

Changes in Physicochemical Properties, Metabolites and Antioxidant Activity of Edible Grass during Spontaneous Fermentation
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Spontaneous Fermentation of Edible Grass
2.3. Determination of Physicochemical Properties of Fermented Edible Grass
2.4. Metabolite Profiling Analysis
2.5. Free Radical Scavenging Activity
2.6. Cell Culture and Cell Viability Assay
2.7. Induction of the H2O2-Mediated Oxidative Damaged HepG2 Cells Model
2.8. Determination of ROS Level and Antioxidant Enzyme Activities
2.9. Data Analysis
3. Results and Discussion
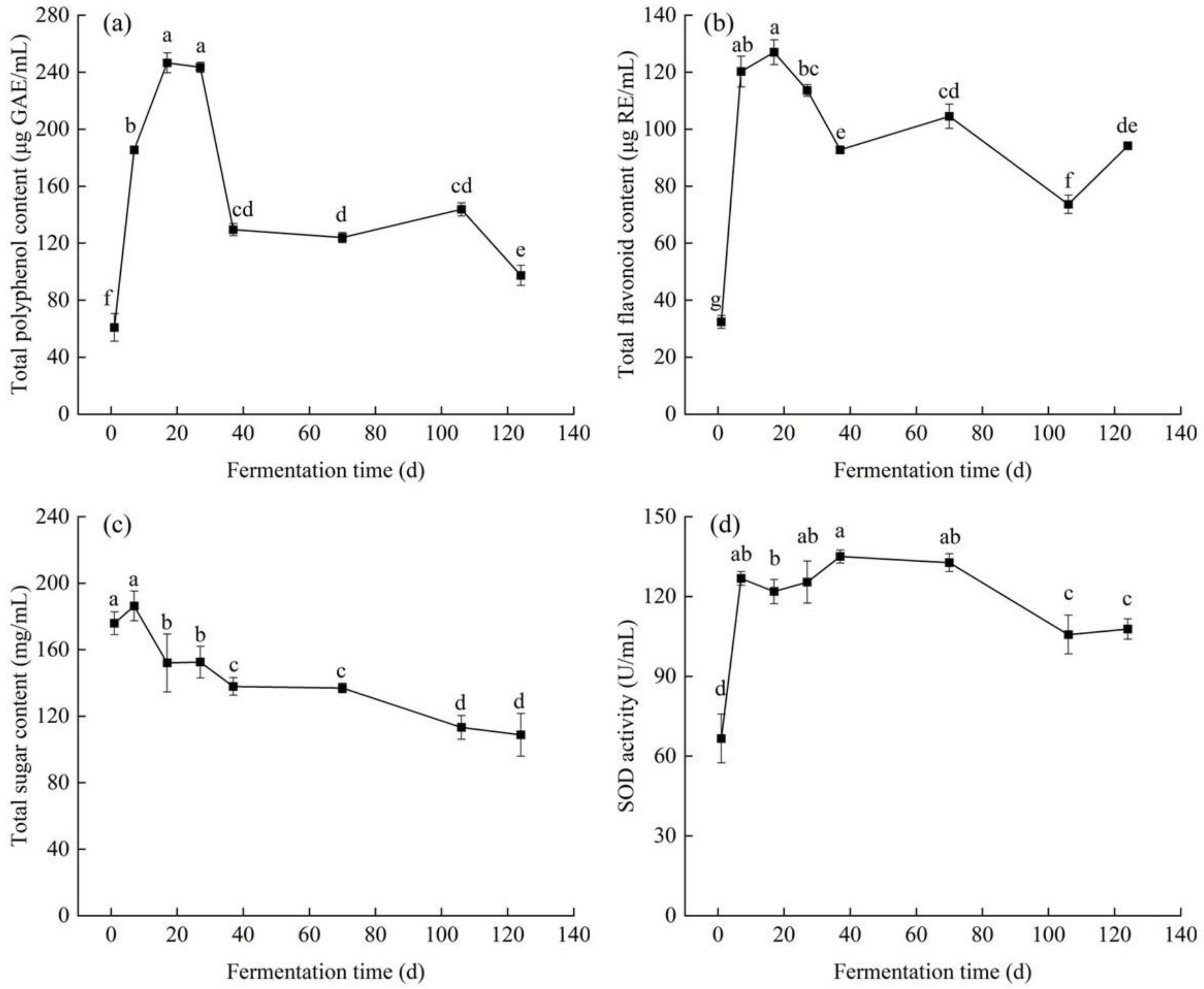
3.1. Changes in Physicochemical Properties of Edible Grass during Spontaneous Fermentation
3.1.1. Total Polyphenol Content and Total Flavonoid Content
3.1.2. Total Sugar Content and Superoxide Dismutase Activity
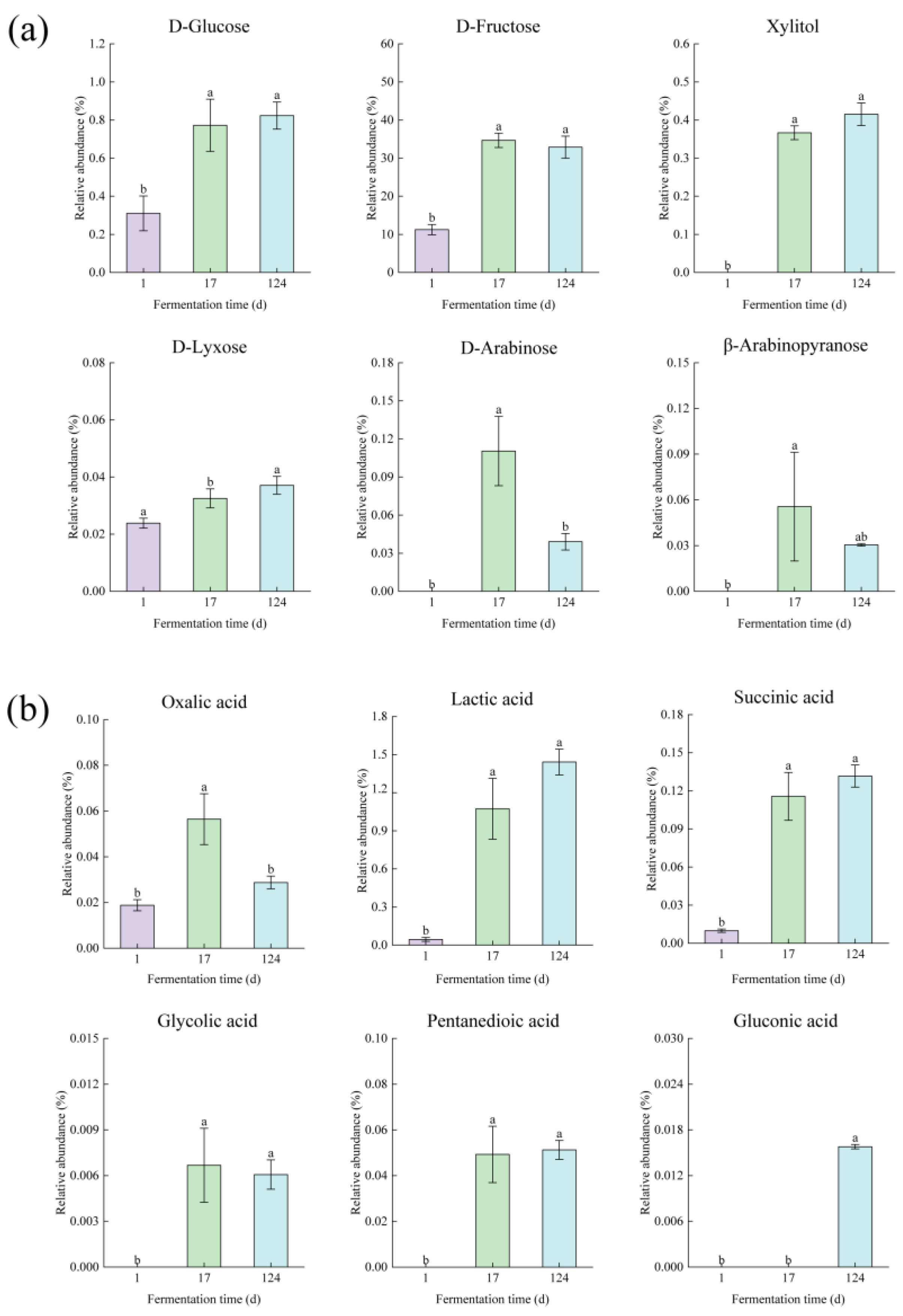
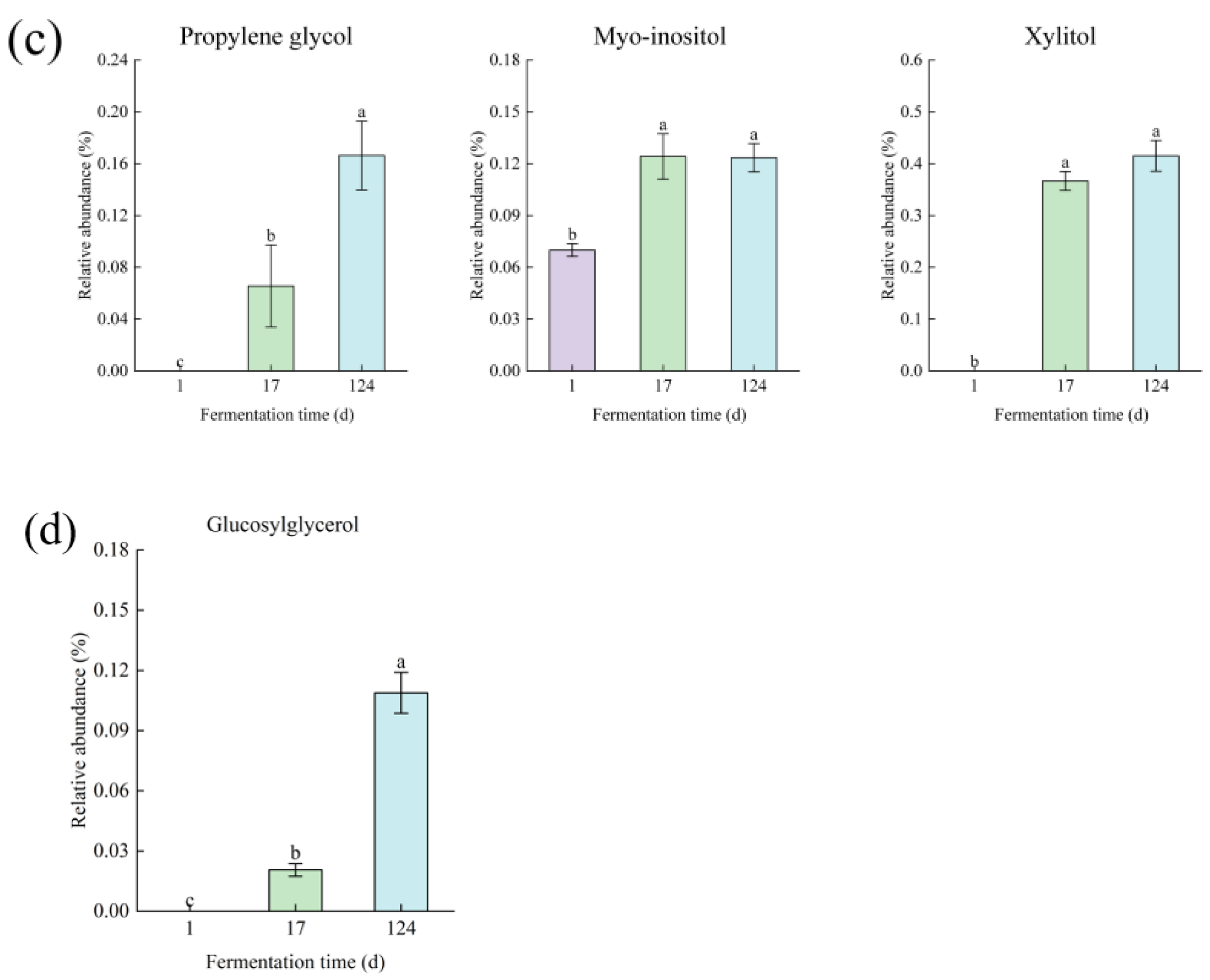
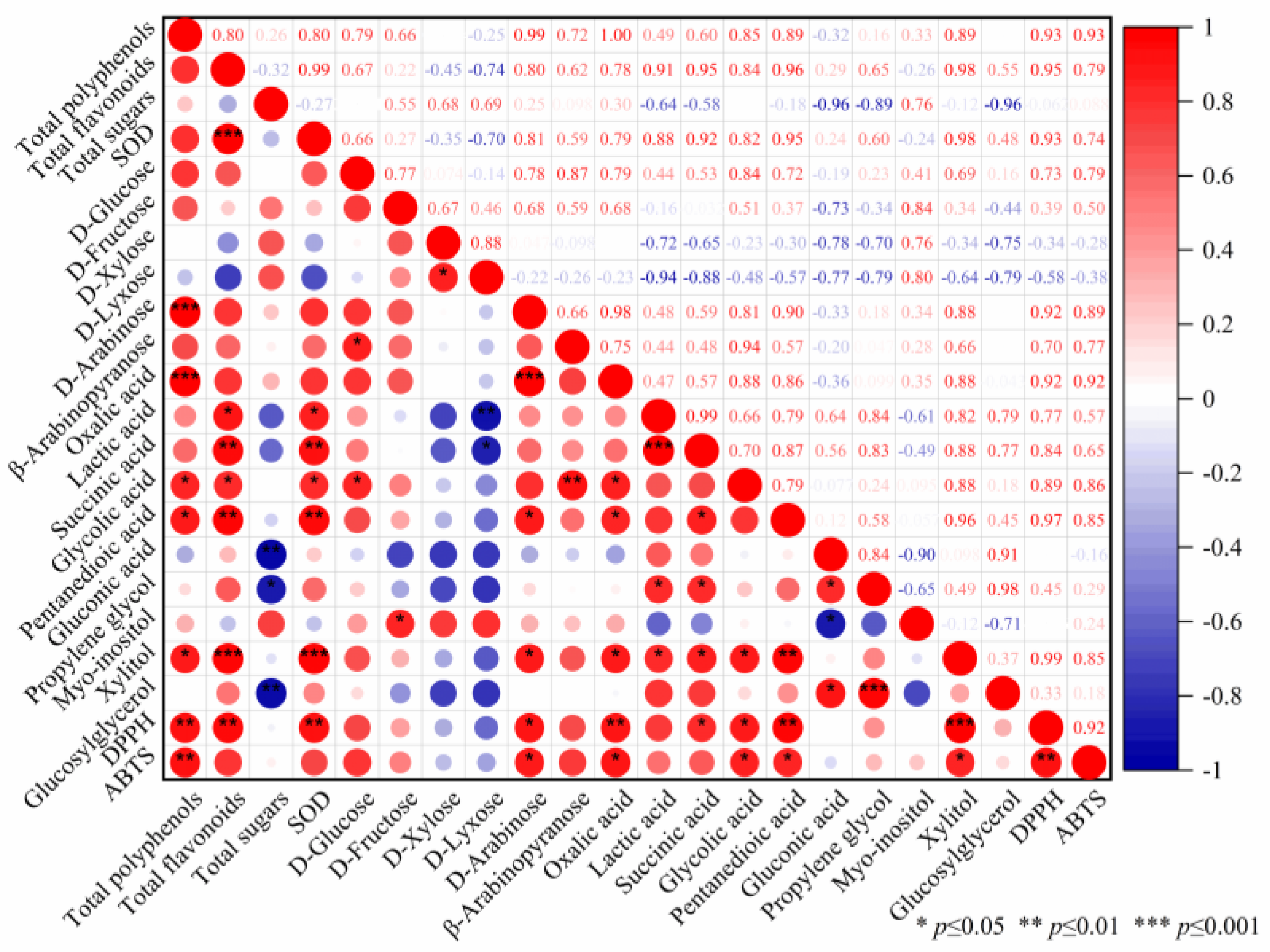
3.2. Metabolomics Analysis in the Fermentation Process of Edible Grass
3.2.1. Multivariate Analysis of Gas Chromatography Mass Spectrometry Based Metabolomics Data
3.2.2. Metabolite Differences in Fermented Edible Grass at Different Fermentation Times
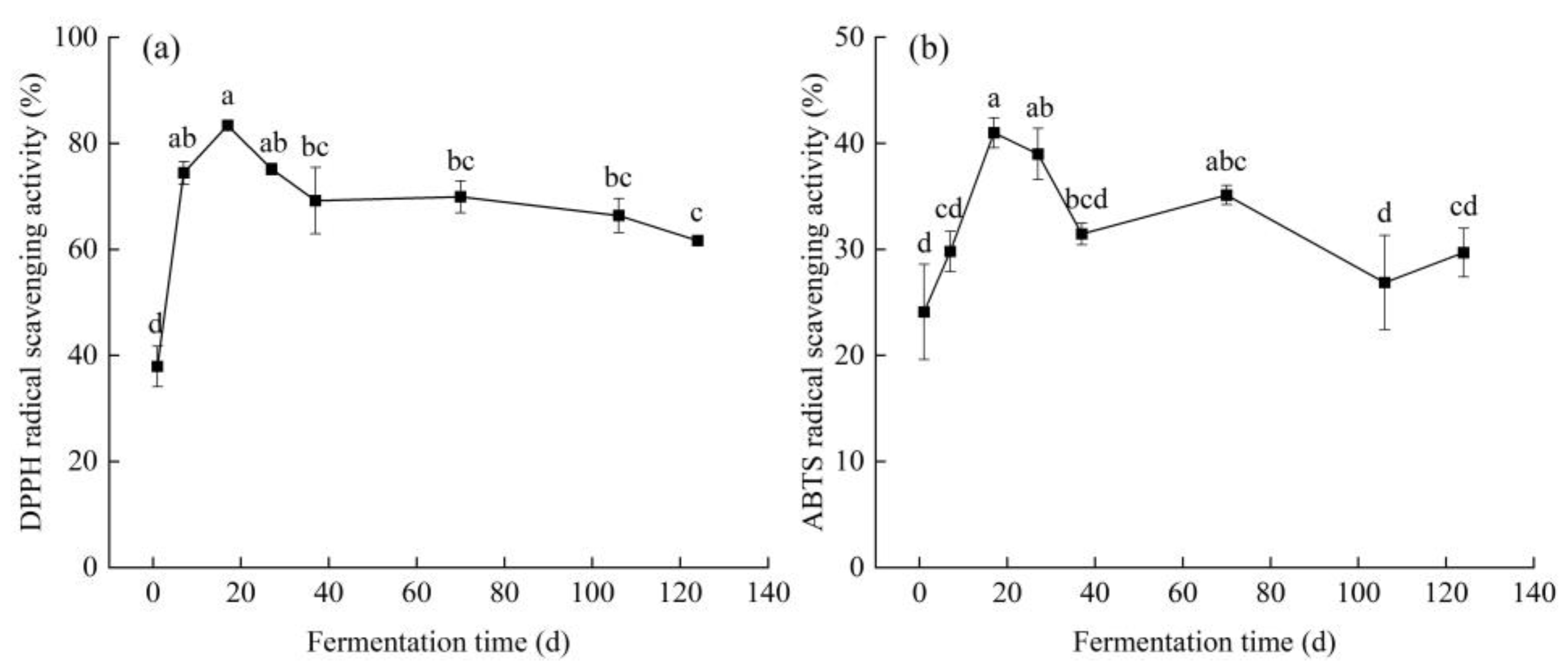
3.3. Antioxidant Activity
3.3.1. Free Radical Scavenging Activity
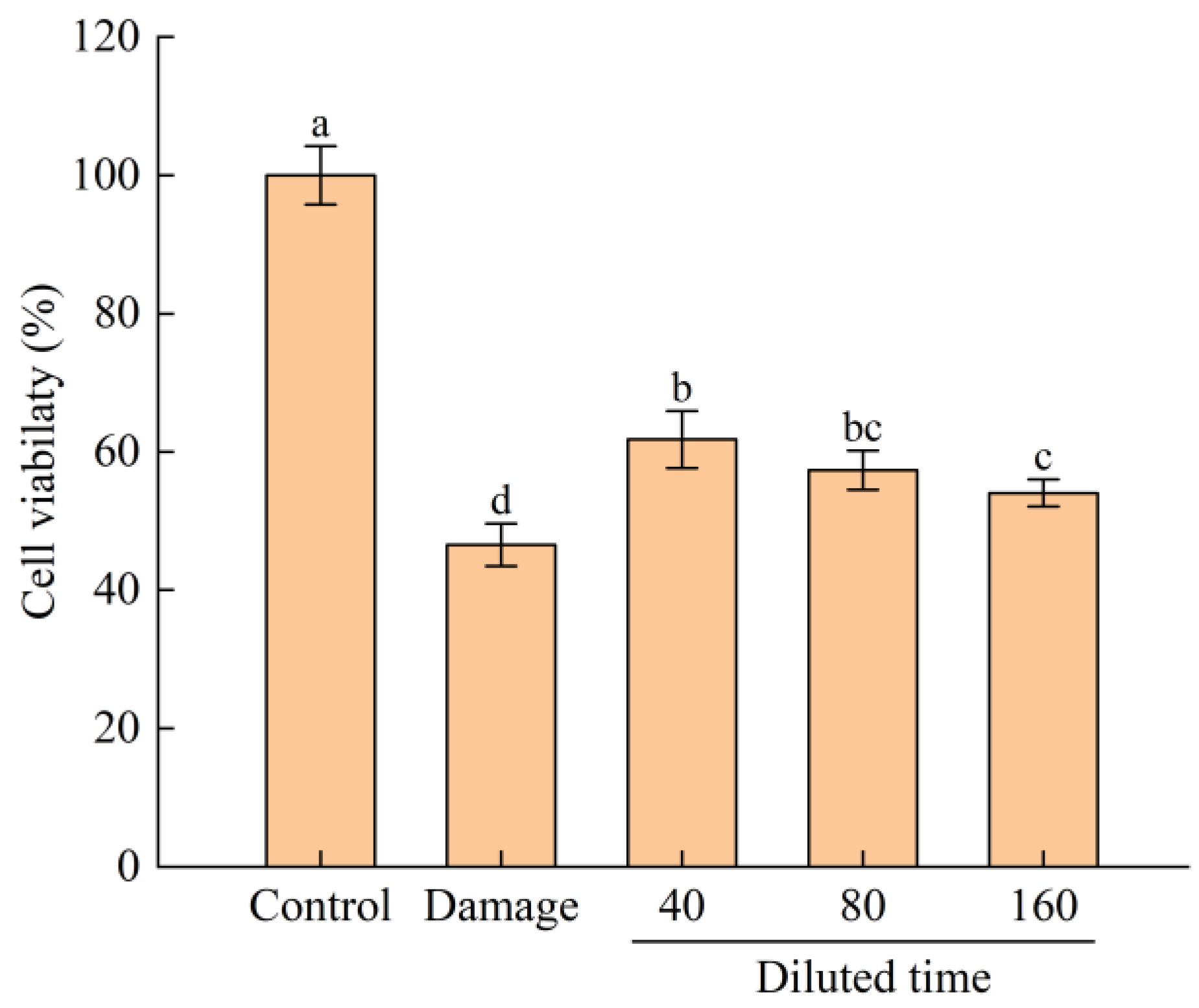
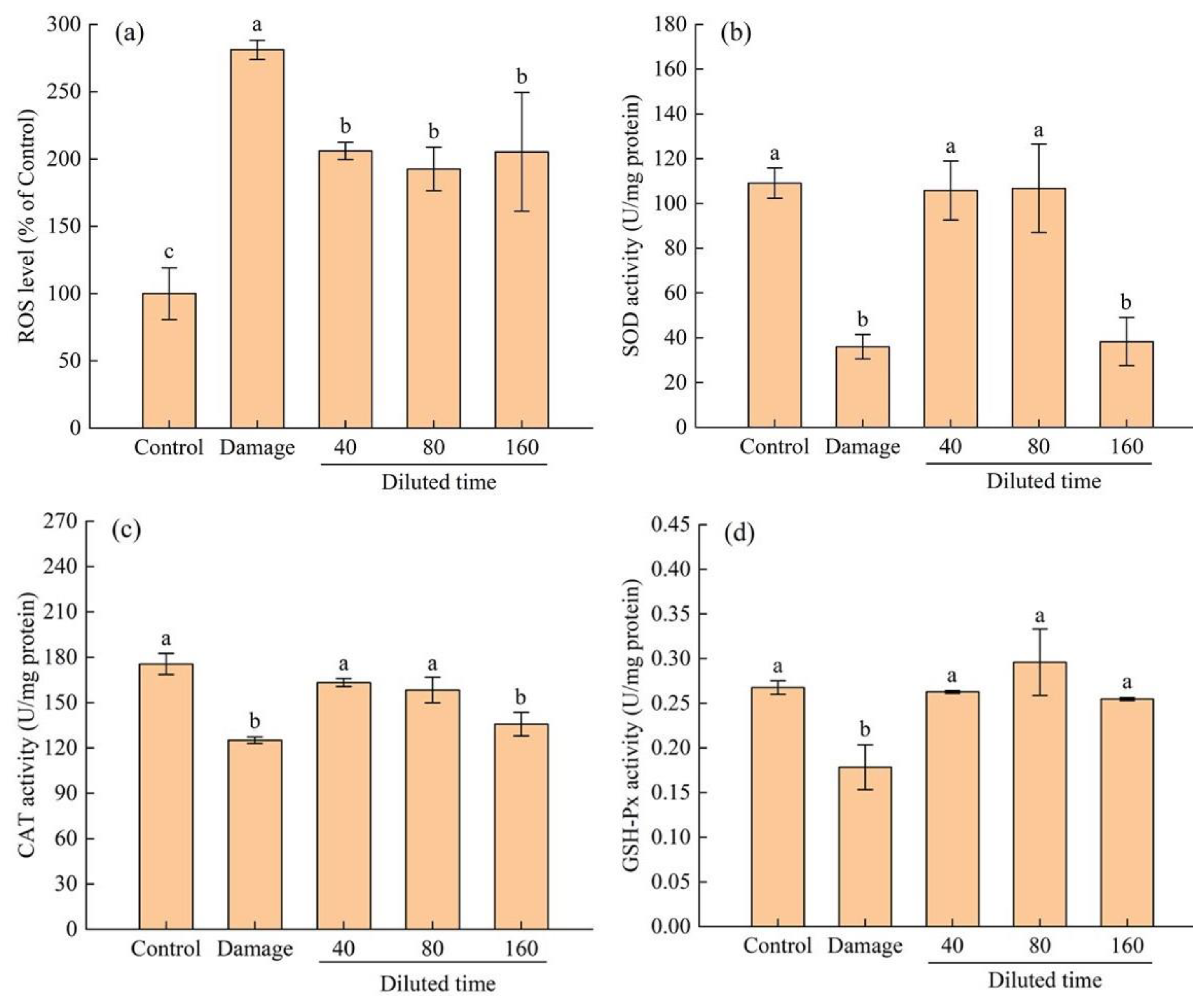
3.3.2. Protective Effect of Fermented Edible Grass on the H2O2-Mediated Oxidative Injured HepG2 Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.; He, T.; Mao, J.; Sha, R. Effects of lactic acid bacteria fermentation on physicochemical properties, functional compounds and antioxidant activity of edible grass. Fermentation 2022, 8, 647. [Google Scholar] [CrossRef]
