
Valorisation of CO2 into Value-Added Products via Microbial Electrosynthesis (MES) and Electro-Fermentation Technology

 ,
, 
 ,
,  ,
,  ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
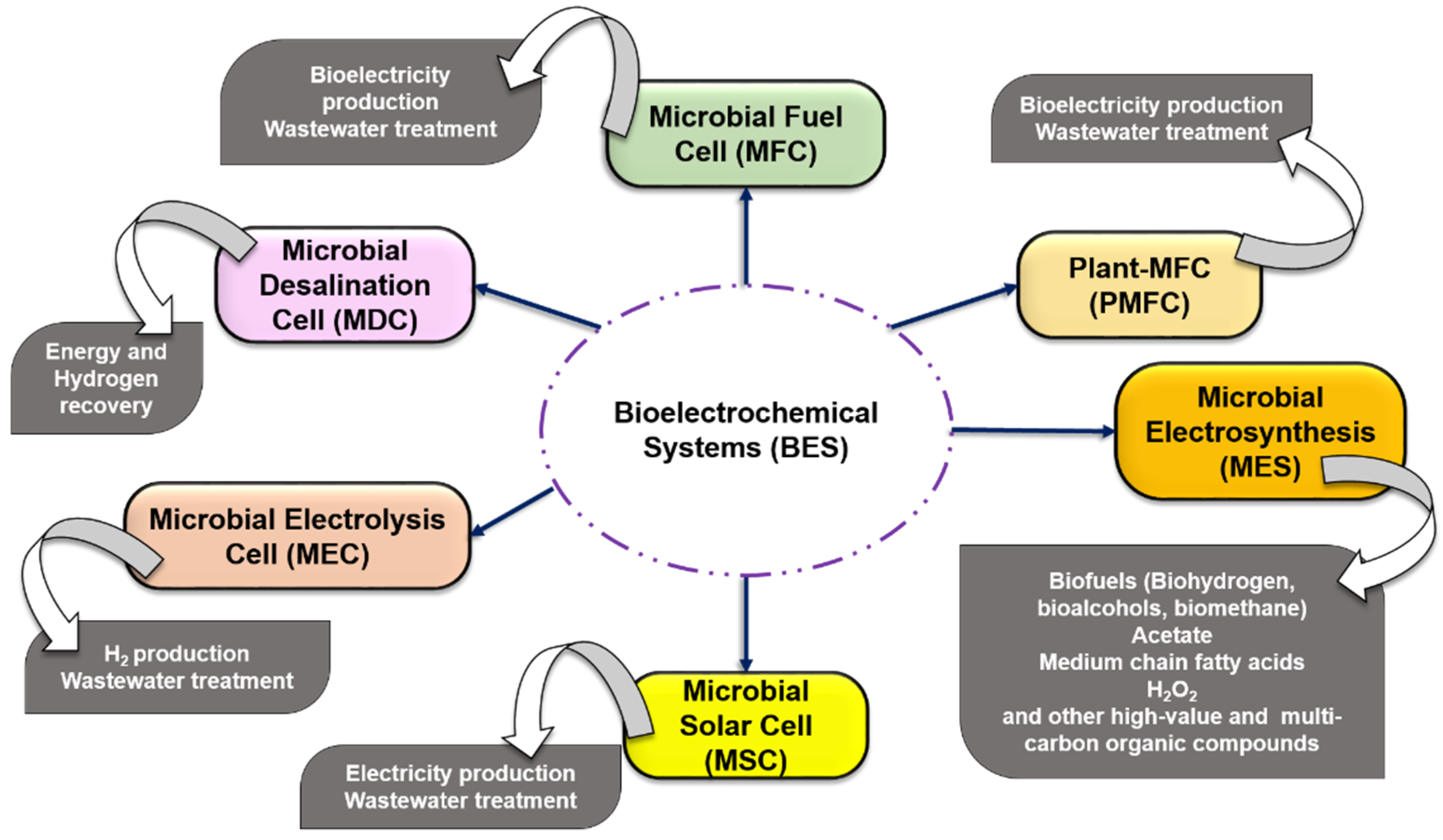
2. Bioelectrochemical System (BES)
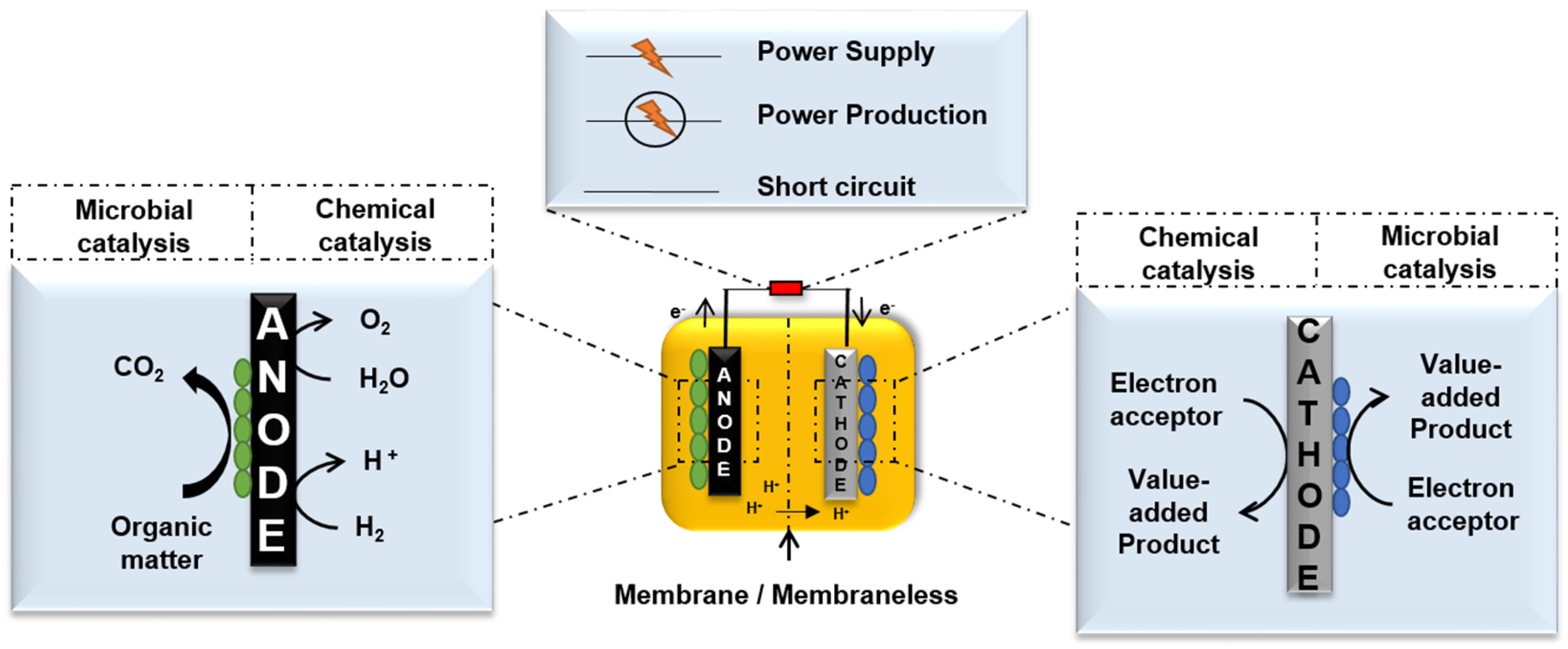
2.1. Transmission of Electrons at the Anode
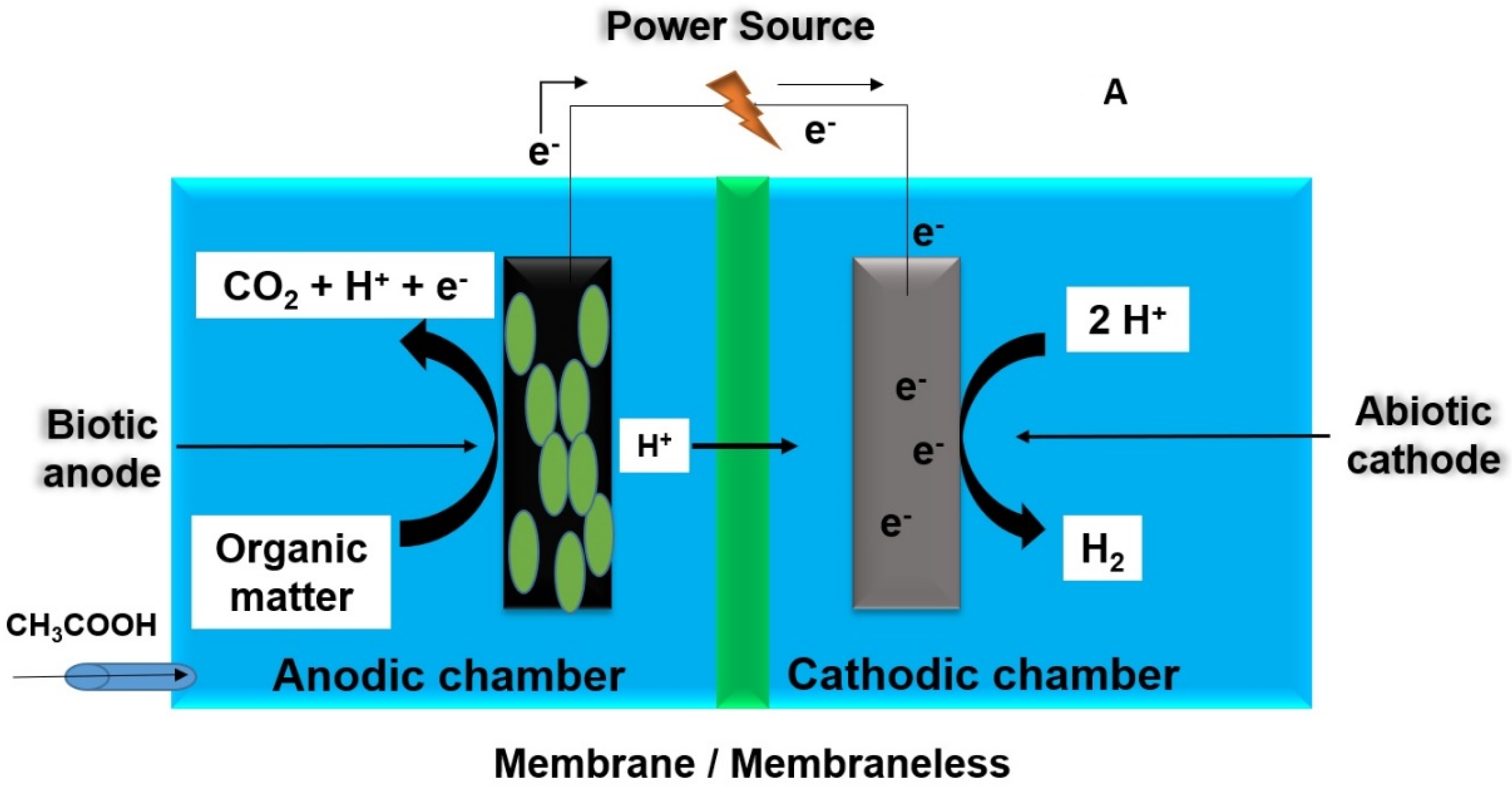
2.2. Transmission of Electrons at the Cathode
2.3. Electrosynthesis Assisted by Microbes
2.4. Electroactive Microbes and Extracellular Electron Transfer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microbe | Substrate | Product | Mechanisms of EET | Genetic Modification of Host | Yield (Y) and/or Titre (T) | Ref. |
|---|---|---|---|---|---|---|
| Anodic Electro-Fermentation | ||||||
| Shewanella oneidensis | Glucose | Acetate | Direct electron transfer (ET) | Introduction of E. coli galactose permease (galP) and glucose kinase (glk) genes. | No | [82] |
| Glycerol | Ethanol; Acetate | Direct ET | Introduction of Zymomonas mobilis ethanol production module and E. coli glycerol utilisation module | Y = 52% ± 4% T = 1.28 ± 0.02 g L−1; Y = 13% ± 6% T = 0.29 ± 0.08 g L−1 | [83] | |
| Lactate | Acetoin | Direct ET | Introduction of Bacillus subtilis acetolactate decarboxylase and acetolactate synthase; Deletion of genomic prophages; Knockout of the phosphotransacetylase and acetate kinase genes | Y = 52% T = 0.24 g L−1 Productivity = 0.91 mg h−1 | [84] | |
| Actinobacillus succinogenes | Glycerol | Succinate; Acetate; Formate | Neutral red mediated ET | Transmembrane mediator transport was improved by atmospheric and room temperature plasma mutagenesis. | Y = 68% T = 23.92 ± 0.08 g L−1; Y = 7% T = 1.15 ± 0.77 g L−1; Y = 19% T = 2.57 ± 0.11 g L−1 | [85] |
| Klebsiella pneumoniae | Glycerol | Acetate; 3-Hydroxypropionic acid; 1,3-Propanediol | Direct ET | Not modified | T = 21.7 mM; T = 7.6 mM; T = 45.5 mM | [86] |
| Clostridium cellobioparum + Geobacter sulfurreducens | Glycerol | Ethanol | Direct ET | Adaptive evolution of C. cellobioparum | T = 10 g L−1 | [87] |
| Propionibacterium freudenreichii | Glycerol & propionate; Only Propionate; Lactate & propionate | Acetate | Ferricyanide mediated ET | Enhanced bacterial growth & substrate consumption | Y = 56% T = 0.38 g L−1; Y = 68% T = 0.47 g L−1; Y = 60% T = 0.42 g L−1 | [81] |
| Enterobacter aerogens NBRC 12010 | Glycerol | Ethanol; Hydrogen | Thionine mediated ET | Increased glycerol consumption | Y = 92% T = 3.93 g L−1; Y = 74% T = 0.14 g L−1 | [81] |
| Cellulomonas uda + Geobacter sulfurreducens | Cellobiose | Ethanol | Direct ET | Adaptive evolution and deleted G. sulfurreducens hydrogenase gene | No | [88] |
| Ralstonia eutropha | Fructose | Poly hydroxy butyrate | poly (2-methacryloyloxyethyl phosphorylcholine-co-vinyl-ferrocene)-mediated ET | Not modified | No | [89] |
| Escherichia coli | Lactate | Acetate; Ethanol | Direct ET | Introduction of S. oneidensis MR-1 Mtr pathway | Productivity = 0.038 mM day−1; T = 40 ± 3 μM | [90] |
| Escherichia coli + Methano bacterium formicicum | Glycerol | Ethanol; Acetate | Methylene blue-mediated ET | Cyt c introduction—CymA, MtrA and STC from S. oneidensis | Y = 35% ± 5% T = 55.25 ± 7.76 g L−1 Productivity = 12.12 ± 1.70 mg h−1; Y = 20% ± 1% T = 40.75 ± 2.37 g L−1 Productivity = 8.94 ± 0.52 mg h−1 | [91] |
| Pseudomonas putida F1 | Glucose | 2-Keto- gluconate | 7 different mediators-based mediated ET | Not modified | Y = 90% ± 2% T = 1.47 ± 0.27 g L−1 Productivity = 1.75 ± 0.33 mg h−1 | [92] |
| Pseudomonas putida | Glucose | 2-ketoglu conic acid | Direct ET | Overexpression of periplasmic glucose dehydrogenase | Productivity = 0.25 ± 0.02 mmol gCDW−1 h−1 | [93] |
| Corynebacterium glutamicum + Zymomonas mobilis | Glucose Glucose | l-lysine; Ethanol | Ferricyanide-mediated ET Methyl naphthoquinone, humic acid, methylene blue, neutral red, 1,4-riboflavin, butane-disulfonate and tempol-mediated ET | Feedback deregulated mutant and overexpressed redox-related genes—ZMO0899, ZMO1116 and ZMO1885 | T = 2.9 Mm Productivity = 0.2 mmol L−1 h−1 Bioelectricity generation = 2.0 m Wm−2; T = ~ 42.5 g L−1 | [94] |
