

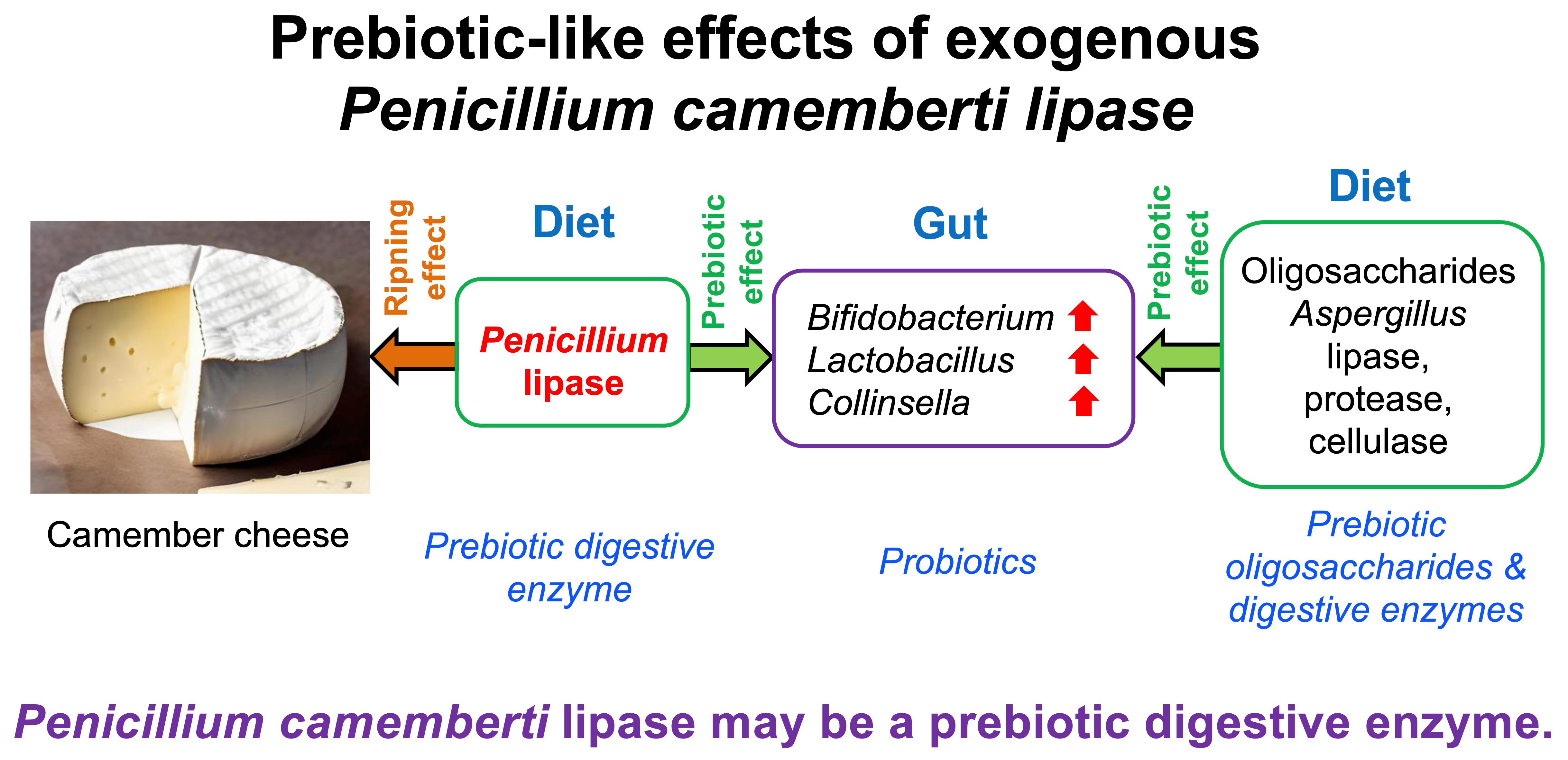
Exogenous Penicillium camemberti Lipase Preparation Exerts Prebiotic-like Effects by Increasing Cecal Bifidobacterium and Lactobacillus Abundance in Rats
 ,
,  , , ,
, , ,
Abstract

{kind=link}
{kind=link}
{kind=link}
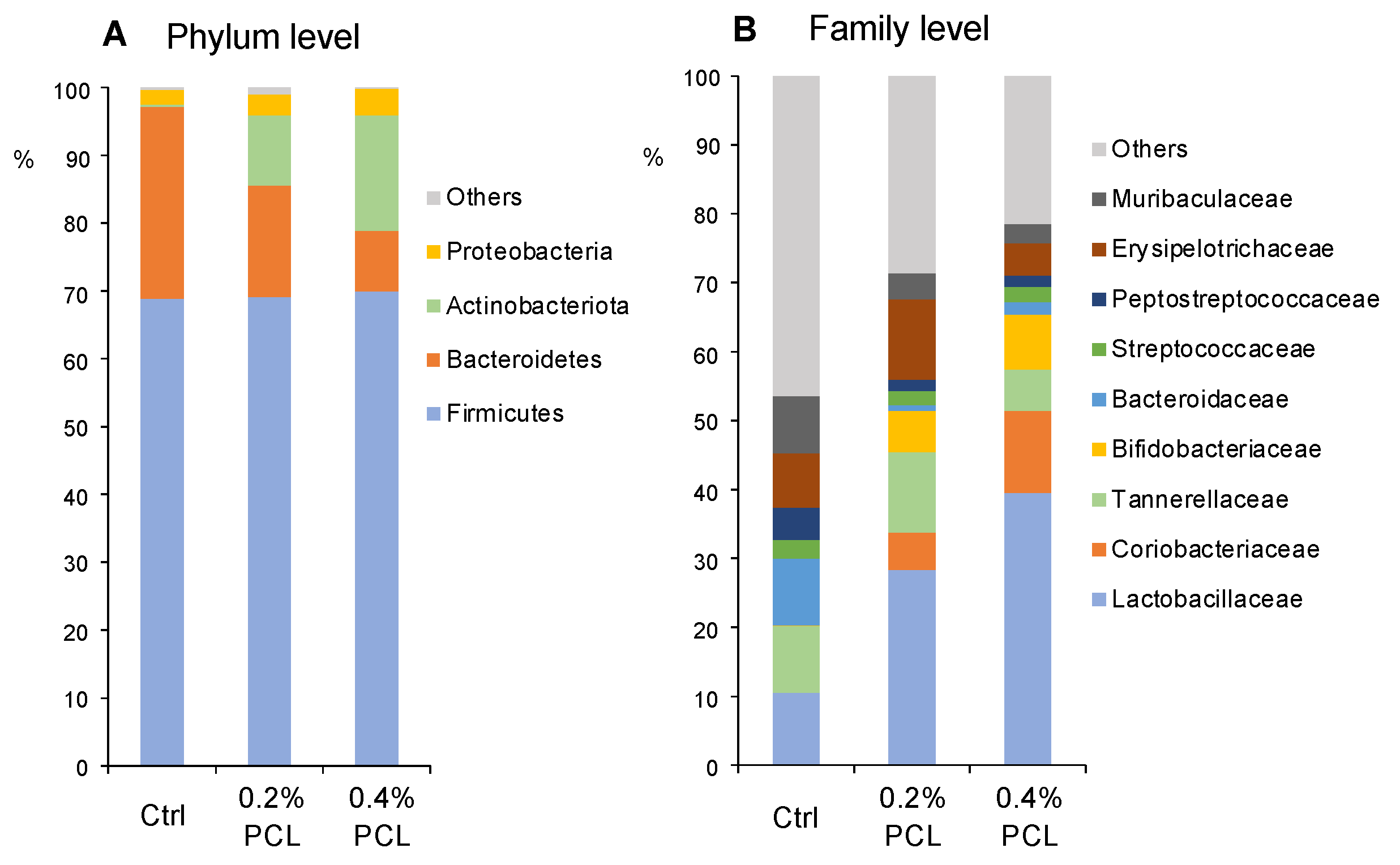{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Sample Collection
2.3. Bacterial DNA Extraction and 16S rRNA Gene-Based Microbiome Analysis
2.4. Bioinformatics Analysis
2.5. Analysis of Cecal Organic Acids
2.6. Data Analysis
3. Results
3.1. Food Intake, Body Weight, and Cecal Content Weight
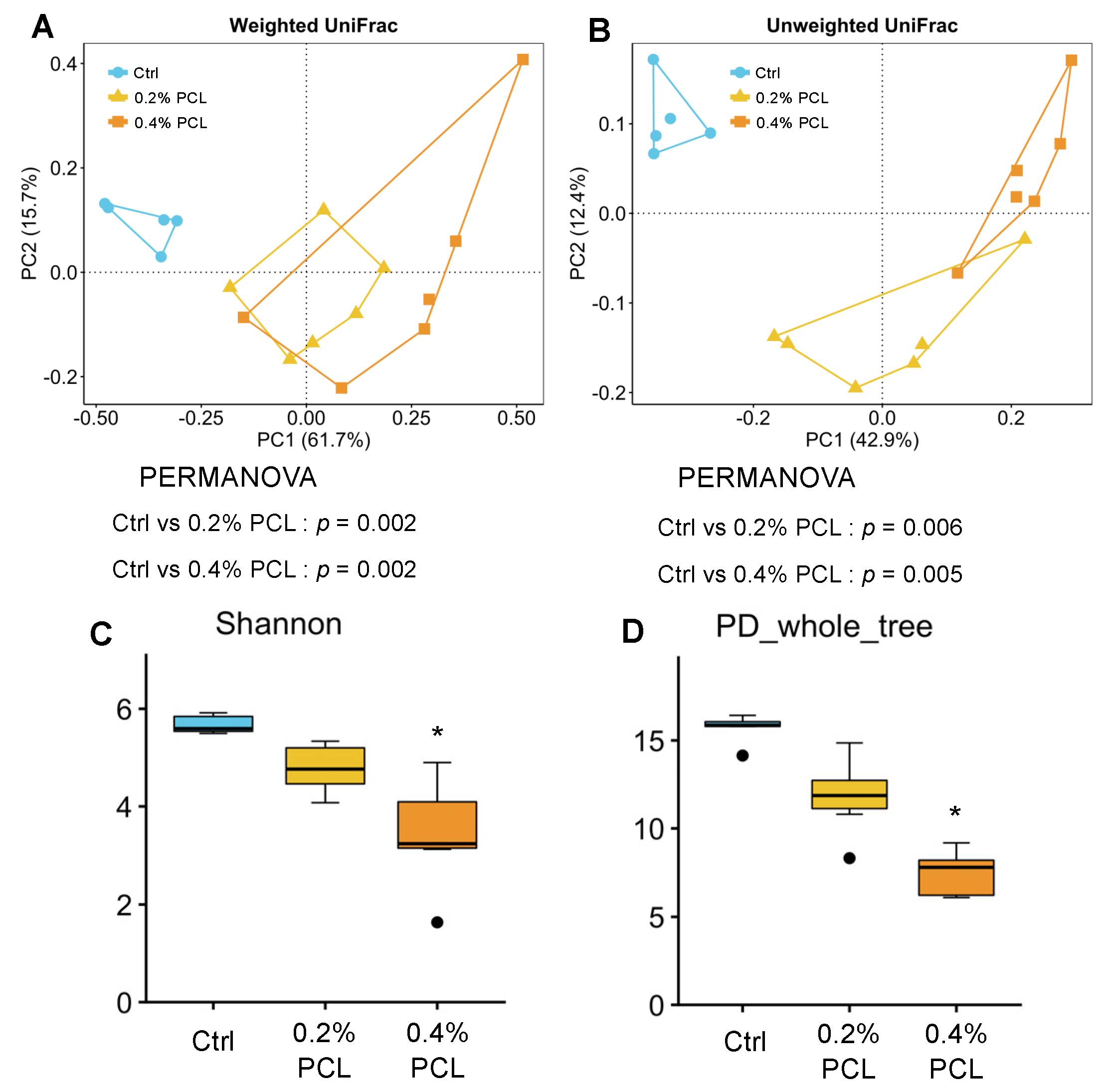
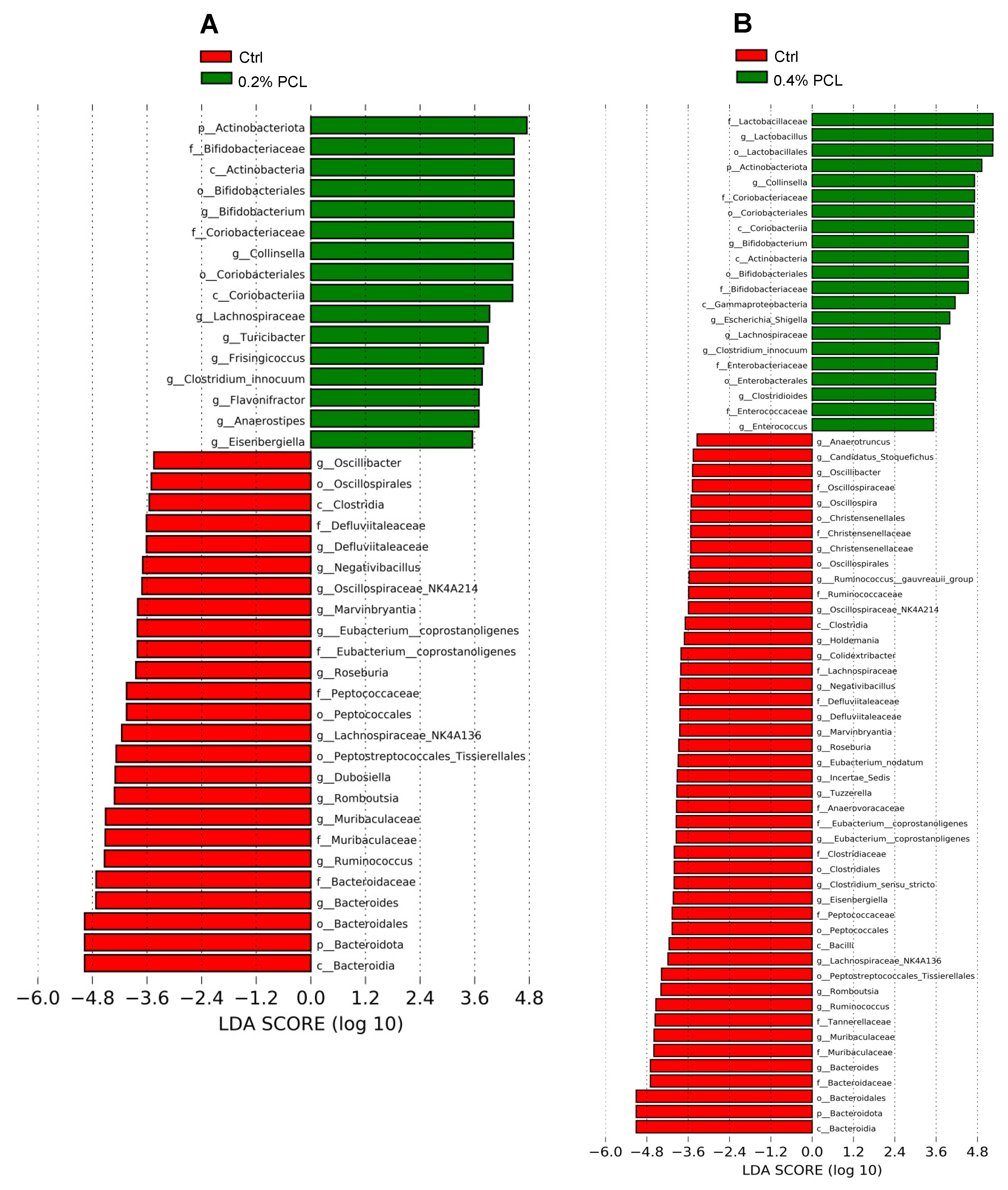
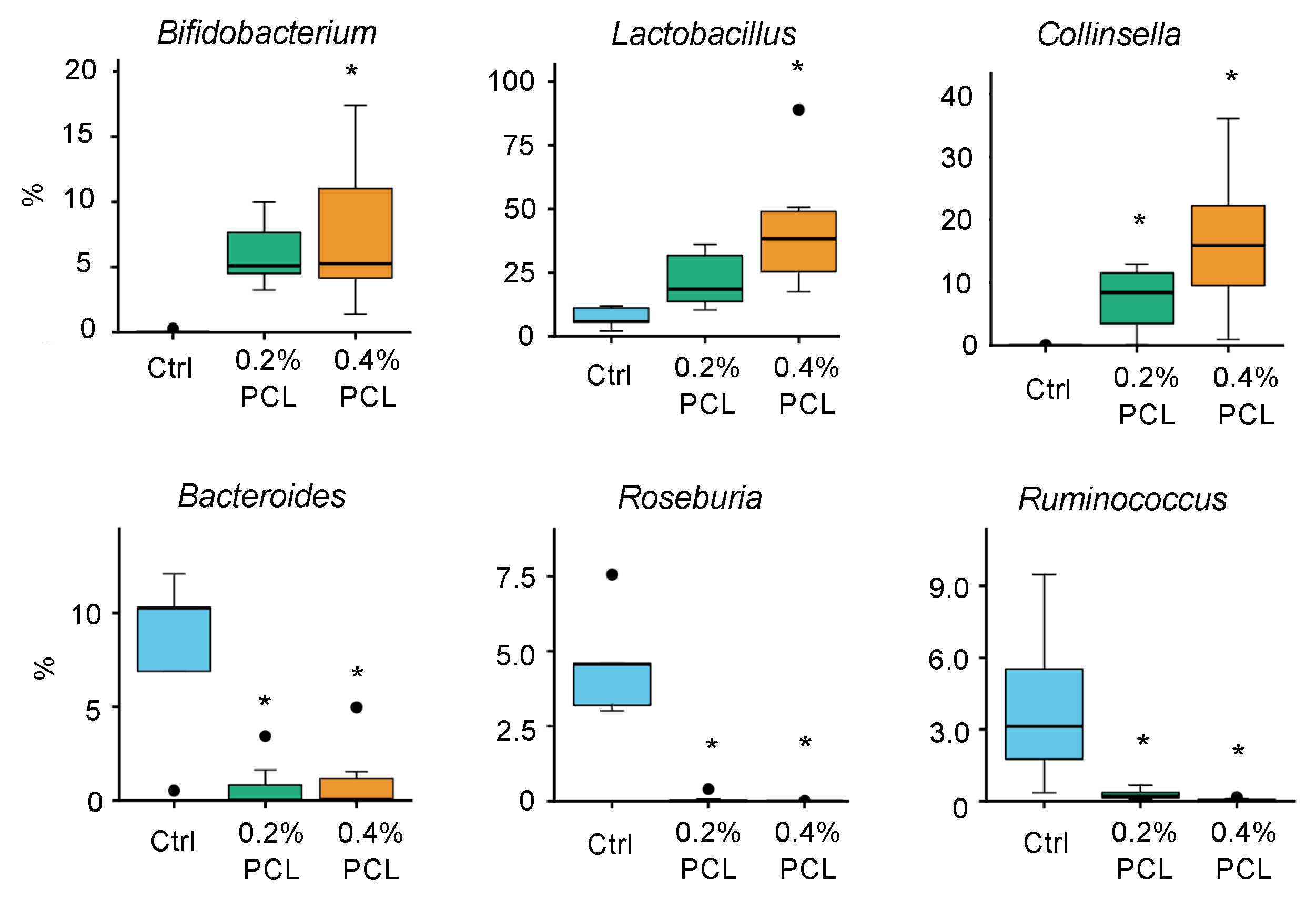
3.2. Cecal Microbiota
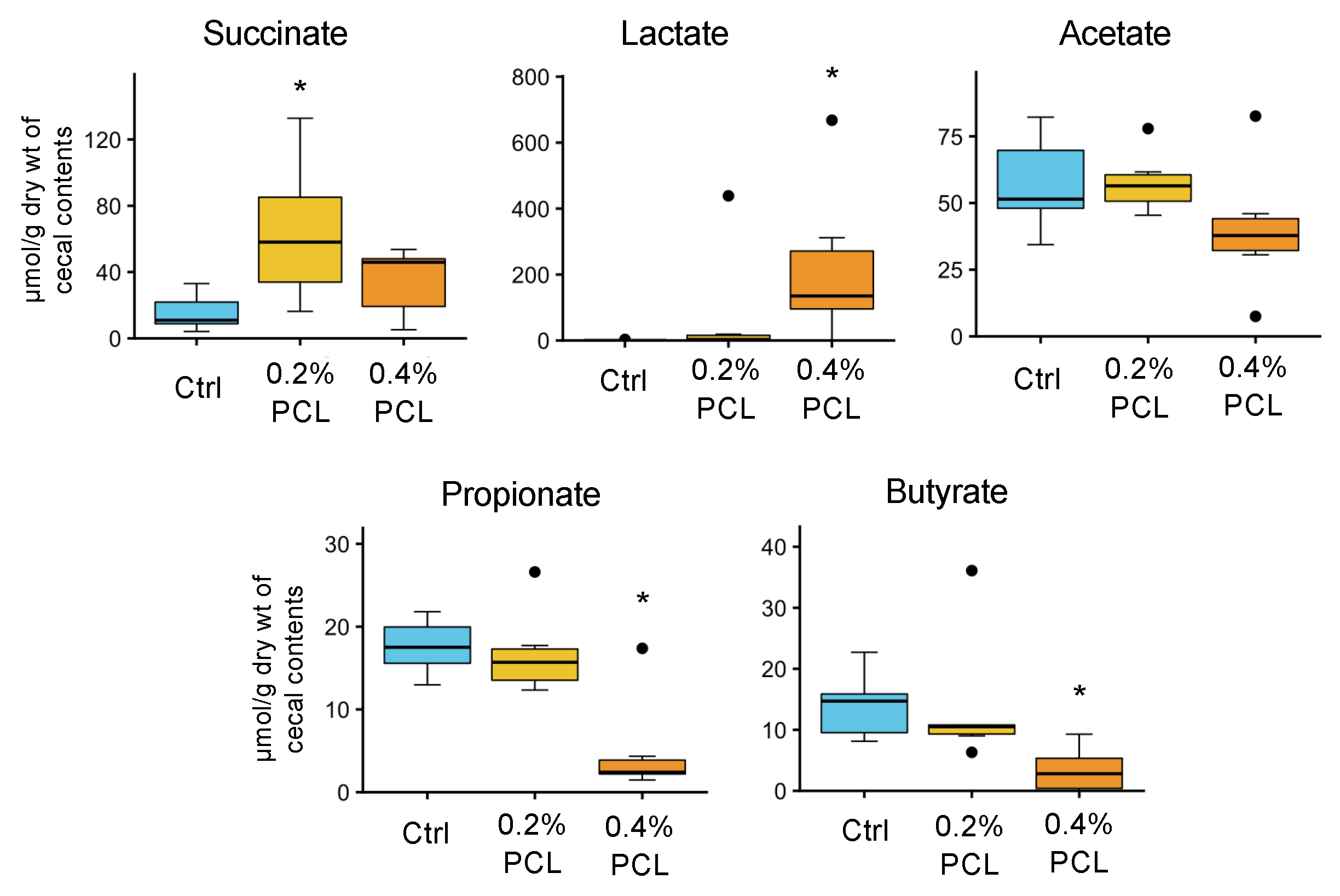
3.3. Cecal Organic Acids
3.4. PICRUSt Analysis
4. Discussion
4.1. Bifidobacterium, Lactobacillus, and Collinsella
4.2. Other Affected Genera
4.3. Bacterial Diversity
4.4. Organic Acids
4.5. PICRUSt Analysis
4.6. Limitations of this Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, M.-J.; Tan, J.-S.; Gao, X.-J.; Yang, J.-G.; Yang, Y.-J. Effect of Cheese Intake on Cardiovascular Diseases and Cardiovascular Biomarkers. Nutrients 2022, 14, 2936. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Bueno, C.; Cavero-Redondo, I.; Martinez-Vizcaino, V.; Sotos-Prieto, M.; Ruiz, J.R.; Gil, A. Effects of Milk and Dairy Product Consumption on Type 2 Diabetes: Overview of Systematic Reviews and Meta-Analyses. Adv. Nutr. 2019, 10 (Suppl. 2), S154–S163. [Google Scholar] [CrossRef] [PubMed]
