
Bacterial Diversity Analysis of Chaozhou Sauerkraut Based on High-Throughput Sequencing of Different Production Methods

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction and PCR Amplification
2.3. Illumina NovaSeq Sequencing
2.4. Bioinformatics Analysis
2.5. Detection and Statistical Analysis of pH Value and Nitrite Content and Reducing Sugar Content
3. Results and Discussion
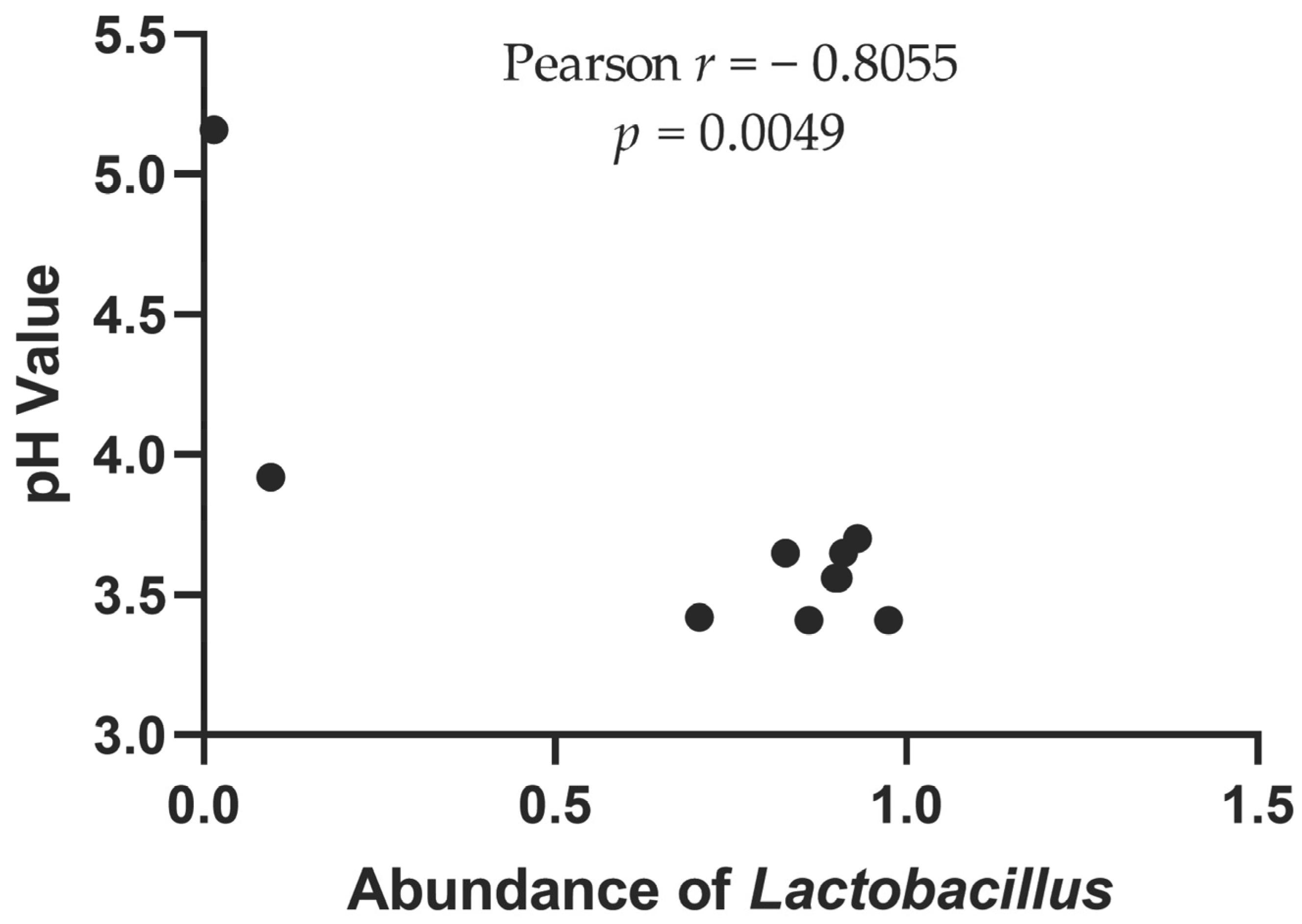
3.1. pH Value, Nitrite Content and Reducing Sugar Content
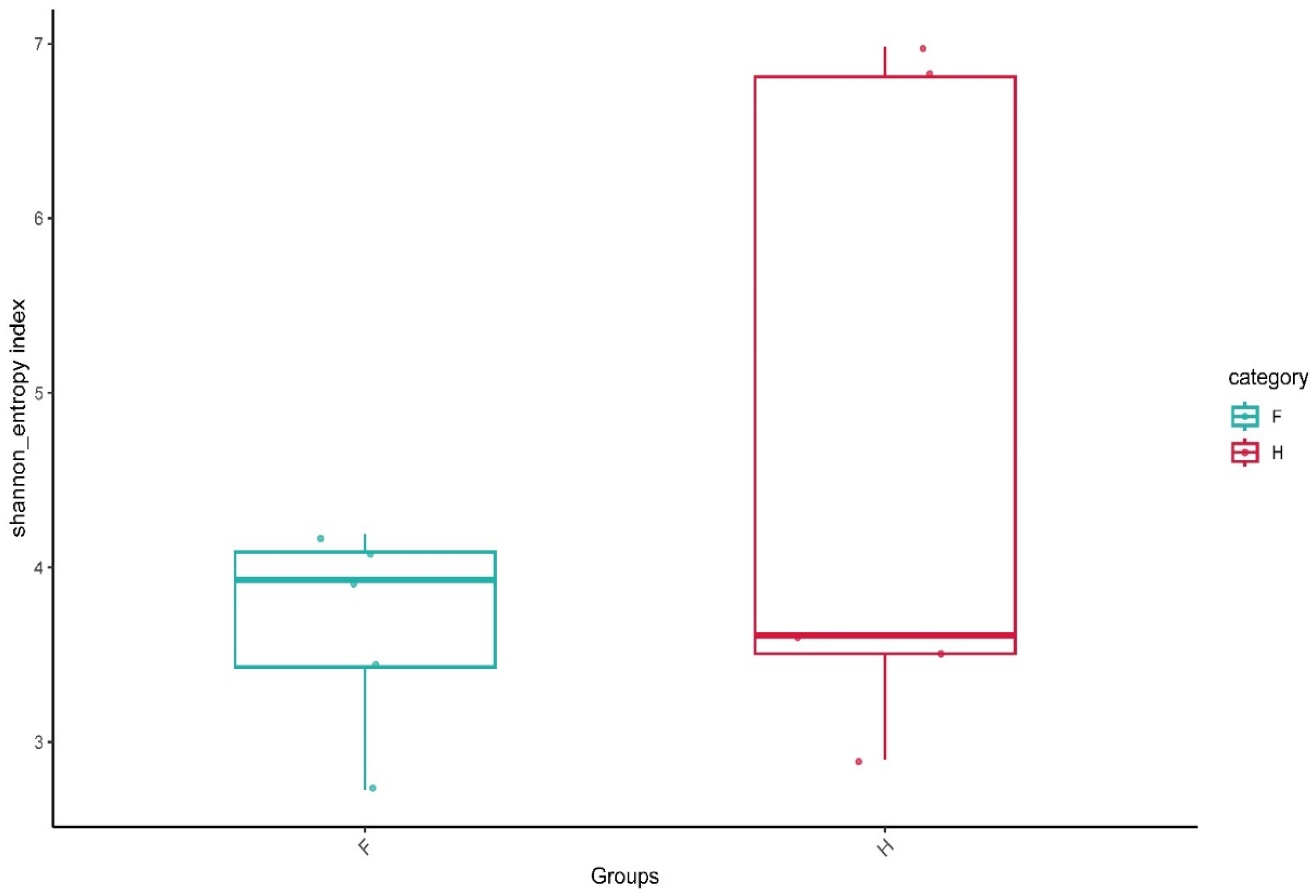
3.2. Statistical Analysis of the Alpha Diversity Index
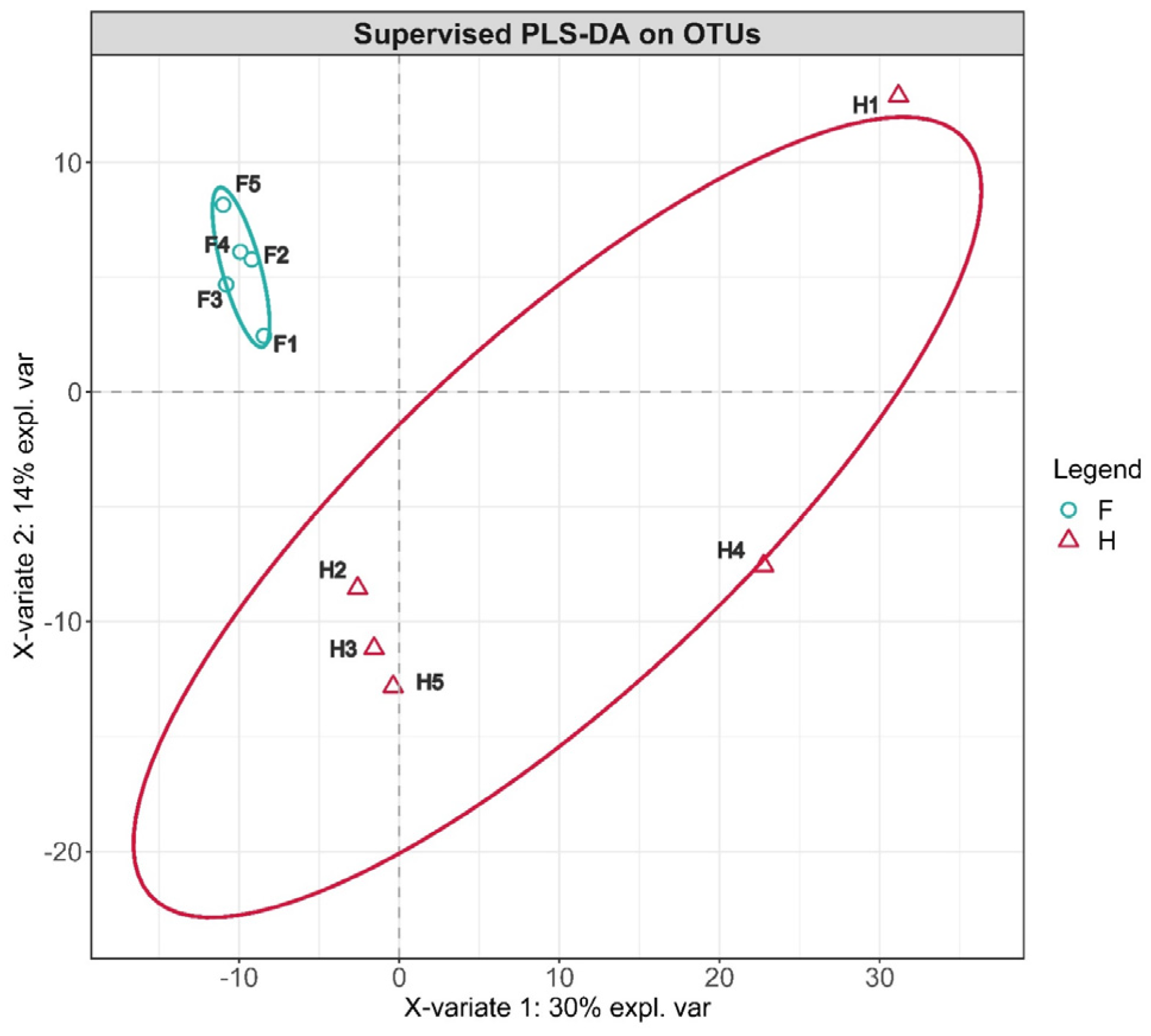
3.3. Beta Diversity Analysis
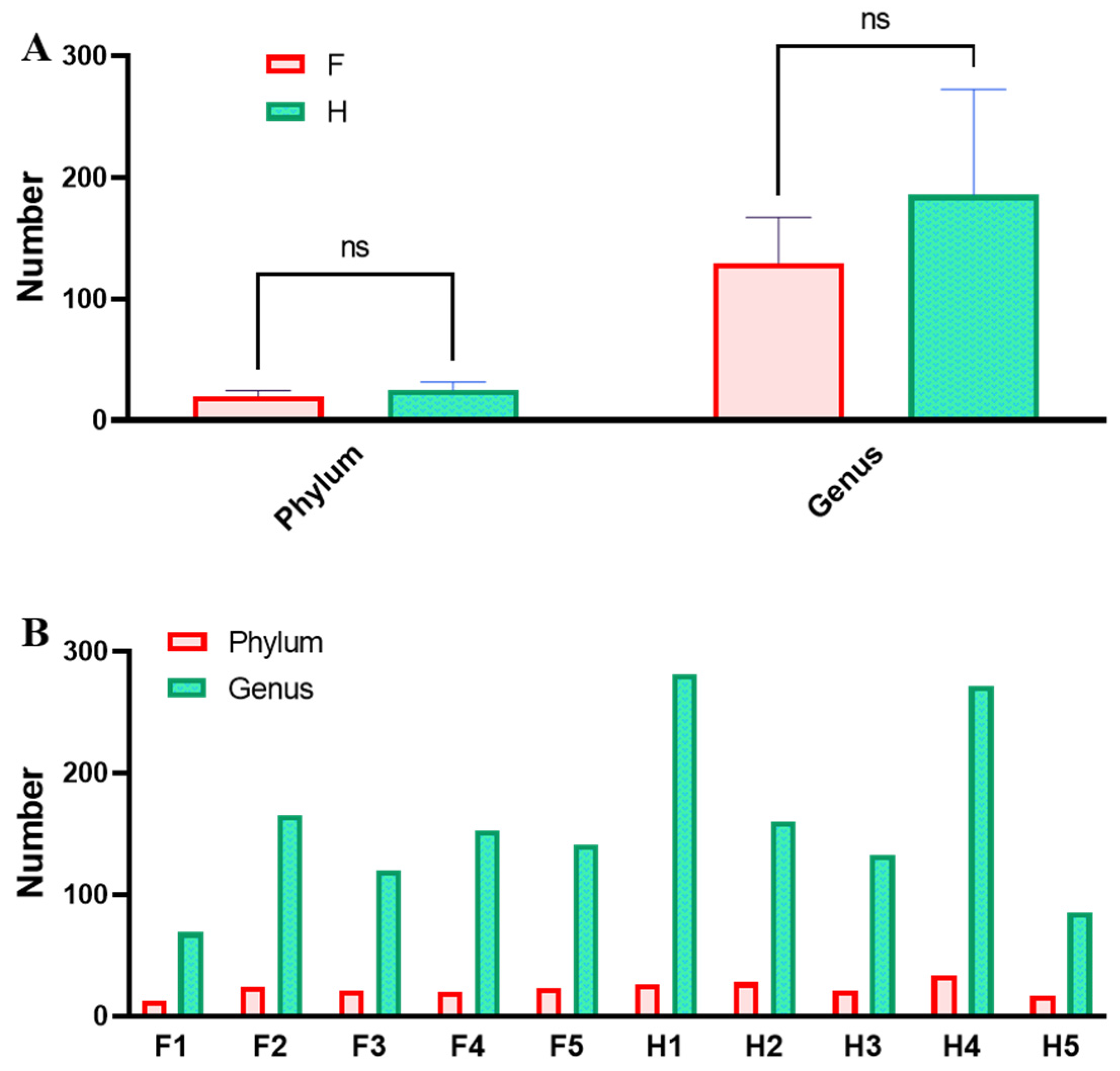
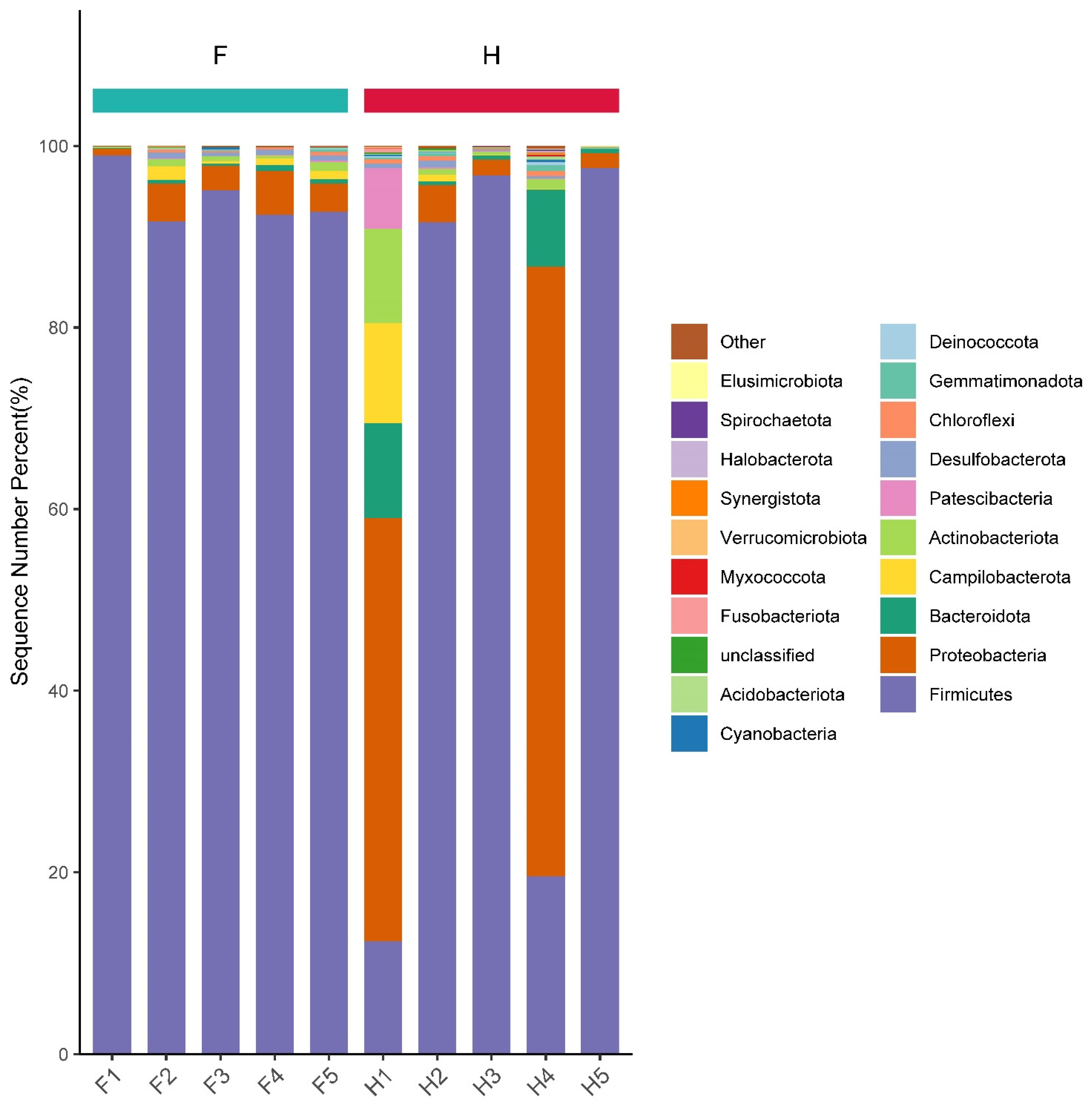
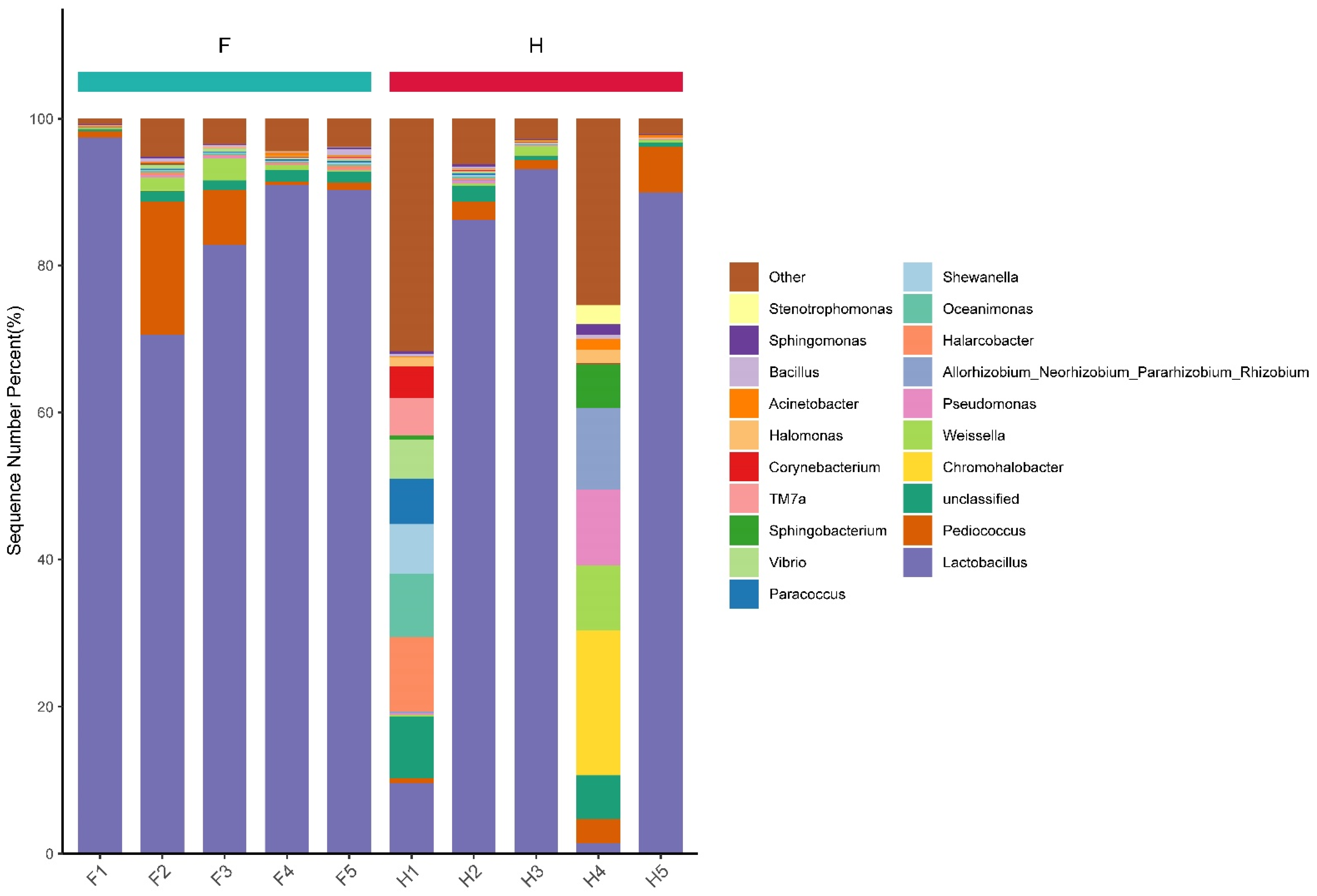
3.4. Analysis of Bacterial Community Structure Based on Phylum and Genus Levels
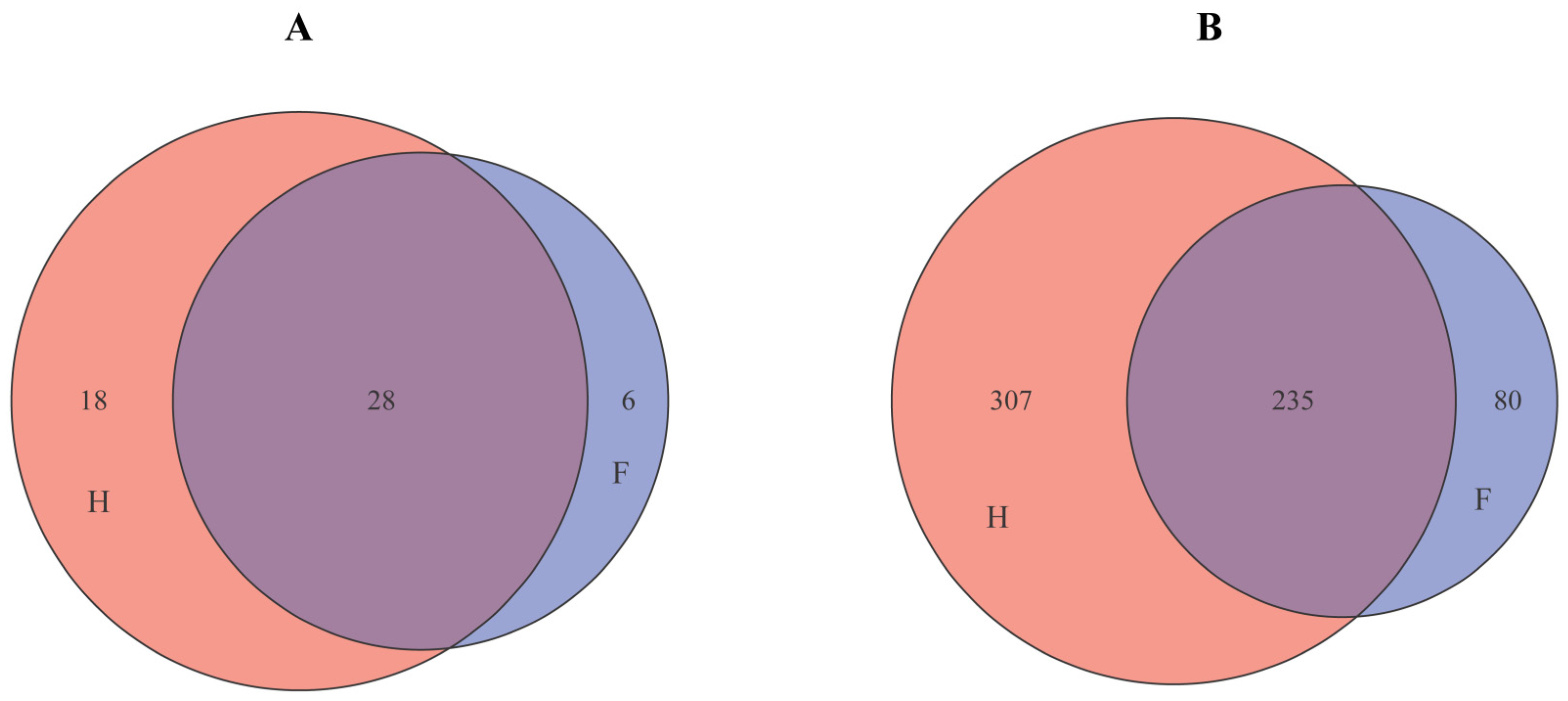
3.5. Cluster Analysis
3.6. Analysis of Dominant Bacteria Based on Phylum Level
3.7. Analysis of Dominant Bacteria Based on Genus Level
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rao, Y.; Chang, W.; Xiang, W.; Li, M.; Che, Z.; Tang, J. Screening and Performance of Lactobacillus plantarum E11 with Bacteriocin-Like Substance Secretion as Fermentation Starter of Sichuan Pickle. J. Food Saf. 2013, 33, 445–452. [Google Scholar] [CrossRef]