| Microbial electrosynthesis (MES) | ||||||
| Clostridium pasteurianumDSM 525 | Glucose; Glycerol | Butanol; 1,3-propanediol | Direct ET | Not modified | T = 1.00 ± 0.20 g L−1; T = 4.74 g L−1 | [47] |
| Geobacter sulfurreducens | CO2; Succinate | Glycerol | Direct ET | Not modified | T = 8.7 ± 0.3 mM | [95] |
| Sporomusa ovate + Methanococcus maripaludis | CO2 | Acetate; CH4 | H2-mediated ET | Not modified | T = 0.2 to 0.3 mM; T = 0.2 to 0.3 mM; | [34] |
| Sporomusa ovate | CO2 | Acetate | Direct ET | Not modified | No | [96] |
| Shewanella onedensis MR-1 | Acetoin | 2,3-butanediol | Direct ET | Heterologous expression of butanediol dehydrogenase (Bdh) gene along with a light-driven proton pump and hydrogenase gene ΔhyaBΔhydA knockout | T = 0.03 mM | [97] |
| Clostridium pasteurianum | Glycerol | 1,3-propanediol; n-butanol | Neutral red and brilliant blue-mediated ET | Not modified | Y = 0.41 mol mol−1 glycerol in brilliant blue-mediated ET; Y = 0.35 mol mol−1 glycerol in Neutral red-mediated ET | [98] |
| Saccharomyces cerevisiae | Dhea | 7α–OH–DHEA | Neutral red and 7α-hydroxylase-mediated ET | Heterogenous expression of 7α-hydroxylase | T = 288.6 ± 7.8 mg L−1 | [99] |
| Ralstonia eutropha | CO2 | Iso- propanol | H2-mediated ET | Not modified | T = 216 mg L−1 | [100] |
| 3-methyl-1-butanol; Isobutanol | Formate mediated ET | Introduction of genes alsS, ilvC, ilvD, kivd and yqhD; Knockout of polyhydroxy butyrate synthesis gene cluster (phaC1, phaA and phaB1) | Both depicted a titre of 140 mg L−1 | [79] | ||
| Xanthobacter autotrophicus | N2 and H2O | NH3 | H2-mediated ET | Not modified | T = ~ 0.8 mM | [101] |
2.5. Increasing Electrode Interaction
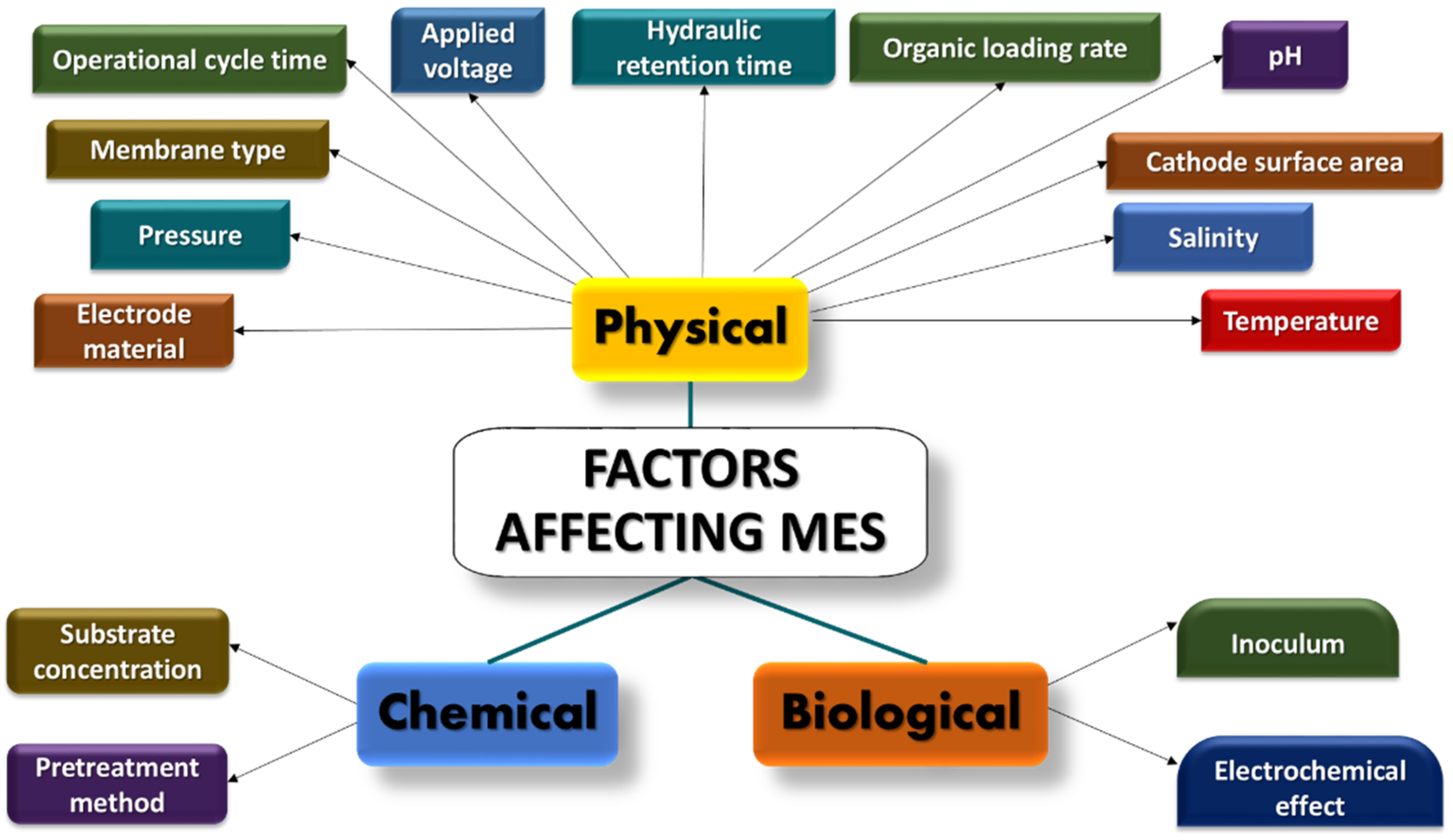
3. Techniques for Improving MES Performance
3.1. Cathode Fabrication
3.2. Anode Fabrication
3.3. MES and Gene Manipulation
3.3.1. Modification of Pathways for Generating Value-Added Products
3.3.2. Host Cell Selection
Escherichia coli
Clostridium ljungdahlii
Cyanobacteria (Cyano)
4. MES Allows Biocatalysts to Utilise CO2 and Generate Electricity
5. Diverse Products Obtained from CO2
5.1. H2 Production via MES
5.2. Acetate Production via MES
| Cathode Material | Ecathode (V vs. SHE) | Current Density (A m−2) | Volumetric Production Rate (g L−1 day−1) | Maximum Acetate Titre (g L−1) | Coulombic Efficiency (%) | Ref. |
|---|---|---|---|---|---|---|
| 12 mg cm−2 Mo2C | −0.85 | −5.2 | 0.19 | 5.72 | 64 | [168] |
| NanoWeb-RVC | −0.85 | −37 | 0.03 | 1.65 | 70 | [61] |
| Graphene-nickel foam | −0.85 | −10.2 | 0.19 | 5.46 | 70 | [177] |
| VITO-CoRE™ electrode fabricated with activated carbon | −0.6 | −0.069 | 0.14 | 4.97 | 45.5 | [178] |
| Carbon felt (CF) | −1.26 | −5.0 | 0.06 | 1.29 | 58 | [179] |
| −0.903 | −2.96 | −0.14 | 4.7 | 89.5 | [180] | |
| CF and stainless steel | −0.78 | −15 | 0.14 | 2 | 22.5 | [7] |
| −0.9 | −10 | 1.3 | 0.6 | 40 | [181] | |
| RVC-EPD | −0.85 | −102 | - | 11 | 100 | [182] |
| rGO-CF | −0.85 | −4.9 | 0.17 | 7.1 | 77 | [177] |
| CF with fluidised GAC (16 g L−1) | −0.85 | −4.08 | 0.14 | 3.9 | 65 | [183] |
| Graphite stick-graphite felt | −0.8 | −20 | 0.14 | 8.28 | - | [184] |
| Graphite granules | −0.6 | - | 1.0 | 10.5 | 69 | [185] |
5.3. Formic Acid Production via MES
5.4. Syngas Production via MES
6. MES Enhancement
7. Downstream Processes Involved in MES
7.1. Process for Conventional Separation
7.2. Pressure and Concentration-Driven Separation Process
7.3. Process of Reactive Extraction
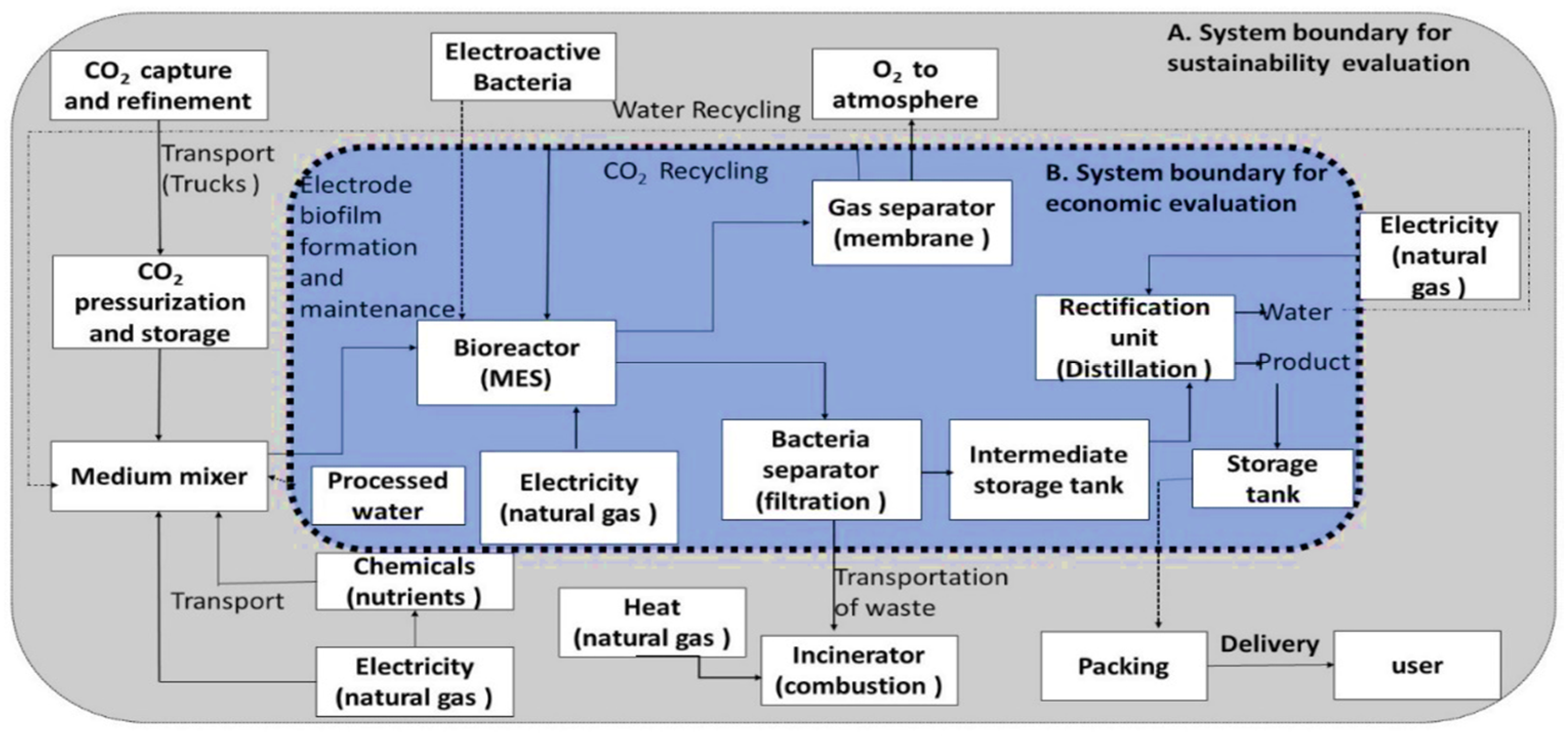
8. Advancing towards Sustainable Development of MES
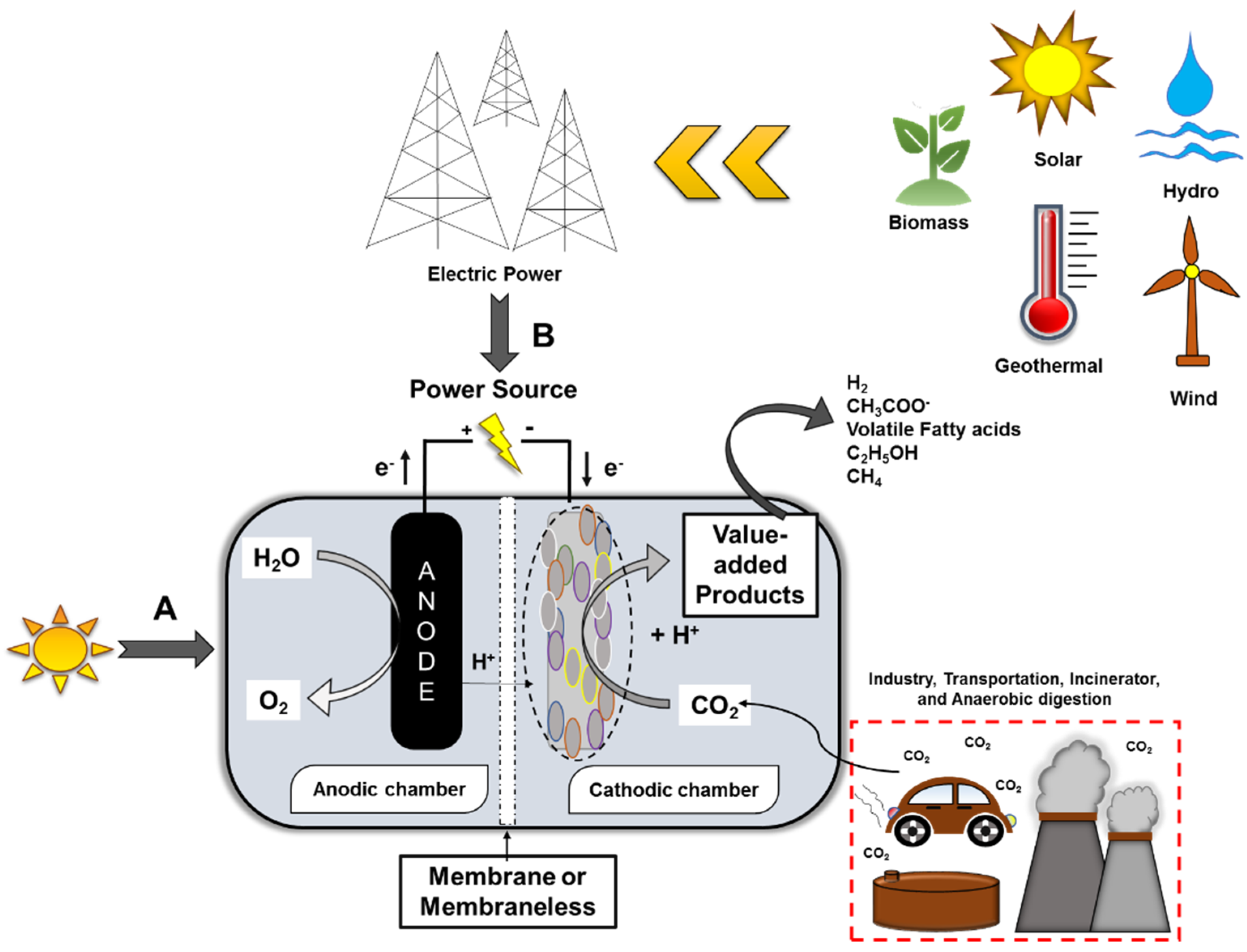
8.1. Uses of Renewable Sources of Energy and Integrated Hybrid Systems
8.2. Electronic Design and Energy Storage for MES
8.3. Commercialisation of MES
9. Prospects
10. Conclusions
- MES and Electro-fermentation are innovations that not only aim at minimising the emissions of greenhouse gases but also contributes to low manufacturing prices boosting the circular bioeconomy, offering a practical solution to lighten the ever-expanding global issues.
