
Physiological and Growth Responses of W. Murcott Tangor Grafted on Four Rootstocks under Water Restriction

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Design
2.2. Leaf Gas Exchange, Relative Chlorophyll Content, and Chlorophyll Fluorescence
2.3. Stem Water Potential
2.4. Trunk Cross-Sectional Area and Shoot Length
2.5. Data Analyses
3. Results
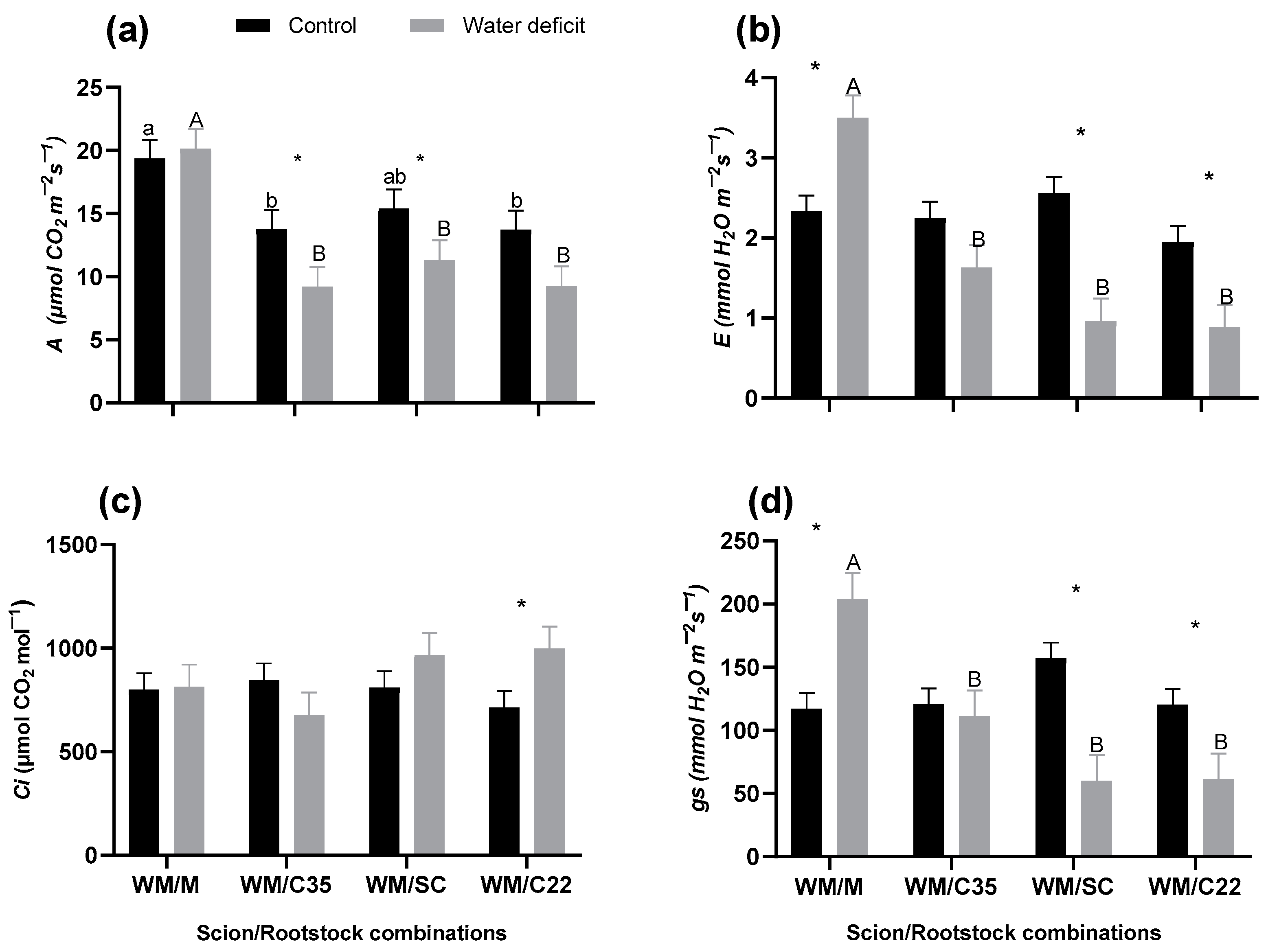
3.1. Physiological Variables
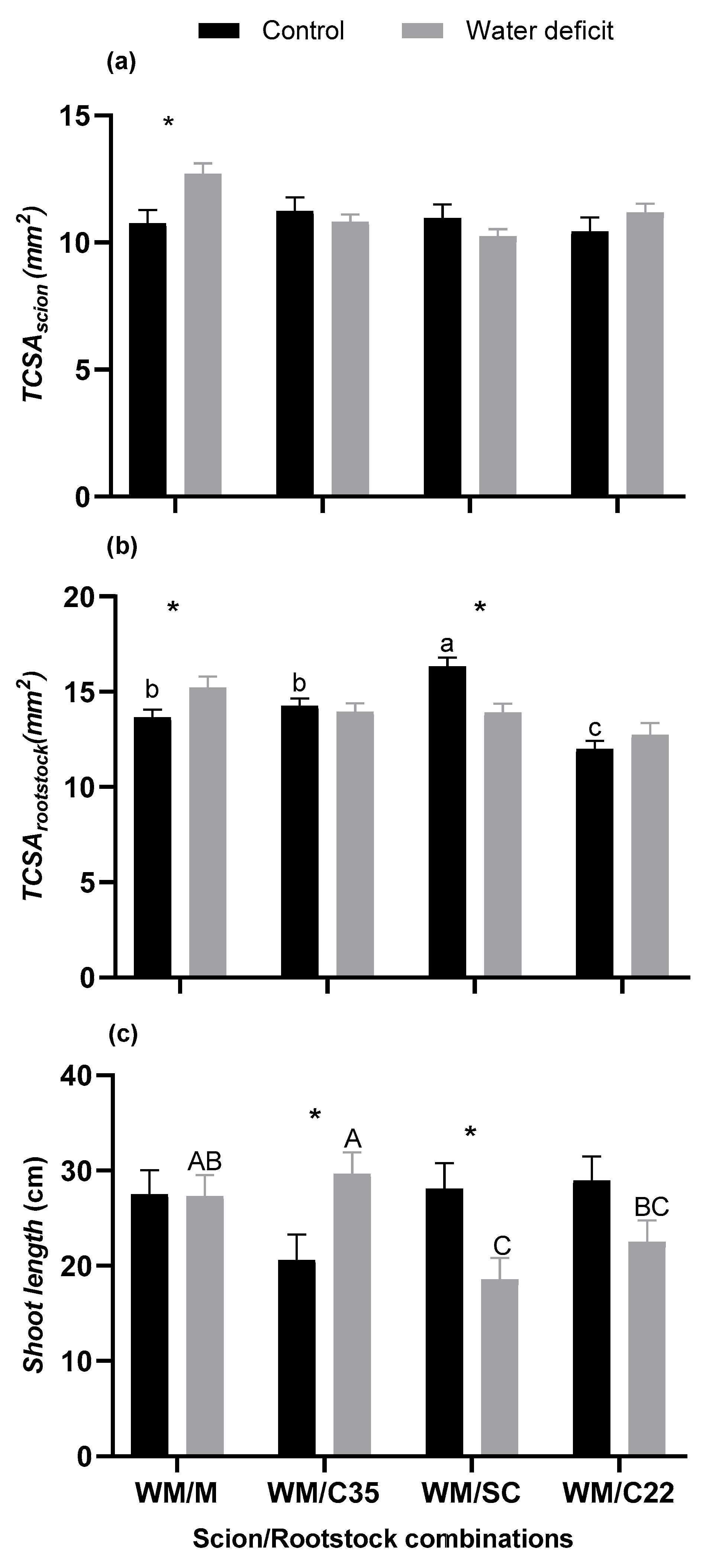
3.2. Vegetative Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO. “Resultados.” Estados Unidos de América Importaciones—Cantidad Tangerinas, Mandarinas, Clementinas. 2021. Available online: https://www.fao.org/faostat/en/#compare (accessed on 5 December 2023).
