Impact on Sensory and Aromatic Profile of Low Ethanol Malbec Wines Fermented by Sequential Culture of Hanseniaspora uvarum and Saccharomyces cerevisiae Native Yeasts

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Yeast Inoculum Preparation
2.3. Grapes and Vineyard Location
2.4. Inoculation and Winemaking
2.5. Chemical Analysis
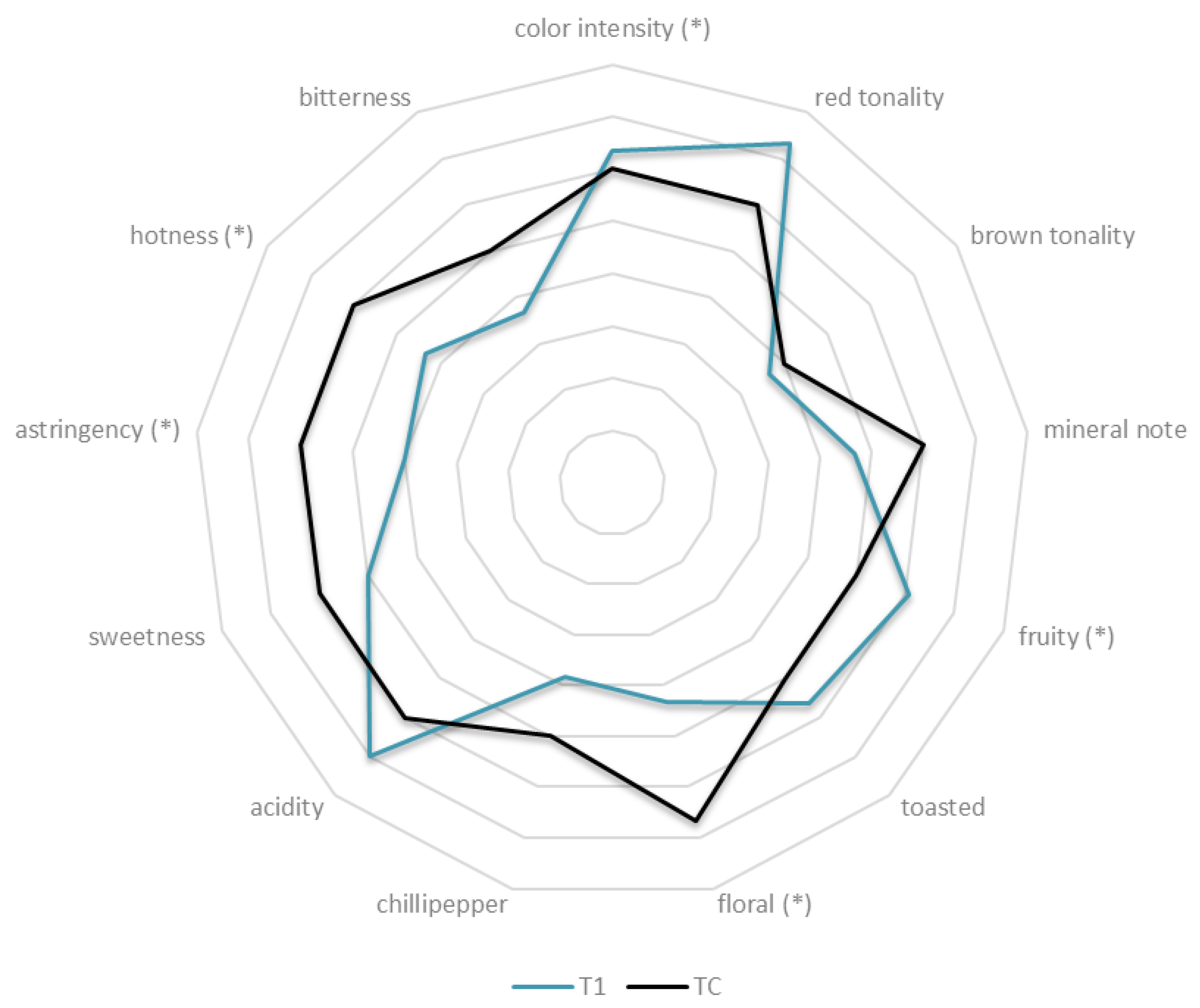
2.6. Sensorial Analysis
2.7. Free aromatic Analyses
2.7.1. Solid Phase Extraction (SPE)
2.7.2. GC-MS Analyses
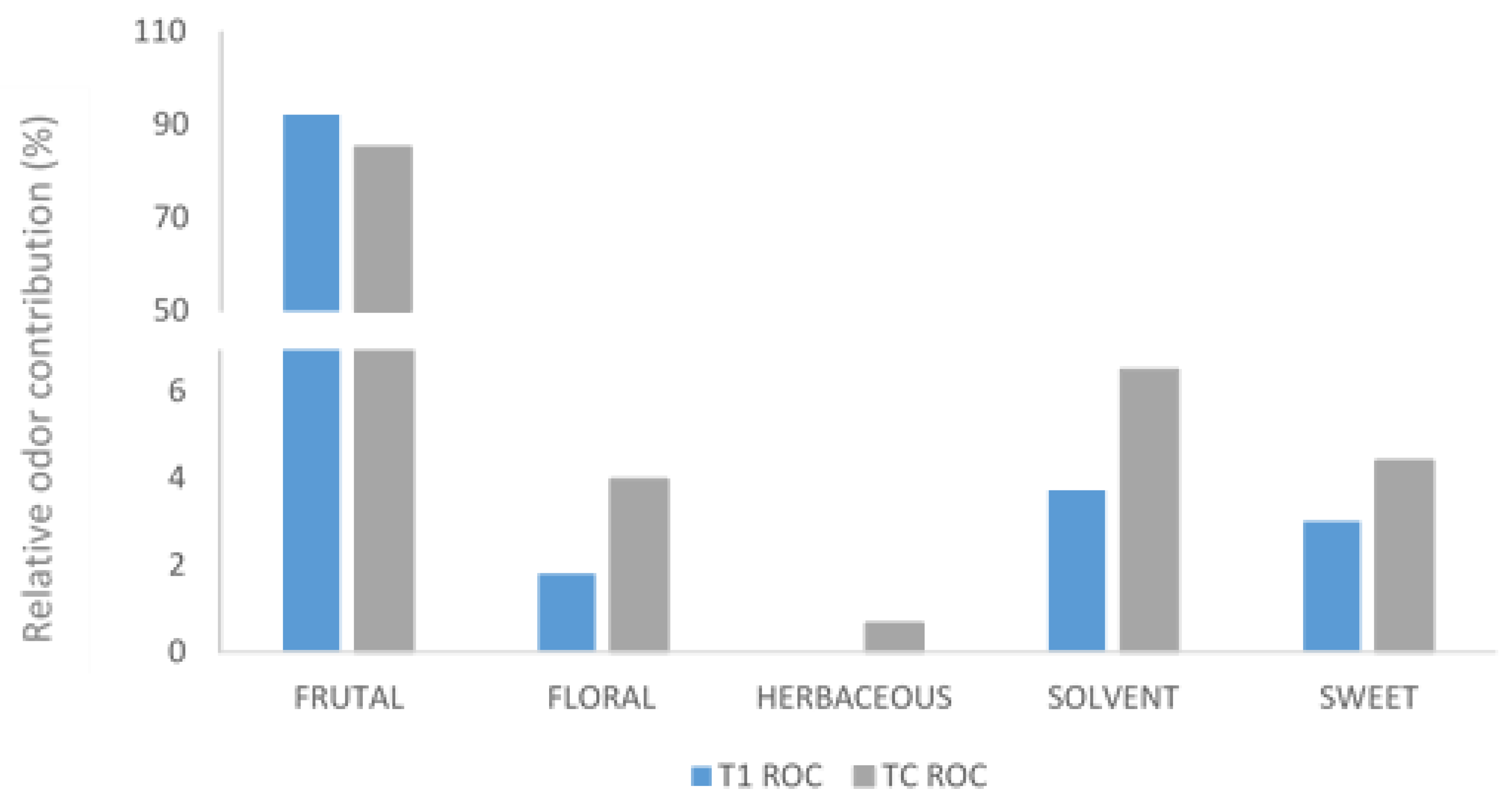
2.8. Odor Activity Value (OAV) and Relative Odor Contributions (ROCs)
2.9. Statistical Analysis
3. Results
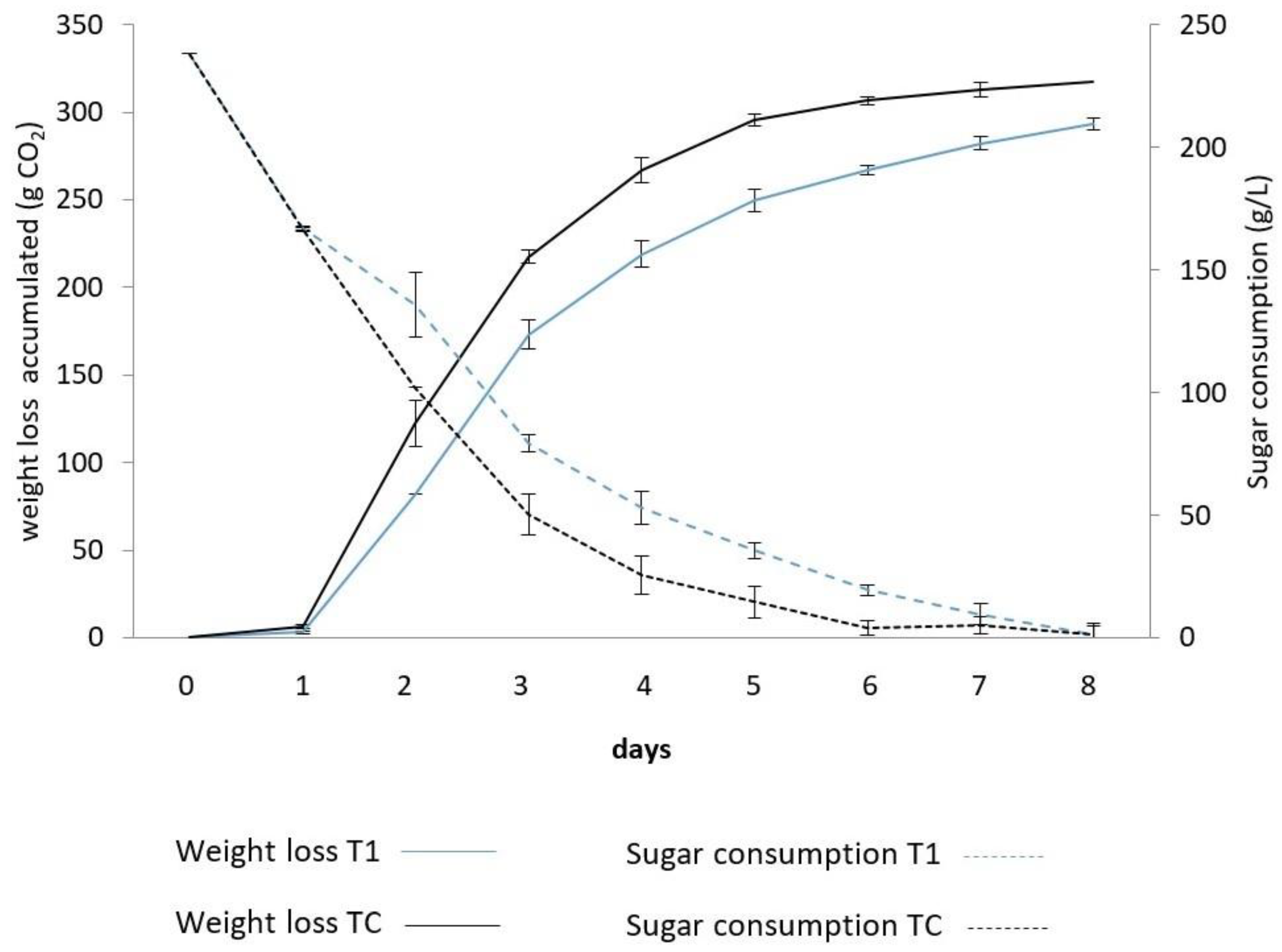
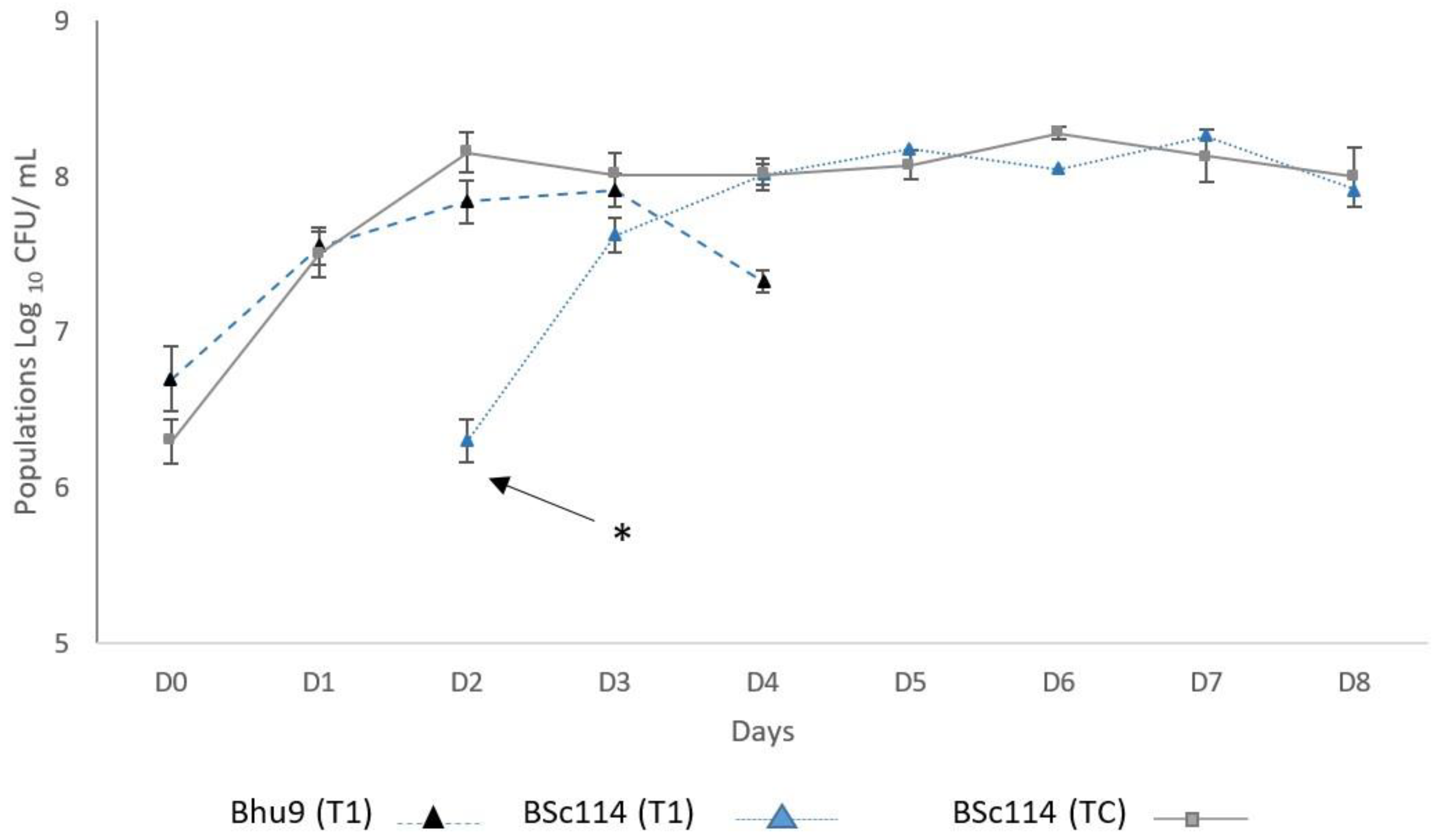
3.1. Fermentative Kinetics and Population Dynamics
3.2. Enological Parameters
3.3. Aromatic Composition
3.4. Sensorial Analysis
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Zamora, F. Dealcoholised Wines and Low-Alcohol Wines. In Wine Safety, Consumer Preference, and Human Health; Springer: Cham, Switzerland, 2016; pp. 163–182. [Google Scholar]
- De Orduna, R.M. Climate change associated effects on grape and wine quality and production. Food Res. Int. 2010, 43, 1844–1855. [Google Scholar] [CrossRef]
