Abstract
Drought stress reduces water availability in plant cells and influences rapeseed yield. Currently, key genetic regulators that contribute to rapeseed response to drought remain largely unexplored, which limits breeding of drought-resistant rapeseed. In this study, we found that Brassica napus JASMONATE ZIM-DOMAIN 5 (BnA.JAZ5), one of the transcriptional repressors functioning in the jasmonate (JA) signaling pathway, was triggered by drought treatment in rapeseed, and drought-susceptibility increased in BnA.JAZ5-overexpressing rapeseed plants as compared to wild-type plants, resulting in a lower survival rate after recovery from dehydration. After recovery for 3 days, 22–40% of p35S::BnA.JAZ5 transgenic plants survived, while approximately 61% of wild-type plants survived. Additionally, seed germination of BnA.JAZ5-overexpressing rapeseed was hyposensitive to abscisic acid (ABA). The germination rate of five transgenic lines was 32~42% under 9 µM ABA treatment, while the germination rate of wild-type plants was 14%. We also found that the average stomatal density of five overexpressing lines was 371~446/mm2, which is higher than that of wild-type (232/mm2) plants under normal conditions. These results indicate that BnA.JAZ5 regulated drought response in an ABA-dependent manner, possibly by affecting stomatal density. Interestingly, methyl jasmonate (MeJA) treatment rescued the ABA-hyposensitive seed germination, revealing crosstalk between JAZ5-meidated JA and the ABA signaling pathway. Taken together, our results suggest that BnA.JAZ5 attenuated drought resistance through the ABA-dependent pathway, which could represent important genetic loci for drought-resistant rapeseed breeding.
1. Introduction
Various environmental stresses affect plant growth and yield. Water availability is the most important abiotic factor contributing to plant evolution [1]. Establishment of seedlings is directly inhibited by drought stress, which reduces plant densities and yields [2]. With the reduction in arable land worldwide, global food production needs to be increased to feed an ever-growing population [3]. Thereby, genetic engineering must be integrated with breeding technologies to develop climate-resilient crops adaptable to environmental changes leading to stress conditions. Rapeseed (Brassica napus, AACC, 2n = 36), as an important oilseed crop used for animal fodder and human consumption, is very sensitive to environmental stresses during its growth and reproductive stages [4,5].
Plants have evolved different strategies for protection against drought. A total of 90% of soil water used by plants is lost through transpiration via the opening of stomata [5], and drought stress causes insufficient uptake of water from soil by roots to meet the requirement of plant transpiration [6,7]. Stomatal density can modulate plant transpiration rates [7,8]. Stomatal density is controlled by both genetic factors and environmental cues [8,9,10]. Overexpression of Arabidopsis thaliana GT-2 LIKE 1 (AtGTL1), AtERECTA, STOMATAL DENSITY AND DISTRIBUTION1 (AtSDD1) and Oryza sativa stress-induced protein kinase gene 1 (OsSIK1) enhances drought tolerance, which is associated with a reduction in stomatal density [7,8,11]. Drought stress induces transcription of dehydration-responsive element-binding proteins (DREBs) and activates genes involved in water movement and chaperone functions, such as late embryogenesis abundant (LEA) proteins [12,13,14,15]. RESPONSIVE TO DESICCATION 29A (RD29A), a stress-responsive marker [16], can be used as a control in stress treatments. Overexpression of the gene encoding Δ1-pyrroline-5-carboxylate synthetase 1 (P5CS1), an enzyme involved in proline biosynthesis, enhances osmotic stress tolerance [17]. Phospholipase C (PLC) works upstream of DREB2 [18].
Jasmonates (JAs) are crucial hormones that regulate plant response to abiotic and biotic stresses [19]. JASMONATE ZIM-DOMAIN (JAZ) proteins are inhibitors of the jasmonic acid signaling pathway [20]. Low jasmonoyl–isoleucine (JA-Ile) levels permit the accumulation of JAZ proteins, which can interact with several bHLH-type transcription factors, including MYC2, MYC3, and MYC4, to activate the transcription of some early JA-responsive genes [21,22,23]. JAZ proteins can also repress MYC activity to recruit NINJA and TOPLESS [24,25,26], which participate in JA-Ile perception and induce JAZ degradation [20,27,28]. Some JAZ members interact with other proteins, such as MYB75, that inhibit trichome initiation and anthocyanin accumulation [29]. Some JAZ members play roles in increasing plant tolerance to abiotic stress, while OsJAZ1 negatively regulates drought resistance by modulating JA and abscisic acid (ABA) signaling in rice [30].
Although JAZ proteins were clearly identified as transcriptional repressors of JA responses in Arabidopsis [20,21,31,32], their functions in rapeseed resistance to abiotic stress are unknown. In this study, we explore the function of BnA.JAZ5, a rapeseed (Brassica napus) homolog of JAZ5, in drought resistance, which could be a potential genetic recourse for stress-resistant plant breeding.
2. Materials and Methods
2.1. Plant Material and Growth Conditions
The rapeseed accession K407 was acquired from the Hybrid Rape Research Center of Shaanxi Province [33]. Seeds of K407 and p35S::BnA.JAZ5 plants were sterilized according to the method of Li et al. (2020) [34] and grown on Murashige and Skoog (MS) plates with 1% sucrose in darkness for 3 days at 4 °C. Then, seedings were transferred to a greenhouse for growth at 22 °C under long-day conditions (16 h light/8 h dark). The seedlings were finally transplanted to Songjiang Farm Station of the Shanghai Institute of Plant Physiology and Ecology, China in early September 2017 and 2018 [35]. To preserve the phenotypic uniformity of individual plants of the wild-type accession, potential aberrant forms were eliminated at several developmental stages.
2.2. Plasmid Construction and Transformation
The coding sequences of BnA.JAZ5 genes were amplified from cDNA of B. napus using appropriate primers (Supplementary Table S1). The sequences were inserted into a chimeric binary vector. We followed the protocol described by Moloney et al. (1989) and Verma et al. (2012) for rapeseed hypocotyl transformation [30,36], and Agrobacterium tumefaciens strain GV3101 pMP90RK was selected [34]. p35S::BnA.JAZ5 was transferred into B. napus accession K407, and transgenic plants were selected on kanamycin after Agrobacterium-mediated transformation. Positive seedlings (T0) were transplanted to the greenhouse. T1-generation p35S::BnA.JAZ5 plants were identified by PCR genotyping using transgene-specific oligos and maintained in a greenhouse as described above.
2.3. RNA Extraction and Real-Time PCR
All plant material was sampled and frozen immediately in liquid nitrogen, and total RNA was extracted with TRIzol (Invitrogen). Total RNA was treated with DNase I (Takara, Shanghai, China), and PrimeScript reverse transcriptase (Takara) was used for cDNA synthesis. Real-time PCR was performed in a 303 MyiQ2 two-color real-time PCR detection system (Bio-Rad, Richmond, CA, USA) [35]. At least three biological replicates per gene were carried out. Transcript levels were normalized relative to those of UBC21 cDNA [37]. PCR amplifications were performed in 20 mL reaction volumes containing SYBR Green PCR Master Mix (Applied Biosystems), cDNA, and the primers (listed in Supplementary Table S1). Quantification of relative expression levels followed a previously reported method [38].
2.4. MeJA and ABA Treatments
The B. napus accession K407 and transgenic lines were subjected to MeJA and ABA treatments. Initial ABA and MeJA concentrations were set according to [39,40]. Seeds were sown on solid MS plates containing MeJA (50 µM), ABA (9 µM), or both MeJA (50 µM) and ABA (9 µM) and treated at 4 °C for 4 d before moving to 22 °C for seed germination. Seed germination was recorded with images taken on the third day after the seed was grown at 22 °C. The emergence of radicle was scored as germination. Cotyledon opening and cotyledon expansion were also recorded [41,42,43]. The experiment was repeated using different batches of seeds, and at least 150 seeds were used per line. At least three biological replicates were carried out. For every biological replicate, we tested the seeds from the same batch at least three times as technical replicates.
2.5. Stress Treatment at Seedling Stage
For drought-stress treatment of seedlings, the seeds were sown in MS solid medium containing 400 mM mannitol, and then seeds were stratified at 4 °C in the dark for 4 d. The number of germinated seeds was recorded after time intervals of 1, 2, 3, 4, and 5 days. Cotyledon opening and cotyledon expansion were also recorded [41,42,43]. For determination of root length, the sterilized seeds were first sown on half-strength MS medium for 3 days, transplanted to fresh medium containing 400 mM mannitol and grown vertically. At least three biological replicates were carried out, and at least 150 seeds were used per line. For every biological replicate, we tested the seeds from the same batch at least three times as technical replicates.
For PEG treatment, the seedlings grown on 1/2 MS medium for 7 days were moved to fresh medium with 20% PEG-6000. Seedling samples were harvested after 0, 12, and 48 h according to the methods of Verslues et al. [44,45]. At least three biological replicates were carried out, and at least 150 seeds were used per line. For every biological replicate, we tested the seeds from the same batch at least three times as technical replicates
2.6. Dehydration Treatments
For seedling dehydration, the seedlings grown on half-strength MS medium for 7 days were transplanted to the surface of a Petri dish, maintained in a greenhouse at 22 °C under dark conditions, and then collected for an assay of dehydration effects after 0, 2, 6, and 12 h. At least three biological replicates were carried out, and at least 150 seeds were used per line.