- Chen, X.; Zheng, M.; Liu, J.; Deng, Z.; Zhang, B.; Li, H. Selection and use of indigenous mixed starter cultures for mustard leaves fermentation and the improvement of cuocai characteristics. J. Sci. Food Agric. 2018, 98, 1773–1786. [Google Scholar] [CrossRef] [PubMed]
- Yoo, J.Y.; Cho, H.J.; Moon, S.; Choi, J.; Lee, S.; Ahn, C.; Yoo, K.Y.; Kim, I.; Ko, K.P.; Lee, J.E. Pickled Vegetable and Salted Fish Intake and the Risk of Gastric Cancer: Two Prospective Cohort Studies and a Meta-Analysis. Cancers 2020, 12, 996. [Google Scholar] [CrossRef] [PubMed]
- Frazie, M.D.; Kim, M.J.; Ku, K.M. Health-Promoting Phytochemicals from 11 Mustard Cultivars at Baby Leaf and Mature Stages. Molecules 2017, 22, 1749. [Google Scholar] [CrossRef]
- Park, S.Y.; Jang, H.L.; Lee, J.H.; Choi, Y.; Kim, H.; Hwang, J.; Seo, D.; Kim, S.; Nam, J.S. Changes in the phenolic compounds and antioxidant activities of mustard leaf (Brassica juncea) kimchi extracts during different fermentation periods. Food Sci. Biotechnol. 2017, 26, 105–112. [Google Scholar] [CrossRef]
