
Chemical and Sensory Profiles of Merlot Wines Produced by Sequential Inoculation of Metschnikowia pulcherrima or Meyerzyma guilliermondii


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. Merlot Grape Must
2.3. Chemical and Microbiological Analyses
2.4. Sensory Analyses
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquin, D.; Calderón, F.; Santos, A.; Benito, S. The genus Metschnikowia in enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The effect of non-Saccharomyces yeasts on fermentation and wine quality. S. Afr. J. Enol. Vitic. 2003, 24, 55–62. [Google Scholar] [CrossRef]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The use of Candida pulcherrima in combination with Saccharomyces cerevisiae for the production of Chenin blanc wine. S. Afr. J. Enol. Vitic. 2003, 24, 63–69. [Google Scholar] [CrossRef] [Green Version]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef]
- Rojas, V.; Gil, J.V.; Piñaga, F.; Manzanares, P. Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 2003, 86, 181–188. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genovés, S.; Vallés, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Ciani, M.; Maccarelli, F. Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 1998, 14, 199–203. [Google Scholar] [CrossRef]
- Bataillon, M.; Rico, A.; Sablayrolles, J.-M.; Salmon, J.-M.; Barre, P. Early thiamin assimilation by yeasts under enological conditions: Impact on alcoholic fermentation kinetics. J. Ferment. Bioeng. 1996, 82, 145–150. [Google Scholar] [CrossRef]
- Medina, K.; Boido, E.; Dellacassa, E.; Carrau, F. Growth of non-Saccharomyces yeasts affects nutrient availability for Saccharomyces cerevisiae during wine fermentation. Int. J. Food Microbiol. 2012, 157, 245–250. [Google Scholar] [CrossRef]
