
Modulation of the Aromatic Profile of Verdejo Wine through Sequential Inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Culture Conditions
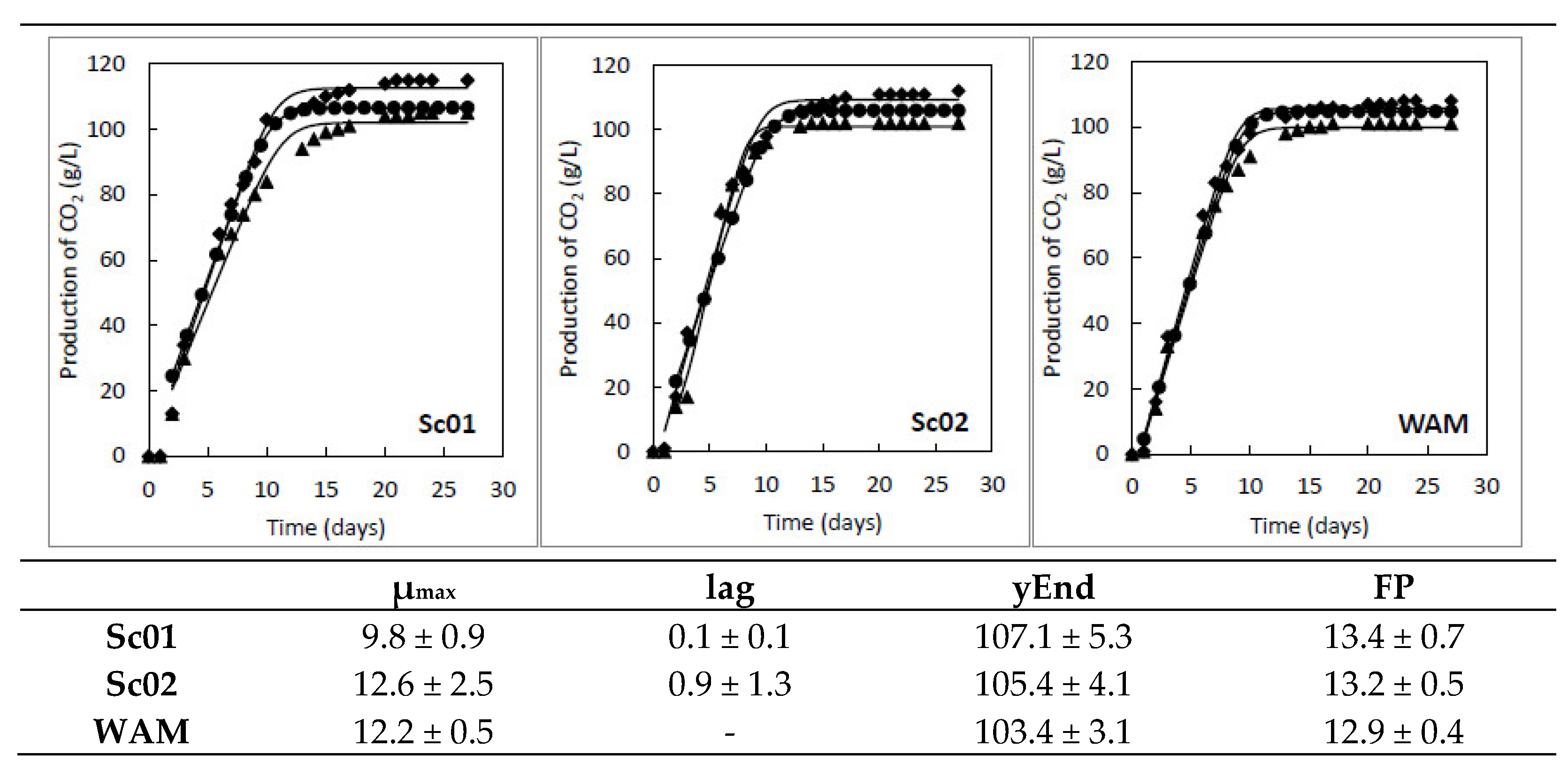
2.2. Kinetic Parameters of S. cerevisiae Strains
2.3. Trial Fermentations in the Winery
2.3.1. Population Dynamic
2.3.2. Implantation Studies
2.3.3. Wine Chemical Composition
2.3.4. Analysis of Volatile Organic Compounds (VOCs)
2.4. Statistical Analysis
3. Results and Discussion
3.1. Kinetic Aptitude of Native S. cerevisiae Strains
3.2. Trial Fermentations in the Winery
3.3. VOCs Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fleet, G.H. Yeast Interactions and Wine Flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Comitini, F.; Capece, A.; Ciani, M.; Romano, P. New Insights on the Use of Wine Yeasts. Curr. Opin. Food Sci. 2017, 13, 44–49. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 2016, 7, 411. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Challenges of the Non-Conventional Yeast Wickerhamomyces anomalus in Winemaking. Fermentation 2018, 4, 68. [Google Scholar] [CrossRef]
- Bagheri, B.; Zambelli, P.; Vigentini, I.; Bauer, F.F.; Setati, M.E. Investigating the Effect of Selected Non-Saccharomyces Species on Wine Ecosystem Function and Major Volatiles. Front. Bioeng. Biotechnol. 2018, 6, 169. [Google Scholar] [CrossRef]
- Lee, S.-B.; Park, H.-D. Isolation and Investigation of Potential Non-Saccharomyces Yeasts to Improve the Volatile Terpene Compounds in Korean Muscat Bailey A Wine. Microorganisms 2020, 8, 1552. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Phylogeny of the Ascomycetous Yeasts and the Renaming of Pichia anomala to Wickerhamomyces anomalus. Antonie Van Leeuwenhoek 2011, 99, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Robnett, C.J.; Basehoar-Powers, E. Phylogenetic Relationships among Species of Pichia, Issatchenkia and Williopsis Determined from Multigene Sequence Analysis, and the Proposal of Barnettozyma Gen. Nov., Lindnera Gen. Nov. and Wickerhamomyces Gen. FEMS Yeast Res. 2008, 8, 939–954. [Google Scholar] [CrossRef] [PubMed]
- Passoth, V.; Fredlund, E.; Druvefors, U.A.; Schnürer, J. Biotechnology, Physiology and Genetics of the Yeast Pichia anomala. FEMS Yeast Res. 2006, 6, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Kurita, O. Increase of Acetate Ester-Hydrolysing Esterase Activity in Mixed Cultures of Saccharomyces cerevisiae and Pichia anomala. J. Appl. Microbiol. 2008, 104, 1051–1058. [Google Scholar] [CrossRef]