For leaf dehydration, leaves were cut from the five-leaf-stage plants, maintaining the same dehydration conditions as seedlings. Leaves were harvested after time intervals of 0, 6, 12, 24, 36, 48, and 60 h [46]. At least three biological replicates were carried out, and at least 30 seeds were used per line.
For the whole plant dehydration treatment, soil was weighed to ensure that there was equal weight of soil in each pot and the same volume of water was poured into each pot. Seven-day-old seedlings were transplanted into the pots and grown until the five-leaf stage. Water was then withheld for 2 weeks. Plant survival rates were recorded based on the number of plants surviving 3 days after rewatering [47]. At least three biological replicates were carried out, and at least 30 seeds were used per line.
2.7. Measurement of Leaf Stomatal Density
Plants at the five-leaf stage were treated with drought stress (no watering for 7 days). Third leaves were collected, cut into leaf sections (0.5 cm in length), starting from the middle of a leaf, and fixed overnight in FAA at 25 °C. The leaf sections were rinsed three times using distilled water, dehydrated in ethanol series (30%, 50%, 70%, 80%, and 95%), and rinsed three times in 100% ethanol. The dehydrated samples were sputter-coated with gold, and stomatal observation was performed with a Hitachi S-2300 electron microscope. The number of stomata per mm2 in middle regions of leaves of five-leaf-stage plants was recorded [48]. At least three biological replicates were carried out, and at least 30 seeds were used per line.
2.8. Water Loss Assay
Leaves from plants at the five-leaf stage were detached and immediately weighed. The leaves were then placed on a laboratory bench and weighed according to the schedule (designated as Wi; Wi represents the weight of leaves at time i). Fresh weight (FW) loss was calculated according to the initial weight of the detached leaves. The water-loss rate (WLR) was also calculated: WLR = (FW − Wt)/FW. At least three biological replicates were carried out, and at least 20 seeds were used per line.
2.9. Measurement of Relative Water Content
Relative water content (RWC) was measured as described by Kumar et al. [49], with minor modification. Leaves were detached from plants at the five-leaf stage and immediately weighed to record their fresh weight (FW). The detached leaves were placed in distilled water for 12 h, blotted dry, and weighed to record their turgid weight (TW). To determine the dry weight (DW), the turgid leaves were subjected to oven drying at 70 °C for 36 h. RWC was calculated using the following equation: RWC = ((FW − DW) × 100)/(TW − DW). At least three biological replicates were carried out, and at least 20 seeds were used per line.
2.10. Sequence Alignment and Phylogenetic Analysis
Arabidopsis JAZ protein sequences were obtained from the Arabidopsis Information Resource (https://www.arabidopsis.org/ (accessed on 3 November 2021). BnA.JAZ5 and BnC.JAZ5 were selected based on their high similarity with AtJAZ5. Full-length amino-acid sequence multiple alignments were performed in ClustalW. Unrooted phylogenetic trees were constructed from the aligned amino-acid sequences in MEGA 6.0, and bootstrapping was carried out with 1000 iterations [50].
2.11. Statistical Analysis
Student’s t-tests were used for statistical analyses, and statistical significance was determined with p value < 0.05.
3. Results
3.1. Expression Patterns of JA Signaling Regulator BnA.JAZ5 Revealed Its Potential Function in Rapeseed Response to Drought Stress
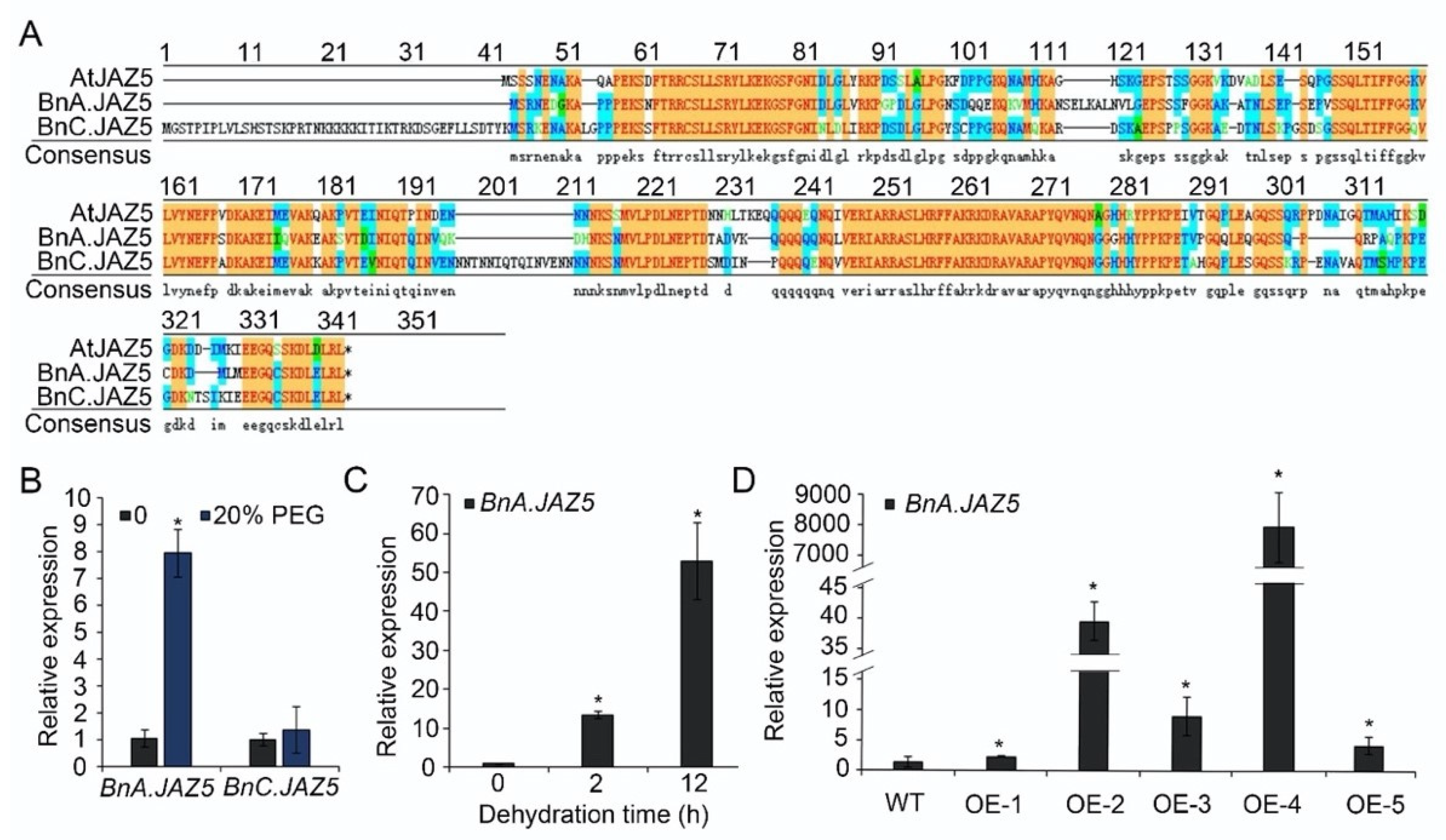
JAs are involved in plant development, reproduction, and defense. Within JA signaling cascades, JAZ proteins play a central role. BnA.JAZ5 and BnC.JAZ5 are two of the JAZ5 homoeologous genes present in polyploid rapeseed (Figure 1A and Figure S1). To investigate whether JAZ5 in rapeseed is involved in drought stress response, we checked the expression profiles of BnA.JAZ5 and BnC.JAZ5 in 18-day-old rapeseed treated with 20% high-molecular-weight polyethylene glycol (PEG-6000) for 48 h using real-time PCR. We found that BnA.JAZ5 was strongly induced by 20% PEG-6000, whereas BnC.JAZ5 was not (Figure 1B and Figure S4E). In addition, the expression level of BnA.JAZ5 was further increased under continued dehydration stress (Figure 1C and Figure S4E). Taken together, these results suggest that BnA.JAZ5 may be involved in plant drought response.
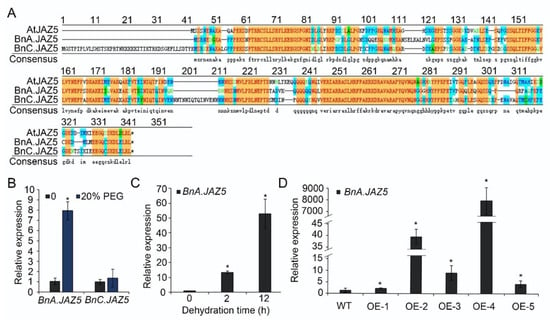
Figure 1.
Homologous alignment of AtJAZ5 in rapeseed and expression levels of BnA.JAZ5 and BnC.JAZ5. (A) Alignment of the protein sequences of AtJAZ5 in Arabidopsis and BnA.JAZ5 and BnC.JAZ5 in B. napus. (B) Expression levels of BnA.JAZ5 and BnC.JAZ5 under 20% PEG-6000. (C) Expression levels of BnA.JAZ5 under dehydration stress. Detached leaves of five-leaf-stage plants were subjected to dehydration treatment for 2 and 12 h. (D) The expression levels of BnA.JAZ5 in p35S::BnA.JAZ5 plants (OE-1, OE-2, OE-3, OE-4 and OE-5) and wild-type plants. Error bars indicate standard errors. Asterisks indicate significant differences (p < 0.05). WT, accession K407, OE-1, OE-2, OE-3, OE-4, OE-5, and p35S::BnA.JAZ5 lines.