- Rollero, S.; Bloem, A.; Ortiz-Julien, A.; Camarasa, C.; Divol, B. Altered fermentation performances, growth, and metabolic footprints reveal competition for nutrients between yeast species inoculated in synthetic grape juice-like medium. Front. Microbiol. 2018, 9, 196. [Google Scholar] [CrossRef]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Canonico, L.; Comitini, F.; Oro, L.; Ciani, M. Sequential fermentations with selected immobilized non-Saccharomyces yeast for reduction of ethanol content in wine. Front. Microbiol. 2016, 7, 278. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef]
- Rossouw, D.; Bauer, F. Exploring the phenotypic space of non-Saccharomyces wine yeast biodiversity. Food Microbiol. 2016, 55, 32–46. [Google Scholar] [CrossRef]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavor profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.J.; White, K.P.; Edwards, C.G. Growth and metabolism of non-Saccharomyces yeasts isolated from Washington state vineyards in media and high sugar grape musts. Food Microbiol. 2019, 77, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Aplin, J.A.; Edwards, C.G. Impacts of non-Saccharomyces species and aeration on sequential inoculation with Saccharomyces cerevisiae to produce lower alcohol Merlot wines. J. Sci. Food Agric. 2021, 101, 1715–1719. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Teresa Arroyo, T. Sequential non-Saccharomyces and Saccharomyces cerevisiae fermentations to reduce the alcohol content in wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Hranilovic, A.; Gambetta, J.M.; Jeffery, D.W.; Grbin, P.R.; Jiranek, V. Lower-alcohol wines produced by Metschnikowia pulcherrima and Saccharomyces cerevisiae co-fermentations: The effect of sequential inoculation timing. Int. J. Food Microbiol. 2020, 329, 108651. [Google Scholar] [CrossRef] [PubMed]