- Barry, G.H.; Caruso, M.; Gmitter, F.G. Commercial Scion Varieties. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Jr., Eds.; Woodhead Publishing: Duxford, UK, 2020; pp. 83–104. [Google Scholar]
- Jamshidi, S.; Zand, S.; Dev, P. Physiological Responses of Orange Trees Subject to Regulated Deficit Irrigation and Partial Root Drying. Irrig. Sci. 2021, 39, 441–455. [Google Scholar] [CrossRef]
- Puig-Sirera, A.; Provenzano, G.; González-Altozano, P.; Intrigliolo, D.S.; Rallo, G. Irrigation Water Saving Strategies in Citrus Orchards: Analysis of the Combined Effects of Timing and Severity of Soil Water Deficit. Agric. Water Manag. 2021, 248, 106773. [Google Scholar] [CrossRef]
- Sampaio, A.; Oliveira Silva, R.; Franca Brito, R.B.; Soares, W.; da Silva, A.; Duarte, L.; Coelho, M. Sweet Orange Acclimatisation to Water Stress: A Rootstock Dependency. Sci. Hortic. 2021, 276, 109727. [Google Scholar] [CrossRef]
- Miranda, T.; Da Silva, S.F.; Silveira, N.; Pereira, L.; Machado, E.C.; Ribeiro, R.V. Root Osmotic Adjustment and Stomatal Control of Leaf Gas Exchange Are Dependent on Citrus Rootstocks Under Water Deficit. J. Plant Growth Regul. 2020, 40, 11–19. [Google Scholar] [CrossRef]
- Puglisi, I.; Nicolosi, E.; Vanella, D.; Lo Piero, A.; Stagno, F.; Saitta, D.; Roccuzzo, G.; Consoli, S.; Baglieri, A. Physiological and Biochemical Responses of Orange Trees to Different Deficit Irrigation Regimes. Plants 2019, 8, 423. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.S.; Neves, D.; Santana-Vieira, D.D.S.; Almeida, L.; Costa, M.; Soares, W.; Pirovani, C.; Coelho, M.; Ferreira, C.; Gesteira, A. Citrus Scion and Rootstock Combinations Show Changes in DNA Methylation Profiles and ABA Insensitivity under Recurrent Drought Conditions. Sci. Hortic. 2020, 267, 109313. [Google Scholar] [CrossRef]
- Dong, T.; Xi, L.; Bo, X.; Qiu, X.; Huang, S.; Xu, W.; Wang, J.; Wang, B.; Yao, Y.; Duan Ch Tang, X.; et al. Drought Resistance in Harumi Tangor Seedlings Grafted onto Different Rootstocks. Funct. Plant Biol. 2021, 48, 529–541. [Google Scholar] [CrossRef] [PubMed]
- Paudel, I.; Cohen, S.; Shlizerman, L.; Jaiswal, A.K.; Shaviv, A.; Sadka, A. Reductions in Root Hydraulic Conductivity in Response to Clay Soil and Treated Waste Water Are Related to PIPs Down-Regulation in Citrus. Sci. Rep. 2017, 7, 15429. [Google Scholar] [CrossRef] [PubMed]
- Robles, J.M.; Botía, P.; Pérez-Pérez, J.G. Sour Orange Rootstock Increases Water Productivity in Deficit Irrigated ‘Verna’ Lemon Trees Compared with Citrus Macrophylla. Agric. Water Manag. 2017, 186, 98–107. [Google Scholar] [CrossRef]
- Jiménez, R.; Zamora, V. Principales Cultivares y Patrones Utilizados En La Citricultura. In Taller Reg. Sobre Viveros De Cítricos; Instituto de Investigaciones en Fruticultura Tropical: La Habana, Cuba, 2010; Volume 44, ISBN 978-959-296-020-6. [Google Scholar]
- Bowman, K.D.; Joubert, J. Citrus Rootstocks. In The Genus Citrus; Talon, M., Caruso, M., Gmitter, F.G., Jr., Eds.; Woodhead Publishing: Duxford, UK, 2020; pp. 105–127. [Google Scholar]
- Evett, S.R.; Schwartz, R.; Casanova, J.; Heng, L. Soil Water Sensing for Water Balance, ET and WUE. Agric. Water Manag. 2012, 104, 1–9. [Google Scholar] [CrossRef]
- Passioura, J.B. Viewpoint: The perils of pot experiments. Funct.Plant Biol. 2006, 33, 1075–1079. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, U. Facilitating Protocols While Maintaining Accuracy in Grapevine Pressure Chamber Measurements-Comments on Levin 2019. Agric. Water Manag. 2020, 227, 2019–2021. [Google Scholar] [CrossRef]