3.2. Overexpression of BnA.JAZ5 Reduced Drought Tolerance
To study the functions of BnA.JAZ5 in response to drought, we constructed BnA.JAZ5-overexpressing transgenic plants by introducing a p35S::BnA.JAZ5 construct into the K407 cultivar of rapeseed (designated as p35S::BnA.JAZ5), which was self-fertilized and further selected to obtain homozygous lines for subsequent experiments. In total, five independent lines (OE-1, OE-2, OE-3 OE-4, and OE-5) were generated, and we found that BnA.JAZ5 was overexpressed from 3-fold to 7000-fold compared to the wild-type plants (Figure 1D).
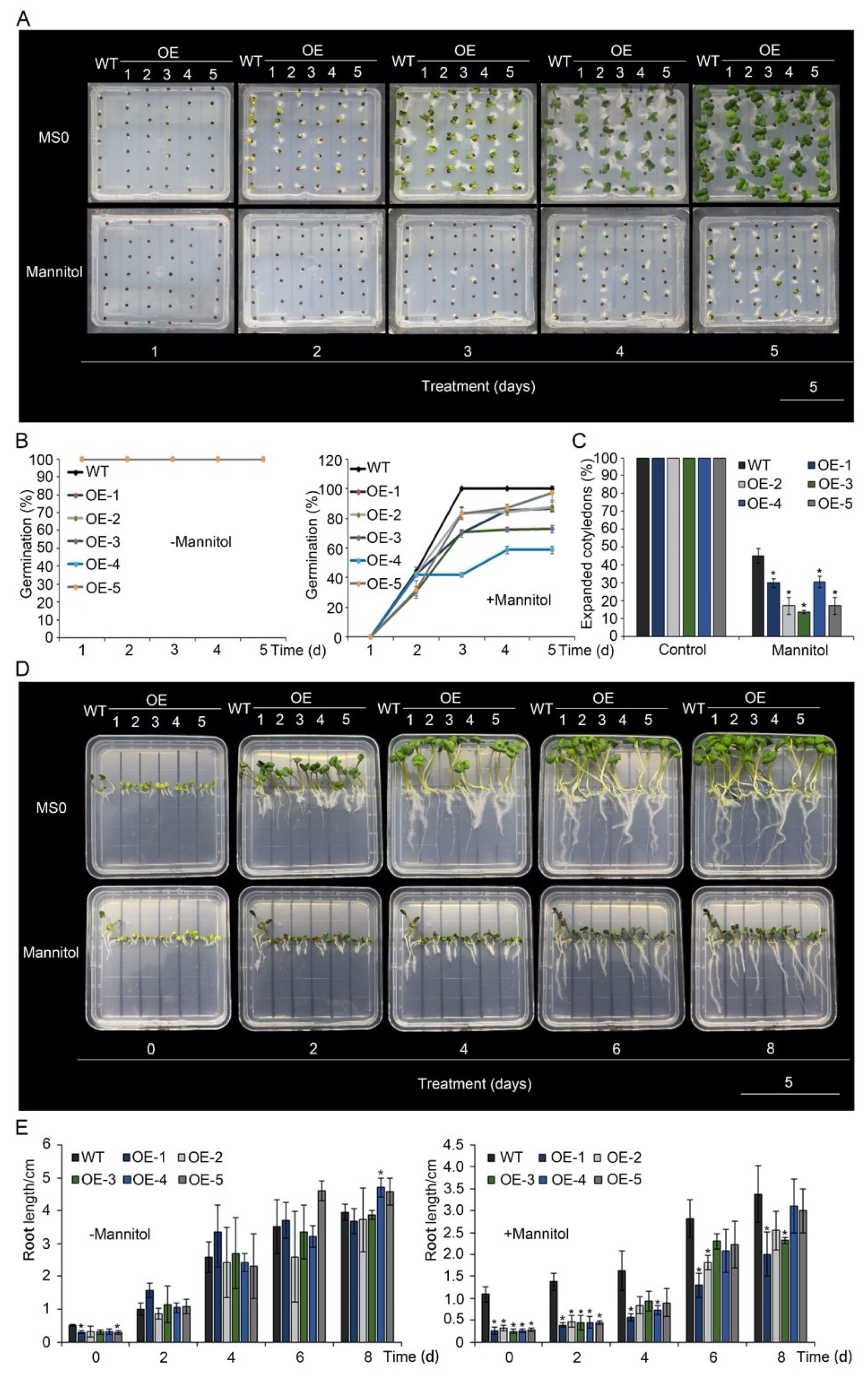
To study the tolerance of p35S::BnA.JAZ5-overexpressing plants to drought stress, seeds of the transgenic plants and K407 were sown on MS medium containing 400 mM mannitol (Figure 2). Compared with the control (0 mM mannitol), germination rates of all five of these transgenic lines were significantly reduced under 400 mM mannitol (Figure 2A,B and Figure S4A) and exhibited much lower percentages of expanded cotyledons on the fifth day after the seed was sown (Figure 2C). Root elongation of p35S::BnA.JAZ5 plants was inhibited compared with wild-type plants (Figure 2D,E). These results indicate that BnA.JAZ5 decreased drought tolerance of B. napus.
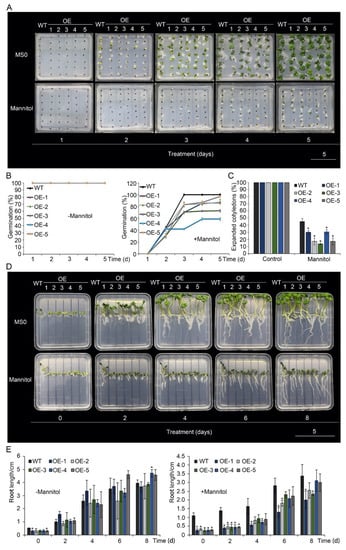
Figure 2.
Drought-stress responses of p35S::BnA.JAZ5 rapeseed plants. (A) Phenotypes of germinated six-day-old overexpressing lines under 400 mM mannitol treatment. (B) Seed germination rate of the five overexpressing lines under 400 mM mannitol treatment. (C) The percentage of wild-type and p35S::BnA.JAZ5 rapeseed seedlings with expanded cotyledons was scored 5 d after stratification on MS medium supplemented with 400 mM mannitol. (D) Phenotypes of root elongation of five overexpressing lines under 400 mM mannitol treatment. (E) Root length of five overexpressing lines under 400 mM mannitol treatment. Error bars represent standard errors. Asterisks indicate significant differences between wild-type and p35S::BnA.JAZ5 lines (p < 0.05).
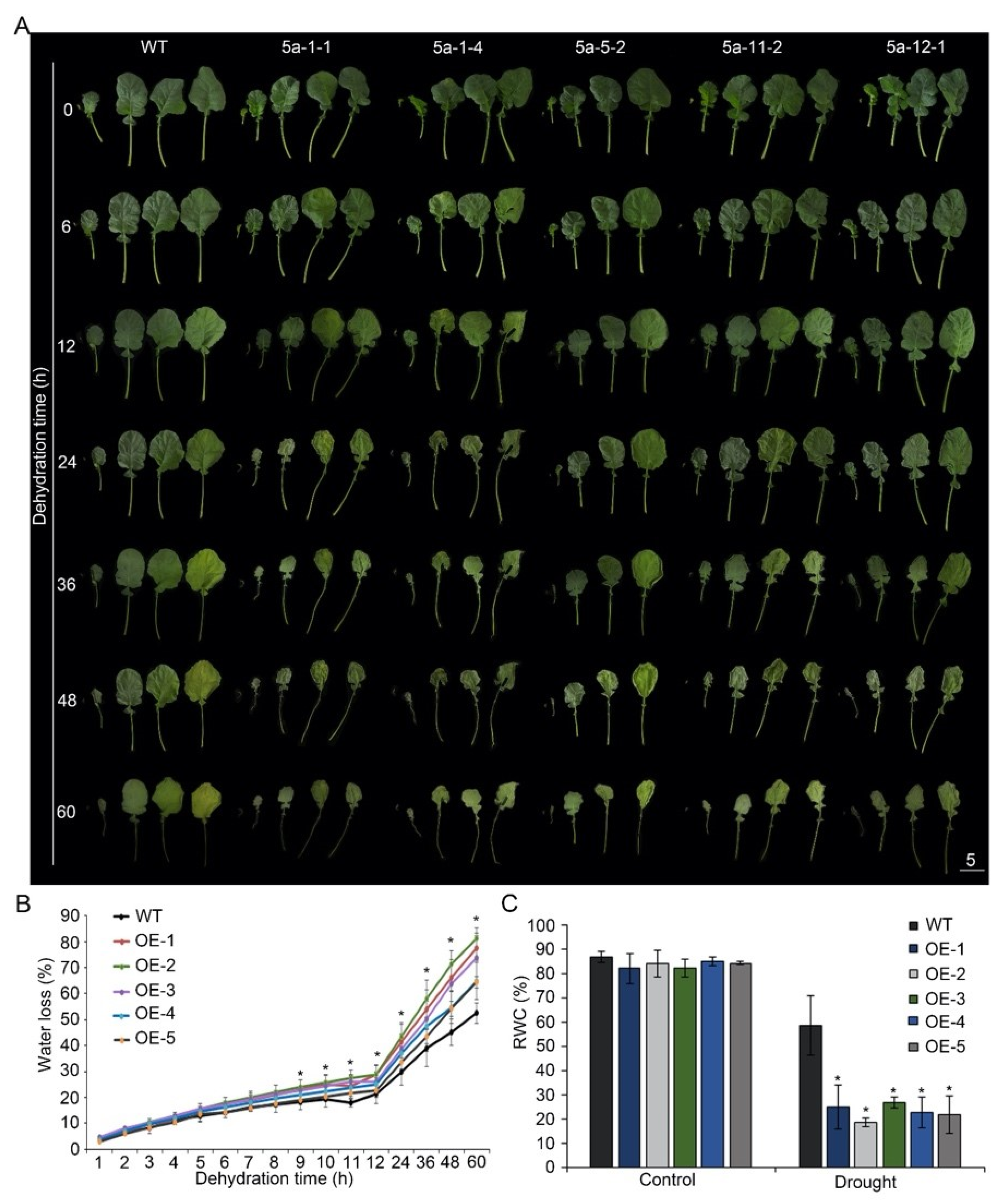
Compared to the wild-type plants, the leaves of p35S::BnA.JAZ5 plants rolled and wilted earlier (Figure 3A). Relative water content (RWC) and water-loss rate (WLR) are two indicators of cell and tissue hydration. Therefore, we tested the WLR and found the WLR of BnA.JAZ5-overexpressing rapeseed plants was significantly higher than that of wild-type (Figure 3B and Figure S4B). Besides, the RWC of BnA.JAZ5-overexpressing rapeseed plants was significantly lower than that of K407 under drought conditions (Figure 3C and Figure S4C). These results suggest that BnA.JAZ5 played a negative role in drought resistance.
