
Sulphate Uptake Plays a Major Role in the Production of Sulphur Dioxide by Yeast Cells during Oenological Fermentations
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Media
2.2. Bacterial Strains and Media
2.3. Plasmids Construction
2.4. Yeast Genetic Modification
2.5. Yeast Functional Assays
2.6. Fermentation Assays
2.7. Chemical Analyses
2.8. Statistical Analysis
3. Results
3.1. Construction of CRISPR/Cas9 Tools for Genome Editing of Natural Yeast Strains
3.2. Yeast Genome Editing
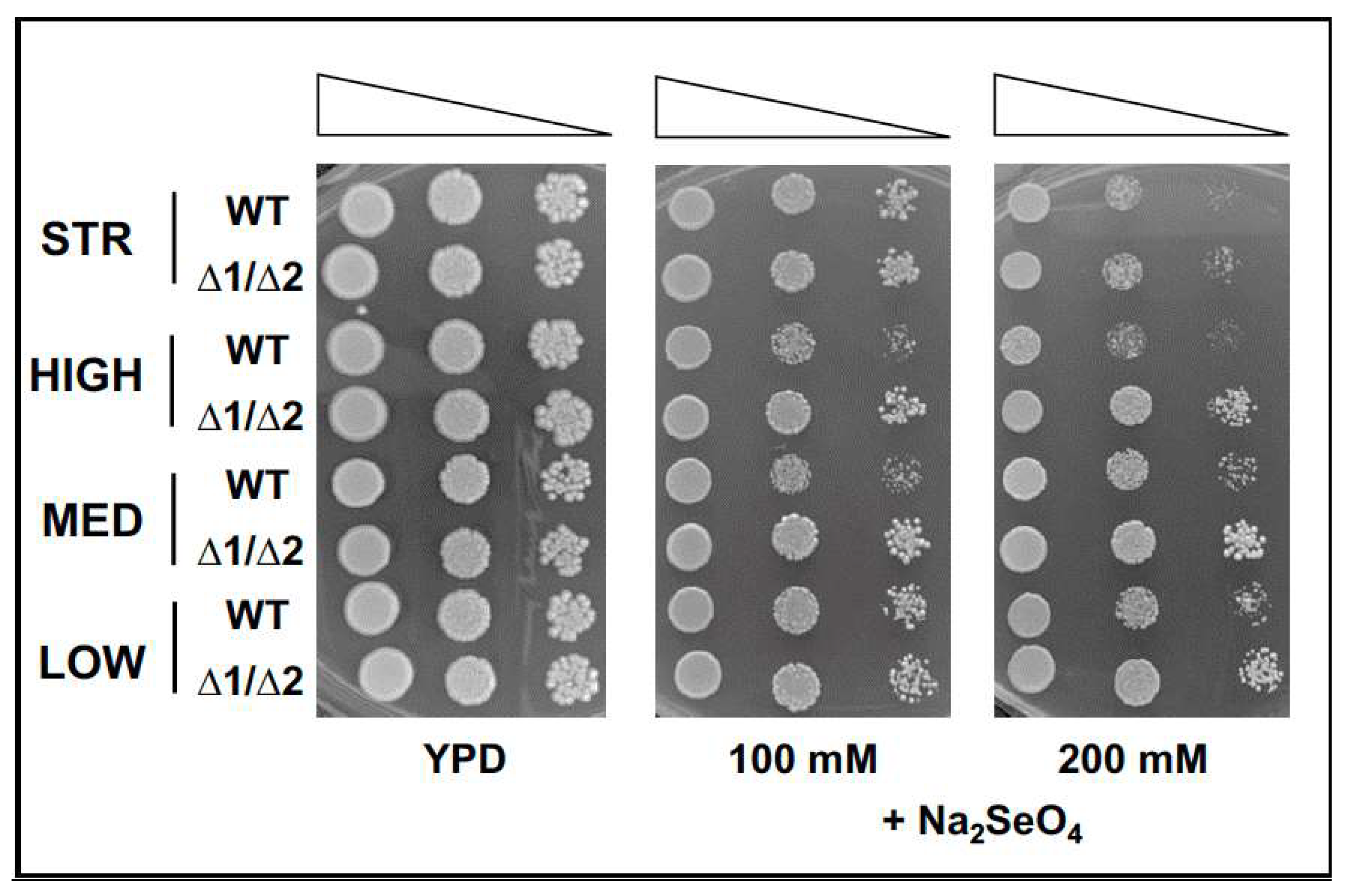
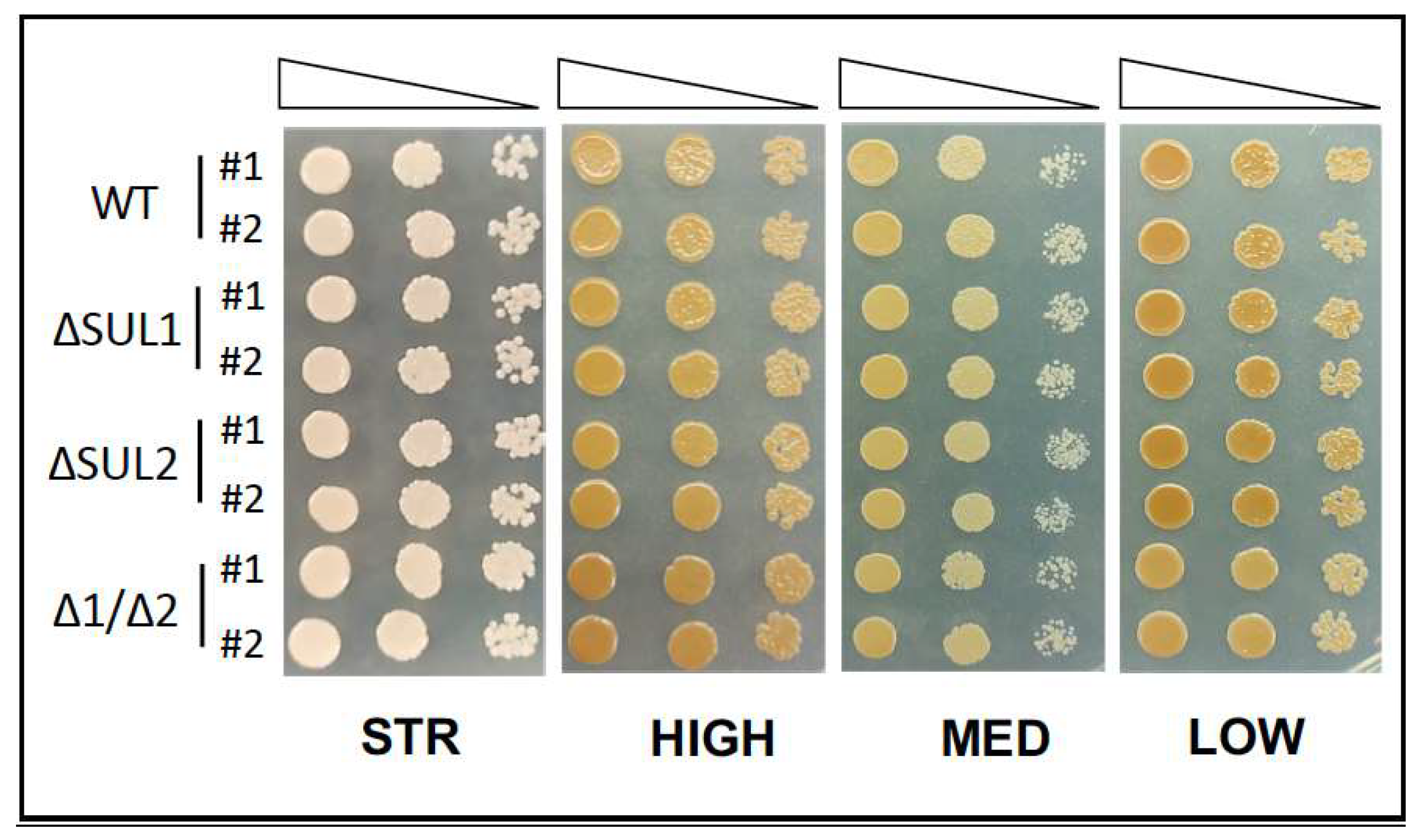
3.3. Functional Characterization of Yeast Mutant Strains
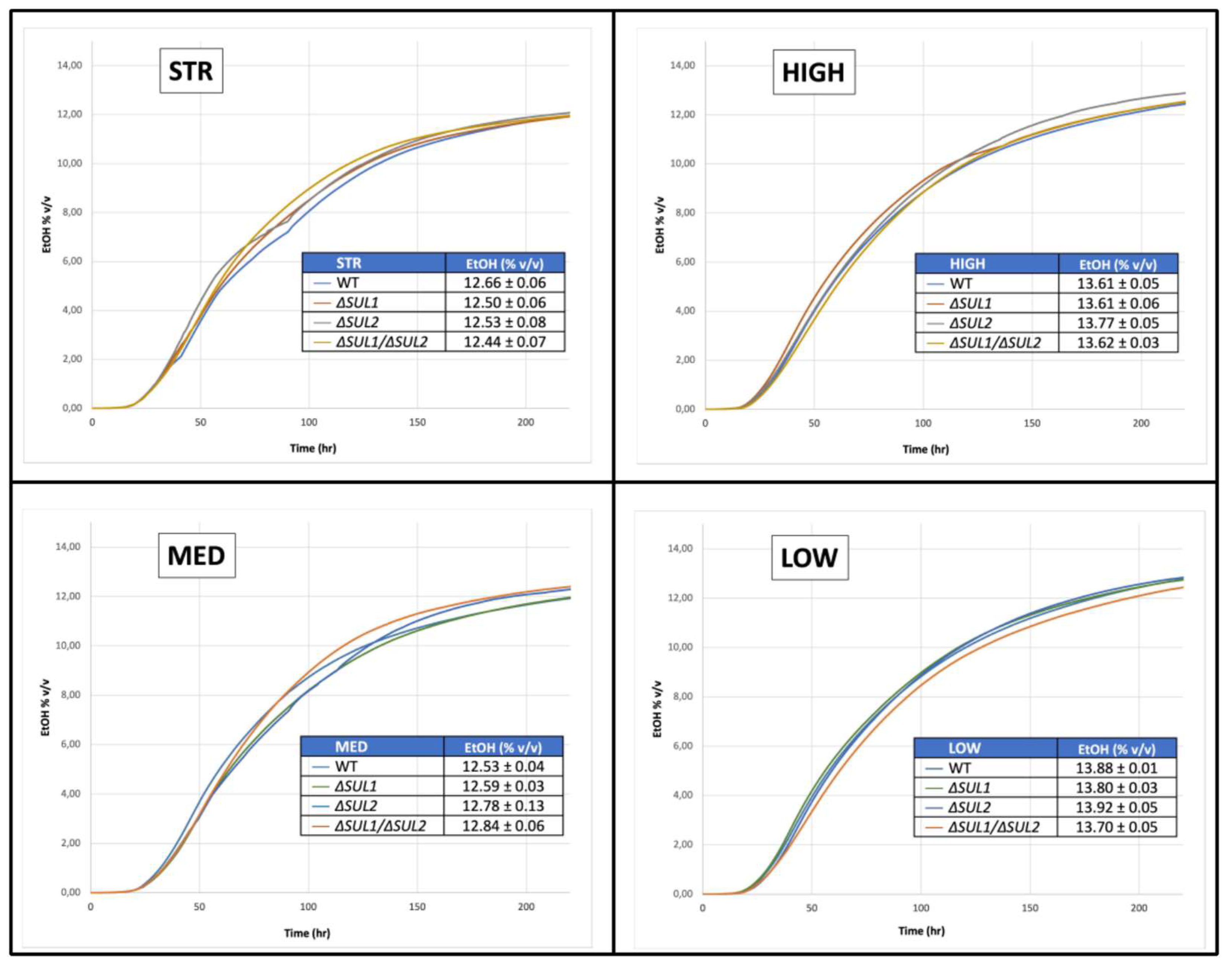
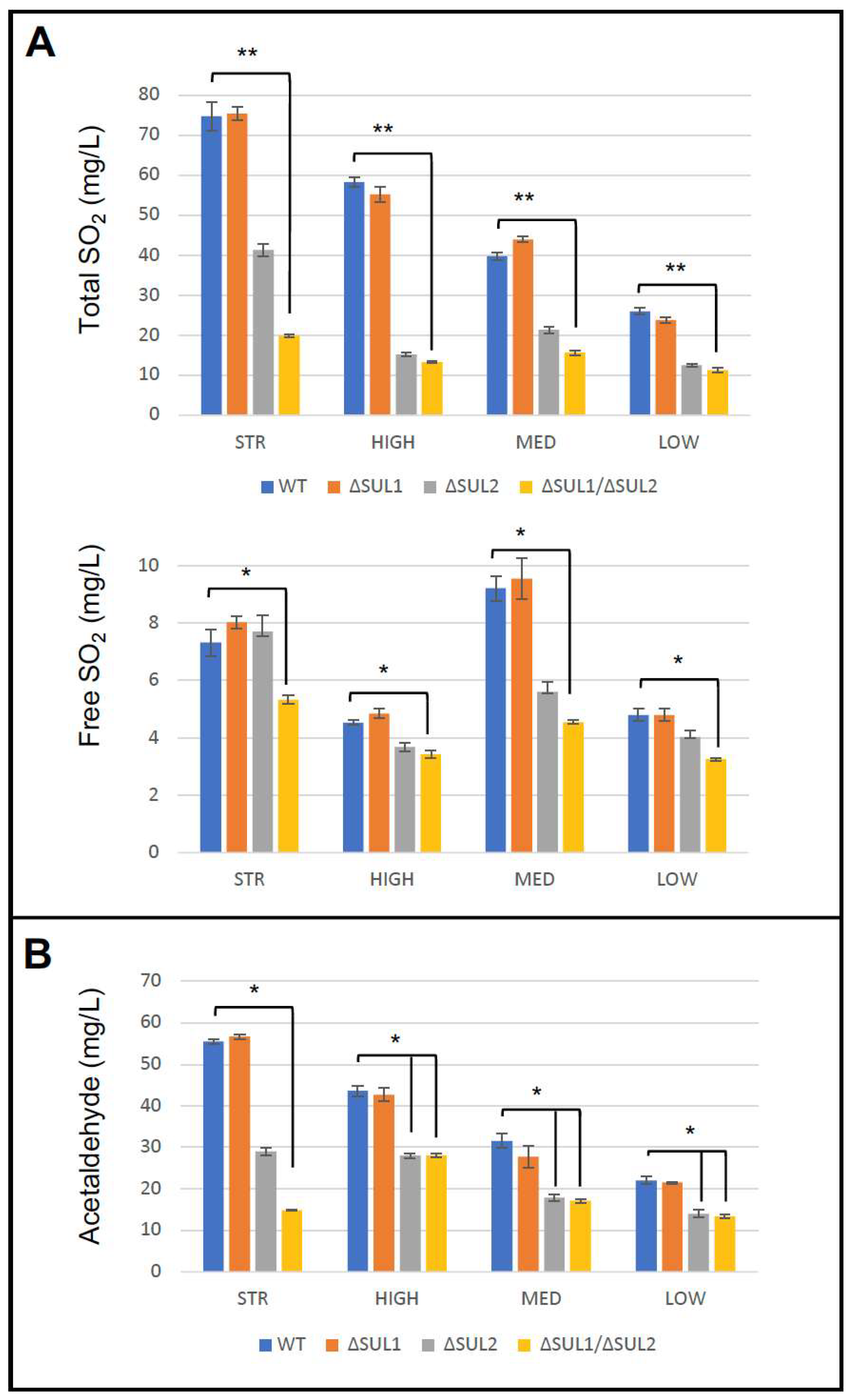
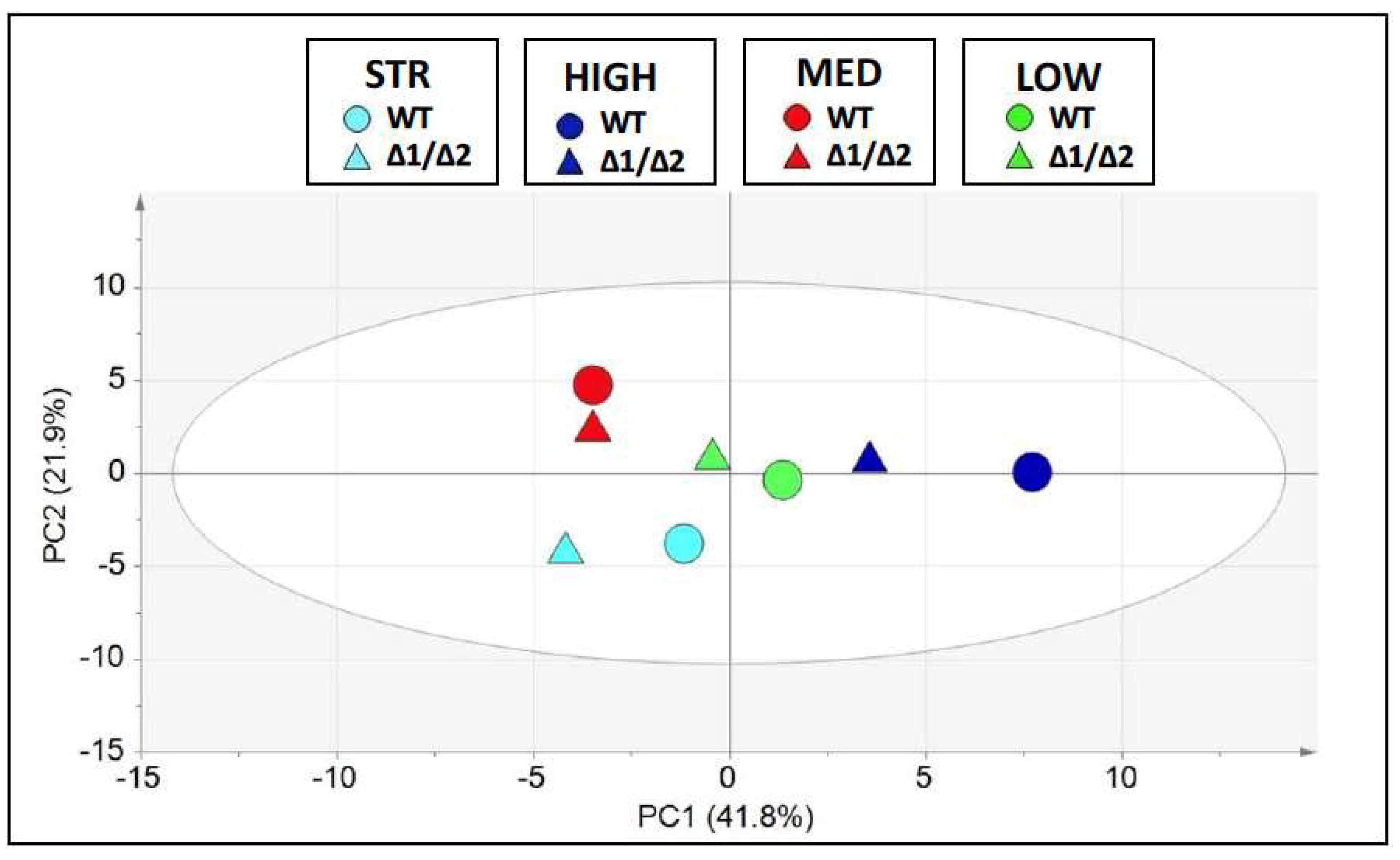
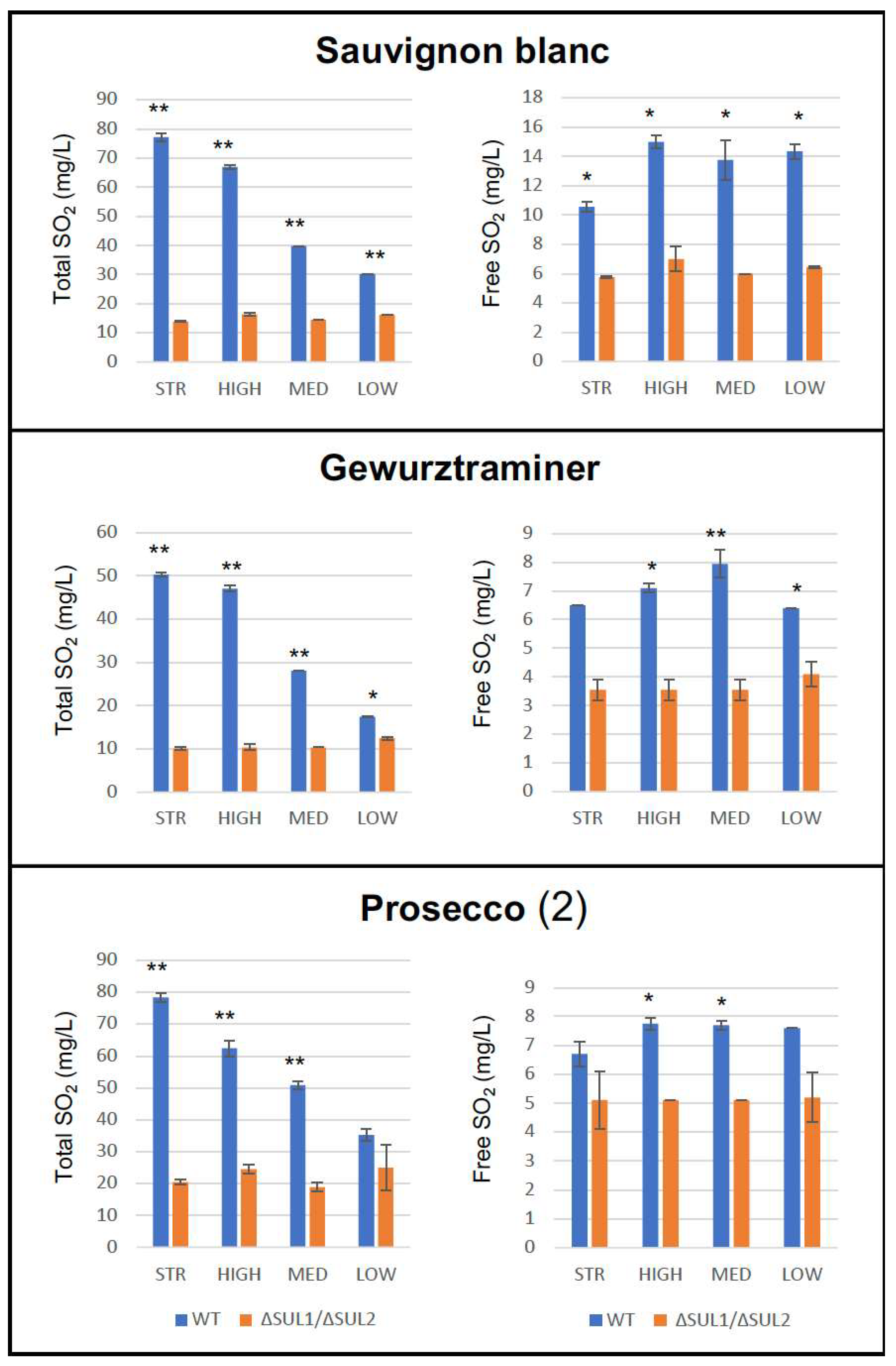
3.4. Fermentation Assays in Natural Must
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blouin, J. Le SO2 en Oenologie; Dunod: Paris, France, 2014; ISBN 978-2-10-071956-3. [Google Scholar]
- Ribereau-Gayon, P.; Dubourdieu, D.; Doneche, B.; Lonvaud, A. Handbook of Enology: The Microbiology of Wine and Vinifications; John Wiley & Sons Ltd.: Chichester, UK, 2006. [Google Scholar]
- Danilewicz, J.C. Review of reaction mechanisms of oxygen and proposed intermediate reaction products in wine: Central role of iron and copper. Am. J. Enol. Vitic. 2003, 54, 73–85. [Google Scholar] [CrossRef]
- Burroughs, L.F.; Sparks, A.H. Sulphite-binding power of wines and ciders. I. Equilibrium constants for the dissociation of carbonyl bisulphite compounds. J. Sci. Food Agric. 1973, 24, 187–198. [Google Scholar] [CrossRef]
- UE. The Maximum Sulphur Dioxide Content of Wines; Official Journal of the European Union, L149 Annex I B.; UE: Brussels, Belgium, 2019; pp. 32–36. [Google Scholar]
- Komarnisky, L.A.; Christopherson, R.J.; Basu, T.K. Sulfur: Its clinical and toxicologic aspects. Nutrition 2003, 19, 54–61. [Google Scholar] [CrossRef]