- Li, X.; He, T.; Yang, F.; Wang, C.; Zhou, Y.; Sha, R.; Mao, J. Analysis of nutritional components, functional components and bioactivity of edible grass. Sci. Technol. Food Ind. 2023, 44, 307–315. [Google Scholar]
- Lou, M.; Qu, X.; Zhang, L.; Liu, Z.; Hu, Z.; Wang, Y. Safety evaluation of edible grass as a new food raw material. J. Food Saf. Qual. 2021, 12, 3919–3926. [Google Scholar]
- Handa, C.L.; de Lima, F.S.; Guelfi, M.F.G.; da Silva Fernandes, M.; Georgetti, S.R.; Ida, E.I. Parameters of the fermentation of soybean flour by Monascus purpureus or Aspergillus oryzae on the production of bioactive compounds and antioxidant activity. Food Chem. 2019, 271, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Luo, Y.; Wu, Y.; Liu, Y.; Wu, Z. Fermentation and complex enzyme hydrolysis for improving the total soluble phenolic contents, flavonoid aglycones contents and bio-activities of guava leaves tea. Food Chem. 2018, 264, 189–198. [Google Scholar] [CrossRef]
- Iñaki, D.-O.; Juaristi, A.O. Fermented foods: An update on evidence-based health benefits and future perspectives. Food Res. Int. 2022, 156, 111133. [Google Scholar]
- Zhao, Y.; Wu, C.; Zhu, Y.; Zhou, C.; Xiong, Z.; Eweys, A.S.; Zhou, H.; Dong, Y.; Xiao, X. Metabolomics strategy for revealing the components in fermented barley extracts with Lactobacillus plantarum dy-1. Food Res. Int. 2020, 139, 109808. [Google Scholar] [CrossRef]
- Villacrés, E.; Quelal, M.B.; Fernández, E.; Garcìa, G.; Cueva, G.; Rosell, C.M. Impact of debittering and fermentation processes on the antinutritional and antioxidant compounds in Lupinus mutabilis sweet. LWT 2020, 131, 109745. [Google Scholar] [CrossRef]
- Sew, S.W.; Lu, Y.; Taniasuri, F.; Liu, S.Q. Chemical analysis and flavour compound changes of vegetable blend slurry fermented with selected probiotic bacteria. Food Biosci. 2022, 47, 101784. [Google Scholar] [CrossRef]