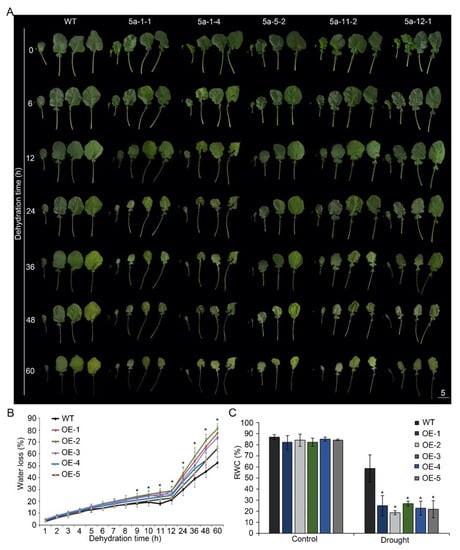
Figure 3.
Dehydration test of p35S::BnA.JAZ5 plants at the five-leaf stage. (A) Phenotypes of detached leaves of p35S::BnA.JAZ5 plants. (B) WLR of five overexpressing lines. Water loss was measured at the indicated time points. Asterisks indicate significant differences between wild-type and p35S::BnA.JAZ5 lines (p < 0.05, Student’s t-test). (C) RWC of five overexpressing lines under drought-stress treatment. Error bars represent standard errors. Asterisks indicate significant differences between control and drought stress (p < 0.05, Student’s t-test). WT, accession K407. OE-1, OE-2, OE-3, OE-4, OE-5, and p35S::BnA.JAZ5 lines. Scale bar, 5 cm.
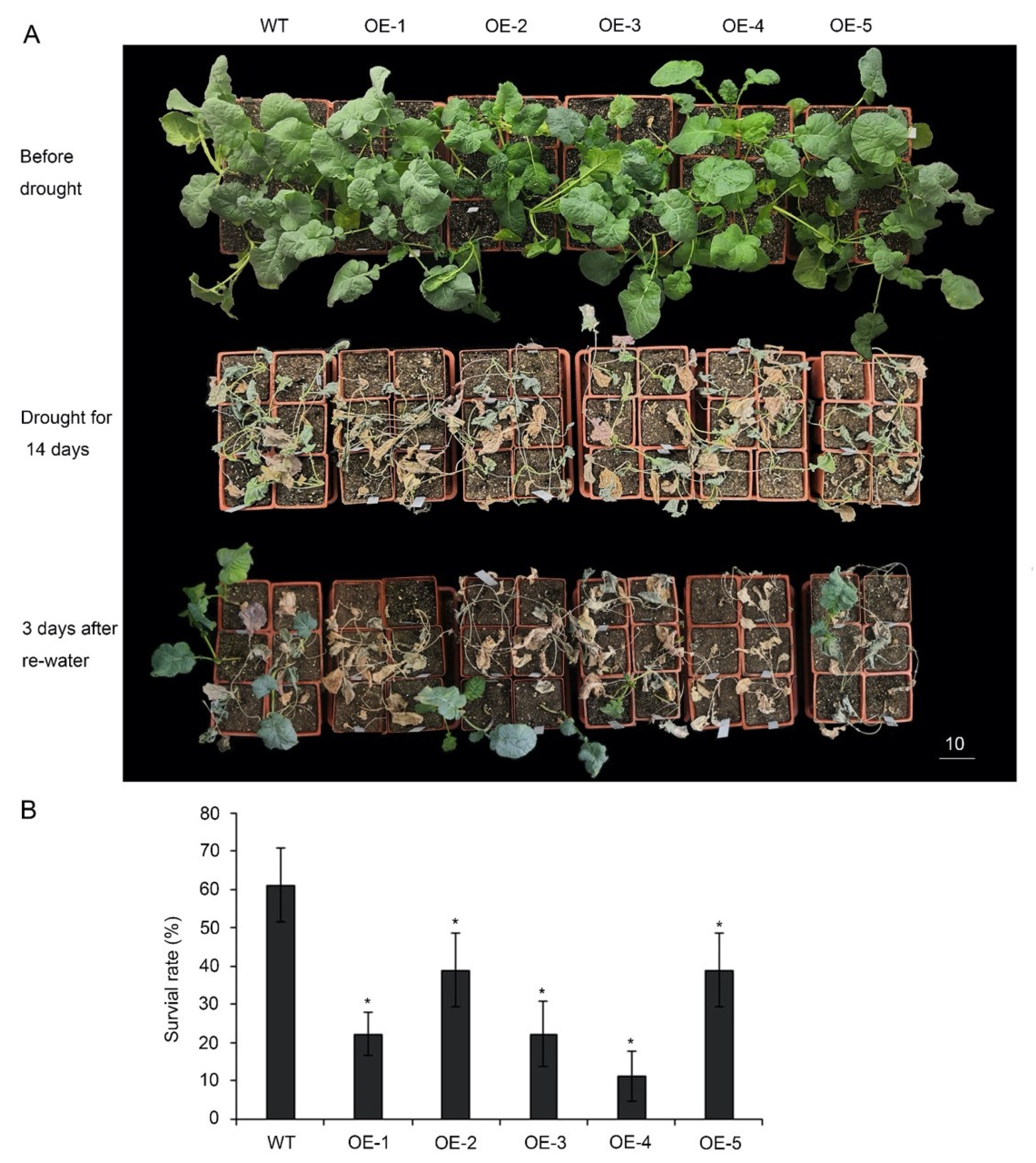
To further investigate the response of plants under drought conditions, five overexpressing lines were planted in the greenhouse for 5 weeks and then subjected to drought stress by withholding water for 14 days (Figure 4A). Drought-associated symptoms, such as leaf rolling and wilting, appeared earlier in p35S::BnA.JAZ5 plants than in the wild-type during drought treatment. After recovery for 3 days, only 22–40% of p35S::BnA.JAZ5 transgenic plants survived, when approximately 61% of wild-type ones survived (Figure 4B and Figure S4D). Overall, these results revealed that BnA.JAZ5 was a negative regulator of drought resistance.
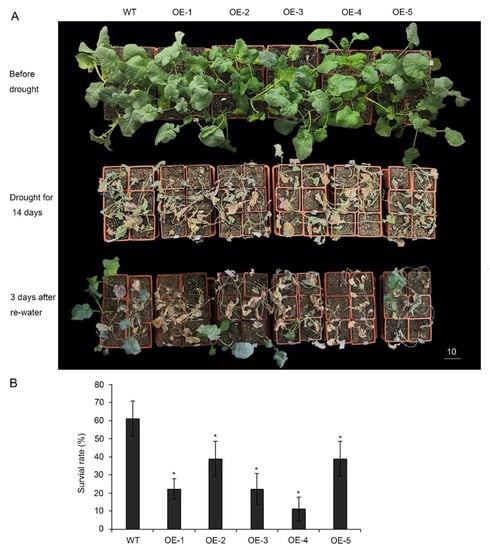
Figure 4.
Drought resistance testing of p35S::BnA.JAZ5 plants at the five-leaf stage. (A) Drought resistance assay of five overexpressing lines. (B) Survival rates of five overexpressing lines after 3-day recovery from drought stress treatment. WT, accession K407. OE-1, OE-2, OE-3, OE-4 and OE-5, p35S::BnA.JAZ5 plants. Asterisks indicate significant differences between wild-type and p35S::BnA.JAZ5 lines (p < 0.05, Student’s t-test). Scale bar, 10 cm.