- Papadimitriou, N.; Bouras, E.; Brandt, P.A.V.D.; Muller, D.C.; Papadopoulou, A.; Heath, A.K.; Critselis, E.; Gunter, M.J.; Vineis, P.; Ferrari, P.; et al. A Prospective Diet-Wide Association Study for Risk of Colorectal Cancer in EPIC. Clin. Gastroenterol. Hepatol. 2022, 20, 864–873.e13. [Google Scholar] [CrossRef] [PubMed]
- Gil, Á.; Ortega, R.M. Introduction and Executive Summary of the Supplement, Role of Milk and Dairy Products in Health and Prevention of Noncommunicable Chronic Diseases: A Series of Systematic Reviews. Adv. Nutr. 2019, 10, S67–S73. [Google Scholar] [CrossRef] [PubMed]
- Park, K.M.; Fulgoni, V.L. The association between dairy product consumption and cognitive function in the National Health and Nutrition Examination Survey. Br. J. Nutr. 2013, 109, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, Z.; Gu, L.; Zhang, K. Fermented dairy foods consumption and depressive symptoms: A meta-analysis of cohort studies. PLoS ONE 2023, 18, e0281346. [Google Scholar] [CrossRef]
- Bruna, J.M.; Hierro, E.M.; de la Hoz, L.; Mottram, D.S.; Fernández, M.; Ordóñez, J.A. Changes in selected biochemical and sensory parameters as affected by the superficial inoculation of Penicillium camemberti on dry fermented sausages. Int. J. Food Microbiol. 2003, 85, 111–125. [Google Scholar] [CrossRef]
- Ano, Y.; Ozawa, M.; Kutsukake, T.; Sugiyama, S.; Uchida, K.; Yoshida, A.; Nakayama, H. Preventive Effects of a Fermented Dairy Product against Alzheimer’s Disease and Identification of a Novel Oleamide with Enhanced Microglial Phagocytosis and Anti-Inflammatory Activity. PLoS ONE 2015, 10, e0118512. [Google Scholar] [CrossRef]