- Loureiro, V.; Malfeito-Ferreira, M. Spoilage Yeasts in the Wine Industry. Int. J. Food Microbiol. 2003, 86, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Sabel, A.; Martens, S.; Petri, A.; Konig, H.; Claus, H. Wickerhamomyces anomalus AS1: A New Strain with Potential to Improve Wine Aroma. Ann. Microbiol. 2014, 64, 483–491. [Google Scholar] [CrossRef]
- Schwentke, J.; Sabel, A.; Petri, A.; König, H.; Claus, H. The Yeast Wickerhamomyces anomalus AS1 Secretes a Multifunctional Exo-β-1, 3-Glucanase with Implications for Winemaking. Yeast 2014, 31, 349–359. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yan, J.J.; Zhang, W.J.; Zhang, Y.X.; Dong, Z.G.; Luo, H.; Liu, M.; Su, J. Comparison of Potential Wickerhamomyces anomalus to Improve the Quality of Cabernet Sauvignon Wines by Mixed Fermentation with Saccharomyces cerevisiae. LWT Food Sci. Technol. 2023, 173, 114285. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genovés, S.; Vallés, S.; Manzanares, P. Rational Selection of Non-Saccharomyces Wine Yeasts for Mixed Starters Based on Ester Formation and Enological Traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef]
- Schlander, M.; Distler, U.; Tenzer, S.; Thines, E.; Claus, H. Purification and Properties of Yeast Proteases Secreted by Wickerhamomyces anomalus 227 and Metschnikovia pulcherrima 446 during Growth in a White Grape Juice. Fermentation 2017, 3, 2. [Google Scholar] [CrossRef]
- Lin, M.; Boss, P.; Walker, M.; Sumby, K.; Grbin, P.; Jiranek, V. Evaluation of Indigenous Non-Saccharomyces Yeasts Isolated from a South Australian Vineyard for Their Potential as Wine Starter Cultures. Int. J. Food Microbiol. 2020, 312, 108373. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial Resources and Enological Significance: Opportunities and Benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a Future for Non-Saccharomyces Yeasts: Selection of Putative Spoilage Wine Strains to Be Used in Association with Saccharomyces cerevisiae for Grape Juice Fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate Ester Formation in Wine by Mixed Cultures in Laboratory Fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Izquierdo-Cañas, P.M.; García-Romero, E.; Heras Manso, J.M.; Fernández-González, M. Influence of Sequential Inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the Quality of Red Wines. Eur. Food Res. Technol. 2014, 239, 279–286. [Google Scholar] [CrossRef]
- Izquierdo-Cañas, P.M.; Palacios García, A.T.; García Romero, E.G. Enhancement of Flavour Properties in Wines Using Sequential Inoculations of Non-Saccharomyces (Hansenula and Torulaspora) and Saccharomyces Yeast Starter. VITIS J. Grapevine Res. 2011, 50, 177–182. [Google Scholar]