- Meillon, S.; Dugas, V.; Urbano, C.; Schlich, P. Preference and acceptability of partially dealcoholized white and red wines by consumers and professionals. Am. J. Enol. Vitic. 2010, 61, 42–52. [Google Scholar]
- Contreras, A.; Hidalgo, C.; Henschke, P.A.; Chambers, P.J.; Curtin, C.; Varela, C. Evaluation of non-Saccharomyces yeasts for the reduction of alcohol content in wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed]
- Fischer, U.; Noble, A.C. The effect of ethanol, catechin concentration, and pH on sourness and bitterness of wine. Am. J. Enol. Vitic. 1994, 45, 6–10. [Google Scholar]
- King, E.S.; Heymann, H. The effect of reduced alcohol on the sensory profiles and consumer preferences of white wine. J. Sens. Stud. 2014, 29, 33–42. [Google Scholar] [CrossRef]
- Kliewer, W.M.; Dokoozlian, N.K. Leaf area/crop weight ratios of grapevines: Influence on fruit composition and wine quality. Am. J. Enol. Vitic. 2005, 56, 170–181. [Google Scholar]
- Walker, R.R.; Blackmore, D.H.; Clingeleffer, P.R.; Tarr, C.R. Rootstock effects on salt tolerance of irrigated field-grown grapevines (Vitis vinifera L. cv. Sultana). 3. Fresh fruit composition and dried grape quality. Aust. J. Grape Wine Res. 2007, 13, 130–141. [Google Scholar] [CrossRef]