- Di, H.; Ma, J.; Zhang, Y.; Wei, J.; Yang, J.; Ma, J.; Bian, J.; Xu, J.; Huang, Z.; Tang, Y. Correlations between flavor and glucosinolates and changes in quality-related physiochemical characteristics of Guizhou suancai during the fermentation process. Food Chem. 2023, 405, 134965. [Google Scholar] [CrossRef]
- Dimidi, E.; Cox, S.R.; Rossi, M.; Whelan, K. Fermented Foods: Definitions and Characteristics, Impact on the Gut Microbiota and Effects on Gastrointestinal Health and Disease. Nutrients 2019, 11, 1806. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligne, B.; Ganzle, M.; Kort, R.; Pasin, G.; Pihlanto, A. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, J.; Yi, R.; Zhou, X.; Long, X.; Pan, Y.; Zhao, X. Preventive Effect of Lactobacillus fermentum CQPC08 on 4-Nitroquineline-1-Oxide Induced Tongue Cancer in C57BL/6 Mice. Foods 2019, 8, 93. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, X.; Li, Y.; Sun, R.; Lin, Y.; Yu, H.; Xue, Y.; Zhou, X.; Liu, W.; Yan, L. The drivers of bacterial community underlying biogeographical pattern in Mollisol area of China. Ecotoxicol. Environ. Saf. 2019, 177, 93–99. [Google Scholar] [CrossRef]
- Zang, J.; Xu, Y.; Xia, W.; Yu, D.; Gao, P.; Jiang, Q.; Yang, F. Dynamics and diversity of microbial community succession during fermentation of Suan yu, a Chinese traditional fermented fish, determined by high throughput sequencing. Food Res. Int. 2018, 111, 565–573. [Google Scholar] [CrossRef]