- López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Screening and Enzymatic Evaluation of Saccharomyces cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines. Foods 2022, 11, 3448. [Google Scholar] [CrossRef] [PubMed]
- López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Non-Saccharomyces Yeasts from Organic Vineyards as Spontaneous Fermentation Agents. Foods 2023, 12, 3644. [Google Scholar] [CrossRef] [PubMed]
- Baranyi, J.; Roberts, T.A. A Dynamic Approach to Predicting Bacterial Growth in Food. Int. J. Food Microbiol. 1994, 23, 277–294. [Google Scholar] [CrossRef] [PubMed]
- OIV. Compendium of International Methods of Wine and Must Analysis; International Organisation of Vine and Wine: Paris, France, 2020; ISBN 978-2-85038-016-7. [Google Scholar]
- Legras, J.-L.; Karst, F. Optimisation of Interdelta Analysis for Saccharomyces cerevisiae Strain Characterisation. FEMS Microbiol. Lett. 2003, 221, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Huerta, C.; Fernández-Fernández, E.; Vila-Crespo, J.; Ruipérez, V.; Moyano, R.; Rodríguez-Nogales, J.M. Impact of Ageing on Ultrasound-Treated Lees on Volatile Composition and Sensory Properties of Red Sparkling Base Wine. Beverages 2023, 9, 23. [Google Scholar] [CrossRef]
- Ayestaran, B.; Martinez-Lapuente, L.; Guadalupe, Z.; Canals, C.; Adell, E.; Vilanova, M. Effect of the Winemaking Process on the Volatile Composition and Aromatic Profile of Tempranillo Blanco Wines. Food Chem. 2019, 276, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, D.; Neira, N.; Blanco, P. Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation 2020, 6, 89. [Google Scholar] [CrossRef]
- Gonzalez, R.; Quirós, M.; Morales, P. Yeast Respiration of Sugars by Non-Saccharomyces Yeast Species: A Promising and Barely Explored Approach to Lowering Alcohol Content of Wines. Trends Food Sci. Technol. 2013, 29, 55–61. [Google Scholar] [CrossRef]
- Sumby, K.M.; Grbin, P.R.; Jiranek, V. Microbial Modulation of Aromatic Esters in Wine: Current Knowledge and Future Prospects. Food Chem. 2010, 121, 1–16. [Google Scholar] [CrossRef]
- Dzialo, M.C.; Park, R.; Steensels, J.; Lievens, B.; Verstrepen, K.J. Physiology, Ecology and Industrial Applications of Aroma Formation in Yeast. FEMS Microbiol. Rev. 2017, 41, S95–S128. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Yeast Modulation of Wine Flavor. In Advances in Applied Microbiology; Laskin, A.I., Bennett, J.W., Gadd, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 57, pp. 131–175. ISBN 0065-2164. [Google Scholar]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Pollon, M.; Fracassetti, D.