- Bourret, T.B.; Grove, G.G.; Vandemark, G.J.; Henick-Kling, T.; Glawe, D.A. Diversity and molecular determination of wild yeasts in a central Washington state vineyard. N. Am. Fungi 2013, 8, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Contreras, A.; Curtin, C.; Varela, C. Yeast population dynamics reveal a potential ‘collaboration’ between Metschnikowia pulcherrima and Saccharomyces uvarum for the production of reduced alcohol wines during Shiraz fermentation. Appl. Microbiol. Biotechnol. 2015, 99, 1885–1895. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef]
- Rodríguez, M.E.; Lopes, C.A.; Barbagelata, R.J.; Barda, N.B.; Caballero, A.C. Influence of Candida pulcherrima Patagonian strain on alcoholic fermentation behaviour and wine aroma. Int. J. Food Microbiol. 2010, 138, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Ough, C.S.; Amerine, M.A. Methods for Analysis of Musts and Wines; John Wiley & Sons: New York, NY, USA, 1988. [Google Scholar]
- Dukes, B.C.; Butzke, C.E. Rapid determination of primary amino acids in grape juice using an o-pthaldialdehyde/N-acetyl-L-cysteine spectrophotometric assay. Am. J. Enol. Vitic. 1998, 49, 125–134. [Google Scholar]
- Aplin, J.A. Use of Non-Saccharomyces Yeasts for Reducing the Ethanol Contents of Red Wine. Ph.D. Thesis, Washington State University, Pullman, WA, USA, 2019. [Google Scholar]
- Andorrà, I.; Berradre, M.; Rozès, N.; Mas, A.; Guillamón, J.M.; Esteve-Zarzoso, B. Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 2010, 231, 215–224. [Google Scholar] [CrossRef] [Green Version]