- Chorti, E.; Guidoni, S.; Ferrandino, A.; Novello, V. Effect of different cluster sunlight exposure levels on ripening and anthocyanin accumulation in Nebbiolo grapes. Am. J. Enol. Vitic. 2010, 61, 23–30. [Google Scholar]
- Palliotti, A.; Panara, F.; Silvestroni, O.; Lanari, V.; Sabbatini, P.; Howell, G.S.; Gatti, M.; Poni, S. Influence of mechanical postveraison leaf removal apical to the cluster zone on delay of fruit ripening in Sangiovese (Vitis vinifera L.) grapevines. Aust. J. Grape Wine Res. 2013, 19, 369–377. [Google Scholar] [CrossRef]
- Schmidtke, L.M.; Blackman, J.W.; Agboola, S.O. Production technologies for reduced alcoholic wines. J. Food Sci. 2012, 77, R25–R41. [Google Scholar] [CrossRef]
- Catarino, M.; Mendes, A. Dealcoholizing wine by membrane separation processes. Innov. Food Sci. Emerg. 2011, 12, 330–337. [Google Scholar] [CrossRef]
- Belisario-Sanchez, Y.Y.; Taboada-Rodriguez, A.; Marin-Iniesta, F.; Lopez-Gomez, A. Dealcoholized wines by spinning cone column distillation: Phenolic compounds and antioxidant activity measured by the 1, 1-diphenyl-2-picrylhydrazyl method. J. Agric. Food Chem. 2009, 57, 6770–6778. [Google Scholar] [CrossRef] [PubMed]