- An, F.; Sun, H.; Wu, J.; Zhao, C.; Li, T.; Huang, H.; Fang, Q.; Mu, E.; Wu, R. Investigating the core microbiota and its influencing factors in traditional Chinese pickles. Food Res. Int. 2021, 147, 110543. [Google Scholar] [CrossRef]
- Rao, Y.; Qian, Y.; Tao, Y.; She, X.; Li, Y.; Chen, X.; Guo, S.; Xiang, W.; Liu, L.; Du, H. Characterization of the microbial communities and their correlations with chemical profiles in assorted vegetable Sichuan pickles. Food Control 2020, 113, 107174. [Google Scholar] [CrossRef]
- Yang, X.; Hu, W.; Xiu, Z.; Jiang, A.; Yang, X.; Sarengaowa; Ji, Y.; Guan, Y.; Feng, K. Comparison of northeast sauerkraut fermentation between single lactic acid bacteria strains and traditional fermentation. Food Res. Int. 2020, 137, 109553. [Google Scholar] [CrossRef]
- Ma, Q.; Zhu, Y.T.; Li, Y.D.; Zhang, Z.L.; Huang, J.; Zuo, Y. Quantification of heavy metals and health risk assessment in Sichuan pickle. J. Food Sci. 2022, 87, 2229–2244. [Google Scholar] [CrossRef]