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Volatile Profile of White Wines Fermented with Sequential Inoculation of Starmerella bacillaris and Saccharomyces cerevisiae. Food Chem. 2018, 257, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Benito, A.; Calderón, F.; Benito, S. The Influence of Non-Saccharomyces Species on Wine Fermentation Quality Parameters. Fermentation 2019, 5, 54. [Google Scholar] [CrossRef]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of Non-Saccharomyces Yeasts to Wine Volatile and Sensory Diversity: A Study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris Strains Isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef]
- Borren, E.; Tian, B. The Important Contribution of Non-Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2021, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine Aromatic Compound Production and Fermentative Behaviour within Different Non-Saccharomyces Species and Clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled Mixed Culture Fermentation: A Newperspective on the Use of Non-Saccharomyces Yeasts in Winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast Interactions in Inoculated Wine Fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef]
- Arcari, S.G.; Caliari, V.; Sganzerla, M.; Godoy, H.T. Volatile Composition of Merlot Red Wine and Its Contribution to the Aroma: Optimization and Validation of Analytical Method. Talanta 2017, 174, 752–766. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Sc01 | Wa/Sc01 | Sc02 | Wa/Sc02 | |
|---|---|---|---|---|
| Reducing sugars (g/L) | 2.1 ± 0.8 | 2.9 ± 0.1 | 1.8 ± 0.2 | 1.9 ± 0.1 |
| pH | 3.29 ± 0.01 | 3.26 ± 0.01 | 3.28 ± 0.01 | 3.26 ± 0.01 |
| Total acidity (g/L) | 6.3 ± 0.0 | 6.7 ± 0.6 | 6.1 ± 0.1 | 6.7 ± 0.0 |
| Volatile acidity (g/L) | 0.66 ± 0.01 | 1.03 ± 0.05 | 0.51 ± 0.05 | 0.91 ± 0.33 |
| Total SO2 (mg/L) | 57 ± 8 | 56 ± 1 | 52 ± 2 | 48 ± 0 |
| Alcoholic content (% v/v) | 15.30 ± 0.00 | 15.30 ± 0.00 | 15.40 ± 0.10 | 15.20 ± 0.10 |
| Strains Inoculated | Stage | Number of Isolates | Interdelta PCR | Sequentiation | Lysine Growth |
|---|---|---|---|---|---|
| Sc01 | 48 h | 6 | Sc01 | - | - |
| Sc01 | 48 h | 6 | Sc01 | - | - |
| Sc02 | 48 h | 6 | Sc02 | - | - |
| Sc02 | 48 h | 6 | Sc02 | - | - |
| Wa | 48 h | 6 | - | W. anomalus | Positive |
| Wa | 48 h | 6 | - | W. anomalus | Positive |
| Sc01 | EF | 3 | Sc01 | - | - |
| Sc01 | EF | 3 | Sc01 | - | - |