- Gobert, A.; Tourdot-Maréchal, R.; Morge, C.; Sparrow, C.; Liu, Y.; Quintanilla-Casas, B.; Vichi, S.; Alexandre, H. Non-Saccharomyces yeasts nitrogen source preference: Impacts on sequential fermentation and wine volatile compounds profile. Front. Microbiol. 2017, 8, 2175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Klerk, J.L. Succinic Acid Production by Wine Yeasts. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 2010. [Google Scholar]
- Jackson, R.S. Wine Science: Principles and Applications, 3rd ed.; Elsevier Science: Amsterdam, The Netherlands, 2008. [Google Scholar]
- Benito, S.; Morata, A.; Palomero, F.; González, M.C.; Suárez-Lepe, J.A. Formation of vinylphenolic pyranoanthocyanins by Saccharomyces cerevisiae and Pichia guillermondii in red wines produced following different fermentation strategies. Food Chem. 2011, 124, 15–23. [Google Scholar] [CrossRef]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbüler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Bende, M.; Nordin, S. Perceptual learning in olfaction: Professional wine tasters versus controls. Physiol. Behav. 1997, 62, 1065–1070. [Google Scholar] [CrossRef]
- Landon, J.L.; Weller, K.; Harbertson, J.F.; Ross, C.F. Chemical and sensory evaluation of astringency in Washington state red wines. Am. J. Enol. Vitic. 2008, 59, 153–158. [Google Scholar]
- McMahon, K.M.; Diako, C.; Aplin, J.; Mattinson, D.S.; Culver, C.; Ross, C.F. Trained and consumer panel evaluation of sparkling wines sweetened to brut or demi sec residual sugar levels with three different sugars. Food Res. Int. 2017, 99, 173–185. [Google Scholar] [CrossRef] [PubMed]
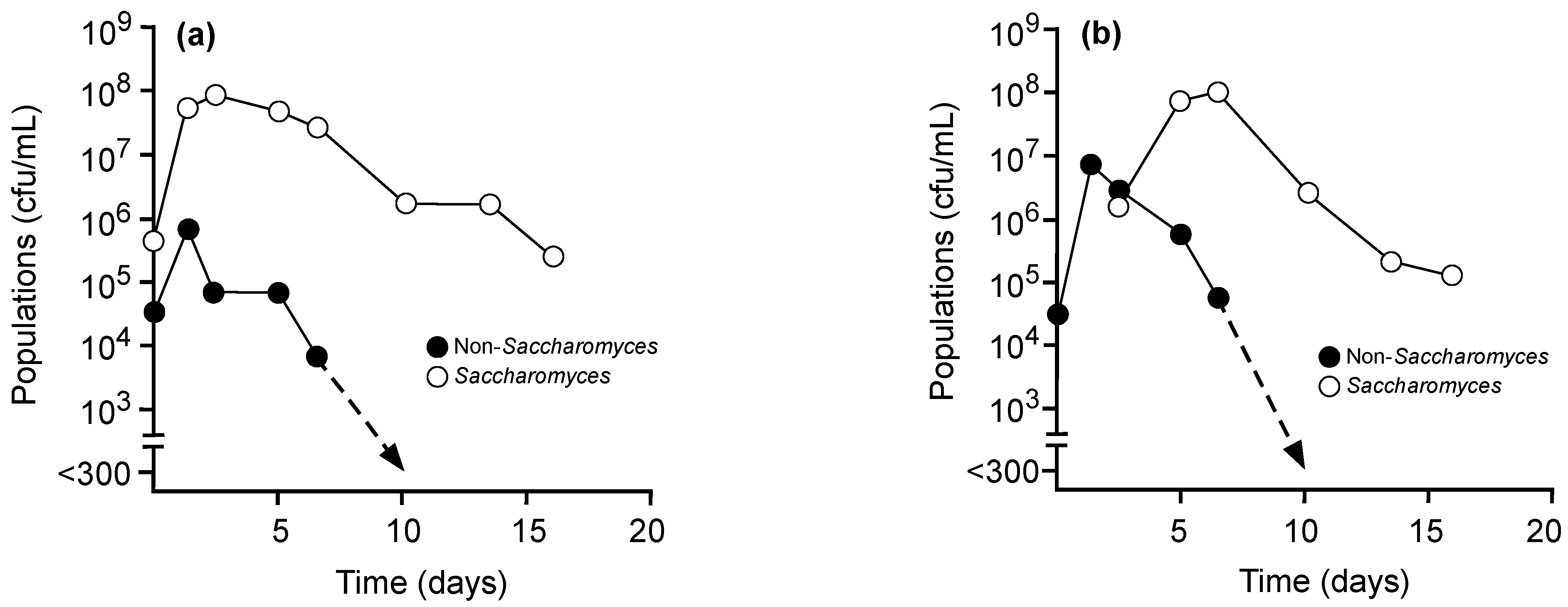

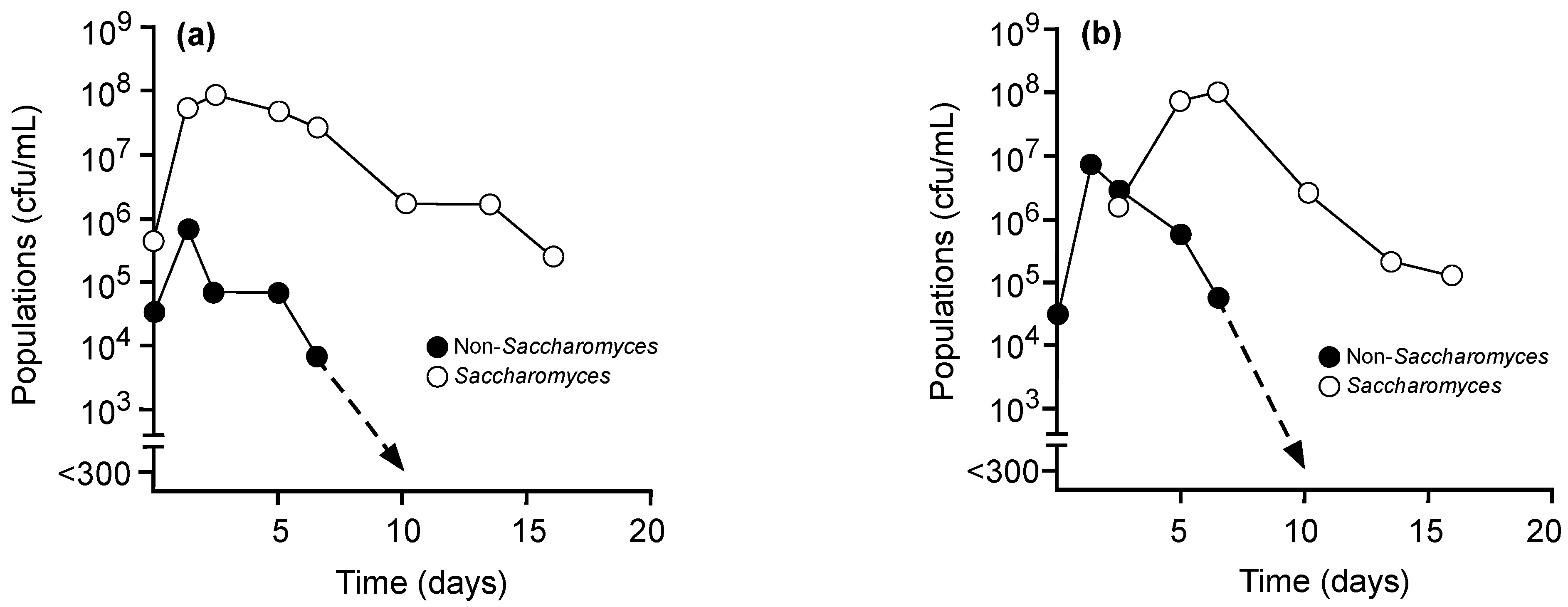{kind=link}
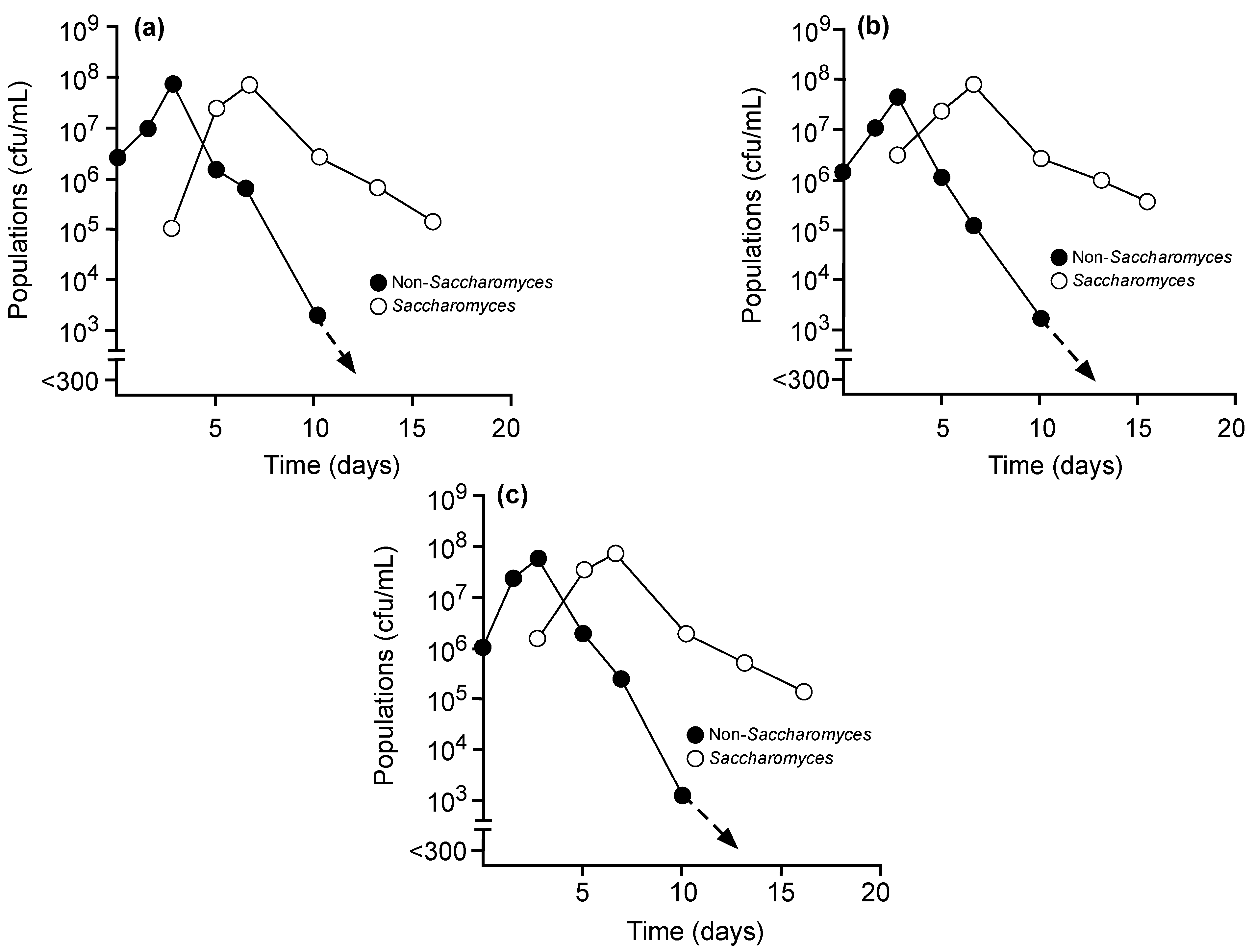
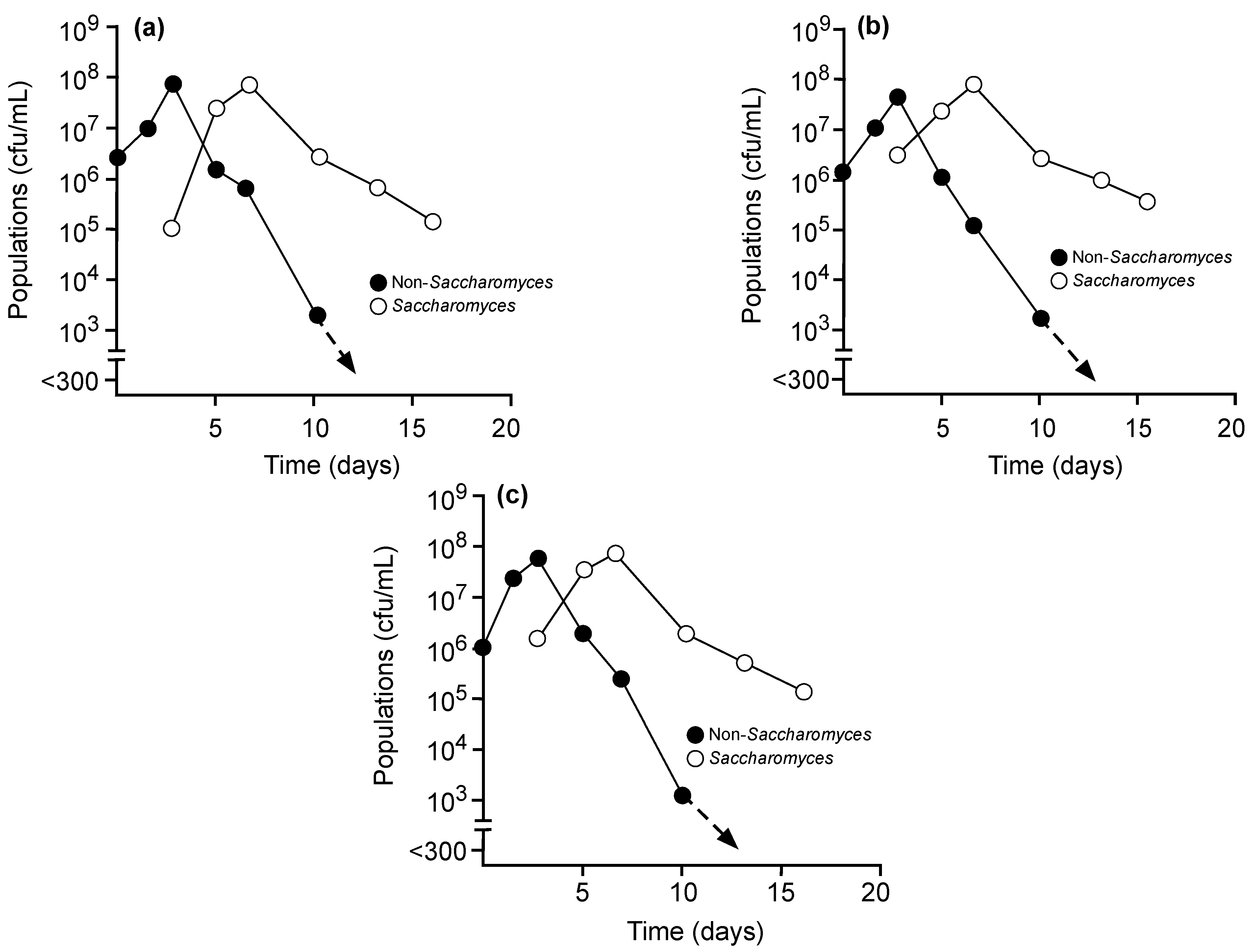{kind=link}
| Constituent | Inoculated Yeasts | ||||