- Pinaev, A.G.; Kichko, A.A.; Aksenova, T.S.; Safronova, V.I.; Kozhenkova, E.V.; Andronov, E.E. RIAM: A Universal Accessible Protocol for the Isolation of High Purity DNA from Various Soils and Other Humic Substances. Methods Protoc. 2022, 5, 99. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Mandal, S.; Van Treuren, W.; White, R.A.; Eggesbo, M.; Knight, R.; Peddada, S.D. Analysis of composition of microbiomes: A novel method for studying microbial composition. Microb. Ecol. Health Dis. 2015, 26, 27663. [Google Scholar] [CrossRef]
- Vázquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. Gigascience 2013, 2, 16. [Google Scholar] [CrossRef] [PubMed]
- Rohart, F.; Gautier, B.; Singh, A.; Le Cao, K.A. mixOmics: An R package for ’omics feature selection and multiple data integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, D.; Tian, J.; Sun, T.; Meng, Q.; Li, J.; Shan, A. The responses of organic acid production and microbial community to different carbon source additions during the anaerobic fermentation of Chinese cabbage waste. Bioresour. Technol. 2023, 371, 128624. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Peng, Z.; Huang, T.; Guan, Q.; Li, J.; Xie, M.; Xiong, T. Bacterial community dynamics and physicochemical characteristics in natural fermentation of jiang-shui, a traditional food made in northwest China. J. Sci. Food Agric. 2019, 99, 3391–3397. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, J.; Wei, B.; Huang, T.; Xiao, Y.; Peng, Z.; Xie, M.; Xiong, T. Bacterial community and composition in Jiang-shui and Suan-cai revealed by high-throughput sequencing of 16S rRNA. Int. J. Food Microbiol. 2019, 306, 108271. [Google Scholar] [CrossRef]
- Sun, X.H.; Qi, X.; Han, Y.D.; Guo, Z.J.; Cui, C.B.; Lin, C.Q. Characteristics of changes in volatile organic compounds and microbial communities during the storage of pickles. Food Chem. 2023, 409, 135285. [Google Scholar] [CrossRef]
- Wang, C.; Du, X.; Xie, T.; Li, H. Label- and modification-free-based in situ selection of bovine serum albumin specific aptamer. J. Sep. Sci. 2019, 42, 3571–3578. [Google Scholar] [CrossRef]
- Chien, H.-I.; Yen, Y.-F.; Lee, Y.-C.; Wei, P.-C.; Huang, C.-Y.; Tseng, C.-H.; Yen, F.-L.; Tsai, Y.-H. Determination of the Bacterial Community of Mustard Pickle Products and Their Microbial and Chemical Qualities. Biology 2023, 12, 258. [Google Scholar] [CrossRef]
- Chen, C.; Liu, Y.; Tian, H.; Ai, L.; Yu, H. Metagenomic analysis reveals the impact of JIUYAO microbial diversity on fermentation and the volatile profile of Shaoxing-jiu. Food Microbiol. 2020, 86, 103326. [Google Scholar] [CrossRef]