- Ano, Y.; Nakayama, H. Preventive Effects of Dairy Products on Dementia and the Underlying Mechanisms. Int. J. Mol. Sci. 2018, 19, 1927. [Google Scholar] [CrossRef]
- Yang, Y.; Kumrungsee, T.; Kato, N.; Fukuda, S.; Kuroda, M.; Yamaguchi, S. Supplemental Aspergillus Lipase and Protease Preparations Display Powerful Bifidogenic Effects and Modulate the Gut Microbiota Community of Rats. Fermentation 2021, 7, 294. [Google Scholar] [CrossRef]
- Yang, Y.; Sitanggang, N.V.; Kato, N.; Inoue, J.; Murakami, T.; Watanabe, T.; Iguchi, T.; Okazaki, Y. Beneficial effects of protease preparations derived from Aspergillus on the colonic luminal environment in rats consuming a high-fat diet. Biomed. Rep. 2015, 3, 715–720. [Google Scholar] [CrossRef]
- Yang, Y.; Kumrungsee, T.; Kuroda, M.; Yamaguchi, S.; Kato, N. Feeding Aspergillus protease preparation combined with adequate protein diet to rats increases levels of cecum gut-protective amino acids, partially linked to Bifidobacterium and Lactobacillus. Biosci. Biotech. Biochem. 2019, 83, 1901–1911. [Google Scholar] [CrossRef]
- Yang, Y.; Kumrungsee, T.; Kato, N.; Fukuda, S.; Kuroda, M.; Yamaguchi, S. Aspergillus-Derived Cellulase Preparation Exhibits Prebiotic-like Effects on Gut Microbiota in Rats. Fermentation 2022, 8, 71. [Google Scholar] [CrossRef]