- Both these processes provide a plethora of premium products like biofuels, bioenergy and can also perform concurrent valorisation of CO2 and wastewater.
- Recently, there have been multiple strategies in optimising the MES process and improving its efficiency, including treating the cultivation substrate to include adequate nutrients, enhancing the architecture and feeding conditions of the reactor, enriching the inoculum mix culture, running reactors in optimised conditions and also boosting the microbial interactions by spatially organising the cathode.
- However, significant challenges need to be tackled before commercialisation. Both these technologies, in general, are still far from practical application and further research into basic operational variables, long-term stability, continuous production, modelling, repeatability and scalability is still necessary.
- Overall, this review paper promotes further studies on promising microbial aspirants to aid advancement in this emergent field, with the subsequent aim of bringing this sustainable technology one step closer to real-world applications.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inglezakis, V.J. Extraterrestrial Environment. Environ. Dev. Basic Princ. Hum. Act. Environ. Implic. 2016, 453–498. [Google Scholar] [CrossRef]
- Zhang, S.; Jiang, J.; Wang, H.; Li, F.; Hua, T.; Wang, W. A review of microbial electrosynthesis applied to carbon dioxide capture and conversion: The basic principles, electrode materials, and bioproducts. J. CO2 Util. 2021, 51, 101640. [Google Scholar] [CrossRef]
- Fawzy, S.; Osman, A.I.; Doran, J.; Rooney, D.W. Strategies for mitigation of climate change: A review. Environ. Chem. Lett. 2020, 18, 2069–2094. [Google Scholar] [CrossRef]
- Brack, D. Background Analytical Study Forests and Climate Change; United Nations Forum on Forest: New York, NY, USA, 2019. [Google Scholar]
- Gielen, D.; Boshell, F.; Saygin, D.; Bazilian, M.D.; Wagner, N.; Gorini, R. The role of renewable energy in the global energy transformation. Energy Strateg. Rev. 2019, 24, 38–50. [Google Scholar] [CrossRef]
- Perera, F. Pollution from fossil-fuel combustion is the leading environmental threat to global pediatric health and equity: Solutions exist. Int. J. Environ. Res. Public Health 2018, 15, 16. [Google Scholar] [CrossRef]
- Bajracharya, S.; Vanbroekhoven, K.; Buisman, C.J.N.; Strik, D.P.B.T.B.; Pant, D. Bioelectrochemical conversion of CO2 to chemicals: CO2 as a next-generation feedstock for electricity-driven bioproduction in batch and continuous modes. Faraday Discuss. 2017, 202, 433–449. [Google Scholar] [CrossRef] [PubMed]
- Dessì, P.; Rovira-Alsina, L.; Sánchez, C.; Dinesh, G.K.; Tong, W.; Chatterjee, P.; Tedesco, M.; Farràs, P.; Hamelers, H.M.V.; Puig, S. Microbial electrosynthesis: Towards sustainable biorefineries for the production of green chemicals from CO2 emissions. Biotechnol. Adv. 2021, 46, 107675. [Google Scholar] [CrossRef] [PubMed]
- Ben-Iwo, J.; Manovic, V.; Longhurst, P. Biomass resources and biofuels potential for the production of transportation fuels in Nigeria. Renew. Sustain. Energy Rev. 2016, 63, 172–192. [Google Scholar] [CrossRef]
- Kumaravel, V.; Bartlett, J.; Pillai, S.C. Photoelectrochemical Conversion of Carbon Dioxide (CO2) into Fuels and Value-Added Products. ACS Energy Lett. 2020, 486–519. [Google Scholar] [CrossRef]
- Kumar, M. Social, Economic, and Environmental Impacts of Renewable Energy Resources. In Wind Solar Hybrid Renewable Energy System; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Cabau-Peinado, O.; Straathof, A.J.J.; Jourdin, L. A General Model for Biofilm-Driven Microbial Electrosynthesis of Carboxylates From CO2. Front. Microbiol. 2021, 12, 669218. [Google Scholar] [CrossRef]
- Badwal, S.P.S.; Giddey, S.S.; Munnings, C.; Bhatt, A.I.; Hollenkamp, A.F. Emerging electrochemical energy conversion and storage technologies. Front. Chem. 2014, 2, 79. [Google Scholar] [CrossRef] [PubMed]
- Enescu, D.; Chicco, G.; Porumb, R.; Seritan, G. Thermal energy storage for grid applications: Current status and emerging trends. Energies 2020, 13, 340. [Google Scholar] [CrossRef]
- Kondaveeti, S.; Abu-Reesh, I.M.; Mohanakrishna, G.; Bulut, M.; Pant, D. Advanced Routes of Biological and Bio-electrocatalytic Carbon Dioxide (CO2) Mitigation Toward Carbon Neutrality. Front. Energy Res. 2020, 8, 94. [Google Scholar] [CrossRef]
- Jourdin, L. Microbial Electrosynthesis from Carbon Dioxide: Performance Enhancement and Elucidation of Mechanisms; The University of Queensland: Brisbane, Australia, 2016. [Google Scholar] [CrossRef]
- Christodoulou, X.; Okoroafor, T.; Parry, S.; Velasquez-Orta, S.B. The use of carbon dioxide in microbial electrosynthesis: Advancements, sustainability and economic feasibility. J. CO2 Util. 2017, 18, 390–399. [Google Scholar] [CrossRef]
- Jourdin, L.; Burdyny, T. Microbial Electrosynthesis: Where Do We Go from Here? Trends Biotechnol. 2021, 39, 359–369. [Google Scholar] [CrossRef]
- Das, S.; Diels, L.; Pant, D.; Patil, S.A.; Ghangrekar, M.M. Review—Microbial Electrosynthesis: A Way Towards The Production of Electro-Commodities Through Carbon Sequestration with Microbes as Biocatalysts. J. Electrochem. Soc. 2020, 167, 155510. [Google Scholar] [CrossRef]
- Vassilev, I.; Hernandez, P.A.; Batlle-Vilanova, P.; Freguia, S.; Krömer, J.O.; Keller, J.; Ledezma, P.; Virdis, B. Microbial Electrosynthesis of Isobutyric, Butyric, Caproic Acids, and Corresponding Alcohols from Carbon Dioxide. ACS Sustain. Chem. Eng. 2018, 6, 8485–8493. [Google Scholar] [CrossRef]
- Shin, H.J.; Jung, K.A.; Nam, C.W.; Park, J.M. A genetic approach for microbial electrosynthesis system as biocommodities production platform. Bioresour. Technol. 2017, 245, 1421–1429. [Google Scholar] [CrossRef]
- Ammam, F.; Tremblay, P.L.; Lizak, D.M.; Zhang, T. Effect of tungstate on acetate and ethanol production by the electrosynthetic bacterium Sporomusa ovata. Biotechnol. Biofuels 2016, 9, 163. [Google Scholar] [CrossRef]
- Jiang, Y.; Jianxiong Zeng, R. Expanding the product spectrum of value-added chemicals in microbial electrosynthesis through integrated process design—A review. Bioresour. Technol. 2018, 269, 503–512. [Google Scholar] [CrossRef]
- Tremblay, P.L.; Zhang, T. Electrifying microbes for the production of chemicals. Front. Microbiol. 2015, 6, 201. [Google Scholar] [CrossRef]
- Wood, J.C.; Grové, J.; Marcellin, E.; Heffernan, J.K.; Hu, S.; Yuan, Z.; Virdis, B. Strategies to improve the viability of a circular carbon bioeconomy-A techno-economic review of microbial electrosynthesis and gas fermentation. Water Res. 2021, 201, 117306. [Google Scholar] [CrossRef]
- Syed, Z.; Sogani, M.; Dongre, A.; Kumar, A.; Sonu, K.; Sharma, G.; Gupta, A.B. Bioelectrochemical systems for environmental remediation of estrogens: A review and way forward. Sci. Total Environ. 2021, 780, 146544. [Google Scholar] [CrossRef]
- Pant, D.; Singh, A.; van Bogaert, G.; Olsen, S.I.; Nigam, P.S.; Diels, L.; Vanbroekhoven, K. Bioelectrochemical systems (BES) for sustainable energy production and product recovery from organic wastes and industrial wastewaters. RSC Adv. 2012, 2, 1248–1263. [Google Scholar] [CrossRef]
- Bajracharya, S.; Sharma, M.; Mohanakrishna, G.; Dominguez Benneton, X.; Strik, D.P.B.T.B.; Sarma, P.M.; Pant, D. An overview on emerging bioelectrochemical systems (BESs): Technology for sustainable electricity, waste remediation, resource recovery, chemical production and beyond. Renew. Energy 2016, 98, 153–170. [Google Scholar] [CrossRef]
- Shaikh, R.; Rizvi, A.; Quraishi, M.; Pandit, S.; Mathuriya, A.S.; Gupta, P.K.; Singh, J.; Prasad, R. Bioelectricity production using plant-microbial fuel cell: Present state of art. S. African J. Bot. 2020. [Google Scholar] [CrossRef]
- Zheng, T.; Li, J.; Ji, Y.; Zhang, W.; Fang, Y.; Xin, F.; Dong, W.; Wei, P.; Ma, J.; Jiang, M. Progress and Prospects of Bioelectrochemical Systems: Electron Transfer and Its Applications in the Microbial Metabolism. Front. Bioeng. Biotechnol. 2020, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Chen, G. Factors affecting the effectiveness of bioelectrochemical system applications: Data synthesis and meta-analysis. Batteries 2018, 4, 34. [Google Scholar] [CrossRef]
- Kadier, A.; Simayi, Y.; Abdeshahian, P.; Azman, N.F.; Chandrasekhar, K.; Kalil, M.S. A comprehensive review of microbial electrolysis cells (MEC) reactor designs and configurations for sustainable hydrogen gas production. Alexandria Eng. J. 2016, 55, 427–443. [Google Scholar] [CrossRef]
- Anukam, A.; Mohammadi, A.; Naqvi, M.; Granström, K. A review of the chemistry of anaerobic digestion: Methods of accelerating and optimizing process efficiency. Processes 2019, 7, 504. [Google Scholar] [CrossRef]
- Kracke, F.; Wong, A.B.; Maegaard, K.; Deutzmann, J.S.; Hubert, M.K.A.; Hahn, C.; Jaramillo, T.F.; Spormann, A.M. Robust and biocompatible catalysts for efficient hydrogen-driven microbial electrosynthesis. Commun. Chem. 2019, 2, 1–9. [Google Scholar] [CrossRef]
- Ucar, D.; Zhang, Y.; Angelidaki, I. An overview of electron acceptors in microbial fuel cells. Front. Microbiol. 2017, 8, 643. [Google Scholar] [CrossRef]
- Cardeña, R.; Cercado, B.; Buitrón, G. Microbial Electrolysis Cell for Biohydrogen Production. In Biohydrogen; Elsevier: Paris, France, 2019; pp. 159–185. [Google Scholar] [CrossRef]
- Rivera, I.; Schröder, U.; Patil, S.A. Microbial electrolysis for biohydrogen production: Technical aspects and scale-up experiences. In Biomass, Biofuels, Biochemicals: Microbial Electrochemical Technology: Sustainable Platform for Fuels, Chemicals and Remediation; Elsevier: Gurgaon, India, 2018; pp. 871–898. [Google Scholar] [CrossRef]
- Caizán-Juanarena, L.; Borsje, C.; Sleutels, T.; Yntema, D.; Santoro, C.; Ieropoulos, I.; Soavi, F.; Ter Heijne, A. Combination of bioelectrochemical systems and electrochemical capacitors: Principles, analysis and opportunities. Biotechnol. Adv. 2020, 39, 107456. [Google Scholar] [CrossRef] [PubMed]
- Rabaey, K.; Rozendal, R.A. Microbial electrosynthesis-Revisiting the electrical route for microbial production. Nat. Rev. Microbiol. 2010, 8, 706–716. [Google Scholar] [CrossRef] [PubMed]
- Drendel, G.; Mathews, E.R.; Semenec, L.; Franks, A.E. Microbial Fuel Cells, Related Technologies, and Their Applications. Appl. Sci. 2018, 8, 2384. [Google Scholar] [CrossRef]
- Borole, A.P.; Reguera, G.; Ringeisen, B.; Wang, Z.W.; Feng, Y.; Kim, B.H. Electroactive biofilms: Current status and future research needs. Energy Environ. Sci. 2011, 4, 4813–4834. [Google Scholar] [CrossRef]
- ter Heijne, A.; Pereira, M.A.; Pereira, J.; Sleutels, T. Electron Storage in Electroactive Biofilms. Trends Biotechnol. 2021, 39, 34–42. [Google Scholar] [CrossRef]
- Patil, S.A.; Hägerhäll, C.; Gorton, L. Electron transfer mechanisms between microorganisms and electrodes in bioelectrochemical systems. Bioanal. Rev. 2014, 1, 71–129. [Google Scholar] [CrossRef]