3.3. BnA.JAZ5 Overexpression Altered Plant Responses to JA and ABA
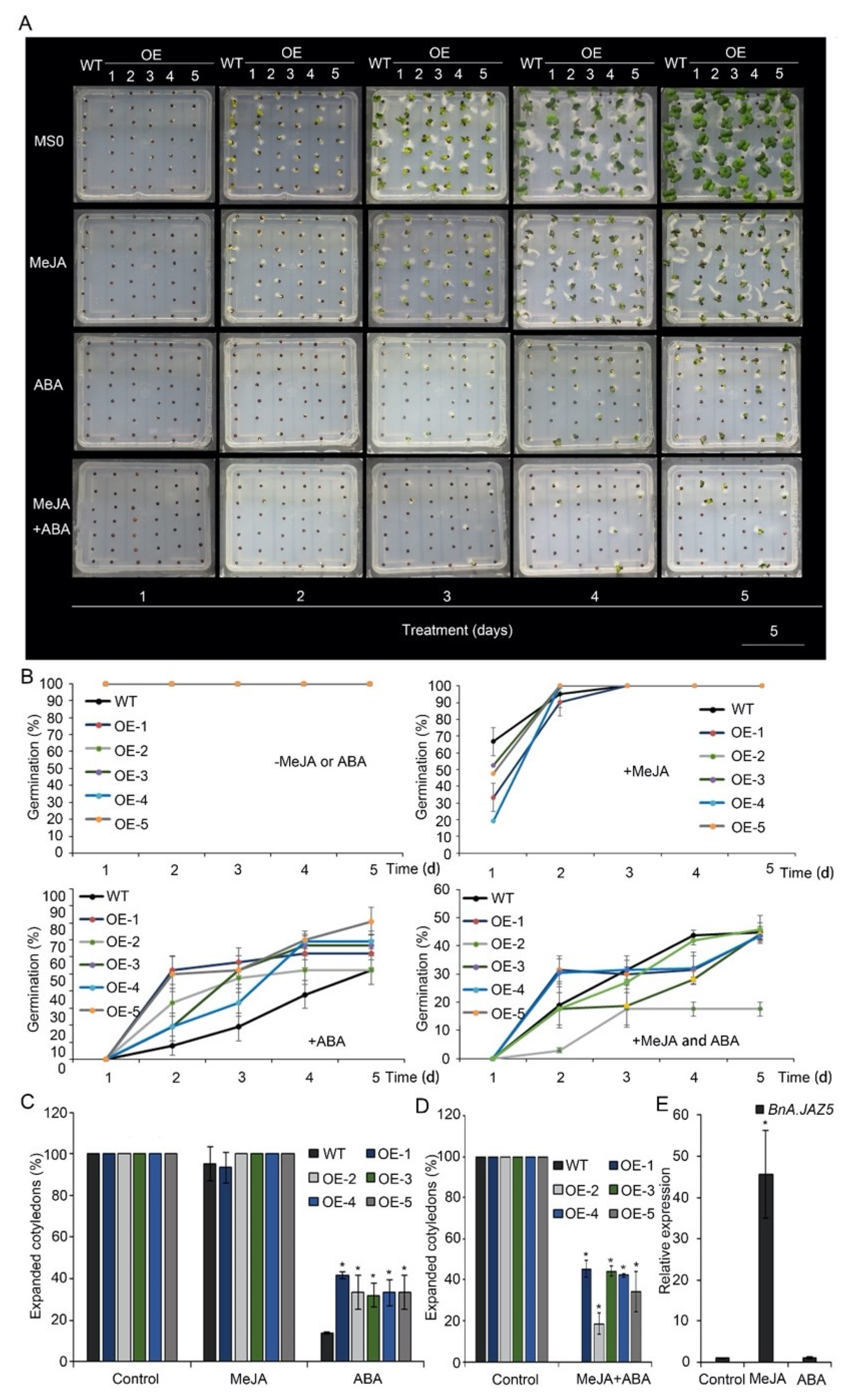
Given that JAZ5 belongs to the JAZ family, whose members are key inhibitors of the JA signaling pathway, we examined the function of JA on seed germination of BnA.JAZ5-overexpressing rapeseed. Overall, methyl jasmonate (MeJA) inhibited the seed germination of both wide-type and BnA.JAZ5-overexpressing rapeseed (Figure 5A–E). However, the germination rate of these five overexpressing lines showed no significant change under treatment with 50 µM MeJA (Figure 5A,B and Figure S4A), and the cotyledon expansion of the overexpressing lines was no significant difference on media containing 50 µM MeJA on the 4th day after seed was sown (Figure 5A,C). In fact, we found that, in wide-type plants, with 50 µM MeJA treatment, BnA.JAZ5 expression increased up to 46-fold as compared with the control (Figure 5E). These results suggested that BnA.JAZ5 overexpression did not affect the seed germination sensitivity to JA.
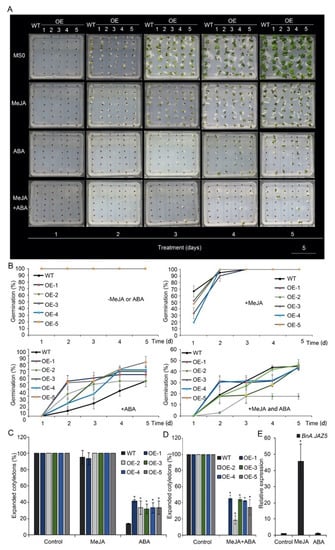
Figure 5.
Seed germination of p35S::BnA.JAZ5 plants on MS medium supplemented with 50 µM MeJA, 9 µM ABA or both 50 µM MeJA and 9 µM ABA, respectively. (A) Phenotypes of seed germination of five overexpressing lines under 50 µM MeJA, 9 µM ABA or both 50 µM MeJA and 9 µM ABA treatments. (B) Seed germination rate of five overexpressing lines under 50 µM MeJA, 9 µM ABA or both 50 µM MeJA and 9 µM ABA treatments. (C) The percentage of wild-type and p35S::BnA.JAZ5 rapeseed seedlings with expanded cotyledons was scored 4 d after stratification on MS medium supplemented with 50 µM MeJA or 9 µM ABA. (D) The percentage of wild-type and p35S::BnA.JAZ5 rapeseed seedlings with expanded cotyledons was scored 9 d after stratification on MS medium supplemented with both 50 µM MeJA and 9 µM ABA. (E) Expression levels of BnA.JAZ5 under 50 µM MeJA or 9 µM ABA treatments. Error bars represent standard errors. Asterisks indicate significant differences between wild-type and p35S::BnA.JAZ5 lines (p < 0.05). WT, accession K407. OE-1, OE-2, OE-3, OE-4 and OE-5, p35S::BnA.JAZ5 lines. Scale bar, 5 cm.
In rapeseed seeds, exogenous application of ABA inhibits germination [51]. To test the possible roles of BnA.JAZ5 in ABA signaling, we treated the wild-type and five overexpressing lines with 9 µM ABA, as suggested by previous studies [52,53]. From this analysis, we found the seed germination of five overexpressing lines exhibited hyposensitivity to ABA as compared to wild-type plants (Figure 5A,B and Figure S4A). Additionally, compared with the wild-type line, the five overexpressing lines exhibited much higher percentages of expanded cotyledons on media containing 9 µM ABA on the 5th day after the seed was sown (Figure 5A,C). In addition, we found no change in the expression levels of BnA.JAZ5 as compared to the control with 9 µM ABA treatment (Figure 5E). These findings indicate that BnA.JAZ5 may regulate drought response in an ABA-dependent manner.
Next, to test the JAZ5 function in JA–ABA crosstalk, we treated the plants with a combination of 50 µM MeJA and 9 µM ABA. As expected, the seed germination of wild-type and p35S::BnA.JAZ5-overexpressing plants was severely inhibited (Figure 5A,B,D and Figure S4A). Interestingly, while the seed germination rate of p35S::BnA.JAZ5-overexpressing plants was higher as compared to wild-type plants under ABA treatment, the seed germination rate showed no difference with the combination of ABA and JA treatment, suggesting that JAZ5 plays a role in crosstalk between JA and ABA. Taken together, these results suggest that BnA.JAZ5 regulated rapeseed plant responses to drought stress through ABA and JA signaling.