|---|---|---|---|---|---|
| S. cerevisiae | S. cerevisiae * | P01A016 *,† | NS-MP *,† | P40D002 *,† | |
| Ethanol (% v/v) | 14.9 b | 14.9 b | 13.8 a | 13.9 a | 15.0 b |
| Glucose + fructose (g/L) | 0.109 a | 0.114 a | 0.119 a | 0.112 a | 0.130 a |
| pH | 3.31 c,d | 3.29 c | 3.23 b | 3.21 a | 3.29 c,d |
| Titratable acidity (g/L) | 5.56 a | 5.61 a | 6.09 b | 6.09 b | 6.81 c |
| Volatile acidity (g/L) | 0.37 b,c | 0.45 d | 0.35 a,b | 0.33 a | 0.40 c |
| Malic acid (g/L) | 2.22 b | 2.18 b | 1.38 a | 1.36 a | 2.29 b |
| Succinic acid (g/L) | 1.75 a | 1.73 a | 1.91 a | 1.74 a | 1.82 a |
| Glycerol (g/L) | 10.0 a | 10.1 a | 10.1 a | 10.1 a | 10.1 a |
| Higher Alcohols (mg/L) | |||||
| 1-Propanol | 2.52 a | 3.98 a | 5.33 a | 3.36 a | 4.60 a |
| 2-Methyl-1-propanol | 29.0 a | 64.5 b | 61.4 b | 64.5 b | 70.4 b |
| 2&3-Methyl-1-butanols | 108 a | 112 a | 137 b | 165 b | 144 b |
| 1-Hexanol | 1.18 a | 1.20 a | 0.975 a | 0.949 a | 1.18 a |
| 1-Octanol | 1.84 a | 0.662 a | 1.52 a | 0.735 a | 1.23 a |
| 2-Phenylethanol | 36.5 a | 37.3 a | 37.9 a | 37.8 a | 45.3 a |
| Esters (mg/L) | |||||
| Ethyl acetate | 52.3 a | 145 b | 73.1 a | 64.1 a | 148 b |
| 2- and 3-Methyl-butyl acetates | 0.712 a | 0.912 a b | 0.808 a,b | 0.858 a,b | 1.02 b |
| Hexyl acetate | 0.007 a | 0.007 a | 0.012 a | 0.009 a | 0.009 a |
| Diethyl succinate | 1.24 b | 0.787 a | 0.992 a,b | 1.00 a,b | 1.05 a,b |
| 2-Phenylethyl acetate | 0.016 a | 0.058 b,c | 0.043 a,b | 0.054 b,c | 0.073 c |
| Ethyl butanoate | 0.223 a | 0.166 a | 0.193 a | 0.190 a | 0.187 a |
| Ethyl hexanoate | 0.089 a | 0.024 a | 0.073 a | 0.038 a | 0.037 a |
| Ethyl octanoate | 0.541 b | 0.353 a | 0.388 a,b | 0.348 a | 0.355 a |
| Acids (mg/L) | |||||
| Hexanoic acid | 4.05 c | 2.39 a | 2.79 a,b | 3.90 b,c | 3.09 a,b,c |