- Vally, H.; Misso, N.L.A.; Madan, V. Clinical effects of sulphite additives. Clin. Exp. Allergy 2009, 39, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Swiegers, J.H.; Bartowsky, E.J.; Henschke, P.A.; Pretorius, I.S. Yeast and bacterial modulation of wine aroma and flavour. Aust. J. Grape Wine Res. 2005, 11, 139–173. [Google Scholar] [CrossRef]
- Thomas, D.; Surdin-Kerjan, Y. Metabolism of sulfur amino acids in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 1997, 61, 503–532. [Google Scholar] [PubMed]
- González Hernández, G.; Hardisson de La Torre, A.; José Arias León, J. Boron, sulphate, chloride and phosphate contents in musts and wines of the Tacoronte-Acentejo D.O.C. region (Canary Islands). Food Chem. 1997, 60, 339–345. [Google Scholar] [CrossRef]
- Amerine, M.A.; Berg, H.W.; Kunkee, R.E.; Ough, C.S.; Singleton, V.L.; Webb, A.D. The composition of grapes. In The Technology of Wine Making, 4th ed.; Company, A.P.: Westport, CT, USA, 1980; pp. 77–139. [Google Scholar]
- Leske, P.A.; Sas, A.N.; Coulter, A.D.; Stockley, C.S.; Lee, T.H. The composition of Australian grape juice: Chloride, sodium and sulfate ions. Aust. J. Grape Wine Res. 1997, 3, 26–30. [Google Scholar] [CrossRef]
- Cherest, H.; Davidian, J.-C.; Thomas, D.; Benes, V.; Ansorge, W.; Surdin-Kerjan, Y. Molecular Characterization of Two High Affinity Sulfate Transporters in Saccharomyces cerevisiae. Genetics 1997, 145, 627–635. [Google Scholar] [CrossRef]
- Kankipati, H.N.; Rubio-Texeira, M.; Castermans, D.; Diallinas, G.; Thevelein, J.M. Sul1 and Sul2 Sulfate Transceptors Signal to Protein Kinase A upon Exit of Sulfur Starvation. J. Biol. Chem. 2015, 290, 10430–10446. [Google Scholar] [CrossRef]
- Smith, F.W.; Hawkesford, M.J.; Prosser, I.M.; Clarkson, D.T. Isolation of a cDNA from Saccharomyces cerevisiae that encodes a high affinity sulphate transporter at the plasma membrane. Mol. Gen. Genet. MGG 1995, 247, 709–715. [Google Scholar] [CrossRef]