- Balcázar-Zumaeta, C.R.; Castro-Alayo, E.M.; Cayo-Colca, I.S.; Idrogo-Vásquez, G.; Muñoz-Astecker, L.D. Metabolomics during the spontaneous fermentation in cocoa (Theobroma cacao L.): An exploraty review. Food Res. Int. 2023, 163, 112190. [Google Scholar] [CrossRef]
- Antoniewicz, J.; Jakubczyk, K.; Kwiatkowski, P.; Maciejewska-Markiewicz, D.; Kochman, J.; Rebacz-Maron, E.; Janda-Milczarek, K. Analysis of antioxidant capacity and antimicrobial properties of selected polish grape vinegars obtained by spontaneous fermentation. Molecules 2021, 26, 4727. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Liu, Y.Q.; Lv, J.W.; Ma, Y.L.; Guan, X.L. Changes in the physicochemical components, polyphenol profile, and flavor of persimmon wine during spontaneous and inoculated fermentation. Food Sci. Nutr. 2020, 8, 2728–2738. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Zhu, Y.; Fang, C.; Wijffels, R.H.; Xu, Y. Can we control microbiota in spontaneous food fermentation?–Chinese liquor as a case example. Trends Food Sci. Technol. 2021, 110, 321–331. [Google Scholar] [CrossRef]
- Leyva, A.; Quintana, A.; Sánchez, M.; Rodríguez, E.N.; Cremata, J.; Sánchez, J.C. Rapid and sensitive anthrone–sulfuric acid assay in microplate format to quantify carbohydrate in biopharmaceutical products: Method development and validation. Biologicals 2007, 36, 134–141. [Google Scholar] [CrossRef]
- Dai, J.; Hu, Y.; Si, Q.; Gu, Y.; Xiao, Z.; Ge, Q.; Sha, R. Antioxidant and hypoglycemic activity of sequentially extracted fractions from pingguoli pear fermentation broth and identification of bioactive compounds. Molecules 2022, 27, 6077. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Song, Y.; Liang, Y.; Li, Y.; Chang, Y.; Ma, R.; Cao, X.; Wang, S. Dynamics of physicochemical properties, functional compounds and antioxidant capacity during spontaneous fermentation of Lycium ruthenicum Murr. (Qinghai-Tibet Plateau) natural vinegar. Foods 2022, 11, 1344. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Chu, S.C.; Chen, C. Effects of origins and fermentation time on the antioxidant activities of kombucha. Food Chem. 2006, 98, 502–507. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, H.; Liu, H.; Ma, R.; Ma, J.; Fang, H.; Ferreira, I.C.F.R.; McPhee, D.J. Fermentation by multiple bacterial strains improves the production of bioactive compounds and antioxidant activity of goji juice. Molecules 2019, 24, 3519. [Google Scholar] [CrossRef]
- Haile, M.; Kang, W.H. Antioxidant activity, total polyphenol, flavonoid and tannin contents of fermented green coffee beans with selected yeasts. Fermentation 2019, 5, 29. [Google Scholar] [CrossRef]
- Adebo, O.A.; Njobeh, P.B.; Adebiyi, J.A.; Kayitesi, E. Co-influence of fermentation time and temperature on physicochemical properties, bioactive components and microstructure of ting (a Southern African food) from whole grain sorghum. Food Biosci. 2018, 25, 118–127. [Google Scholar] [CrossRef]
- Liu, W.; Dun, M.; Liu, X.; Zhang, G.; Ling, J. Effects on total phenolic and flavonoid content, antioxidant properties, and angiotensin I-converting enzyme inhibitory activity of beans by solid-state fermentation with Cordyceps militaris. Int. J. Food Prop. 2022, 25, 477–491. [Google Scholar] [CrossRef]
- Lopusiewicz, L.; Drozlowska, E.; Trocer, P.; Kwiatkowski, P.; Bartkowiak, A.; Gefrom, A.; Sienkiewicz, M. The effect of fermentation with kefir grains on the physicochemical and antioxidant properties of beverages from blue lupin (Lupinus angustifolius L.) seeds. Molecules 2020, 25, 5791. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H.; Kim, M.-J.; Kim, D.-W.; Kim, G.-Y.; Kim, J.