- He, Z.; Chen, H.; Wang, X.; Lin, X.; Ji, C.; Li, S.; Liang, H. Effects of different temperatures on bacterial diversity and volatile flavor compounds during the fermentation of suancai, a traditional fermented vegetable food from northeastern China. LWT 2020, 118, 108773. [Google Scholar] [CrossRef]
- Kwon, E.A.; Kim, M. Microbial evaluation of commercially packed Kimchi products. Food Sci. Biotechnol. 2007, 16, 615–620. [Google Scholar]
- Gu, C.T.; Li, C.Y.; Yang, L.J.; Huo, G.C. Lactobacillus heilongjiangensis sp. nov., isolated from Chinese pickle. Int. J. Syst. Evol. Microbiol. 2013, 63, 4094–4099. [Google Scholar] [CrossRef]
- Karruli, A.; Catalini, C.; D’Amore, C.; Foglia, F.; Mari, F.; Harxhi, A.; Galdiero, M.; Durante-Mangoni, E. Evidence-Based Treatment of Pseudomonas aeruginosa Infections: A Critical Reappraisal. Antibiotics 2023, 12, 399. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Label | pH Value | Nitrite Content (mg/kg) | Reducing Sugar Content (mg/mL) |
|---|---|---|---|
| F1 | 3.41 | 0.66 | 0.0976 |
| F2 | 3.42 | 0.12 | 0.0980 |
| F3 | 3.65 | 0.24 | 0.0976 |
| F4 | 3.65 | 0.35 | 0.0981 |
| F5 | 3.56 | 0.38 | 0.0985 |
| F group | 3.54 ± 0.12 a | 0.35 ± 0.2 a | 0.098 ± 0.0004 a |
| H1 | 3.92 | 0.15 | 0.1022 |