- Mao, L.; Verwoerd, W.S. Selection of organisms for systems biology study of microbial electricity generation: A review. Int. J. Energy Environ. Eng. 2013, 4, 1–18. [Google Scholar] [CrossRef]
- Kumar, R.; Singh, L.; Zularisam, A.W.; Hai, F.I. Microbial fuel cell is emerging as a versatile technology: A review on its possible applications, challenges and strategies to improve the performances. Int. J. Energy Res. 2018, 42, 369–394. [Google Scholar] [CrossRef]
- Pawar, A.A.; Karthic, A.; Lee, S.; Pandit, S.; Jung, S.P. Microbial electrolysis cells for electromethanogenesis: Materials, configurations and operations. Environ. Eng. Res. 2022, 27, 53–57. [Google Scholar] [CrossRef]
- Choi, O.; Sang, B.I. Extracellular electron transfer from cathode to microbes: Application for biofuel production. Biotechnol. Biofuels 2016, 9, 11. [Google Scholar] [CrossRef]
- Aryal, N.; Ammam, F.; Patil, S.A.; Pant, D. An overview of cathode materials for microbial electrosynthesis of chemicals from carbon dioxide. Green Chem. 2017, 19, 5748–5760. [Google Scholar] [CrossRef]
- Werth, C.J.; Yan, C.; Troutman, J.P. Factors Impeding Replacement of Ion Exchange with (Electro)Catalytic Treatment for Nitrate Removal from Drinking Water. ACS EST Eng. 2021, 1, 6–20. [Google Scholar] [CrossRef]
- Lim, S.S.; Kim, B.H.; Li, D.; Feng, Y.; Daud, W.R.W.; Scott, K.; Yu, E.H. Effects of Applied Potential and Reactants to Hydrogen-Producing Biocathode in a Microbial Electrolysis Cell. Front. Chem. 2018, 6, 318. [Google Scholar] [CrossRef] [PubMed]
- Ali Shah, F.; Mahmood, Q.; Maroof Shah, M.; Pervez, A.; Ahmad Asad, S. Microbial ecology of anaerobic digesters: The key players of anaerobiosis. Sci. World J. 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Spiess, S.; Kucera, J.; Seelajaroen, H.; Sasiain, A.; Thallner, S.; Kremser, K.; Novak, D.; Guebitz, G.M.; Haberbauer, M. Impact of Carbon Felt Electrode Pretreatment on Anodic Biofilm Composition in Microbial Electrolysis Cells. Biosensors 2021, 11, 170. [Google Scholar] [CrossRef]
- Dincer, I.; Zamfirescu, C. Sustainable Hydrogen Production; Elsevier Inc.: North York, ON, Canada, 2016; ISBN 9780128017487. [Google Scholar]
- Krishnan, S.; Kadier, A.; Fadhil Bin MD Din, M.; Nasrullah, M.; Najiha, N.N.; Taib, S.M.; Wahid, Z.A.; Li, Y.Y.; Qin, Y.; Pant, K.K.; et al. Application of bioelectrochemical systems in wastewater treatment and hydrogen production. In Delivering Low-Carbon Biofuels with Bioproduct Recovery; Elsevier: Gurgaon, India, 2021; pp. 31–44. [Google Scholar] [CrossRef]
- Hussain, S.; Aneggi, E.; Goi, D. Catalytic activity of metals in heterogeneous Fenton-like oxidation of wastewater contaminants: A review. Environ. Chem. Lett. 2021, 19, 2405–2424. [Google Scholar] [CrossRef]
- Hassan, M.; Olvera-Vargas, H.; Zhu, X.; Zhang, B.; He, Y. Microbial electro-Fenton: An emerging and energy-efficient platform for environmental remediation. J. Power Source 2019, 424, 220–244. [Google Scholar] [CrossRef]
- Song, T.; Wang, G.; Wang, H.; Huang, Q.; Xie, J. Experimental evaluation of the influential factors of acetate production driven by a DC power system via CO2 reduction through microbial electrosynthesis. Bioresour. Bioprocess. 2019, 6, 1–10. [Google Scholar] [CrossRef]
- Saheb-Alam, S.; Singh, A.; Hermansson, M.; Persson, F.; Schnürer, A.; Wilén, B.M.; Modin, O. Effect of start-up strategies and electrode materials on carbon dioxide reduction on biocathodes. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef]
- Thatikayala, D.; Min, B. Copper ferrite supported reduced graphene oxide as cathode materials to enhance microbial electrosynthesis of volatile fatty acids from CO2. Sci. Total Environ. 2021, 768, 144477. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.; Cho, S.K.; Sivagurunathan, P.; Anburajan, P.; Mahapatra, D.M.; Park, J.H.; Pugazhendhi, A. Insights into evolutionary trends in molecular biology tools in microbial screening for biohydrogen production through dark fermentation. Int. J. Hydrogen Energy 2018, 43, 19885–19901. [Google Scholar] [CrossRef]
- Jourdin, L.; Freguia, S.; Donose, B.C.; Chen, J.; Wallace, G.G.; Keller, J.; Flexer, V. A novel carbon nanotube modified scaffold as an efficient biocathode material for improved microbial electrosynthesis. J. Mater. Chem. A 2014, 2, 13093–13102. [Google Scholar] [CrossRef]
- Zhang, T.; Ghosh, D.; Tremblay, P.-L. Synthetic Biology Strategies to Improve Electron Transfer Rate at the Microbe–Anode Interface in Microbial Fuel Cells. Bioelectrochem. Interface Eng. 2019, 187–208. [Google Scholar] [CrossRef]
- Alfonta, L. Genetically Engineered Microbial Fuel Cells. Electroanalysis 2010, 22, 822–831. [Google Scholar] [CrossRef]
- Kerzenmacher, S. Engineering of microbial electrodes. In Advances in Biochemical Engineering/Biotechnology; Harnisch, F., Holtmann, D., Eds.; Springer: Cham, Switzerland, 2019; Volume 167, pp. 135–180. [Google Scholar] [CrossRef]
- Fruehauf, H.M.; Enzmann, F.; Harnisch, F.; Ulber, R.; Holtmann, D. Microbial Electrosynthesis—An Inventory on Technology Readiness Level and Performance of Different Process Variants. Biotechnol. J. 2020, 15, 2000066. [Google Scholar] [CrossRef]
- Simões, M.; Simões, L.C.; Vieira, M.J. A review of current and emergent biofilm control strategies. LWT Food Sci. Technol. 2010, 43, 573–583. [Google Scholar] [CrossRef]
- Nealson, K.H.; Rowe, A.R. Electromicrobiology: Realities, grand challenges, goals and predictions. Microb. Biotechnol. 2016, 9, 595. [Google Scholar] [CrossRef] [PubMed]
- Ali, J.; Sohail, A.; Wang, L.; Haider, M.R.; Mulk, S.; Pan, G. Electro-microbiology as a promising approach towards renewable energy and environmental sustainability. Energies 2018, 11, 1822. [Google Scholar] [CrossRef]
- Kumar, P.; Chandrasekhar, K.; Kumari, A.; Sathiyamoorthi, E.; Kim, B.S. Electro-Fermentation in Aid of Bioenergy and Biopolymers. Energies 2018, 11, 343. [Google Scholar] [CrossRef]
- Ishii, T.; Kawaichi, S.; Nakagawa, H.; Hashimoto, K.; Nakamura, R. From chemolithoautotrophs to electrolithoautotrophs: CO2 fixation by Fe(II)-oxidizing bacteria coupled with direct uptake of electrons from solid electron sources. Front. Microbiol. 2015, 6, 994. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Singh, L.; Zularisam, A.W. Exoelectrogens: Recent advances in molecular drivers involved in extracellular electron transfer and strategies used to improve it for microbial fuel cell applications. Renew. Sustain. Energy Rev. 2016, 56, 1322–1336. [Google Scholar] [CrossRef]
- Pillot, G.; Davidson, S.; Shintu, L.; Amin Ali, O.X.; Godfroy, A.; Combet-Blanc, Y.; Liebgott, P.-P. Electrotrophy as potential primary metabolism for colonization of conductive surfaces in deep-sea hydrothermal chimneys. 2 Key-words. bioRxiv 2020. [Google Scholar] [CrossRef]
- Yee, M.O.; Deutzmann, J.; Spormann, A.; Rotaru, A.E. Cultivating electroactive microbes-from field to bench. Nanotechnology 2020, 31, 174003. [Google Scholar] [CrossRef]
- Ajunwa, O.M.; Audu, J.O.; Kumar, P.; Marsili, E.; Onilude, A.A. Electrotrophs and Electricigens; Key Players in Microbial Electrophysiology. In Bioelectrochemical Systems; Springer: Singapore, 2020; pp. 299–326. [Google Scholar] [CrossRef]
- Cao, Y.; Mu, H.; Liu, W.; Zhang, R.; Guo, J.; Xian, M.; Liu, H. Electricigens in the anode of microbial fuel cells: Pure cultures versus mixed communities. Microb. Cell Fact. 2019, 18, 1–14. [Google Scholar] [CrossRef]
- Hernandez, C.A.; Osma, J.F. Microbial Electrochemical Systems: Deriving Future Trends From Historical Perspectives and Characterization Strategies. Front. Environ. Sci. 2020, 8, 44. [Google Scholar] [CrossRef]
- Saavedra, A.; Aguirre, P.; Gentina, J.C. Biooxidation of Iron by Acidithiobacillus ferrooxidans in the Presence of D-Galactose: Understanding Its Influence on the Production of EPS and Cell Tolerance to High Concentrations of Iron. Front. Microbiol. 2020, 11, 759. [Google Scholar] [CrossRef]
- Kracke, F.; Vassilev, I.; Krömer, J.O. Microbial electron transport and energy conservation-The foundation for optimizing bioelectrochemical systems. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Yu, H.; Zhang, J.; Li, F.; Song, H. Microbial electro-fermentation for the synthesis of chemicals and biofuels driven by bi-directional extracellular electron transfer. Synth. Syst. Biotechnol. 2020, 5, 304–313. [Google Scholar] [CrossRef]
- Zhao, J.; Li, F.; Cao, Y.; Zhang, X.; Chen, T.; Song, H.; Wang, Z. Microbial extracellular electron transfer and strategies for engineering electroactive microorganisms. Biotechnol. Adv. 2020, 107682. [Google Scholar] [CrossRef]
- Vassilev, I.; Averesch, N.J.H.; Ledezma, P.; Kokko, M. Anodic electro-fermentation: Empowering anaerobic production processes via anodic respiration. Biotechnol. Adv. 2021, 48, 107728. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, G.; Kouzuma, A.; Hirose, A.; Kasai, T.; Yoshida, G.; Watanabe, K. Metabolic Characteristics of a Glucose-Utilizing Shewanella oneidensis Strain Grown under Electrode-Respiring Conditions. PLoS ONE 2015, 10, e0138813. [Google Scholar] [CrossRef]
- Flynn, J.M.; Ross, D.E.; Hunt, K.A.; Bond, D.R.; Gralnick, J.A. Enabling unbalanced fermentations by using engineered electrode-interfaced bacteria. MBio 2010, 1. [Google Scholar] [CrossRef] [PubMed]
- Bursac, T.; Gralnick, J.A.; Gescher, J. Acetoin production via unbalanced fermentation in Shewanella oneidensis. Biotechnol. Bioeng. 2017, 114, 1283–1289. [Google Scholar] [CrossRef]
- Zheng, T.; Xu, B.; Ji, Y.; Zhang, W.; Xin, F.; Dong, W.; Wei, P.; Ma, J.; Jiang, M. Microbial fuel cell-assisted utilization of glycerol for succinate production by mutant of Actinobacillus succinogenes. Biotechnol. Biofuels 2021, 14, 1–10. [Google Scholar] [CrossRef]
- Kim, M.Y.; Kim, C.; Ainala, S.K.; Bae, H.; Jeon, B.H.; Park, S.; Kim, J.R. Metabolic shift of Klebsiella pneumoniae L17 by electrode-based electron transfer using glycerol in a microbial fuel cell. Bioelectrochemistry 2019, 125, 1–7. [Google Scholar] [CrossRef]
- Speers, A.M.; Young, J.M.; Reguera, G. Fermentation of Glycerol into Ethanol in a Microbial Electrolysis Cell Driven by a Customized Consortium. Environ. Sci. Technol. 2014, 48, 6350–6358. [Google Scholar] [CrossRef]
- Awate, B.; Steidl, R.J.; Hamlischer, T.; Reguera, G. Stimulation of electro-fermentation in single-chamber microbial electrolysis cells driven by genetically engineered anode biofilms. J. Power Source 2017, 356, 510–518. [Google Scholar] [CrossRef]
- Nishio, K.; Kimoto, Y.; Song, J.; Konno, T.; Ishihara, K.; Kato, S.; Hashimoto, K.; Nakanishi, S. Extracellular Electron Transfer Enhances Polyhydroxybutyrate Productivity in Ralstonia eutropha. Environ. Sci. Technol. Lett. 2013, 1, 40–43. [Google Scholar] [CrossRef]
- TerAvest, M.A.; Zajdel, T.J.; Ajo-Franklin, C.M. The Mtr Pathway of Shewanella oneidensis MR-1 Couples Substrate Utilization to Current Production in Escherichia coli. ChemElectroChem 2014, 1, 1874–1879. [Google Scholar] [CrossRef]