- Jiménez, S.; Fattahi, M.; Bedis, K.; Nasrolahpour-Moghadam, S.; Irigoyen, J.; Gogorcena, Y. Interactional Effects of Climate Change Factors on the Water Status, Photosynthetic Rate, and Metabolic Regulation in Peach. Front. Plant Sci. 2020, 11, 43. [Google Scholar] [CrossRef]
- Tu, A.; Xie, S.; Mo, M.; Song, Y.; Li, Y. Water Budget Components Estimation for a Mature Citrus Orchard of Southern China Based on HYDRUS-1D Model. Agric. Water Manag. 2021, 243, 106426. [Google Scholar] [CrossRef]
- Singh, G.; Singh, H.; Gupta, M.; Singh Sidhu, G. Standardization of Stage Wise Water Requirement in Drip Irrigated Kinnow Mandarin Orchards under Sub-Tropical Conditions. J. Agrometeorol. 2020, 22, 305–312. [Google Scholar] [CrossRef]
- Nagaz, K.; El Mokh, F.; Ben Hassen, N.; Masmoudi, M.M.; Ben Mechlia, N.; Baba Sy, M.O.; Belkheiri, O.; Ghiglieri, G. Impact of Deficit Irrigation on Yield and Fruit Quality of Orange Trees (Citrus sinensis, L. Osbeck, CV. Meski maltaise) in Southern Tunisia. Irrig. Drain. 2020, 69, 186–193. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, F.; Gou, X.; Fonti, P.; Xia, J.; Cao, Z.; Liu, J.; Wang, Y.; Zhang, J. Seasonal Variations in Leaf-Level Photosynthesis and Water Use Efficiency of Three Isohydric to Anisohydric Conifers on the Tibetan Plateau. Agric. For. Meteorol. 2021, 308–309, 108581. [Google Scholar] [CrossRef]
- Hochberg, U.; Rockwell, F.E.; Michele Holbrook, N.; Cochard, H. Iso/Anisohydry: A Plant–Environment Interaction Rather Than a Simple Hydraulic Trait. Trends Plant Sci. 2018, 23, 112–120. [Google Scholar] [CrossRef] [PubMed]
- Salmon, Y.; Lintunen, A.; Dayet, A.; Chan, T.; Dewar, R.; Vesala, T.; Hölttä, T. Leaf Carbon and Water Status Control Stomatal and Nonstomatal Limitations of Photosynthesis in Trees. New Phytol. 2020, 226, 690–703. [Google Scholar] [CrossRef]
- Caruso, M.; Continella, A.; Modica, G.; Pannitteri, C.; Russo, R.; Salonia, F.; Arlotta, C.; Gentile, A.; Russo, G. Rootstocks Influence Yield Precocity, Productivity, and Pre-Harvest Fruit Drop of Mandared Pigmented Mandarin. Agronomy 2020, 10, 1305. [Google Scholar] [CrossRef]
- Bhusal, N.; Lee, M.; Reum Han, A.; Han, A.; Seok Kim, H. Responses to Drought Stress in Prunus Sargentii and Larix Kaempferi Seedlings Using Morphological and Physiological Parameters. For. Ecol. Manag. 2019, 465, 118099. [Google Scholar] [CrossRef]
- Martínez-Vilalta, J.; Garcia-Forner, N. Water Potential Regulation, Stomatal Behaviour and Hydraulic Transport under Drought: Deconstructing the Iso/Anisohydric Concept. Plant Cell Environ. 2017, 40, 962–976. [Google Scholar] [CrossRef] [PubMed]
- Livellara, N.; Saavedra, F.; Salgado, E. Plant Based Indicators for Irrigation Scheduling in Young Cherry Trees. Agric. Water Manag. 2011, 98, 684–690. [Google Scholar] [CrossRef]
- Meza, F.J.; Montes, C.; Bravo-Martínez, F.; Serrano-Ortiz, P.; Kowalski, A.S. Soil Water Content Effects on Net Ecosystem CO2 Exchange and Actual Evapotranspiration in a Mediterranean Semiarid Savanna of Central Chile. Sci. Rep. 2018, 8, 8570. [Google Scholar] [CrossRef] [PubMed]
- Serra, I.; Strever, A.; Myburgh, P.A.; Deloire, A. Review: The Interaction between Rootstocks and Cultivars (Vitis vinifera L.) to Enhance Drought Tolerance in Grapevine. Aust. J. Grape Wine Res. 2014, 20, 1–14. [Google Scholar] [CrossRef]
- Nankishore, A.; Farrell, A.D. The Response of Contrasting Tomato Genotypes to Combined Heat and Drought Stress. J. Plant Physiol. 2016, 202, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Landi, S.; Nurcato, R.; De Lillo, A.; Lentini, M.; Grillo, S.; Esposito, S. Glucose-6-Phosphate Dehydrogenase Plays a Central Role in the Response of Tomato (Solanum lycopersicum) Plants to Short and Long-Term Drought. Plant Physiol. Biochem. 2016, 105, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.C. Turgor Maintenance by Osmotic Adjustment, an Adaptive Mechanism for Coping with Plant Water Deficits. Plant Cell Environ. 2017, 40, 1–3. [Google Scholar] [CrossRef]