| Sc02 | EF | 3 | Sc02 | - | - |
| Sc02 | EF | 3 | Sc02 | - | - |
| Wa/Sc01 | EF | 3 | Sc01 | - | Negative |
| Wa/Sc01 | EF | 3 | Sc01 | - | Negative |
| Wa/Sc02 | EF | 3 | Sc02 | - | Negative |
| Wa/Sc02 | EF | 3 | Sc02 | - | Negative |
| Code | IUPAC Name | Sc01 | Wa/Sc01 | Sc02 | Wa/Sc02 |
|---|---|---|---|---|---|
| Acetate Esters | |||||
| A1 | 2-methylpropyl acetate | 0.182 ± 0.035 | 0.211 ± 0.014 | 0.224 ± 0.073 | 0.166 ± 0.058 |
| A2 | 3-methylbutyl acetate | 27.231 ± 5.201 | 15.494 ± 1.771 | 27.863 ± 1.672 | 17.186 ± 6.791 |
| A3 | Hexyl acetate | 2.602 ± 0.026 b | 0.803 ± 0.355 a | 2.742 ± 0.544 b | 0.645 ± 0.237 a |
| A4 | 2-phenylethyl acetate | 1.955 ± 0.488 | 1.040 ± 0.268 | 1.761 ± 0.165 | 1.072 ± 0.005 |
| ∑ A | Acetate esters sum | 31.970 ± 5.750 | 17.548 ± 2.380 | 32.589 ± 2.308 | 19.070 ± 7.082 |
| Ethyl Esters | |||||
| Et1 | Ethyl acetate | 16.083 ± 3.722 | 44.738 ± 4.996 | 31.742 ± 23.043 | 41.159 ± 6.942 |
| Et2 | Ethyl propanoate | 0.151 ± 0.048 | 0.248 ± 0.039 | 0.194 ± 0.071 | 0.257 ± 0.066 |
| Et3 | Ethyl 2-methylpropanoate | 0.113 ± 0.018 | 0.126 ± 0.064 | 0.078 ± 0.003 | 0.121 ± 0.022 |
| Et4 | Ethyl 3-methylbutanoate | 0.190 ± 0.040 | 0.145 ± 0.072 | 0.144 ± 0.050 | 0.140 ± 0.044 |
| Et5 | Ethyl butanoate | 1.310 ± 0.027 | 1.180 ± 0.116 | 1.505 ± 0.004 | 1.138 ± 0.298 |
| Et6 | Ethyl hexanoate | 25.543 ± 0.213 | 17.227 ± 4.980 | 27.064 ± 5.198 | 14.448 ± 6.022 |
| Et7 | Ethyl heptanoate | 0.209 ± 0.054 | 0.665 ± 0.539 | 0.224 ± 0.064 | 0.183 ± 0.060 |
| Et8 | Ethyl octanoate | 25.044 ± 5.255 ab | 14.280 ± 7.548 ab | 34.298 ± 1.284 b | 11.130 ± 5.385 a |
| Et9 | Ethyl decanoate | 9.447 ± 2.899 | 7.016 ± 3.277 | 16.735 ± 7.066 | 4.027 ± 1.403 |
| Et10 | Ethyl dec-9-enoate | 3.849 ± 0.996 ab | 3.354 ± 1.423 ab | 7.493 ± 0.525 b | 3.059 ± 1.057 a |
| Et11 | Ethyl dodecanoate | 0.496 ± 0.156 | 0.395 ± 0.108 | 0.639 ± 0.203 | 0.300 ± 0.021 |
| ∑ Et | Ethyl esters sum | 82.436 ± 12.946 | 89.374 ± 22.083 | 120.118 ± 37.404 | 75.964 ± 21.322 |
| Acids | |||||
| Ac1 | 2-methylpropanoic acid | 0.220 ± 0.013 | 0.257 ± 0.080 | 0.197 ± 0.008 | 0.264 ± 0.008 |
| Ac2 | 3-methylbutanoic acid | 0.773 ± 0.102 | 3.778 ± 4.062 | 0.600 ± 0.132 | 4.056 ± 4.547 |
| Ac3 | Octanoic acid | 3.918 ± 1.191 | 2.715 ± 1.015 | 3.937 ± 0.012 | 2.079 ± 0.089 |
| Ac4 | Nonanoic acid | 0.210 ± 0.114 | 0.242 ± 0.017 | 0.304 ± 0.083 | 0.221 ± 0.021 |
| Ac5 | Dec-9-enoic acid | 0.330 ± 0.082 | 0.233 ± 0.039 | 0.418 ± 0.142 | 0.209 ± 0.004 |
| Ac6 | Decanoic acid | 2.159 ± 0.375 | 1.709 ± 0.238 | 2.471 ± 0.835 | 0.837 ± 0.043 |