3.4. BnA.JAZ5 Regulates ABA-Dependent Stress-Responsive Genes
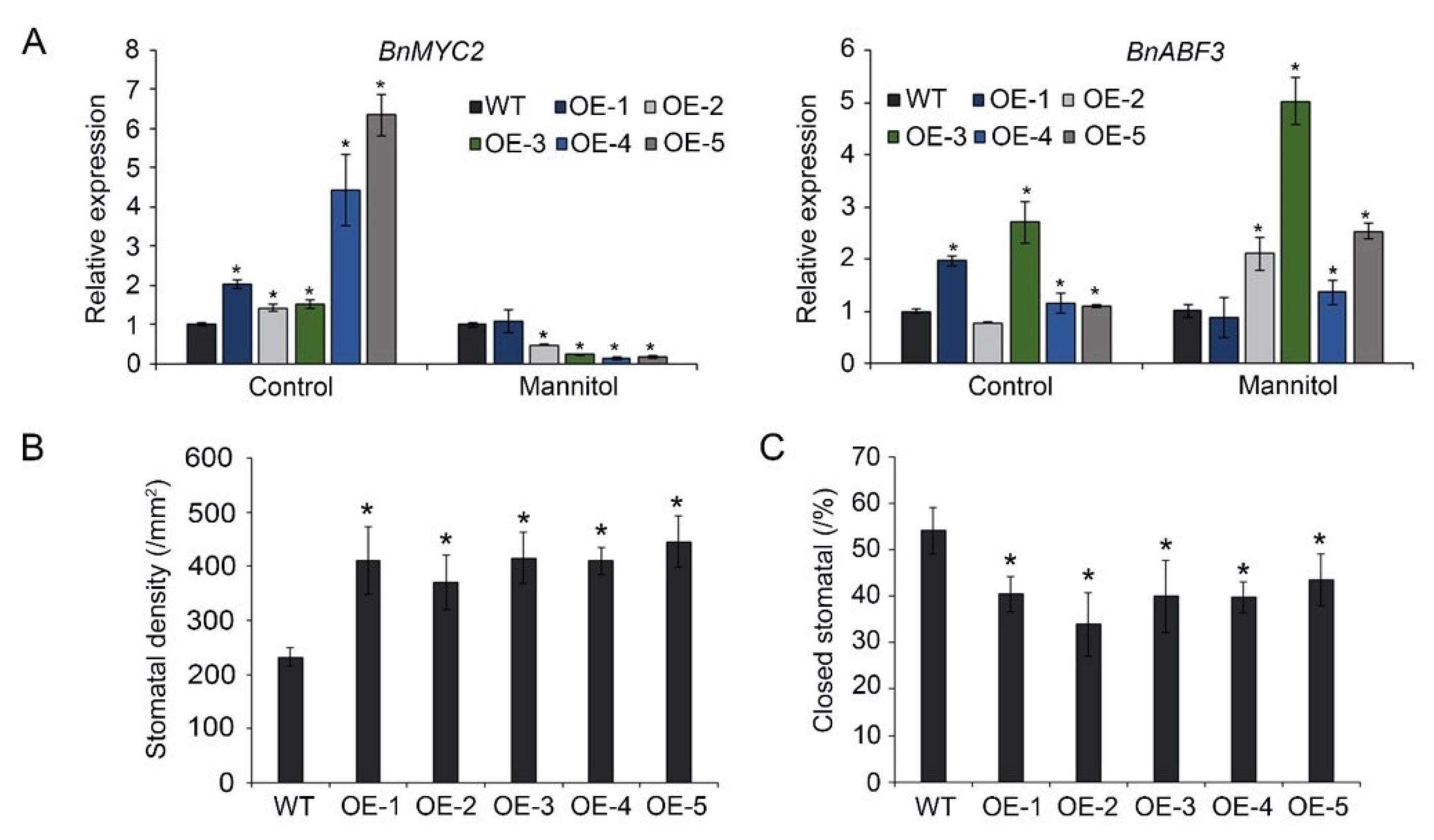
To explore the role of BnA.JAZ5 in drought stress through the ABA signaling pathway, we examined JA- and ABA-responsive marker genes in plants under 400 mM mannitol treatment (Figure 6A). Our real-time PCR results revealed that BnA.JAZ5 overexpression led to downregulation of the JA-responsive gene BnMYC2 and upregulation of the ABA-responsive gene BnABF3 under 400 mM mannitol treatment (Figure 6A). In addition, we examined stress-related marker genes in plants under dehydration treatment (Figure S2A). BnA.JAZ5 overexpression led to downregulation of the stress-related genes BnP5CS, BnCTSTM1, and BnLEA76 (Figure S2B), and the expression of BnPLC was repressed in BnA.JAZ5-overexpressing lines after 6 h and 12 h drought treatment as compared to wide-types. This finding suggests that BnA.JAZ5 attenuated the drought tolerance of the transgenic plants through downregulation of ABA-dependent stress-responsive genes.
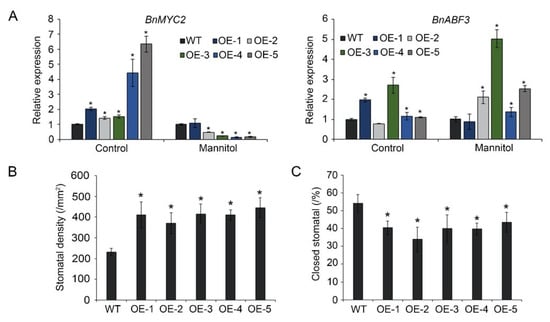
Figure 6.
Changes in stomatal density and expression levels of stress-related marker genes. (A) Expression patterns of JA- and ABA-responsive genes under 400 mM mannitol stress. Error bars represent standard errors. Asterisks indicate significant differences (p < 0.05). (B) Stomatal density of OE-1, OE-2, OE-3, OE-4, and OE-5 lines. (C) Percentage of closed stomata under drought stress. Asterisks indicate significant differences between WT and p35S::BnA.JAZ5 lines (p < 0.05, Student’s t-test). WT, accession K407; OE-1, OE-2, OE-3, OE-4, OE-5, and p35S::BnA.JAZ5 transgenic lines.
3.5. BnA.JAZ5 Overexpression Increased Stomatal Density and Reduced Stomatal Closure
Guard-cell signaling plays a critical role in plant drought response, which is regulated by ABA [1]. We measured stomatal density under normal and drought-stress conditions. Under normal conditions, the average stomatal density of five overexpressing lines was higher than that of wild-type plants (Figure 6B and Figure S3). After drought stress, the percentage of closed stomata of five overexpressing lines was lower than that of the wild-type plants (Figure 6C and Figure S3). These results suggest that BnA.JAZ5 overexpression increased stomatal density under normal conditions and reduced stomatal closure when transgenic plants were subjected to drought stress.
4. Discussion
Rapeseed (B. napus), soybean, and oil palm are the most extensively grown oil crop species worldwide. Rapeseed, however, is very sensitive to water stress, a main factor of crop failure in rapeseed [5]. Identifying genes related to dehydration stress or markers linked to these genes is a key step in genomics-assisted breeding in rapeseed [54]. JAZ proteins are key components of the JA signaling pathway. The OsJAZ genes were induced upon abiotic stress in OsbHLH148-overexpressing plants [55]. OsJAZ9 can increase salt and cold tolerance in rice by inhibiting the expression of OsbHLH062 and OsMYB30 [56,57]. GsJAZ2 overexpression in Arabidopsis reduces plant sensitivity to salt stress [58]. In rice, interaction of OsJAZ proteins with a basic helix–loop–helix protein leads to drought tolerance [55,56,57], whereas OsJAZ1 negatively regulates drought resistance by modulating JA and ABA signaling [52]. The varied roles of JAZ members indicate that JAZ protein family members regulate abiotic stresses differentially. However, little is known about JAZ functional roles and their mechanisms in rapeseed. We collected cotyledon, first true leaf, root (five-leaf stage), third leaf (five-leaf stage), bud, flower, and young silique samples from the rapeseed accession K407 for real-time PCR analysis. We found that BnA.JAZ5 was generally expressed in all these tissues of rapeseed plants (Figure S2C). Furthermore, both 20% PEG-6000 treatment and dehydration stress significantly induced the expression of BnA.JAZ5. The germination rate of p35S::BnA.JAZ5 transgenic seeds was weaker than that of wild-type seeds on medium containing 400 M mannitol. Fewer p35S::BnA.JAZ5 plants survived than wild-type plants following withholding of water for 14 days and recovery for 3 days. Thus, we thought that BnA.JAZ5 may have a negative role in drought resistance, and this conclusion is in agreement with those concerning OsJAZ1 [52].
Most plant transpiration occurs in stomata, and stomatal density and/or movement can adjust the transpiration rate [59,60]. Leaves can respond to water status by changing stomata density and guard-cell size under water stress [61]. We found that stomatal density was significantly increased in p35S::BnA.JAZ5 plants, and a lower proportion of closed stomata was observed in p35S::BnA.JAZ5 plants under drought stress. This change caused the p35S::BnA.JAZ5 plants to lose water more rapidly under drought stress, thereby accelerating death and reducing drought resistance. Consistently, p35S::BnA.JAZ5 plants had lower RWC under drought stress and higher WLR than wild-type plants. Stomatal closure is one of the ABA-regulated pathways activated by water-deficit conditions [62,63]. A smaller stomatal aperture and stomatal density contribute to reduced water loss from plant cells, thereby enhancing drought or osmotic stress tolerance [8,9,10,64]. BnA.JAZ5 gene did not respond to exogenous ABA treatment, but p35S::BnA.JAZ5 plants were ABA-hyposensitive, and the germination rate of p35S::BnA.JAZ5 seeds on MS medium containing ABA was higher than that of wild-type seeds. Studies have shown that JAZ proteins interact with ABA-responsive transcription factors in Arabidopsis [15,41]. The expression of JA-responsive gene BnMYC2 was downregulated, and the expression of ABA-responsive gene BnABF3 was upregulated in p35S::BnA.JAZ5 plants under 400 mM mannitol treatment. At the same time, BnA.JAZ5 could be induced by MeJA. In addition, the proline synthesis gene BnP5CS and stress tolerance-related genes BnRD29A, BnLEA, and BnCYSTM1 tend to be downregulated in p35S::BnA.JAZ5 plants under drought-stress conditions. These results suggest that BnA.JAZ5 negatively regulates drought resistance in rapeseed through JA–ABA crosstalk, and this process affects stomatal development. BnA.JAZ5 can be used as a candidate gene for improving drought resistance of rapeseed. We can design sgRNA to edit these BnA.JAZ5 homologs using the CRISPR/Cas9 system to obtain rape germplasm with higher drought resistance.