-K.; Gebru, Y.A.; Choi, H.-S.; Kim, Y.-H.; Kim, M.-K. Changes of phytochemical components (urushiols, polyphenols, gallotannins) and antioxidant capacity during fomitella fraxinea-mediated fermentation of Toxicodendron vernicifluum Bark. Molecules 2019, 24, 683. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Wang, T.; Wang, R.; Luo, X. Effects of five different lactic acid bacteria on bioactive components and volatile compounds of oat. Foods 2022, 11, 3230. [Google Scholar] [CrossRef]
- Kiai, H.; Hafidi, A. Chemical composition changes in four green olive cultivars during spontaneous fermentation. Lwt-Food Sci. Technol. 2014, 57, 663–670. [Google Scholar] [CrossRef]
- Zhang, P.W.; Li, H.; Zhao, W.P.; Xiong, K.; Wen, H.; Yang, H.L.; Wang, X.L. Dynamic analysis of physicochemical characteristics and microbial communities of Aspergillus-type douchi during fermentation. Food Res. Int. 2022, 153, 110932. [Google Scholar] [CrossRef]
- do Carmo Brito, B.d.N.; Campos Chisté, R.; Santos Lopes, A.; Abreu Glória, M.B.; da Silva Pena, R. Influence of spontaneous fermentation of manipueira on bioactive amine and carotenoid profiles during tucupi production. Food Res. Int. 2019, 120, 209–216. [Google Scholar] [CrossRef]
- Quan, Q.; Liu, W.; Guo, J.; Ye, M.; Zhang, J. Effect of six lactic acid bacteria strains on physicochemical characteristics, antioxidant activities and sensory properties of fermented orange juices. Foods 2022, 11, 1920. [Google Scholar] [CrossRef]
- Fan, X.; Li, X.; Zhang, T.; Guo, Y.; Shi, Z.; Wu, Z.; Zeng, X.; Pan, D. Novel millet-based flavored yogurt enriched with superoxide dismutase. Front. Nutr. 2022, 8, 791886. [Google Scholar] [CrossRef]
- Kong, L.H.; Xiong, Z.Q.; Song, X.; Xia, Y.J.; Zhang, H.; Yang, Y.; Ai, L.Z. Enhanced antioxidant activity in Streptococcus thermophilus by high-level expression of superoxide dismutase. Front. Microbiol. 2020, 11, 579804. [Google Scholar] [CrossRef]