| Octanoic acid | 4.15 a | 3.13 a | 3.79 a | 3.69 a | 3.63 a |
| Source of Variation | Pr > F (p) | Panelist (F) | Yeast (F) | Replicate (F) | Panelist * Yeast (F) |
|---|---|---|---|---|---|
| Degrees Freedom | 9 | 4 | 1 | 36 | |
| Aroma | |||||
| ‘Estery’ | 0.108 | 5.21 *** | 0.895 | 0.709 | 0.567 |
| ‘Pungency’ | 0.350 | 3.13 ** | 1.96 | 0.716 | 0.532 |
| ‘Fruity’ | 0.0004 † | 7.81 *** | 0.695 | 0.038 | 1.70 |
| ‘Dried fruit’ | 0.056 | 5.69 *** | 1.48 | 0.393 | 0.597 |
| ‘Green’ | 0.709 | 1.58 | 0.306 | 0.005 | 0.759 |
| ‘Yeasty’ | 0.095 | 4.56 ** | 0.861 | 0.685 | 0.768 |
| ‘Ethanol’ | <0.0001 † | 17.5 *** | 2.10 | 2.92 | 0.707 |
| ‘Berry’ | 0.018 † | 4.99 *** | 1.39 | 0.043 | 1.15 |
| ‘Earthy’ | 0.778 | 0.852 | 0.847 | 1.33 | 0.772 |
| ‘Woody’ | 0.096 | 4.59 ** | 0.952 | 3.88 | 0.659 |
| ‘Sulfur’ | 0.496 | 3.97 ** | 0.374 | 4.94 * | 0.221 |
| ‘Sweaty’ | 0.054 | 1.91 | 0.957 | 0.590 | 1.61 |
| ‘Chemical’ | <0.0001 † | 12.2 *** | 0.911 | 0.756 | 1.12 |
| ‘Vegetal’ | 0.190 | 1.66 | 2.01 | 0.017 | 1.15 |
| ‘Barn’ | 0.044 † | 4.72 ** | 0.949 | 1.06 | 0.952 |
| Taste | |||||
| ‘Sweet’ | 0.014 † | 7.03 *** | 0.356 | 0.041 | 0.815 |
| ‘Bitter’ | 0.0002 † | 12.5 *** | 0.934 | 0.478 | 0.678 |
| ‘Sour’ | 0.001 † | 9.02 *** | 1.37 | 0.527 | 1.057 |
| Mouthfeel | |||||
| ‘Viscosity’ | 0.041 † | 4.23 ** | 1.46 | 0.177 | 1.06 |
| ‘Hot’ | 0.008 † | 7.06 *** | 1.62 | 0.824 | 0.803 |
| ‘Astringent’ | <0.0001 † | 16.6 *** | 1.01 | 0.857 | 1.05 |
| ‘Roughness’ | <0.0001 † | 18.5 *** | 1.55 | 0.482 | 1.25 |
| Flavor | |||||
| ‘Estery’ | 0.094 | 5.25 *** | 0.740 | 2.99 | 0.550 |
| ‘Pungency’ | 0.001 † | 9.88 *** | 0.857 | 0.157 | 0.958 |
| ‘Fruity’ | 0.222 | 3.72 ** | 0.235 | 0.872 | 0.749 |
| ‘Dried fruit’ | 0.001 † | 8.09 *** | 1.72 | 2.06 | 1.12 |