- Sturm-Richter, K.; Golitsch, F.; Sturm, G.; Kipf, E.; Dittrich, A.; Beblawy, S.; Kerzenmacher, S.; Gescher, J. Unbalanced fermentation of glycerol in Escherichia coli via heterologous production of an electron transport chain and electrode interaction in microbial electrochemical cells. Bioresour. Technol. 2015, 186, 89–96. [Google Scholar] [CrossRef]
- Lai, B.; Yu, S.; Bernhardt, P.V.; Rabaey, K.; Virdis, B.; Krömer, J.O. Anoxic metabolism and biochemical production in Pseudomonas putida F1 driven by a bioelectrochemical system. Biotechnol. Biofuels 2016, 9, 1–13. [Google Scholar] [CrossRef]
- Yu, S.; Lai, B.; Plan, M.R.; Hodson, M.P.; Lestari, E.A.; Song, H.; Krömer, J.O. Improved performance of Pseudomonas putida in a bioelectrochemical system through overexpression of periplasmic glucose dehydrogenase. Biotechnol. Bioeng. 2018, 115, 145–155. [Google Scholar] [CrossRef]
- Vassilev, I.; Gießelmann, G.; Schwechheimer, S.K.; Wittmann, C.; Virdis, B.; Krömer, J.O. Anodic electro-fermentation: Anaerobic production of L-Lysine by recombinant Corynebacterium glutamicum. Biotechnol. Bioeng. 2018, 115, 1499–1508. [Google Scholar] [CrossRef]
- Soussan, L.; Riess, J.; Erable, B.; Delia, M.L.; Bergel, A. Electrochemical reduction of CO2 catalysed by Geobacter sulfurreducens grown on polarized stainless steel cathodes. Electrochem. Commun. 2013, 28, 27–30. [Google Scholar] [CrossRef]
- Nevin, K.P.; Woodard, T.L.; Franks, A.E.; Summers, Z.M.; Lovley, D.R. Microbial electrosynthesis: Feeding microbes electricity to convert carbon dioxide and water to multicarbon extracellular organic compounds. MBio 2010, 1. [Google Scholar] [CrossRef]
- Tefft, N.M.; TerAvest, M.A. Reversing an Extracellular Electron Transfer Pathway for Electrode-Driven Acetoin Reduction. ACS Synth. Biol. 2019, 8, 1590–1600. [Google Scholar] [CrossRef] [PubMed]
- Utesch, T.; Sabra, W.; Prescher, C.; Baur, J.; Arbter, P.; Zeng, A.-P. Enhanced electron transfer of different mediators for strictly opposite shifting of metabolism in Clostridium pasteurianum grown on glycerol in a new electrochemical bioreactor. Biotechnol. Bioeng. 2019, 116, 1627–1643. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, F.; Cao, Y.; Tian, Y.; Li, J.; Zong, Y.; Song, H. Electricity-driven 7α-hydroxylation of a steroid catalyzed by a cytochrome P450 monooxygenase in engineered yeast. Catal. Sci. Technol. 2019, 9, 4877–4887. [Google Scholar] [CrossRef]
- Torella, J.P.; Gagliardi, C.J.; Chen, J.S.; Bediako, D.K.; Colón, B.; Way, J.C.; Silver, P.A.; Nocera, D.G. Efficient solar-to-fuels production from a hybrid microbial–water-splitting catalyst system. Proc. Natl. Acad. Sci. USA 2015, 112, 2337–2342. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sakimoto, K.K.; Colón, B.C.; Silver, P.A.; Nocera, D.G. Ambient nitrogen reduction cycle using a hybrid inorganic–biological system. Proc. Natl. Acad. Sci. USA 2017, 114, 6450–6455. [Google Scholar] [CrossRef]
- Kouzuma, A.; Kasai, T.; Hirose, A.; Watanabe, K. Catabolic and regulatory systems in Shewanella oneidensis MR-1 involved in electricity generation in microbial fuel cells. Front. Microbiol. 2015, 6, 609. [Google Scholar] [CrossRef]
- Li, F.; Li, Y.X.; Cao, Y.X.; Wang, L.; Liu, C.G.; Shi, L.; Song, H. Modular engineering to increase intracellular NAD(H/+) promotes rate of extracellular electron transfer of Shewanella oneidensis. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Goldbeck, C.P.; Jensen, H.M.; TerAvest, M.A.; Beedle, N.; Appling, Y.; Hepler, M.; Cambray, G.; Mutalik, V.; Angenent, L.T.; Ajo-Franklin, C.M. Tuning Promoter Strengths for Improved Synthesis and Function of Electron Conduits in Escherichia coli. ACS Synth. Biol. 2013, 2, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Andrade, A.; Hernández-Eligio, A.; Tirado, A.L.; Vega-Alvarado, L.; Olvera, M.; Morett, E.; Juárez, K. Specialization of the Reiterated Copies of the Heterodimeric Integration Host Factor Genes in Geobacter sulfurreducens. Front. Microbiol. 2021, 12, 626443. [Google Scholar] [CrossRef]
- Hernández-Eligio, A.; Pat-Espadas, A.; Vega-Alvarado, L.; Huerta-Amparán, M.; Cervantes, F.; Juárez, K. Global transcriptional analysis of Geobacter sulfurreducens under palladium reducing conditions reveals new key cytochromes involved. bioRxiv 2018, 319194. [Google Scholar] [CrossRef] [PubMed]
- Mevers, E.; Su, L.; Pishchany, G.; Baruch, M.; Cornejo, J.; Hobert, E.; Dimise, E.; Ajo-Franklin, C.M.; Clardy, J. An elusive electron shuttle from a facultative anaerobe. Elife 2019, 8. [Google Scholar] [CrossRef]
- Thirumurthy, M.A.; Jones, A.K. Geobacter cytochrome OmcZs binds riboflavin: Implications for extracellular electron transfer. Nanotechnology 2020, 31, 124001. [Google Scholar] [CrossRef]
- Hassan, R.Y.A.; Febbraio, F.; Andreescu, S. Microbial electrochemical systems: Principles, construction and biosensing applications. Sensors 2021, 21, 1279. [Google Scholar] [CrossRef]
- Logan, B.E.; Rossi, R.; Ragab, A.; Saikaly, P.E. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef]
- Hirose, A.; Kasai, T.; Koga, R.; Suzuki, Y.; Kouzuma, A.; Watanabe, K. Understanding and engineering electrochemically active bacteria for sustainable biotechnology. Bioresour. Bioprocess. 2019, 6, 1–15. [Google Scholar] [CrossRef]
- Leang, C.; Malvankar, N.S.; Franks, A.E.; Nevin, K.P.; Lovley, D.R. Engineering Geobacter sulfurreducens to produce a highly cohesive conductive matrix with enhanced capacity for current production. Energy Environ. Sci. 2013, 6, 1901–1908. [Google Scholar] [CrossRef]
- Kouzuma, A.; Oba, H.; Tajima, N.; Hashimoto, K.; Watanabe, K. Electrochemical selection and characterization of a high current-generating Shewanella oneidensis mutant with altered cell-surface morphology and biofilm-related gene expression. BMC Microbiol. 2014, 14, 190. [Google Scholar] [CrossRef] [PubMed]
- Jourdin, L.; Sousa, J.; van Stralen, N.; Strik, D.P.B.T.B. Techno-economic assessment of microbial electrosynthesis from CO2 and/or organics: An interdisciplinary roadmap towards future research and application. Appl. Energy 2020, 279, 115775. [Google Scholar] [CrossRef]
- Mahadevan, A.; Gunawardena, D.A.; Fernando, S. Biochemical and Electrochemical Perspectives of the Anode of a Microbial Fuel Cell. In Technology and Application of Microbial Fuel Cells; Wang, C.-T., Ed.; InTech: Hsinchu City, Taiwan, 2014. [Google Scholar] [CrossRef]
- Aryal, N.; Kvist, T.; Ammam, F.; Pant, D.; Ottosen, L.D.M. An overview of microbial biogas enrichment. Bioresour. Technol. 2018, 264, 359–369. [Google Scholar] [CrossRef]
- Selembo, P.A.; Merrill, M.D.; Logan, B.E. Hydrogen production with nickel powder cathode catalysts in microbial electrolysis cells. Int. J. Hydrogen Energy 2010, 35, 428–437. [Google Scholar] [CrossRef]
- Yaqoob, A.A.; Ibrahim, M.N.M.; Rodríguez-Couto, S. Development and modification of materials to build cost-effective anodes for microbial fuel cells (MFCs): An overview. Biochem. Eng. J. 2020, 164, 107779. [Google Scholar] [CrossRef]
- Nie, H.; Zhang, T.; Cui, M.; Lu, H.; Lovley, D.R.; Russell, T.P. Improved cathode for high efficient microbial-catalyzed reduction in microbial electrosynthesis cells. Phys. Chem. Chem. Phys. 2013, 15, 14290–14294. [Google Scholar] [CrossRef]
- Kellon, J.E.; Young, S.L.; Hutchison, J.E. Engineering the Nanoparticle–Electrode Interface. Chem. Mater. 2019, 31, 2685–2701. [Google Scholar] [CrossRef]
- Bian, B.; Alqahtani, M.F.; Katuri, K.P.; Liu, D.; Bajracharya, S.; Lai, Z.; Rabaey, K.; Saikaly, P.E. Porous nickel hollow fiber cathodes coated with CNTs for efficient microbial electrosynthesis of acetate from CO2 using Sporomusa ovata. J. Mater. Chem. A 2018, 6, 17201–17211. [Google Scholar] [CrossRef]
- Cai, P.J.; Xiao, X.; He, Y.R.; Li, W.W.; Zang, G.L.; Sheng, G.P.; Hon-Wah Lam, M.; Yu, L.; Yu, H.Q. Reactive oxygen species (ROS) generated by cyanobacteria act as an electron acceptor in the biocathode of a bio-electrochemical system. Biosens. Bioelectron. 2013, 39, 306–310. [Google Scholar] [CrossRef]
- Kondaveeti, S.; Min, B. Bioelectrochemical reduction of volatile fatty acids in anaerobic digestion effluent for the production of biofuels. Water Res. 2015, 87, 137–144. [Google Scholar] [CrossRef]
- Tahir, K.; Miran, W.; Jang, J.; Maile, N.; Shahzad, A.; Moztahida, M.; Ghani, A.A.; Kim, B.; Jeon, H.; Lee, D.S. MXene-coated biochar as potential biocathode for improved microbial electrosynthesis system. Sci. Total Environ. 2021, 773, 145677. [Google Scholar] [CrossRef]
- Yaqoob, A.A.; Ibrahim, M.N.M.; Rafatullah, M.; Chua, Y.S.; Ahmad, A.; Umar, K. Recent advances in anodes for microbial fuel cells: An overview. Materials 2020, 13, 2078. [Google Scholar] [CrossRef]
- Saito, T.; Mehanna, M.; Wang, X.; Cusick, R.D.; Feng, Y.; Hickner, M.A.; Logan, B.E. Effect of nitrogen addition on the performance of microbial fuel cell anodes. Bioresour. Technol. 2011, 102, 395–398. [Google Scholar] [CrossRef]
- Nakanishi, E.Y.; Palacios, J.H.; Godbout, S.; Fournel, S. Interaction between Biofilm Formation, Surface Material and Cleanability Considering Different Materials Used in Pig Facilities—An Overview. Sustainability 2021, 13, 5836. [Google Scholar] [CrossRef]
- Angelaalincy, M.J.; Navanietha Krishnaraj, R.; Shakambari, G.; Ashokkumar, B.; Kathiresan, S.; Varalakshmi, P. Biofilm Engineering Approaches for Improving the Performance of Microbial Fuel Cells and Bioelectrochemical Systems. Front. Energy Res. 2018, 6, 63. [Google Scholar] [CrossRef]
- Uria, N.; Ferrera, I.; Mas, J. Electrochemical performance and microbial community profiles in microbial fuel cells in relation to electron transfer mechanisms. BMC Microbiol. 2017, 17, 208. [Google Scholar] [CrossRef]
- Tuson, H.H.; Weibel, D.B. Bacteria-surface interactions. Soft Matter 2013, 9, 4368–4380. [Google Scholar] [CrossRef]
- Schechter, M.; Schechter, A.; Rozenfeld, S.; Efrat, E.; Cahan, R. Anode Biofilm. In Technology and Application of Microbial Fuel Cells; Wang, C.-T., Ed.; InTech: Hsinchu City, Taiwan, 2014. [Google Scholar] [CrossRef]
- Zhang, T.; Nie, H.; Bain, T.S.; Lu, H.; Cui, M.; Snoeyenbos-West, O.L.; Franks, A.E.; Nevin, K.P.; Russell, T.P.; Lovley, D.R. Improved cathode materials for microbial electrosynthesis. Energy Environ. Sci. 2012, 6, 217–224. [Google Scholar] [CrossRef]
- Nevin, K.P.; Hensley, S.A.; Franks, A.E.; Summers, Z.M.; Ou, J.; Woodard, T.L.; Snoeyenbos-West, O.L.; Lovley, D.R. Electrosynthesis of organic compounds from carbon dioxide is catalyzed by a diversity of acetogenic microorganisms. Appl. Environ. Microbiol. 2011, 77, 2882–2886. [Google Scholar] [CrossRef]
- Bhagchandanii, D.D.; Babu, R.P.; Sonawane, J.M.; Khanna, N.; Pandit, S.; Jadhav, D.A.; Khilari, S.; Prasad, R. A comprehensive understanding of electro-fermentation. Fermentation 2020, 6, 92. [Google Scholar] [CrossRef]
- Leang, C.; Ueki, T.; Nevin, K.P.; Lovley, D.R. A Genetic system for Clostridium ljungdahlii: A chassis for autotrophic production of biocommodities and a model homoacetogen. Appl. Environ. Microbiol. 2013, 79, 1102–1109. [Google Scholar] [CrossRef]