- Zhou, X.; Guan, Q.; Qin, Y.; Qin, Z.; Lin, D. Dynamic changes in physic-chemical properties and bacterial community during natural fermentation of tomatoes. Food Sci. Technol. 2021, 42, 63520. [Google Scholar] [CrossRef]
- Yang, X.; Zhou, J.; Fan, L.; Qin, Z.; Chen, Q.; Zhao, L. Antioxidant properties of a vegetable-fruit beverage fermented with two Lactobacillus plantarum strains. Food Sci. Biotechnol. 2018, 27, 1719–1726. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-E.; Yoo, S.-A.; Seo, S.-H.; Lee, K.-I.; Na, C.-S.; Son, H.-S. GC–MS based metabolomics approach of Kimchi for the understanding of Lactobacillus plantarum fermentation characteristics. Lwt-Food Sci. Technol. 2016, 68, 313–321. [Google Scholar] [CrossRef]
- Zhao, N.; Zhang, C.; Yang, Q.; Guo, Z.; Yang, B.; Lu, W.; Li, D.; Tian, F.; Liu, X.; Zhang, H.; et al. Selection of taste markers related to lactic acid bacteria microflora metabolism for Chinese traditional paocai: A gas chromatography-mass spectrometry-based metabolomics approach. J. Agric. Food Chem. 2016, 64, 2415–2422. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Mu, Y.; Wei, S.; Mu, Y.; Zhao, C. Study on the dynamic changes and formation pathways of metabolites during the fermentation of black waxy rice wine. Food Sci. Nutr. 2020, 8, 2288–2298. [Google Scholar] [CrossRef]
- Vitas, J.S.; Cvetanović, A.D.; Mašković, P.Z.; Švarc-Gajić, J.V.; Malbaša, R.V. Chemical composition and biological activity of novel types of kombucha beverages with yarrow. J. Funct. Foods 2018, 44, 95–102. [Google Scholar] [CrossRef]
- Lian, Z.; Dai, L.; Zhang, R.; Liu, Y.; Zhou, X.; Xu, Y. Efficient aerobic fermentation of gluconic acid by high tension oxygen supply strategy with reusable Gluconobacter oxydans HG19 cells. Bioprocess Biosyst. Eng. 2022, 45, 1849–1855. [Google Scholar] [CrossRef]