- Holt, S.; Kankipati, H.; De Graeve, S.; Van Zeebroeck, G.; Foulquié-Moreno, M.R.; Lindgreen, S.; Thevelein, J.M. Major sulfonate transporter Soa1 in Saccharomyces cerevisiae and considerable substrate diversity in its fungal family. Nat. Commun. 2017, 8, 14247. [Google Scholar] [CrossRef]
- Donalies, U.E.B.; Stahl, U. Increasing sulphite formation inSaccharomyces cerevisiae by overexpression ofMET14 andSSU1. Yeast 2002, 19, 475–484. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Modulation of volatile sulfur compounds by wine yeast. Appl. Microbiol. Biotechnol. 2007, 74, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-W.; Walker, M.E.; Fedrizzi, B.; Gardner, R.C.; Jiranek, V. Hydrogen sulfide and its roles in Saccharomyces cerevisiae in a winemaking context. FEMS Yeast Res. 2017, 17, fox058. [Google Scholar] [CrossRef]
- Eschenbruch, R.; Bonish, P. Production of sulphite and sulphide by low-and high-sulphite forming wine yeasts. Arch. Microbiol. 1976, 107, 299–302. [Google Scholar] [CrossRef]
- Ugliano, M.; Kwiatkowski, M.; Vidal, S.; Capone, D.; Siebert, T.; Dieval, J.-B.; Aagaard, O.; Waters, E.J. Evolution of 3-Mercaptohexanol, Hydrogen Sulfide, and Methyl Mercaptan during Bottle Storage of Sauvignon blanc Wines. Effect of Glutathione, Copper, Oxygen Exposure, and Closure-Derived Oxygen. J. Agric. Food Chem. 2011, 59, 2564–2572. [Google Scholar] [CrossRef]
- Ugliano, M.; Fedrizzi, B.; Siebert, T.; Travis, B.; Magno, F.; Versini, G.; Henschke, P.A. Effect of Nitrogen Supplementation and Saccharomyces Species on Hydrogen Sulfide and Other Volatile Sulfur Compounds in Shiraz Fermentation and Wine. J. Agric. Food Chem. 2009, 57, 4948–4955. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-Q.; Pilone, G.J. An overview of formation and roles of acetaldehyde in winemaking with emphasis on microbiological implications. Int. J. Food Sci. Technol. 2000, 35, 49–61. [Google Scholar] [CrossRef]