| ‘Green’ | 0.007 † | 7.26 *** | 0.635 | 0.022 | 0.949 |
| ‘Yeasty’ | 0.0003 † | 8.26 *** | 1.29 | 2.35 | 1.57 |
| ‘Ethanol’ | <0.0001 † | 16.5 *** | 1.71 | 6.72 * | 1.67 * |
| ‘Berry’ | 0.013 † | 4.29 ** | 1.64 | 3.40 | 1.29 |
| ‘Earthy’ | 0.017 † | 1.96 | 1.33 | 0.21 | 1.90 * |
| ‘Woody’ | 0.287 | 3.01 ** | 0.740 | 0.961 | 0.771 |
| ‘Sulfur’ | 0.088 | 4.12 ** | 0.114 | 8.66 ** | 0.764 |
| ‘Sweaty’ | 0.020 † | 2.41 * | 1.43 | 2.41 | 1.69 * |
| ‘Chemical’ | <0.0001 † | 22.3 *** | 0.676 | 0.097 | 1.31 |
| ‘Vegetal’ | 0.802 | 0.753 | 0.645 | 0.122 | 0.827 |
| ‘Barn’ | 0.118 | 4.32 ** | 1.20 | 1.27 | 0.703 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aplin, J.J.; Paup, V.D.; Ross, C.F.; Edwards, C.G. Chemical and Sensory Profiles of Merlot Wines Produced by Sequential Inoculation of Metschnikowia pulcherrima or Meyerzyma guilliermondii. Fermentation 2021, 7, 126. https://doi.org/10.3390/fermentation7030126
Aplin JJ, Paup VD, Ross CF, Edwards CG. Chemical and Sensory Profiles of Merlot Wines Produced by Sequential Inoculation of Metschnikowia pulcherrima or Meyerzyma guilliermondii. Fermentation. 2021; 7(3):126. https://doi.org/10.3390/fermentation7030126
Chicago/Turabian StyleAplin, Jesse J., Victoria D. Paup, Carolyn F. Ross, and Charles G. Edwards. 2021. "Chemical and Sensory Profiles of Merlot Wines Produced by Sequential Inoculation of Metschnikowia pulcherrima or Meyerzyma guilliermondii" Fermentation 7, no. 3: 126. https://doi.org/10.3390/fermentation7030126
APA StyleAplin, J. J., Paup, V. D., Ross, C. F., & Edwards, C. G. (2021). Chemical and Sensory Profiles of Merlot Wines Produced by Sequential Inoculation of Metschnikowia pulcherrima or Meyerzyma guilliermondii. Fermentation, 7(3), 126. https://doi.org/10.3390/fermentation7030126