- Yang, Y.; Iwamoto, A.; Kumrungsee, T.; Okazaki, Y.; Kuroda, M.; Yamaguchi, S.; Kato, N. Consumption of an acid protease derived from Aspergillus oryzae causes bifidogenic effect in rats. Nutr. Res. 2017, 44, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable and Extensible Microbiome Data Science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PIC-RUSt2: An improved and customizable approach for metagenome inference. BioRxiv 2020, 672295. [Google Scholar] [CrossRef]
- Ramli, N.S.; Jia, H.; Sekine, A.; Lyu, W.; Furukawa, K.; Saito, K.; Hasebe, Y.; Kato, H. Eggshell membrane powder lowers plasma triglyceride and liver total cholesterol by modulating gut microbiota and accelerating lipid metabolism in high-fat diet-fed mice. Food Sci. Nutr. 2020, 8, 2512–2523. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Cani, P.; Claus, S.; Fuentes, S.; Puylaert, P.G.B.; Neyrinck, A.; Bindels, L.B.; De Vos, W.M.; Gibson, G.R.; Thissen, J.-P.; et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2013, 62, 1112. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial lipases and their industrial applications: A comprehensive review. Microb. Cell Fact. 2020, 19, 169. [Google Scholar] [CrossRef]
- Torossian, K.; Bell, A.W. Purification and characterization of an acid-resistant triacylglycerol lipase from Aspergillus niger. Biotechnol. Appl. Biochem. 1991, 13, 205–211. [Google Scholar]
- Tan, H.; Zhai, Q.; Chen, W. Investigations of Bacteroides spp. towards next-generation probiotics. Food Res. Int. 2019, 116, 637–644. [Google Scholar] [CrossRef] [PubMed]