- Köpke, M.; Held, C.; Hujer, S.; Liesegang, H.; Wiezer, A.; Wollherr, A.; Ehrenreich, A.; Liebl, W.; Gottschalk, G.; Dürre, P. Clostridium ljungdahlii represents a microbial production platform based on syngas. Proc. Natl. Acad. Sci. USA 2010, 107, 13087–13092. [Google Scholar] [CrossRef]
- Kolesinska, B.; Fraczyk, J.; Binczarski, M.; Modelska, M.; Berlowska, J.; Dziugan, P.; Antolak, H.; Kaminski, Z.J.; Witonska, I.A.; Kregiel, D. Butanol synthesis routes for biofuel production: Trends and perspectives. Materials 2019, 12, 350. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, L.; Ke, C.; Pang, Z.; Liu, L. Pathway dissection, regulation, engineering and application: Lessons learned from biobutanol production by solventogenic clostridia. Biotechnol. Biofuels 2020, 13, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, A.; Leang, C.; Ueki, T.; Nevin, K.P.; Lovley, D.R. Lactose-inducible system for metabolic engineering of Clostridium ljungdahlii. Appl. Environ. Microbiol. 2014, 80, 2410–2416. [Google Scholar] [CrossRef]
- Kracke, F.; Krömer, J.O. Identifying target processes for microbial electrosynthesis by elementary mode analysis. BMC Bioinformatics 2014, 15, 410. [Google Scholar] [CrossRef]
- Khan, S.; Ullah, M.W.; Siddique, R.; Nabi, G.; Manan, S.; Yousaf, M.; Hou, H. Role of recombinant DNA technology to improve life. Int. J. Genom. 2016, 2016. [Google Scholar] [CrossRef]
- Adrio, J.L.; Demain, A.L. Microbial enzymes: Tools for biotechnological processes. Biomolecules 2014, 4, 117–139. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Wang, J.; Liu, J.; Wang, Y.; Bi, C.; Zhang, X. Engineering an electroactive Escherichia coli for the microbial electrosynthesis of succinate from glucose and CO2. Microb. Cell Fact. 2019, 18, 1–14. [Google Scholar] [CrossRef]
- Costa, N.L.; Carlson, H.K.; Coates, J.D.; Louro, R.O.; Paquete, C.M. Heterologous expression and purification of a multiheme cytochrome from a Gram-positive bacterium capable of performing extracellular respiration. Protein Expr. Purif. 2015, 111, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Phillips, J.R.; Huhnke, R.L.; Atiyeh, H.K. Syngas fermentation: A microbial conversion process of gaseous substrates to various products. Fermentation 2017, 3, 28. [Google Scholar] [CrossRef]
- Hamilton, T.L.; Bryant, D.A.; Macalady, J.L. The role of biology in planetary evolution: Cyanobacterial primary production in low-oxygen Proterozoic oceans. Environ. Microbiol. 2016, 18, 325–340. [Google Scholar] [CrossRef]
- Parmar, A.; Singh, N.K.; Pandey, A.; Gnansounou, E.; Madamwar, D. Cyanobacteria and microalgae: A positive prospect for biofuels. Bioresour. Technol. 2011, 102, 10163–10172. [Google Scholar] [CrossRef]
- Alqahtani, M.F.; Katuri, K.P.; Bajracharya, S.; Yu, Y.; Lai, Z.; Saikaly, P.E. Porous Hollow Fiber Nickel Electrodes for Effective Supply and Reduction of Carbon Dioxide to Methane through Microbial Electrosynthesis. Adv. Funct. Mater. 2018, 28, 1804860. [Google Scholar] [CrossRef]
- Bradley, R.W.; Bombelli, P.; Rowden, S.J.L.; Howe, C.J. Biological photovoltaics: Intra-and extra-cellular electron transport by cyanobacteria. Biochem. Soc. Trans. 2012, 40, 1302–1307. [Google Scholar] [CrossRef]
- Pandit, A.V.; Mahadevan, R. In silico characterization of microbial electrosynthesis for metabolic engineering of biochemicals. Microb. Cell Fact. 2011, 10, 1–14. [Google Scholar] [CrossRef]
- Cassia, R.; Nocioni, M.; Correa-Aragunde, N.; Lamattina, L. Climate change and the impact of greenhouse gasses: CO2 and NO, friends and foes of plant oxidative stress. Front. Plant Sci. 2018, 9, 273. [Google Scholar] [CrossRef]
- Das, S.; Das, S.; Das, I.; Ghangrekar, M.M. Application of bioelectrochemical systems for carbon dioxide sequestration and concomitant valuable recovery: A review. Mater. Sci. Energy Technol. 2019, 2, 687–696. [Google Scholar] [CrossRef]
- Butti, S.K.; Mohan, S.V. Autotrophic biorefinery: Dawn of the gaseous carbon feedstock. FEMS Microbiol. Lett. 2017, 364, 166. [Google Scholar] [CrossRef]
- Nitopi, S.; Bertheussen, E.; Scott, S.B.; Liu, X.; Engstfeld, A.K.; Horch, S.; Seger, B.; Stephens, I.E.L.; Chan, K.; Hahn, C.; et al. Progress and Perspectives of Electrochemical CO2 Reduction on Copper in Aqueous Electrolyte. Chem. Rev. 2019, 119, 7610–7672. [Google Scholar] [CrossRef] [PubMed]
- Tahir, K.; Miran, W.; Jang, J.; Woo, S.H.; Lee, D.S. Enhanced product selectivity in the microbial electrosynthesis of butyrate using a nickel ferrite-coated biocathode. Environ. Res. 2021, 196, 110907. [Google Scholar] [CrossRef]
- del Pilar Anzola Rojas, M.; Mateos, R.; Sotres, A.; Zaiat, M.; Gonzalez, E.R.; Escapa, A.; De Wever, H.; Pant, D. Microbial electrosynthesis (MES) from CO2 is resilient to fluctuations in renewable energy supply. Energy Convers. Manag. 2018, 177, 272–279. [Google Scholar] [CrossRef]
- Mateos, R.; Sotres, A.; Alonso, R.M.; Morán, A.; Escapa, A. Enhanced CO2 Conversion to Acetate through Microbial Electrosynthesis (MES) by Continuous Headspace Gas Recirculation. Energies 2019, 12, 3297. [Google Scholar] [CrossRef]
- Batlle-Vilanova, P.; Ganigué, R.; Ramió-Pujol, S.; Bañeras, L.; Jiménez, G.; Hidalgo, M.; Balaguer, M.D.; Colprim, J.; Puig, S. Microbial electrosynthesis of butyrate from carbon dioxide: Production and extraction. Bioelectrochemistry 2017, 117, 57–64. [Google Scholar] [CrossRef]
- Igarashi, K.; Kato, S. Reductive Transformation of Fe(III) (oxyhydr) Oxides by Mesophilic Homoacetogens in the Genus Sporomusa. Front. Microbiol. 2021, 12, 28. [Google Scholar] [CrossRef]
- Gavilanes, J.; Noori, M.T.; Min, B. Enhancing bio-alcohol production from volatile fatty acids by suppressing methanogenic activity in single chamber microbial electrosynthesis cells (SCMECs). Bioresour. Technol. Rep. 2019, 7, 100292. [Google Scholar] [CrossRef]
- Gavilanes, J.; Reddy, C.N.; Min, B. Microbial Electrosynthesis of Bioalcohols through Reduction of High Concentrations of Volatile Fatty Acids. Energy Fuels 2019, 33, 4264–4271. [Google Scholar] [CrossRef]
- Zhuang, W.-Q.; Yi, S.; Bill, M.; Brisson, V.L.; Feng, X.; Men, Y.; Conrad, M.E.; Tang, Y.J.; Alvarez-Cohen, L. Incomplete Wood–Ljungdahl pathway facilitates one-carbon metabolism in organohalide-respiring Dehalococcoides mccartyi. Proc. Natl. Acad. Sci. USA 2014, 111, 6419–6424. [Google Scholar] [CrossRef]
- Philips, J. Extracellular Electron Uptake by Acetogenic Bacteria: Does H2 Consumption Favor the H2 Evolution Reaction on a Cathode or Metallic Iron? Front. Microbiol. 2020, 10, 2997. [Google Scholar] [CrossRef] [PubMed]
- Vassilev, I.; Kracke, F.; Freguia, S.; Keller, J.; Krömer, J.O.; Ledezma, P.; Virdis, B. Microbial electrosynthesis system with dual biocathode arrangement for simultaneous acetogenesis, solventogenesis and carbon chain elongation. Chem. Commun. 2019, 55, 4351–4354. [Google Scholar] [CrossRef]
- Müller, V.; Wiechmann, A. Synthesis of Acetyl-CoA from Carbon Dioxide in Acetogenic Bacteria. In Biogenesis of Fatty Acids, Lipids and Membranes; Geiger, O., Ed.; Springer: Cham, Switzerland, 2017; pp. 1–18. [Google Scholar] [CrossRef]
- Liu, K.; Atiyeh, H.K.; Stevenson, B.S.; Tanner, R.S.; Wilkins, M.R.; Huhnke, R.L. Mixed culture syngas fermentation and conversion of carboxylic acids into alcohols. Bioresour. Technol. 2014, 152, 337–346. [Google Scholar] [CrossRef]
- Jin, S.; Jeon, Y.; Jeon, M.S.; Shin, J.; Song, Y.; Kang, S.; Bae, J.; Cho, S.; Lee, J.-K.; Kim, D.R.; et al. Acetogenic bacteria utilize light-driven electrons as an energy source for autotrophic growth. Proc. Natl. Acad. Sci. USA 2021, 118, 2020552118. [Google Scholar] [CrossRef]
- Tian, S.; Wang, H.; Dong, Z.; Yang, Y.; Yuan, H.; Huang, Q.; Song, T.S.; Xie, J. Mo2C-induced hydrogen production enhances microbial electrosynthesis of acetate from CO2 reduction. Biotechnol. Biofuels 2019, 12, 71. [Google Scholar] [CrossRef]
- Jourdin, L.; Lu, Y.; Flexer, V.; Keller, J.; Freguia, S. Biologically Induced Hydrogen Production Drives High Rate/High Efficiency Microbial Electrosynthesis of Acetate from Carbon Dioxide. ChemElectroChem 2016, 3, 581–591. [Google Scholar] [CrossRef]
- Perona-Vico, E.; Feliu-Paradeda, L.; Puig, S.; Bañeras, L. Bacteria coated cathodes as an in-situ hydrogen evolving platform for microbial electrosynthesis. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Chenevier, P.; Mugherli, L.; Darbe, S.; Darchy, L.; Dimanno, S.; Tran, P.D.; Valentino, F.; Iannello, M.; Volbeda, A.; Cavazza, C.; et al. Hydrogenase enzymes: Application in biofuel cells and inspiration for the design of noble-metal free catalysts for H2 oxidation. C. R. Chim. 2013, 16, 491–505. [Google Scholar] [CrossRef]
- Chen, H.; Dong, F.; Minteer, S.D. The progress and outlook of bioelectrocatalysis for the production of chemicals, fuels and materials. Nat. Catal. 2020, 3, 225–244. [Google Scholar] [CrossRef]
- ter Heijne, A.; Geppert, F.; Sleutels, T.H.J.A.; Batlle-Vilanova, P.; Liu, D.; Puig, S. Mixed culture biocathodes for production of hydrogen, methane, and carboxylates. In Advances in Biochemical Engineering/Biotechnology; Harnisch, F., Holtmann, D., Eds.; Springer: Cham, Switzerland, 2019; Volume 167, pp. 203–229. [Google Scholar] [CrossRef]
- Lalau, C.C.; Low, C.T.J. Electrophoretic Deposition for Lithium-Ion Battery Electrode Manufacture. Batter. Supercaps 2019, 2, 551–559. [Google Scholar] [CrossRef]
- Atiq Ur Rehman, M.; Chen, Q.; Braem, A.; Shaffer, M.S.P.; Boccaccini, A.R. Electrophoretic deposition of carbon nanotubes: Recent progress and remaining challenges. Int. Mater. Rev. 2020. [Google Scholar] [CrossRef]
- Huang, J.; Wang, J.; Xie, R.; Tian, Z.; Chai, G.; Zhang, Y.; Lai, F.; He, G.; Liu, C.; Liu, T.; et al. A universal pH range and a highly efficient Mo2C-based electrocatalyst for the hydrogen evolution reaction. J. Mater. Chem. A 2020, 8, 19879–19886. [Google Scholar] [CrossRef]
- Song, T.; Fei, K.; Zhang, H.; Yuan, H.; Yang, Y.; Ouyang, P.; Xie, J. High efficiency microbial electrosynthesis of acetate from carbon dioxide using a novel graphene–nickel foam as cathode. J. Chem. Technol. Biotechnol. 2018, 93, 457–466. [Google Scholar] [CrossRef]
- Bajracharya, S.; van den Burg, B.; Vanbroekhoven, K.; De Wever, H.; Buisman, C.J.N.; Pant, D.; Strik, D.P.B.T.B. In situ acetate separation in microbial electrosynthesis from CO2 using ion-exchange resin. Electrochim. Acta 2017, 237, 267–275. [Google Scholar] [CrossRef]
- Patil, S.A.; Arends, J.B.A.; Vanwonterghem, I.; van Meerbergen, J.; Guo, K.; Tyson, G.W.; Rabaey, K. Selective Enrichment Establishes a Stable Performing Community for Microbial Electrosynthesis of Acetate from CO2. Environ. Sci. Technol. 2015, 49, 8833–8843. [Google Scholar] [CrossRef]
- Mohanakrishna, G.; Vanbroekhoven, K.; Pant, D. Impact of dissolved carbon dioxide concentration on the process parameters during its conversion to acetate through microbial electrosynthesis. React. Chem. Eng. 2018, 3, 371–378. [Google Scholar] [CrossRef]