- Kosar, F.; Aisha Akram, N.; Sadiq, M.; Al-Qurainy, F.; Ashraf, M. Trehalose: A Key Organic Osmolyte Effectively Involved in Plant Abiotic Stress Tolerance. J. Plant Growth Regul. 2019, 38, 606–618. [Google Scholar] [CrossRef]
- Yuan, W.; Zhang, Q.; Li, Y.; Wang, Q.; Xu, F.; Dang, X.; Xu, W.; Zhang, J.; Miao, R. Abscisic Acid Is Required for Root Elongation Associated With Ca2+ Influx in Response to Water Stress. Front. Plant Sci. 2020, 11, 127–137. [Google Scholar] [CrossRef]
- Fàbregas, N.; Fernie, A.R. The Metabolic Response to Drought. J. Exp. Bot. 2019, 70, 1077–1085. [Google Scholar] [CrossRef] [PubMed]
- Carminati, A.; Javaux, M. Soil Rather Than Xylem Vulnerability Controls Stomatal Response to Drought. Trends Plant Sci. 2020, 25, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Zuo, Q.; Shi, J.; Wang, J.; Xue, X.; Ben-Gal, A. Introducing Water Stress Hysteresis to the Feddes Empirical Macroscopic Root Water Uptake Model. Agric. Water Manag. 2020, 240, 106293. [Google Scholar] [CrossRef]
- Takata, K.; Matsuzaki, T.; Tajika, Y. Aquaporins: Water Channel Proteins of the Cell Membrane. Prog. Histochem. Cytochem. 2004, 39, 1–83. [Google Scholar] [CrossRef]
- Wei, Q.; Ma, Q.; Ma, Z.; Zhou, G.; Feng, F.; Le, S.; Lei Ch Gu, Q. Genome-Wide Identification and Characterization of Sweet Orange (Citrus sinensis) Aquaporin Genes and Their Expression in Two Citrus Cultivars Differing in Drought Tolerance. Tree Genet. Genomes 2019, 15, 17. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Rootstocks | Growth | Frost | Drought | Flooding | Salinity | Alkalinity | Clay Soil | Silty Soil | Sandy Soil |
|---|---|---|---|---|---|---|---|---|---|
| Swingle Citrumelo | Vigorous | 1 | 1 | 2 | 1 | 3 | nif | x | nif |
| Citrange C-35 | Semi-dwarfing | 3 | nif | 1 | nif | 2 | nif | x | nif |
| Macrophylla | Vigorous | 3 | 1 | 3 | 1 | 1 | x | x | nif |
| Bitter Citrandarin (C-22) | Semi-dwarfing | nif | nif | 1 | 1 | 1 | x | x | nif |
| ET (%) | Number of Drippers per Plant (Efficiency of 90%) | Flow Rate per Plant (L/h) | Mean Precipitation per Treatment (mm/h) | Irrigation Time (min/day) | Volumetric Water Content at Pot Capacity (θpt m3/m3) |
|---|---|---|---|---|---|
| 100 | 1 | 2 | 7.94 | 3 | 0.40 |
| 75 | 1 | 2 | 5.32 | 2.3 | 0.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tobar, S.; Gil, P.M.; Schaffer, B.; Schwember, A.R.; Cautín, R.; Mártiz, J. Physiological and Growth Responses of W. Murcott Tangor Grafted on Four Rootstocks under Water Restriction. Horticulturae 2024, 10, 352. https://doi.org/10.3390/horticulturae10040352
Tobar S, Gil PM, Schaffer B, Schwember AR, Cautín R, Mártiz J. Physiological and Growth Responses of W. Murcott Tangor Grafted on Four Rootstocks under Water Restriction. Horticulturae. 2024; 10(4):352. https://doi.org/10.3390/horticulturae10040352
Chicago/Turabian StyleTobar, Sophia, Pilar M. Gil, Bruce Schaffer, Andrés R. Schwember, Ricardo Cautín, and Johanna Mártiz. 2024. "Physiological and Growth Responses of W. Murcott Tangor Grafted on Four Rootstocks under Water Restriction" Horticulturae 10, no. 4: 352. https://doi.org/10.3390/horticulturae10040352
APA StyleTobar, S., Gil, P. M., Schaffer, B., Schwember, A. R., Cautín, R., & Mártiz, J. (2024). Physiological and Growth Responses of W. Murcott Tangor Grafted on Four Rootstocks under Water Restriction. Horticulturae, 10(4), 352. https://doi.org/10.3390/horticulturae10040352