- Burokas, A.; Arboleya, S.; Moloney, R.D.; Peterson, V.L.; Murphy, K.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Targeting the Microbiota-Gut-Brain Axis: Prebiotics Have Anxiolytic and Antidepressant-like Effects and Reverse the Impact of Chronic Stress in Mice. Biol. Psychiatry 2017, 82, 472–487. [Google Scholar] [CrossRef]
- Yang, X.-D.; Wang, L.-K.; Wu, H.-Y.; Jiao, L. Effects of prebiotic galacto-oligosaccharide on postoperative cognitive dysfunction and neuroinflammation through targeting of the gut-brain axis. BMC Anesthesiol. 2018, 18, 177. [Google Scholar] [CrossRef]
- Liu, F.; Li, P.; Chen, M.; Luo, Y.; Prabhakar, M.; Zheng, H.; He, Y.; Qi, Q.; Long, H.; Zhang, Y.; et al. Fructooligosaccharide (FOS) and Galactooligosaccharide (GOS) Increase Bifidobacterium but Reduce Butyrate Producing Bacteria with Adverse Glycemic Metabolism in healthy young population. Sci. Rep. 2017, 7, 11789. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; Jonkers, D.M.; Salonen, A.; Hanevik, K.; Raes, J.; Jalanka, J.; de Vos, W.M.; Manichanh, C.; Golic, N.; Enck, P.; et al. Intestinal Microbiota and Diet in IBS: Causes, Consequences, or Epiphenomena? Am. J. Gastroenterol. 2015, 110, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Henke, M.T.; Kenny, D.J.; Cassilly, C.D.; Vlamakis, H.; Xavier, R.J.; Clardy, J. Ruminococcus gnavus, a member of the human gut microbiome associated with Crohn’s disease, produces an inflammatory polysaccharide. Proc. Natl. Acad. Sci. USA 2019, 116, 12672–12677. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Brunt, J.; Crost, E.; Vaux, L.; Nepravishta, R.; Owen, C.D.; Latousakis, D.; Xiao, A.; Li, W.; Chen, X.; et al. Elucidation of a sialic acid metabolism pathway in mucus-foraging Ruminococcus gnavus unravels mechanisms of bacterial adaptation to the gut. Nat. Microbiol. 2019, 4, 2393–2404. [Google Scholar] [CrossRef]
- Kageyama, Y.; Akiyama, T.; Nakamura, T. Intestinal Dysbiosis and Probiotics in COVID-19. J. Clin. Trials 2020, 10, 421. [Google Scholar]
- Nie, K.; Ma, K.; Luo, W.; Shen, Z.; Yang, Z.; Xiao, M.; Tong, T.; Yang, Y.; Wang, X. Roseburia intestinalis: A Beneficial Gut Organism from the Discoveries in Genus and Species. Front. Cell. Infect. Microbiol. 2021, 11, 757718. [Google Scholar] [CrossRef]
- Song, L.; Sun, Q.; Zheng, H.; Zhang, Y.; Wang, Y.; Liu, S.; Duan, L. Roseburia hominis Alleviates Neuroinflammation via Short-Chain Fatty Acids through Histone Deacetylase Inhibition. Mol. Nutr. Food Res. 2022, 66, 2200164. [Google Scholar] [CrossRef]
- Cai, H.; Wen, Z.; Li, X.; Meng, K.; Yang, P. Lactobacillus plantarum FRT10 alleviated high-fat diet–induced obesity in mice through regulating the PPARα signal pathway and gut microbiota. Appl. Microbiol. Biotechnol. 2020, 104, 5959–5972. [Google Scholar] [CrossRef] [PubMed]
- Sjöberg, F.; Barkman, C.; Nookaew, I.; Östman, S.; Adlerberth, I.; Saalman, R.; Wold, A.E. Low-complexity microbiota in the duodenum of children with newly diagnosed ulcerative colitis. PLoS ONE 2017, 12, e0186178. [Google Scholar] [CrossRef] [PubMed]