- Quirós, M.; Rojas, V.; Gonzalez, R.; Morales, P. Selection of non-Saccharomyces yeast strains for reducing alcohol levels in wine by sugar respiration. Int. J. Food Microbiol. 2014, 181, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Dry, P.R.; Kutyna, D.R.; Francis, I.L.; Henschke, P.A.; Curtin, C.D.; Chambers, P.J. Strategies for reducing alcohol concentration in wine. Aust. J. Grape Wine Res. 2015, 21, 670–679. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef] [PubMed]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef]
- Gonzalez, R.; Quirós, M.; Morales, P. Yeast respiration of sugars by non-Saccharomyces yeast species: A promising and barely explored approach to lowering alcohol content of wines. Trends Food Sci. Technol. 2013, 29, 55–61. [Google Scholar] [CrossRef]
- Martin, V.; Valera, M.; Medina, K.; Boido, E.; Carrau, F. Oenological impact of the Hanseniaspora/Kloeckera yeast genus on wines—A review. Fermentation 2018, 4, 76. [Google Scholar] [CrossRef]
- Mestre Furlani, M.V.; Maturano, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of non-Saccharomyces yeasts to be used in grape musts with high alcoholic potential: A strategy to obtain wines with reduced ethanol content. FEMS Yeast Res. 2017, 17. [Google Scholar] [CrossRef]