- Bajracharya, S.; Ter Heijne, A.; Dominguez Benetton, X.; Vanbroekhoven, K.; Buisman, C.J.N.; Strik, D.P.B.T.B.; Pant, D. Carbon dioxide reduction by mixed and pure cultures in microbial electrosynthesis using an assembly of graphite felt and stainless steel as a cathode. Bioresour. Technol. 2015, 195, 14–24. [Google Scholar] [CrossRef]
- Jourdin, L.; Grieger, T.; Monetti, J.; Flexer, V.; Freguia, S.; Lu, Y.; Chen, J.; Romano, M.; Wallace, G.G.; Keller, J. High Acetic Acid Production Rate Obtained by Microbial Electrosynthesis from Carbon Dioxide. Environ. Sci. Technol. 2015, 49, 13566–13574. [Google Scholar] [CrossRef]
- Dong, Z.; Wang, H.; Tian, S.; Yang, Y.; Yuan, H.; Huang, Q.; Song, T.S.; Xie, J. Fluidized granular activated carbon electrode for efficient microbial electrosynthesis of acetate from carbon dioxide. Bioresour. Technol. 2018, 269, 203–209. [Google Scholar] [CrossRef]
- Xie, S.; Liang, P.; Chen, Y.; Xia, X.; Huang, X. Simultaneous carbon and nitrogen removal using an oxic/anoxic-biocathode microbial fuel cells coupled system. Bioresour. Technol. 2011, 102, 348–354. [Google Scholar] [CrossRef]
- Marshall, C.W.; Ross, D.E.; Fichot, E.B.; Norman, R.S.; May, H.D. Long-term operation of microbial electrosynthesis systems improves acetate production by autotrophic microbiomes. Environ. Sci. Technol. 2013, 47, 6023–6029. [Google Scholar] [CrossRef]
- Ganesh, I. Conversion of carbon dioxide into methanol-A potential liquid fuel: Fundamental challenges and opportunities (a review). Renew. Sustain. Energy Rev. 2014, 31, 221–257. [Google Scholar] [CrossRef]
- Centi, G.; Quadrelli, E.A.; Perathoner, S. Catalysis for CO2 conversion: A key technology for rapid introduction of renewable energy in the value chain of chemical industries. Energy Environ. Sci. 2013, 6, 1711–1731. [Google Scholar] [CrossRef]
- Reymond, H.; Corral-Pérez, J.J.; Urakawa, A.; Rudolf Von Rohr, P. Towards a continuous formic acid synthesis: A two-step carbon dioxide hydrogenation in flow. React. Chem. Eng. 2018, 3, 912–919. [Google Scholar] [CrossRef]
- Rumayor, M.; Dominguez-Ramos, A.; Irabien, A. Formic Acid manufacture: Carbon dioxide utilization alternatives. Appl. Sci. 2018, 8, 914. [Google Scholar] [CrossRef]
- Álvarez, A.; Bansode, A.; Urakawa, A.; Bavykina, A.V.; Wezendonk, T.A.; Makkee, M.; Gascon, J.; Kapteijn, F. Challenges in the Greener Production of Formates/Formic Acid, Methanol, and DME by Heterogeneously Catalyzed CO2 Hydrogenation Processes. Chem. Rev. 2017, 117, 9804–9838. [Google Scholar] [CrossRef]
- Bahmanpour, A.M.; Signorile, M.; Kröcher, O. Recent progress in syngas production via catalytic CO2 hydrogenation reaction. Appl. Catal. B Environ. 2021, 295, 120319. [Google Scholar] [CrossRef]
- Sivalingam, V.; Ahmadi, V.; Babafemi, O.; Dinamarca, C. Integrating syngas fermentation into a single-cell microbial electrosynthesis (MES) reactor. Catalysts 2021, 11, 40. [Google Scholar] [CrossRef]
- Bustan, M.D.; Haryati, S.; Hadiah, F.; Selpiana, S.; Huda, A. Syngas Production Improvement of Sugarcane Bagasse Conversion Using an Electromagnetic Modified Vacuum Pyrolysis Reactor. Processes 2020, 8, 252. [Google Scholar] [CrossRef]
- El-Nagar, R.A.; Ghanem, A.A. Syngas Production, Properties, and Its Importance. Sustain. Altern. Syngas Fuel 2019. [Google Scholar] [CrossRef]
- Nelabhotla, A.B.T.; Dinamarca, C. Bioelectrochemical CO2 Reduction to Methane: MES Integration in Biogas Production Processes. Appl. Sci. 2019, 9, 1056. [Google Scholar] [CrossRef]
- Enzmann, F.; Mayer, F.; Rother, M.; Holtmann, D. Methanogens: Biochemical background and biotechnological applications. AMB Express 2018, 8, 1. [Google Scholar] [CrossRef]
- Munasinghe, P.C.; Khanal, S.K. Syngas fermentation to biofuel: Evaluation of carbon monoxide mass transfer coefficient (kLa) in different reactor configurations. Biotechnol. Prog. 2010, 26, 1616–1621. [Google Scholar] [CrossRef]
- Kumar, G.; Saratale, R.G.; Kadier, A.; Sivagurunathan, P.; Zhen, G.; Kim, S.H.; Saratale, G.D. A review on bio-electrochemical systems (BESs) for the syngas and value-added biochemicals production. Chemosphere 2017, 177, 84–92. [Google Scholar] [CrossRef]
- Annie Modestra, J.; Katakojwala, R.; Venkata Mohan, S. CO2 fermentation to short-chain fatty acids using selectively enriched chemolithoautotrophic acetogenic bacteria. Chem. Eng. J. 2020, 394, 124759. [Google Scholar] [CrossRef]
- Aryal, N.; Ghimire, N.; Bajracharya, S. Chapter 3—Coupling of microbial electrosynthesis with anaerobic digestion for waste valorization. In Advances in Bioenergy; Li, Y., Khanal, S.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 101–127. [Google Scholar] [CrossRef]
- Vu, H.T.; Min, B. Enhanced methane fermentation of municipal sewage sludge by microbial electrochemical systems integrated with anaerobic digestion. Int. J. Hydrogen Energy 2019, 44, 30357–30366. [Google Scholar] [CrossRef]
- Yang, H.Y.; Hou, N.N.; Wang, Y.X.; Liu, J.; He, C.S.; Wang, Y.R.; Li, W.H.; Mu, Y. Mixed-culture biocathodes for acetate production from CO2 reduction in the microbial electrosynthesis: Impact of temperature. Sci. Total Environ. 2021, 790, 148128. [Google Scholar] [CrossRef]
- Names, A.; Whitman, J.D.; Hiatt, J.; Mowery, C.T.; Shy, B.R.; Yu, R.; Yamamoto, T.N.; Rathore, U.; Goldgof, G.M.; Whitty, C.; et al. Title: Test performance evaluation of SARS-CoV-2 serological assays. medRxiv 2020, 29, 30. [Google Scholar] [CrossRef]
- Satinover, S.J.; Rodriguez, M.; Campa, M.F.; Hazen, T.C.; Borole, A.P. Performance and community structure dynamics of microbial electrolysis cells operated on multiple complex feedstocks. Biotechnol. Biofuels 2020, 13, 1–21. [Google Scholar] [CrossRef]
- Saboe, P.O.; Manker, L.P.; Monroe, H.R.; Michener, W.E.; Haugen, S.; Tan, E.C.D.; Prestangen, R.L.; Beckham, G.T.; Karp, E.M. Energy and techno-economic analysis of bio-based carboxylic acid recovery by adsorption. Green Chem. 2021, 23, 4386–4402. [Google Scholar] [CrossRef]
- Clarke, C.J.; Tu, W.C.; Levers, O.; Bröhl, A.; Hallett, J.P. Green and Sustainable Solvents in Chemical Processes. Chem. Rev. 2018, 118, 747–800. [Google Scholar] [CrossRef] [PubMed]
- Keshav, A.; Wasewar, K.L.; Chand, S. Extraction of propionic acid using different extractants (tri-n-butylphosphate, tri-n-octylamine, and Aliquat 336). Ind. Eng. Chem. Res. 2008, 47, 6192–6196. [Google Scholar] [CrossRef]
- Li, Q.-Z.; Jiang, X.-L.; Feng, X.-J.; Wang, J.-M.; Sun, C.; Zhang, H.-B.; Xian, M.; Liu, H.-Z. Recovery Processes of Organic Acids from Fermentation Broths in the Biomass-Based Industry. J. Microbiol. Biotechnol 2016, 26, 1–8. [Google Scholar] [CrossRef]
- Woo, H.C.; Kim, Y.H. Eco-efficient recovery of bio-based volatile C2-6 fatty acids. Biotechnol. Biofuels 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Zhu, X.; Leininger, A.; Jassby, D.; Tsesmetzis, N.; Ren, Z.J. Will Membranes Break Barriers on Volatile Fatty Acid Recovery from Anaerobic Digestion? ACS EST Eng. 2021, 1, 141–153. [Google Scholar] [CrossRef]
- Zhu, Y. Evaluation of Nanofiltration for the Extraction of Volatile Fatty Acids from Fermentation Broth. 2020. Available online: https://tel.archives-ouvertes.fr/tel-03008506 (accessed on 8 August 2021).
- Andersen, S.J.; Berton, J.K.E.T.; Naert, P.; Gildemyn, S.; Rabaey, K.; Stevens, C.V. Extraction and esterification of low-titer short-chain volatile fatty acids from anaerobic fermentation with ionic liquids. ChemSusChem 2016, 9, 2059–2063. [Google Scholar] [CrossRef]
- Montesantos, N.; Maschietti, M. Supercritical carbon dioxide extraction of lignocellulosic bio-oils: The potential of fuel upgrading and chemical recovery. Energies 2020, 13, 1600. [Google Scholar] [CrossRef]
- Murali, N.; Srinivas, K.; Ahring, B.K. Increasing the production of volatile fatty acids from corn stover using bioaugmentation of a mixed rumen culture with homoacetogenic bacteria. Microorganisms 2021, 9, 337. [Google Scholar] [CrossRef]
- Sprakel, L.M.J.; Schuur, B. Solvent developments for liquid-liquid extraction of carboxylic acids in perspective. Sep. Purif. Technol. 2019, 211, 935–957. [Google Scholar] [CrossRef]
- Naik, S.N.; Goud, V.V.; Rout, P.K.; Dalai, A.K. Production of first and second-generation biofuels: A comprehensive review. Renew. Sustain. Energy Rev. 2010, 14, 578–597. [Google Scholar] [CrossRef]
- Ho, D.P.; Ngo, H.H.; Guo, W. A mini-review on renewable sources for biofuel. Bioresour. Technol. 2014, 169, 742–749. [Google Scholar] [CrossRef]
- Savla, N.; Shinde, A.; Sonawane, K.; Mekuto, L.; Chowdhary, P.; Pandit, S. Microbial hydrogen production: Fundamentals to application. In Microorganisms for Sustainable Environment and Health; Elsevier: Gurgaon, India, 2020; pp. 343–365. [Google Scholar] [CrossRef]
- Committee of America’s Energy Future. America’s Energy Future: Technology and Transformation; National Academies Press: Washington, DC, USA, 2010; ISBN 0309116023. [Google Scholar]
- IRENA. Renewable Power Generation Costs in 2019; International Renewable Energy Agency: Abu Dhabi, United Arab Emirates, 2020; ISBN 978-92-9260-244-4. [Google Scholar]
- Owusu, P.A.; Asumadu-Sarkodie, S. A review of renewable energy sources, sustainability issues and climate change mitigation. Cogent Eng. 2016, 3. [Google Scholar] [CrossRef]
- Zsiborács, H.; Baranyai, N.H.; Vincze, A.; Zentkó, L.; Birkner, Z.; Máté, K.; Pintér, G. Intermittent renewable energy sources: The role of energy storage in the european power system of 2040. Electronics 2019, 8, 729. [Google Scholar] [CrossRef]
- Lund, H.; Kempton, W. Integration of renewable energy into the transport and electricity sectors through V2G. Energy Policy 2008, 36, 3578–3587. [Google Scholar] [CrossRef]
- Popp, J.; Lakner, Z.; Harangi-Rákos, M.; Fári, M. The effect of bioenergy expansion: Food, energy, and environment. Renew. Sustain. Energy Rev. 2014, 32, 559–578. [Google Scholar] [CrossRef]
- Bajracharya, S.; Srikanth, S.; Mohanakrishna, G.; Zacharia, R.; Strik, D.P.; Pant, D. Biotransformation of carbon dioxide in bioelectrochemical systems: State of the art and future prospects. J. Power Source 2017, 356, 256–273. [Google Scholar] [CrossRef]
- Molina, M.G. Energy Storage and Power Electronics Technologies: A Strong Combination to Empower the Transformation to the Smart Grid. Proc. IEEE 2017, 105, 2191–2219. [Google Scholar] [CrossRef]
- Chaudhary, D.K.; Kim, J. New insights into bioremediation strategies for oil-contaminated soil in cold environments. Int. Biodeterior. Biodegrad. 2019, 142, 58–72. [Google Scholar] [CrossRef]
- Huang, C.H.; Tan, C.S. A review: CO2 utilization. Aerosol Air Qual. Res. 2014, 14, 480–499. [Google Scholar] [CrossRef]
- Singh Saharan, B.; Sharma, D.; Sahu, R.; Sahin, O.; Warren, A.; Biomonitoring, P.; Millipore, M.; Caddesi, K.; Ciftligi, K.; Kar, Y. Towards Algal Biofuel Production: A Concept of Green Bio Energy Development. Innov. Rom. Food Biotechnol. 2013, 12. Available online: http://www.bioaliment.ugal.ro/ejournal.htm (accessed on 8 November 2021).