- Ochando, T.; Mouret, J.-R.; Humbert-Goffard, A.; Aguera, E.; Sablayrolles, J.-M.; Farines, V. Comprehensive study of the dynamic interaction between SO2 and acetaldehyde during alcoholic fermentation. Food Res. Int. 2020, 136, 109607. [Google Scholar] [CrossRef]
- Linderholm, A.; Dietzel, K.; Hirst, M.; Bisson, L.F. Identification of MET10—932 and Characterization as an Allele Reducing Hydrogen Sulfide Formation in Wine Strains of Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2010, 76, 7699–7707. [Google Scholar] [CrossRef]
- Cordente, A.G.; Heinrich, A.; Pretorius, I.S.; Swiegers, J.H. Isolation of sulfite reductase variants of a commercial wine yeast with significantly reduced hydrogen sulfide production. FEMS Yeast Res. 2009, 9, 446–459. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Canonico, L.; Comitini, F.; Ciani, M. Reduction of Sulfur Compounds through Genetic Improvement of Native Saccharomyces cerevisiae Useful for Organic and Sulfite-Free Wine. Foods 2020, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- Mezzetti, F.; De Vero, L.; Giudici, P. Evolved Saccharomyces cerevisiae wine strains with enhanced glutathione production obtained by an evolution-based strategy. FEMS Yeast Res. 2014, 14, 977–987. [Google Scholar] [CrossRef]
- Wysocki, R.; Tamás, M.J. How Saccharomyces cerevisiae copes with toxic metals and metalloids. FEMS Microbiol. Rev. 2010, 34, 925–951. [Google Scholar] [CrossRef] [PubMed]
- De Vero, L.; Solieri, L.; Giudici, P. Evolution-based strategy to generate non-genetically modified organisms Saccharomyces cerevisiae strains impaired in sulfate assimilation pathway. Lett. Appl. Microbiol. 2011, 53, 572–575. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.E.; Zhang, J.; Sumby, K.M.; Lee, A.; Houlès, A.; Li, S.; Jiranek, V. Sulfate transport mutants affect hydrogen sulfide and sulfite production during alcoholic fermentation. Yeast 2021, 38, 367–381. [Google Scholar] [CrossRef]
