, A.C.; Hernández-Adasme, C.; Escalona Contreras, V.H. Effect of the Ozone Application in the Nutrient Solution and the Yield and Oxidative Stress of Hydroponic Baby Red Chard. Horticulturae 2023, 9, 1234. https://doi.org/10.3390/horticulturae9111234
Machuca Vargas A, Silveira Gómez AC, Hernández-Adasme C, Escalona Contreras VH. Effect of the Ozone Application in the Nutrient Solution and the Yield and Oxidative Stress of Hydroponic Baby Red Chard. Horticulturae. 2023; 9(11):1234. https://doi.org/10.3390/horticulturae9111234
Chicago/Turabian StyleMachuca Vargas, Alejandra, Ana Cecilia Silveira Gómez, Cristian Hernández-Adasme, and Víctor Hugo Escalona Contreras. 2023. "Effect of the Ozone Application in the Nutrient Solution and the Yield and Oxidative Stress of Hydroponic Baby Red Chard" Horticulturae 9, no. 11: 1234. https://doi.org/10.3390/horticulturae9111234
APA StyleMachuca Vargas, A., Silveira Gómez, A. C., Hernández-Adasme, C., & Escalona Contreras, V. H. (2023). Effect of the Ozone Application in the Nutrient Solution and the Yield and Oxidative Stress of Hydroponic Baby Red Chard. Horticulturae, 9(11), 1234. https://doi.org/10.3390/horticulturae9111234