- Shade, A. Diversity is the question, not the answer. ISME J. 2017, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Duncan, S.H.; Scott, K.P.; Louis, P. Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 2015, 74, 13–22. [Google Scholar] [CrossRef]
- Manosalva, C.; Quiroga, J.; Hidalgo, A.I.; Alarcón, P.; Anseoleaga, N.; Hidalgo, M.A.; Burgos, R.A. Role of Lactate in Inflammatory Processes: Friend or Foe. Front. Immunol. 2022, 12, 808799. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Zhang, B.; Lin, X.; Fu, X.; An, Y.; Zou, Y.; Wang, J.X.; Wang, Z.; Yu, T. Lactate metabolism in human health and disease. Signal Transduct. Target. Ther. 2022, 7, 305. [Google Scholar] [CrossRef]
- Park, B.O.; Kang, J.S.; Paudel, S.; Park, S.G.; Park, B.C.; Han, S.B.; Kwak, Y.S.; Kim, J.H.; Kim, S. Novel GPR43 Agonists Exert an Anti-Inflammatory Effect in a Colitis Model. Biomol. Ther. 2022, 30, 48–54. [Google Scholar] [CrossRef]
- Wei, X.; Yu, L.; Zhang, C.; Ni, Y.; Zhao, J.; Zhang, H.; Chen, W.; Zhai, Q.; Tian, F. Prebiotic activity of chitooligosaccharides and their ability to alleviate necrotizing enterocolitis in newborn rats. Carbohydr. Polym. 2023, 299, 120156. [Google Scholar] [CrossRef]
- Matsumoto, M.; Kunisawa, A.; Hattori, T.; Kawana, S.; Kitada, Y.; Tamada, H.; Kawano, S.; Hayakawa, Y.; Iida, J.; Fukusaki, E. Free D-amino acids produced by commensal bacteria in the colonic lumen. Sci. Rep. 2018, 8, 17915. [Google Scholar] [CrossRef]