- Basile, A.; De Pascale, F.; Bianca, F.; Rossi, A.; Frizzarin, M.; De Bernardini, N.; Bosaro, M.; Baldisseri, A.; Antoniali, P.; Lopreiato, R.; et al. Large-scale sequencing and comparative analysis of oenological Saccharomyces cerevisiae strains supported by nanopore refinement of key genomes. Food Microbiol. 2021, 97, 103753. [Google Scholar] [CrossRef]
- Mans, R.; van Rossum, H.M.; Wijsman, M.; Backx, A.; Kuijpers, N.G.A.; van den Broek, M.; Daran-Lapujade, P.; Pronk, J.T.; van Maris, A.J.A.; Daran, J.M.G. CRISPR/Cas9: A molecular Swiss army knife for simultaneous introduction of multiple genetic modifications in Saccharomyces cerevisiae. FEMS Yeast Res. 2015, 15, fov004. [Google Scholar] [CrossRef]
- Amberg, D.C.; Burke, D.; Strathern, J.N.; Burke, D.; Laboratory, C.S.H. Methods in Yeast Genetics: A Cold Spring Harbor Laboratory Course Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2005; ISBN 0879697288 9780879697280. [Google Scholar]
- Vokes, M.; Carpenter, A. Using CellProfiler for Automatic Identification and Measurement of Biological Objects in Images. Curr. Protoc. Mol. Biol. 2008, 82, 14.17.1–14.17.12. [Google Scholar] [CrossRef]
- Gietz, R.D.; Woods, R.A. Yeast transformation by the LiAc/SS Carrier DNA/PEG method. Methods Mol. Biol. 2006, 313, 107–120. [Google Scholar]
- Legras, J.-L.; Karst, F. Optimisation of interdelta analysis for Saccharomyces cerevisiae strain characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef]
- Mirisola, M.G.; Braun, R.J.; Petranovic, D. Approaches to study yeast cell aging and death. FEMS Yeast Res. 2014, 14, 109–118. [Google Scholar] [CrossRef]
- Iland, P.; Bruer, N.; Edwards, G.; Caloghiris, S.; Wilkes, E. Chemical Analysis of Grapes and Wine: Techniques and Concepts, 2nd ed.; Patrick Iland Wine Promotions Pty Ltd.: Adelaide, SA, Australia, 2013; ISBN 0958160575. [Google Scholar]