- Irfan, M.; Bai, Y.; Zhou, L.; Kazmi, M.; Yuan, S.; Maurice Mbadinga, S.; Yang, S.Z.; Liu, J.F.; Sand, W.; Gu, J.D.; et al. Direct microbial transformation of carbon dioxide to value-added chemicals: A comprehensive analysis and application potentials. Bioresour. Technol. 2019, 288, 121401. [Google Scholar] [CrossRef]
- Johnson, M.C.; Heim, C.J.; Billingham, J.F.; Malczewski, M.L. Method for Analyzing Impurities in Carbon Dioxide 2006. Available online: https://patents.google.com/patent/US7064834B2 (accessed on 8 October 2021).
- Lovley, D.R.; Nevin, K.P. Microbial Production of Multi-Carbon Chemicals and Fuels from Water and Carbon Dioxide Using Electric Current 2008. Available online: https://patents.google.com/patent/US9856449B2 (accessed on 8 October 2021).
- Lajua Jo Eun-cheol. Cathode for Production of Biofuel, and Microbial Electrosynthesis System for Production of Biofuel Comprising the Same 2018. Available online: https://patents.google.com/patent/KR101892982B1 (accessed on 8 October 2021).
- Deutzmann, J.S.; Spormann, A.M. Enhanced microbial electrosynthesis by using co-cultures. ISME J. 2016. [Google Scholar] [CrossRef]
- May, H.D.; Labelle, E.V. Bioelectrosynthesis of Organic Compounds 2019. Available online: https://patents.google.com/patent/US20190301029A1 (accessed on 8 October 2021).
- Bergel, A.; Bernet, N.; Blanchet, E.; Bouchez, T.; Erable, B.; Etcheverry, L.; Huyard, A.; Quemener, E.L.; Mauricrace, P.; Moreau, S.; et al. Bioelectrochemical Reactor with Double Bioanode, Method for Anodic Regeneration and Use of the Reactor for Microbial Electrosynthesis 2020. Available online: https://patents.google.com/patent/WO2020053529A1 (accessed on 8 October 2021).
- Li, W.; Menggen, L.; Yang, Y.; Ning, H.; Jiafang, Z. Microbial Cathode Catalytic Reduction CO2 Method for Electrosynthesis of Organic Matter 2020. Available online: https://patents.google.com/patent/CN111961691A (accessed on 8 October 2021).
- Lovley, D.R.; Ueki, T.; LOVLEY, K.N. Microbial Strain for Electrosynthesis and Electrofermentation 2020. Available online: https://patents.google.com/patent/US10711318B2 (accessed on 8 October 2021).
- Kun, G. Electrolytic Hydrogen Bubble Column Microbial Electrosynthesis Reactor and Use Method Thereof 2019. Available online: https://patents.google.com/patent/CN110528017B (accessed on 8 October 2021).
- Prévoteau, A.; Carvajal-Arroyo, J.M.; Ganigué, R.; Rabaey, K. Microbial electrosynthesis from CO2: Forever a promise? Curr. Opin. Biotechnol. 2020, 62, 48–57. [Google Scholar] [CrossRef] [PubMed]







| Carbon Cloth Cathode Treatment | Average Current Consumption Density (mA m−2) | Acetate (mM m−2 day−1) | Coulombic Efficiency |
|---|---|---|---|
| Carbon cloth | −71 ± 11 | 30 ± 7 | 76 ± 14 |
| 3-Aminopropyltriethoxysilane | −206 ± 11 | 95 ± 20 | 82 ± 11 |
| Ni | −302 ± 48 | 136 ± 33 | 80 ± 15 |
| Melamine | −69 ± 9 | 31 ± 08 | 80 ± 15 |
| Carbon Nanotube-cotton | −220 ± 1 | 102 ± 25 | 83 ± 10 |
| Cyanuric chloride | −451 ± 79 | 205 ± 50 | 81 ± 16 |
| Ammonia | −60 ± 21 | 28 ± 14 | 82 ± 8 |
| Pd | −320 ± 64 | 141 ± 35 | 79 ± 16 |
| Chitosan | −475 ± 18 | 229 ± 56 | 86 ± 12 |
| Polyaniline | −189 ± 18 | 90 ± 22 | 85 ± 7 |
| Au | −388 ± 43 | 181 ± 44 | 83 ± 14 |
| Carbon Nanotube-polyester | −210 ± 13 | 96 ± 24 | 82 ± 8 |
| Species | Electron Consumption? (EC) | EC Rate vs. S. ovata | Electron Recovery in Products | Products Formed |
|---|---|---|---|---|
| Moorella thermoacetica | Yes | - | 85% ± 7% (n = 3) | Acetate |
| Clostridium lijungdahli | Yes | - | 82% ± 10% (n = 3) | Acetate + Minor formate and 2-oxobutyrate over time |
| Sporomusa ovata | Yes | 100% | 86% ± 21% | Acetate + Trace of 2-oxobutyrate |
| Acetobacterium woodii | No | - | - | - |
| Sporomusa silvacetica | Yes | 10% | 48% ± 6% | Acetate + Trace of 2-oxobutyrate + non-identified products |
| Clostridium aceticum | Yes | - | 53% ± 4% (n = 2) | 2-oxobutyrate and acetate as prime products and other non-identified |
| Sporomusa sphaeroides | Yes | 5% | 84% ± 26% (n = 3) | Acetate |
| Microbial Inoculum | Cathode Material | Ecathode (V vs. SHE) | Current Density (A m−2) | Acetate Production (g−2 day−1) | Max Acetate (g L−2) | Electron Recovery into Acetate % |
|---|---|---|---|---|---|---|
| S. ovata (continuous) | Graphite rods | −0.4 | −0.208 | 1.3 | 0.063 | 86 ± 21 |
| C. ljungdahlii (continuous) | Graphite rods | −0.4 | −0.029 | 0.14 | - | 72 |
| Brewery WW sludge (fed-batch) | Graphite granules | −0.590 | - | - | 1.71 | 67 |
| Enriched Brewery WW sludge (fed-batch) | Graphite granules | −0.590 | - | - | 10.5 | 69 |
| Enriched WWTP sludge (fed-batch) | Carbon felt | −0.9 | - | 34.5 | - | 89.5 |
| S. ovata (continuous) | Carbon cloth chitosan | −0.4 | −0.475 | 2.7 | 0.118 | 86 ± 12 |
| S. ovata (continuous) | CNT cotton CNT polyester | −0.4 | −0.215 | ~1.2 | 0.059 | 83 ± 10 |
| S. ovate (continuous) | Network coated graphite | −0.4 | ~−0.625 | 3.3 | - | 82 ± 14 |
| Enriched Brewery WW sludge (fed-batch) | Graphite rods | −0.6 | −0.92 ± 0.12 | 8.56 ± 3.22 | - | 40 |
| Mesophilic Brewery WW anaerobic sludge (fed-batch) | Graphite felt | −1.1 | ~−2.8 | 10.1 | 1.4 | 65 |
| Anaerobic digester | Graphite granules | −0.6 | - | - | - | 28.9 ± 6.1 |
| Mixed natural & engineered sludge (fed-batch) | NanoWeb-RVC | −0.85 | −37 | 192 | 1.65 | 70 ± 11 |
| Enriched Mixed natural & engineered sludge (fed-batch) | EPD-3D | −0.85 | −102 | 685 | 11 | 100 ± 4 |
| Inoculum | Cell Design | Cathode | Anode | Current (mA/cm2) | Main Product (Yield/Final Concentration) | Coulombic Efficiency (%) | Solar Conversion Efficiency (%) |
|---|---|---|---|---|---|---|---|
| Engineered Ralstonia eutropha | Single chamber | NiMoZn or stainless steel | CoPi | 0.5–1.1 | Isopropanol (216 mg/L) | 3.9 | 0.7 |
| Enriched methanogenic community | Dual chambers | Carbon cloth | TiO2 nanowire array | 0.07–0.09 | Methane (1.92 L/(m2 d)) | 95 | 0.1 |
| Sporomusa ovata | Dual chamber | Si and TiO2 nanowires arrays | TiO2 nanowires | 0.3 | Acetic acid (6 g/L) | 86 | 0.38 |
| Effluent from methanogenic MES | Dual chamber | Chitosan modified carbon cloth | TiO2/CdS on Fluorine-doped tin oxide (with copper zinc tin sulphide sensitiser) | 0.6 | Methane (15 L/(m2 d), 20.8 L/(m2 d) with copper zinc tin sulphide) | 93 | 0.62 (0.86 with copper zinc tin sulphide) |
| Patent No. | Description | Ref. |
|---|---|---|
| US9856449B2 | The innovation offers mechanisms and approaches for the generation of various organic compounds by utilising CO2 as an origin of carbon and electricity as an energy’s point of origin. A reaction cell is supplied with an anode and a cathode (containing microbial biofilm) distinguished by the utilisation of a selectively porous membrane and conjugated to an electrical power source. The microbial biofilm contains an electrogen that can accept electrons and, in a cathode half-reaction, it can convert CO2 to an organic compound and water, which is then decomposed into free molecular O2 and p+ in an anode half-reaction. The half-reactions are powered by electricity from an external source. Butanol, ethanol, formate, acetate and 2-oxobutyrate are the compounds that can be produced using this technology. | [232] |
| KR101892982B1 | As per the current innovation, a traditional carbon electrode surface has been altered with a positive amine compound to surge the amount of adsorption of an EAM biofilm on the surface while simultaneously, improving the efficiency of transfer of electrons by adding metal nanoparticles, thereby maximising the generation of several biofuels such as biomethanol and hexanol. | [233] |
| US10494596B2 | In specific, the system refers to MES, via which a microbial strain capable of collecting electrons from an electrode is used to generate CO2, formate or H2 in co-cultivation with a strain of microbial development such as methanogen, acetogen or other microbes capable of producing those products. | [234] |
| US20190301029A1 | A system of bioelectric processing of organic molecules like acetate is studied in this current disclosure. In addition, it also proposes strategies for generating a hydrocarbon-based product using CO2 as the source of carbon. | [235] |
| WO2020053529A1 | The innovation discloses the process for the regeneration of the reactor’s bioanode operation and the application of the reactor for the electrosynthesis of organic acids and organic waste alcohols. | [236] |
| CN111961691A | This innovation discloses the utilisation of a biocathode in MES for catalytic CO2 reduction and synthesis of organic compounds. Preparation of the biocathode with Ruminococcus, Clostridium and Lachnospiraceae culture and injection of CO2 into the cathode chamber, circulating aeration and setting the theoretical range for polarisation to be −0.8 V to 1.2 V (vs. Ag/AgCl). The invention reduces the CO2 content drastically with a simultaneous high synthesising rate of organic compounds. | [237] |
| US10711318B2 | In comparison to the wild type of strain, a GM Geobacter sulfurreducens strain demonstrates enhanced functionality as a cathode biofilm. This strain is effective in utilising CO2 as an origin of carbon and electricity as an origin of energy, employing a reverse tricarboxylic acid mechanism to produce a carbonaceous chemical. | [238] |
| CN110528017B | This paper demonstrates the bubbling tower of an electrolytic H2 MES reactor. An electrolytic bath configured below the reactor supplies the bubble tower with micro-nano H2 bubbles. H2 and CO2 are supplied by the microbes suspended in the bubble tower and then processed into organic compounds. This innovation is ideal for the method of H2 Induced microbial CO2 fixation, which is also relevant to the process of H2-driven microbial sewage denitrification. This has perks of high coulombic performance, fast reactor start time, high current density, high output intensity, high system stability, compared to the conventional MES system dependent on the electroactive surface biofilm. | [239] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quraishi, M.; Wani, K.; Pandit, S.; Gupta, P.K.; Rai, A.K.; Lahiri, D.; Jadhav, D.A.; Ray, R.R.; Jung, S.P.; Thakur, V.K.; et al. Valorisation of CO2 into Value-Added Products via Microbial Electrosynthesis (MES) and Electro-Fermentation Technology. Fermentation 2021, 7, 291. https://doi.org/10.3390/fermentation7040291
Quraishi M, Wani K, Pandit S, Gupta PK, Rai AK, Lahiri D, Jadhav DA, Ray RR, Jung SP, Thakur VK, et al. Valorisation of CO2 into Value-Added Products via Microbial Electrosynthesis (MES) and Electro-Fermentation Technology. Fermentation. 2021; 7(4):291. https://doi.org/10.3390/fermentation7040291
Chicago/Turabian StyleQuraishi, Marzuqa, Kayinath Wani, Soumya Pandit, Piyush Kumar Gupta, Ashutosh Kumar Rai, Dibyajit Lahiri, Dipak A. Jadhav, Rina Rani Ray, Sokhee P. Jung, Vijay Kumar Thakur, and et al. 2021. "Valorisation of CO2 into Value-Added Products via Microbial Electrosynthesis (MES) and Electro-Fermentation Technology" Fermentation 7, no. 4: 291. https://doi.org/10.3390/fermentation7040291
APA StyleQuraishi, M., Wani, K., Pandit, S., Gupta, P. K., Rai, A. K., Lahiri, D., Jadhav, D. A., Ray, R. R., Jung, S. P., Thakur, V. K., & Prasad, R. (2021). Valorisation of CO2 into Value-Added Products via Microbial Electrosynthesis (MES) and Electro-Fermentation Technology. Fermentation, 7(4), 291. https://doi.org/10.3390/fermentation7040291