- Sasabe, J.; Suzuki, M. Emerging Role of D-Amino Acid Metabolism in the Innate Defense. Front. Microbiol. 2018, 9, 933. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Jia, H.; Ando, C.; Kato, H.; Kumrungsee, T.; Kato, N.; Kimoto, A.; Fukuda, S.; Kuroda, M.; Nishio, K.; et al. Exogenous Penicillium camemberti Lipase Preparation Exerts Prebiotic-like Effects by Increasing Cecal Bifidobacterium and Lactobacillus Abundance in Rats. Fermentation 2023, 9, 227. https://doi.org/10.3390/fermentation9030227
Yang Y, Jia H, Ando C, Kato H, Kumrungsee T, Kato N, Kimoto A, Fukuda S, Kuroda M, Nishio K, et al. Exogenous Penicillium camemberti Lipase Preparation Exerts Prebiotic-like Effects by Increasing Cecal Bifidobacterium and Lactobacillus Abundance in Rats. Fermentation. 2023; 9(3):227. https://doi.org/10.3390/fermentation9030227
Chicago/Turabian StyleYang, Yongshou, Huijuan Jia, Chika Ando, Hisanori Kato, Thanutchaporn Kumrungsee, Norihisa Kato, Akiko Kimoto, Shinji Fukuda, Manabu Kuroda, Kyoichi Nishio, and et al. 2023. "Exogenous Penicillium camemberti Lipase Preparation Exerts Prebiotic-like Effects by Increasing Cecal Bifidobacterium and Lactobacillus Abundance in Rats" Fermentation 9, no. 3: 227. https://doi.org/10.3390/fermentation9030227
APA StyleYang, Y., Jia, H., Ando, C., Kato, H., Kumrungsee, T., Kato, N., Kimoto, A., Fukuda, S., Kuroda, M., Nishio, K., & Yamaguchi, S. (2023). Exogenous Penicillium camemberti Lipase Preparation Exerts Prebiotic-like Effects by Increasing Cecal Bifidobacterium and Lactobacillus Abundance in Rats. Fermentation, 9(3), 227. https://doi.org/10.3390/fermentation9030227