| H2 | 3.41 | 0.04 | 0.1945 |
| H3 | 3.7 | 0.31 | 0.1020 |
| H4 | 5.16 | 1.12 | 0.8639 |
| H5 | 3.56 | 0.09 | 0.1015 |
| H group | 3.95 ± 0.7 a | 0.34 ± 0.44 a | 0.273 ± 0.3329 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, W.; Peng, H.; Chen, J.; Yan, X.; Zhang, Y. Bacterial Diversity Analysis of Chaozhou Sauerkraut Based on High-Throughput Sequencing of Different Production Methods. Fermentation 2023, 9, 282. https://doi.org/10.3390/fermentation9030282
Huang W, Peng H, Chen J, Yan X, Zhang Y. Bacterial Diversity Analysis of Chaozhou Sauerkraut Based on High-Throughput Sequencing of Different Production Methods. Fermentation. 2023; 9(3):282. https://doi.org/10.3390/fermentation9030282
Chicago/Turabian StyleHuang, Wuying, Heng Peng, Junsheng Chen, Xiantao Yan, and Yanyan Zhang. 2023. "Bacterial Diversity Analysis of Chaozhou Sauerkraut Based on High-Throughput Sequencing of Different Production Methods" Fermentation 9, no. 3: 282. https://doi.org/10.3390/fermentation9030282
APA StyleHuang, W., Peng, H., Chen, J., Yan, X., & Zhang, Y. (2023). Bacterial Diversity Analysis of Chaozhou Sauerkraut Based on High-Throughput Sequencing of Different Production Methods. Fermentation, 9(3), 282. https://doi.org/10.3390/fermentation9030282