- Pereira, Y.; Lagniel, G.; Godat, E.; Baudouin-Cornu, P.; Junot, C.; Labarre, J. Chromate Causes Sulfur Starvation in Yeast. Toxicol. Sci. 2008, 106, 400–412. [Google Scholar] [CrossRef]
- Jennings, M.L.; Cui, J. Inactivation of Saccharomyces cerevisiae Sulfate Transporter Sul2p: Use It and Lose It. Biophys. J. 2012, 102, 768–776. [Google Scholar] [CrossRef]
- Jackowetz, J.N.; Dierschke, S.; Mira de Orduña, R. Multifactorial analysis of acetaldehyde kinetics during alcoholic fermentation by Saccharomyces cerevisiae. Food Res. Int. 2011, 44, 310–316. [Google Scholar] [CrossRef]
- Comuzzo, P.; Zironi, R. Biotechnological Strategies for Controlling Wine Oxidation. Food Eng. Rev. 2013, 5, 217–229. [Google Scholar] [CrossRef]
- Lisanti, M.T.; Blaiotta, G.; Nioi, C.; Moio, L. Alternative Methods to SO2 for Microbiological Stabilization of Wine. Compr. Rev. Food Sci. Food Saf. 2019, 18, 455–479. [Google Scholar] [CrossRef] [PubMed]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romano, P. Biotechnological Approach Based on Selected Saccharomyces cerevisiae Starters for Reducing the Use of Sulfur Dioxide in Wine. Microorganisms 2020, 8, 738. [Google Scholar] [CrossRef] [PubMed]
- Piłsyk, S.; Paszewski, A. Sulfate permeases phylogenetic diversity of sulfate transport. Acta Biochim. Pol. 2009, 56, 375–384. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Residual Glucose | Residual Fructose | Glycerol | Acetic Acid | Citric Acid | Tartaric Acid | Malic Acid | Succinic Acid |
|---|---|---|---|---|---|---|---|---|
| (g/L) | (g/L) | (g/L) | (g/L) | (g/L) | (g/L) | (g/L) | (g/L) | |
| STR | ||||||||
| WT | 0.13 ± 0.01 | 7.27 ± 0.18 | 5.48 ± 0.03 | 0.14 ± 0.01 | 0.14 ± 0.01 | 1.93 ± 0.01 | 3.61 ± 0.03 | 0.92 ± 0.02 |
| DSUL1 | 0.22 ± 0.02 | 8.75 ± 0.36 | 5.55 ± 0.04 | 0.14 ± 0.02 | 0.15 ± 0.00 | 1.95 ± 0.01 | 3.67 ± 0.05 | 0.94 ± 0.01 |