- Chen, W.C.; Chuang, C.J.; Chang, J.S.; Wang, L.F.; Soo, P.C.; Wu, H.S.; Tsai, S.L.; Wei, Y.H. Exploring dual-substrate cultivation strategy of 1,3-propanediol production using Klebsiella pneumoniae. Appl. Biochem. Biotechnol. 2020, 191, 346–359. [Google Scholar] [CrossRef]
- Ma, Z.; Shentu, X.P.; Bian, Y.L.; Yu, X.P. 1,3-propanediol production from glucose by mixed-culture fermentation of Zygosacharomyces rouxii and Klebsiella pneumonia. Eng. Life Sci. 2012, 12, 553–559. [Google Scholar] [CrossRef]
- You, R.; Wang, L.; Shi, C.R.; Chen, H.; Zhang, S.S.; Hu, M.R.; Tao, Y. Efficient production of myo-inositol in Escherichia coli through metabolic engineering. Microb. Cell Factories 2020, 19, 109. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; Utsumi, Y.; Sawayama, S.; Watanabe, Y. Identification and characterization of d-arabinose reductase and d-arabinose transporters from Pichia stipitis. Biosci. Biotechnol. Biochem. 2016, 80, 2151–2158. [Google Scholar] [CrossRef]
- Qi, J.; Huang, H.; Wang, J.; Liu, N.; Chen, X.; Jiang, T.; Xu, H.; Lei, H. Insights into the improvement of bioactive phytochemicals, antioxidant activities and flavor profiles in Chinese wolfberry juice by select lactic acid bacteria. Food Biosci. 2021, 43, 101264. [Google Scholar] [CrossRef]
- McCue, P.P.; Shetty, K. Phenolic antioxidant mobilization during yogurt production from soymilk using Kefir cultures. Process Biochem. 2004, 40, 1791–1797. [Google Scholar] [CrossRef]
- Hu, Y.-m.; Lu, S.-z.; Li, Y.-s.; Wang, H.; Shi, Y.; Zhang, L.; Tu, Z.-c. Protective effect of antioxidant peptides from grass carp scale gelatin on the H2O2-mediated oxidative injured HepG2 cells. Food Chem. 2022, 373, 131539. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, H.; Li, H.; Lai, F.; Li, X.; Tang, Y.; Min, T.; Wu, H. Antioxidant mechanism of betaine without free radical scavenging ability. J. Agric. Food Chem. 2016, 64, 7921–7930. [Google Scholar] [CrossRef]