5. Conclusions
In this study, the function of rapeseed BnA.JAZ5 in drought response was characterized. BnA.JAZ5-overexpressing rapeseed plants were further investigated under drought conditions and plant hormone (JA and ABA) treatment, revealing the role of BnA.JAZ5 in attenuating rapeseed drought resistance in an ABA-dependent manner. Molecular study showed that BnA.JAZ5 regulated ABA-mediated stress-responsive genes MYC2 and ABF3. Finally, significantly increased stomatal density and reduced stomatal closure were observed in BnA.JAZ5-overexpressing rapeseed plants. Taken together, these results suggest that BnA.JAZ5-mediated crosstalk between JA and ABA signaling pathways contributed to the rapeseed response to drought stress. BnA.JAZ5 could be used as CRISPR-editing genetic loci for drought-resistant rapeseed breeding.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/horticulturae8020131/s1, Table S1. List of primers used in this study. Figure S1. Phylogeny tree of rapeseed BnA.JAZ5 (GSBRNA2T00103000001) and BnC.JAZ5 (GSBRNA2T00089260001) with Arabidopsis JAZ genes. Figure S2. Changes in plant phenotype and gene expression under dehydration stress. Figure S3. SEM images showing adaxial epidermal cells of OE-1, OE-2, OE-3, OE-4, and OE-5 lines at 350× magnification. Figure S4. Molecular data of another two biological replicates per experiment.
Author Contributions
Y.H. and S.H. designed the project. B.C., X.W., J.B. and Y.Z. performed the experiments. B.C., X.Y., S.H. and Y.H. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Key Research and Development Program of China (grant Nos. 2016YFD0101900 and 2016YFD0100500) and the Natural Science Foundation of China (grant Nos. 31771442 and 31571261).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Gene sequences generated in this study were uploaded to NCBI (AtJAZ5: MW233040; BnA8.JAZ5: MW233041; BnC8.JAZ5: MW233042; BnP5CS: MW233043; BnPLC: MW233044; BnCYSTM1: MW233045; and BnLEA76: MW233046).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook, E.R.; Seager, R.; Cane, M.A.; Stahle, D.W. North American drought: Reconstructions, causes, and consequences. Earth-Sci. Rev. 2007, 81, 93–134. [Google Scholar] [CrossRef]
- Tester, M.; Langridge, P. Breeding technologies to increase crop production in a changing world. Science 2010, 327, 818–822. [Google Scholar] [CrossRef] [PubMed]
- Allender, C.J.; King, G.J. Origins of the amphiploid species Brassica napus L. investigated by chloroplast and nuclear molecular markers. BMC Plant Biol. 2010, 10, 54. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.X.; Griffiths, R.; Ying, J.F.; McCourt, P.; Huang, Y.F. Development of drought-tolerant Canola (Brassica napus L.) through genetic modulation of ABA-mediated stomatal responses. Crop Sci. 2009, 49, 1539–1554. [Google Scholar] [CrossRef]
- Jeong, J.S.; Kim, Y.S.; Baek, K.H.; Jung, H.; Ha, S.H.; Do Choi, Y.; Kim, M.; Reuzeau, C.; Kim, J.K. Root-specific expression of OsNAC10 improves drought tolerance and grain yield in rice under field drought conditions. Plant Physiol. 2010, 153, 185–197. [Google Scholar] [CrossRef] [Green Version]
- Yoo, C.Y.; Pence, H.E.; Jin, J.B.; Miura, K.; Gosney, M.J.; Hasegawa, P.M.; Mickelbart, M.V. The Arabidopsis GTL1 transcription factor regulates water use efficiency and drought tolerance by modulating stomatal density via transrepression of SDD1. Plant Cell 2010, 22, 4128–4141. [Google Scholar] [CrossRef] [Green Version]
- Masle, J.; Gilmore, S.R.; Farquhar, G.D. The ERECTA gene regulates plant transpiration efficiency in Arabidopsis. Nature 2005, 436, 866–870. [Google Scholar] [CrossRef]
- Berger, D.; Altmann, T. A subtilisin-like serine protease involved in the regulation of stomatal density and distribution in Arabidopsis thaliana. Genes Dev. 2000, 14, 1119–1131. [Google Scholar] [CrossRef]
- Shpak, E.D.; McAbee, J.M.; Pillitteri, L.J.; Torii, K.U. Stomatal patterning and differentiation by synergistic interactions of receptor kinases. Science 2005, 309, 290–293. [Google Scholar] [CrossRef]
- Ouyang, S.Q.; Liu, Y.F.; Liu, P.; Lei, G.; He, S.J.; Ma, B.; Zhang, W.K.; Zhang, J.S.; Chen, S.Y. Receptor-like kinase OsSIK1 improves drought and salt stress tolerance in rice (Oryza sativa) plants. Plant J. 2010, 62, 316–329. [Google Scholar] [CrossRef]
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Zhu, J.K. Molecular and genetic aspects of plant responses to osmotic stress. Plant Cell Environ. 2002, 25, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, T.; Mogami, J.; Yamaguchi-Shinozaki, K. ABA-dependent and ABA-independent signaling in response to osmotic stress in plants. Curr. Opin. Plant Biol. 2014, 21, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Characterization of the expression of a desiccation-responsive rd29 gene of Arabidopsis thaliana and analysis of its promoter in transgenic plants. Mol. Gen. Genet. 1993, 236, 331–340. [Google Scholar] [CrossRef]
- Kishor, P.; Hong, Z.; Miao, G.H.; Hu, C.; Verma, D. Overexpression of Δ1-Pyrroline-5-Carboxylate Synthetase lncreases Proline Production and Confers Osmotolerance in Transgenic Plants. Plant Physiol. 1995, 108, 1387–1394. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; van Wijk, R.; Zarza, X.; Shahbaz, M.; van Hooren, M.; Guardia, A.; Scuffi, D.; Garcia-Mata, C.; Van den Ende, W.; Hoffmann-Benning, S.; et al. Knock-down of Arabidopsis PLC5 reduces primary root growth and secondary root formation while overexpression improves drought tolerance and causes stunted root hair growth. Plant Cell Physiol. 2018, 59, 2004–2019. [Google Scholar] [CrossRef]
- Devoto, A.; Turner, J.G. Jasmonate-regulated Arabidopsis stress signalling network. Physiol. Plant 2005, 123, 161–172. [Google Scholar] [CrossRef]
- Thines, B.; Katsir, L.; Melotto, M.; Niu, Y.; Mandaokar, A.; Liu, G.; Nomura, K.; He, S.Y.; Howe, G.A.; Browse, J. JAZ repressor proteins are targets of the SCFCOI1 complex during jasmonate signalling. Nature 2007, 448, 661–665. [Google Scholar] [CrossRef]
- Fernandez-Calvo, P.; Chini, A.; Fernandez-Barbero, G.; Chico, J.M.; Gimenez-Ibanez, S.; Geerinck, J.; Eeckhout, D.; Schweizer, F.; Godoy, M.; Franco-Zorrilla, J.M.; et al. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell 2011, 23, 701–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo, O.; Chico, J.M.; Sanchez-Serrano, J.J.; Solano, R. JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 2004, 16, 1938–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, Y.; Figueroa, P.; Browse, J. Characterization of JAZ-interacting bHLH transcription factors that regulate jasmonate responses in Arabidopsis. J. Exp. Bot. 2011, 62, 2143–2154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, C.; Li, L.; Zhai, Q.; You, Y.; Deng, L.; Wu, F.; Chen, R.; Jiang, H.; Wang, H.; Chen, Q.; et al. Mediator subunit MED25 links the jasmonate receptor to transcriptionally active chromatin. Proc. Natl. Acad. Sci. USA 2017, 114, E8930–E8939. [Google Scholar] [CrossRef] [Green Version]
- Pauwels, L.; Barbero, G.F.; Geerinck, J.; Tilleman, S.; Grunewald, W.; Perez, A.C.; Chico, J.M.; Bossche, R.V.; Sewell, J.; Gil, E.; et al. NINJA connects the co-repressor TOPLESS to jasmonate signalling. Nature 2010, 464, 788–791. [Google Scholar] [CrossRef] [Green Version]
- Shyu, C.; Figueroa, P.; Depew, C.L.; Cooke, T.F.; Sheard, L.B.; Moreno, J.E.; Katsir, L.; Zheng, N.; Browse, J.; Howe, G.A. JAZ8 lacks a canonical degron and has an EAR motif that mediates transcriptional repression of jasmonate responses in Arabidopsis. Plant Cell 2012, 24, 536–550. [Google Scholar] [CrossRef] [Green Version]
- Katsir, L.; Schilmiller, A.L.; Staswick, P.E.; He, S.Y.; Howe, G.A. COI1 is a critical component of a receptor for jasmonate and the bacterial virulence factor coronatine. Proc. Natl. Acad. Sci. USA 2008, 105, 7100–7105. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Yao, R.; Chen, L.; Li, S.; Gu, M.; Nan, F.; Xie, D. Dynamic perception of jasmonates by the F-Box protein COI1. Mol. Plant 2018, 11, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Qi, T.; Song, S.; Ren, Q.; Wu, D.; Huang, H.; Chen, Y.; Fan, M.; Peng, W.; Ren, C.; Xie, D. The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 2011, 23, 1795–1814. [Google Scholar] [CrossRef] [Green Version]
- Chini, A.; Fonseca, S.; Fernandez, G.; Adie, B.; Chico, J.M.; Lorenzo, O.; Garcia-Casado, G.; Lopez-Vidriero, I.; Lozano, F.M.; Ponce, M.R.; et al. The JAZ family of repressors is the missing link in jasmonate signalling. Nature 2007, 448, 666–671. [Google Scholar] [CrossRef]