| DSUL2 | 0.10 ± 0.09 | 6.75 ± 1.70 | 5.64 ± 0.04 | 0.13 ± 0.03 | 0.13 ± 0.02 | 1.94 ± 0.05 | 3.69 ± 0.06 | 0.97 ± 0.02 |
| DSUL1 /DSUL2 | 0.15 ± 0.07 | 7.37 ± 1.00 | 5.64 ± 0.03 | 0.13 ± 0.01 | 0.18 ± 0.01 | 2.01 ± 0.03 | 3.66 ± 0.07 | 0.99 ± 0.02 |
| HIGH | ||||||||
| WT | 0.23 ± 0.04 | 6.68 ± 0.21 | 5.74 ± 0.05 | 0.38 ± 0.01 | 0.18 ± 0.02 | 2.64 ± 0.04 | 3.62 ± 0.06 | 0.85 ± 0.01 |
| DSUL1 | 0.22 ± 0.04 | 6.64 ± 0.21 | 5.78 ± 0.03 | 0.39 ± 0.01 | 0.17 ± 0.00 | 2.62 ± 0.01 | 3.57 ± 0.04 | 0.88 ± 0.02 |
| DSUL2 | 0.13 ± 0.00 | 5.44 ± 0.52 | 5.87 ± 0.02 | 0.48 ± 0.03 | 0.16 ± 0.00 | 2.63 ± 0.01 | 3.34 ± 0.05 | 0.88 ± 0.01 |
| DSUL1 /DSUL2 | 0.32 ± 0.04 | 7.88 ± 1.22 | 5.82 ± 0.02 | 0.46 ± 0.02 | 0.18 ± 0.02 | 2.64 ± 0.04 | 3.30 ± 0.01 | 0.87 ± 0.01 |
| MED | ||||||||
| WT | 0.38 ± 0.05 | 5.20 ± 0.49 | 6.12 ± 0.06 | 0.13 ± 0.00 | 0.19 ± 0.00 | 2.14 ± 0.02 | 3.44 ± 0.04 | 0.80 ± 0.03 |
| DSUL1 | 0.33 ± 0.04 | 4.69 ± 0.99 | 6.14 ± 0.03 | 0.12 ± 0.01 | 0.17 ± 0.01 | 2.07 ± 0.02 | 3.51 ± 0.05 | 0.81 ± 0.01 |
| DSUL2 | 0.23 ± 0.06 | 3.86 ± 1.27 | 6.10 ± 0.05 | 0.12 ± 0.00 | 0.18 ± 0.00 | 2.09 ± 0.00 | 3.35 ± 0.03 | 0.81 ± 0.02 |
| DSUL1 /DSUL2 | 0.16 ± 0.01 | 2.65 ± 0.14 | 5.70 ± 0.02 | 0.12 ± 0.01 | 0.18 ± 0.02 | 1.99 ± 0.05 | 3.29 ± 0.04 | 0.72 ± 0.01 |
| LOW | ||||||||
| WT | 0.11 ± 0.03 | 3.45 ± 0.21 | 6.20 ± 0.10 | 0.37 ± 0.00 | 0.13 ± 0.01 | 2.54 ± 0.05 | 3.25 ± 0.07 | 0.93 ± 0.02 |
| DSUL1 | 0.10 ± 0.05 | 3.70 ± 0.49 | 6.25 ± 0.10 | 0.35 ± 0.01 | 0.11 ± 0.01 | 2.50 ± 0.03 | 3.26 ± 0.10 | 0.89 ± 0.03 |
| DSUL2 | 0.00 ± 0.04 | 2.69 ± 0.61 | 6.07 ± 0.06 | 0.37 ± 0.00 | 0.11 ± 0.00 | 2.49 ± 0.06 | 3.14 ± 0.10 | 0.88 ± 0.00 |
| DSUL1/DSUL2 | 0.19 ± 0.01 | 5.05 ± 0.19 | 6.00 ± 0.08 | 0.42 ± 0.01 | 0.13 ± 0.01 | 2.54 ± 0.01 | 3.12 ± 0.03 | 0.90 ± 0.02 |
| Strain | Alcohols | Carboxylic Acids | Esters | Terpenes |
|---|---|---|---|---|
| (mg/L) | (mg/L) | (mg/L) | (mg/L) | |
| STR | ||||