- Xiao, Z.; Li, J.; Wang, H.; Zhang, Q.; Ge, Q.; Mao, J.; Sha, R. Hemicellulosic polysaccharides from bamboo leaves promoted by phosphotungstic acids and its attenuation of oxidative stress in HepG2 Cells. Front. Nutr. 2022, 9, 917432. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Wu, L.; Zhong, W.; Zheng, Q.; Tan, W.; Feng, K.; Feng, X.; Meng, F. Cellular antioxidant properties of ischnoderma resinosum polysaccharide. Molecules 2022, 27, 7717. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; He, T.; Mao, Y.; Mao, J.; Lai, X.; Tu, H.; Zhou, Y.; Sha, R. Changes in Physicochemical Properties, Metabolites and Antioxidant Activity of Edible Grass during Spontaneous Fermentation. Fermentation 2023, 9, 377. https://doi.org/10.3390/fermentation9040377
Li X, He T, Mao Y, Mao J, Lai X, Tu H, Zhou Y, Sha R. Changes in Physicochemical Properties, Metabolites and Antioxidant Activity of Edible Grass during Spontaneous Fermentation. Fermentation. 2023; 9(4):377. https://doi.org/10.3390/fermentation9040377
Chicago/Turabian StyleLi, Xianxiu, Tao He, Yangchen Mao, Jianwei Mao, Xiaojin Lai, Hangjia Tu, Yi Zhou, and Ruyi Sha. 2023. "Changes in Physicochemical Properties, Metabolites and Antioxidant Activity of Edible Grass during Spontaneous Fermentation" Fermentation 9, no. 4: 377. https://doi.org/10.3390/fermentation9040377
APA StyleLi, X., He, T., Mao, Y., Mao, J., Lai, X., Tu, H., Zhou, Y., & Sha, R. (2023). Changes in Physicochemical Properties, Metabolites and Antioxidant Activity of Edible Grass during Spontaneous Fermentation. Fermentation, 9(4), 377. https://doi.org/10.3390/fermentation9040377