- Song, S.; Qi, T.; Huang, H.; Ren, Q.; Wu, D.; Chang, C.; Peng, W.; Liu, Y.; Peng, J.; Xie, D. The Jasmonate-ZIM domain proteins interact with the R2R3-MYB transcription factors MYB21 and MYB24 to affect Jasmonate-regulated stamen development in Arabidopsis. Plant Cell 2011, 23, 1000–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, X.; Zhang, W.; Yu, F.; Tian, J.; Li, D.; Guo, A. Functional analysis of the two Brassica AP3 genes involved in apetalous and stamen carpelloid phenotypes. PLoS ONE 2011, 6, e20930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, X.; Yang, J.; He, Y. Natural antisense transcripts of MIR398 genes suppress microR398 processing and attenuate plant thermotolerance. Nat. Commun. 2020, 11, 5351. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Wu, F.; Bai, J.; Li, X.; Yang, X.; Xue, W.; Liu, H.; He, Y. BcpLH organizes a specific subset of microRNAs to form a leafy head in Chinese cabbage (Brassica rapa ssp. pekinensis). Hortic. Res. 2020, 7, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, S.S.; Yajima, W.R.; Rahman, M.H.; Shah, S.; Liu, J.J.; Ekramoddoullah, A.K.; Kav, N.N. A cysteine-rich antimicrobial peptide from Pinus monticola (PmAMP1) confers resistance to multiple fungal pathogens in canola (Brassica napus). Plant Mol. Biol. 2012, 79, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Moloney, M.M.; Walker, J.M.; Sharma, K.K. High-efficiency transformation of Brassica napus using Agrobacterium vectors. Plant Cell Rep. 1989, 8, 238–242. [Google Scholar] [CrossRef]
- Lv, J.; Huang, Q.; Sun, Y.; Qu, G.; Guo, Y.; Zhang, X.; Zhao, H.; Hu, S. Male sterility of an AHAS-mutant induced by tribenuron-methyl solution correlated with the decrease of AHAS activity in Brassica napus L. Front. Plant Sci. 2018, 9, 1014. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhu, Z.; Sun, B.; Xu, X.; Chen, H.; Zou, L.; Chen, G.; Cao, B.; Chen, C.; Lei, J. Overexpression of AtEDT1/HDG11 in Chinese Kale (Brassica oleracea var. alboglabra) enhances drought and osmotic stress tolerance. Front. Plant Sci. 2016, 7, 1285. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Xia, Z.; Cai, Z.; Li, L.; Cheng, Y.; Liu, J.; Nian, H. GmWRKY16 enhances drought and salt tolerance through an ABA-mediated pathway in Arabidopsis thaliana. Front. Plant Sci. 2018, 9, 1979. [Google Scholar] [CrossRef] [Green Version]
- Pan, J.; Hu, Y.; Wang, H.; Guo, Q.; Chen, Y.; Howe, G.A.; Yu, D. Molecular mechanism underlying the synergetic effect of jasmonate on abscisic acid signaling during seed germination in Arabidopsis. Plant Cell 2020, 32, 3846–3865. [Google Scholar] [CrossRef] [PubMed]
- Dave, A.; Hernandez, M.L.; He, Z.; Andriotis, V.M.; Vaistij, F.E.; Larson, T.R.; Graham, I.A. 12-oxo-phytodienoic acid accumulation during seed development represses seed germination in Arabidopsis. Plant Cell 2011, 23, 583–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piskurewicz, U.; Lopez-Molina, L. Basic techniques to assess seed germination responses to abiotic stress in Arabidopsis thaliana. Methods Mol. Biol. 2016, 1398, 183–196. [Google Scholar]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Van der Weele, C.M.; Spollen, W.G.; Sharp, R.E.; Baskin, T.I. Growth of Arabidopsis thaliana seedlings under water deficit studied by control of water potential in nutrient-agar media. J. Exp. Bot. 2000, 51, 1555–1562. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Suzuki, T.; Takahashi, F.; Shinozaki, K.; Yamaguchi-Shinozaki, K. NF-YB2 and NF-YB3 have functionally diverged and differentially induce drought and heat stress-specific genes. Plant Physiol. 2019, 180, 1677–1690. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Nolan, T.M.; Ye, H.; Zhang, M.; Tong, H.; Xin, P.; Chu, J.; Chu, C.; Li, Z.; Yin, Y. Arabidopsis WRKY46, WRKY54, and WRKY70 transcription factors are involved in Brassinosteroid-regulated plant growth and drought responses. Plant Cell 2017, 29, 1425–1439. [Google Scholar] [CrossRef]
- Cao, W.; Cao, B.; Wang, X.; Bai, J.; Xu, Y.Z.; Zhao, J.; Li, X.; He, Y.; Hu, S. Alternatively spliced BobCAL transcripts alter curd morphotypes in a collection of Chinese cauliflower accessions. Hortic. Res. 2020, 7, 160. [Google Scholar] [CrossRef]
- Kumar, D.; Yusuf, M.A.; Singh, P.; Sardar, M.; Sarin, N.B. Modulation of antioxidant machinery in α-tocopherol-enriched transgenic Brassica juncea plants tolerant to abiotic stress conditions. Protoplasma 2013, 250, 1079–1089. [Google Scholar] [CrossRef]
- Danisman, S.; van Dijk, A.D.; Bimbo, A.; van der Wal, F.; Hennig, L.; de Folter, S.; Angenent, G.C.; Immink, R.G. Analysis of functional redundancies within the Arabidopsis TCP transcription factor family. J. Exp. Bot. 2013, 64, 5673–5685. [Google Scholar] [CrossRef] [Green Version]
- Schopfer, P.; Plachy, C. Control of seed germination by abscisic acid. II. Effect on embryo water uptake in Brassica napus L. Plant Physiol. 1984, 76, 155–160. [Google Scholar] [CrossRef] [Green Version]
- Fu, J.; Wu, H.; Ma, S.; Xiang, D.; Liu, R.; Xiong, L. OsJAZ1 attenuates drought resistance by regulating JA and ABA signaling in rice. Front. Plant Sci. 2017, 8, 2108. [Google Scholar] [CrossRef] [Green Version]
- Ju, L.; Jing, Y.; Shi, P.; Liu, J.; Chen, J.; Yan, J.; Chu, J.; Chen, K.M.; Sun, J. JAZ proteins modulate seed germination through interaction with ABI5 in bread wheat and Arabidopsis. New Phytol. 2019, 223, 246–260. [Google Scholar] [CrossRef]
- Xue, Y.; Warburton, M.L.; Sawkins, M.; Zhang, X.; Setter, T.; Xu, Y.; Grudloyma, P.; Gethi, J.; Ribaut, J.M.; Li, W.; et al. Genome-wide association analysis for nine agronomic traits in maize under well-watered and water-stressed conditions. Theor. Appl. Genet. 2013, 126, 2587–2596. [Google Scholar] [CrossRef]
- Seo, J.S.; Joo, J.; Kim, M.J.; Kim, Y.K.; Nahm, B.H.; Song, S.I.; Cheong, J.J.; Lee, J.S.; Kim, J.K.; Choi, Y.D. OsbHLH148, a basic helix-loop-helix protein, interacts with OsJAZ proteins in a jasmonate signaling pathway leading to drought tolerance in rice. Plant J. 2011, 65, 907–921. [Google Scholar] [CrossRef]
- Wu, H.; Ye, H.; Yao, R.; Zhang, T.; Xiong, L. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling and modulates salt stress tolerance in rice. Plant Sci. 2015, 232, 1–12. [Google Scholar] [CrossRef]
- Lv, Y.; Yang, M.; Hu, D.; Yang, Z.; Ma, S.; Li, X.; Xiong, L. The OsMYB30 transcription factor suppresses cold tolerance by interacting with a JAZ protein and suppressing β-amylase expression. Plant Physiol. 2017, 173, 1475–1491. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Cai, H.; Luo, X.; Bai, X.; Deyholos, M.K.; Chen, Q.; Chen, C.; Ji, W.; Zhu, Y. Over-expression of a novel JAZ family gene from Glycine soja, increases salt and alkali stress tolerance. Biochem. Biophys. Res. Commun. 2012, 426, 273–279. [Google Scholar] [CrossRef]
- Kim, T.H.; Bohmer, M.; Hu, H.; Nishimura, N.; Schroeder, J.I. Guard cell signal transduction network: Advances in understanding abscisic acid, CO2, and Ca2+ signaling. Annu. Rev. Plant Biol. 2010, 61, 561–591. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Chen, X.; Hong, Y.Y.; Wang, Y.; Xu, P.; Ke, S.D.; Liu, H.Y.; Zhu, J.K.; Oliver, D.J.; Xiang, C.B. Activated expression of an Arabidopsis HD-START protein confers drought tolerance with improved root system and reduced stomatal density. Plant Cell 2008, 20, 1134–1151. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Zhou, G. Responses of leaf stomatal density to water status and its relationship with photosynthesis in a grass. J. Exp. Bot. 2008, 59, 3317–3325. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Guzman, M.; Pizzio, G.A.; Antoni, R.; Vera-Sirera, F.; Merilo, E.; Bassel, G.W.; Fernandez, M.A.; Holdsworth, M.J.; Perez-Amador, M.A.; Kollist, H.; et al. Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stomatal aperture and transcriptional response to abscisic acid. Plant Cell 2012, 24, 2483–2496. [Google Scholar] [CrossRef] [Green Version]
- Leung, J.; Giraudat, J. Aabscisic acid signal transduction. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 199–222. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.Y.; Chao, D.Y.; Gao, J.P.; Zhu, M.Z.; Shi, M.; Lin, H.X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control. Genes Dev. 2009, 23, 1805–1817. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).