| WT | 154.49 ± 9.66 | 14.60 ± 2.15 | 9.13 ± 0.76 | 143.04 ± 12.81 |
| DSUL1 /DSUL2 | 145.75 ± 4.27 | 16.80 ± 0.16 | 9.84 ± 0.27 | 149.98 ± 15.39 |
| HIGH | ||||
| WT | 176.80 ± 9.19 | 13.74 ± 0.96 | 6.84 ± 0.46 | 85.63 ± 11.40 |
| DSUL1 /DSUL2 | 143.90 ± 4.45 | 16.48 ± 1.42 | 7.41 ± 0.28 | 104.73 ± 8.94 |
| MED | ||||
| WT | 167.70 ± 3.63 | 32.35 ± 1.30 | 8.84 ± 0.24 | 139.75 ± 7.37 |
| DSUL1 /DSUL2 | 174.17 ± 3.18 | 22.94 ± 1.57 | 8.32 ± 0.23 | 142.84 ± 5.17 |
| LOW | ||||
| WT | 174.88 ± 11.18 | 14.00 ± 0.76 | 12.15 ± 1.10 | 134.88 ± 5.39 |
| DSUL1 /DSUL2 | 135.51 ± 6.38 | 15.67 ± 1.00 | 10.53 ± 0.76 | 131.79 ± 6.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Granuzzo, S.; Righetto, F.; Peggion, C.; Bosaro, M.; Frizzarin, M.; Antoniali, P.; Sartori, G.; Lopreiato, R. Sulphate Uptake Plays a Major Role in the Production of Sulphur Dioxide by Yeast Cells during Oenological Fermentations. Fermentation 2023, 9, 280. https://doi.org/10.3390/fermentation9030280
Granuzzo S, Righetto F, Peggion C, Bosaro M, Frizzarin M, Antoniali P, Sartori G, Lopreiato R. Sulphate Uptake Plays a Major Role in the Production of Sulphur Dioxide by Yeast Cells during Oenological Fermentations. Fermentation. 2023; 9(3):280. https://doi.org/10.3390/fermentation9030280
Chicago/Turabian StyleGranuzzo, Sara, Francesca Righetto, Caterina Peggion, Matteo Bosaro, Martina Frizzarin, Paolo Antoniali, Geppo Sartori, and Raffaele Lopreiato. 2023. "Sulphate Uptake Plays a Major Role in the Production of Sulphur Dioxide by Yeast Cells during Oenological Fermentations" Fermentation 9, no. 3: 280. https://doi.org/10.3390/fermentation9030280
APA StyleGranuzzo, S., Righetto, F., Peggion, C., Bosaro, M., Frizzarin, M., Antoniali, P., Sartori, G., & Lopreiato, R. (2023). Sulphate Uptake Plays a Major Role in the Production of Sulphur Dioxide by Yeast Cells during Oenological Fermentations. Fermentation, 9(3), 280. https://doi.org/10.3390/fermentation9030280