| ∑ Ac | Acids sum | 7.611 ± 1.851 | 8.934 ± 5.213 | 7.927 ± 0.923 | 7.666 ± 4.381 |
| Alcohols | |||||
| C6-1 | Hexan-1-ol | 2.774 ± 0.269 | 3.038 ± 0.265 | 2.941 ± 0.023 | 3.085 ± 0.223 |
| Alc1 | Propan-1-ol | 0.292 ± 0.016 a | 0.489 ± 0.082 ab | 0.345 ± 0.053 ab | 0.602 ± 0.088 b |
| Alc2 | 2-methylpropan-1-ol | 5.207 ± 0.415 | 5.866 ± 0.318 | 5.518 ± 0.070 | 5.932 ± 0.015 |
| Alc3 | Butan-1-ol | 0.105 ± 0.002 a | 0.155 ± 0.006 b | 0.164 ± 0.002 b | 0.179 ± 0.011 b |
| Alc4 | 3-methylbutan-1-ol | 126.497 ± 6.016 | 127.159 ± 2.544 | 139.490 ± 23.582 | 144.977 ± 11.145 |
| Alc5 | (3S)-3-methylpentan-1-ol | 0.394 ± 0.085 | 0.343 ± 0.039 | 0.416 ± 0.180 | 0.385 ± 0.058 |
| Alc6 | 4-methylpentan-1-ol | 0.135 ± 0.030 ab | 0.094 ± 0.011 a | 0.217 ± 0.043 b | 0.109 ± 0.002 ab |
| Alc7 | Heptan-1-ol | 0.312 ± 0.080 | 0.151 ± 0.057 | 0.268 ± 0.045 | 0.168 ± 0.026 |
| Alc8 | 2-phenylethanol | 19.437 ± 0.146 ab | 11.963 ± 0.867 a | 24.868 ± 2.262 b | 15.763 ± 3.893 ab |
| ∑ Alc | Alcohols sum | 155.154 ± 6.286 | 149.259 ± 3.311 | 174.227 ± 25.924 | 171.201 ± 7.671 |
| Aldehydes | |||||
| Ald1 | Benzaldehyde | 0.221 ± 0.002 | 0.330 ± 0.007 | 0.323 ± 0.054 | 0.356 ± 0.041 |
| Ald2 | Acetaldehyde | 0.107 ± 0.019 | 0.145 ± 0.020 | 0.142 ± 0.031 | 0.144 ± 0.028 |
| ∑ Ald | Aldehydes sum | 0.328 ± 0.021 | 0.475 ± 0.026 | 0.465 ± 0.085 | 0.501 ± 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Modulation of the Aromatic Profile of Verdejo Wine through Sequential Inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae. Fermentation 2023, 9, 977. https://doi.org/10.3390/fermentation9110977
López-Enríquez L, Vila-Crespo J, Rodríguez-Nogales JM, Fernández-Fernández E, Ruipérez V. Modulation of the Aromatic Profile of Verdejo Wine through Sequential Inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae. Fermentation. 2023; 9(11):977. https://doi.org/10.3390/fermentation9110977
Chicago/Turabian StyleLópez-Enríquez, Lorena, Josefina Vila-Crespo, José Manuel Rodríguez-Nogales, Encarnación Fernández-Fernández, and Violeta Ruipérez. 2023. "Modulation of the Aromatic Profile of Verdejo Wine through Sequential Inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae" Fermentation 9, no. 11: 977. https://doi.org/10.3390/fermentation9110977
APA StyleLópez-Enríquez, L., Vila-Crespo, J., Rodríguez-Nogales, J. M., Fernández-Fernández, E., & Ruipérez, V. (2023). Modulation of the Aromatic Profile of Verdejo Wine through Sequential Inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae. Fermentation, 9(11), 977. https://doi.org/10.3390/fermentation9110977