- Maturano, Y.P.; Mestre, M.V.; Kuchen, B.; Toro, M.E.; Mercado, L.A.; Vazquez, F.; Combina, M. Optimization of fermentation-relevant factors: A strategy to reduce ethanol in red wine by sequential culture of native yeasts. Int. J. Food Microbiol. 2019, 289, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL medium to profile native flora fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- Lawless, H.T.; Heymann, H. Physiological and psychological foundations of sensory function. In Sensory Evaluation of Food; Springer: New York, NY, USA, 2010; pp. 19–56. [Google Scholar]
- Boido, E.; Lloret, A.; Medina, K.; Fariña, L.; Carrau, F.; Versini, G.; Dellacassa, E. Aroma composition of Vitis vinifera cv. Tannat: The typical red wine from Uruguay. J. Agric. Food Chem. 2003, 51, 5408–5413. [Google Scholar] [CrossRef] [PubMed]
- Mondello, L. Mass Spectra of Flavors and Fragrances of Natural and Synthetic Compounds, 3rd ed.; MS Wil GmbH: Zurich, Switzerland, 2015. [Google Scholar]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured publishing corporation: Carol Stream, IL, USA, 2007; Volume 456. [Google Scholar]
- McLafferty, F.W.; Stauffer, D.B. The Wiley/NBS Registry of Mass Spectral Data; Wiley and Sons: New York, NY, USA, 1991. [Google Scholar]
- Hellin, P.; Manso, A.; Flores, P.; Fenoll, J. Evolution of aroma and phenolic compounds during ripening of superior seedless grapes. J. Agric. Food Chem. 2010, 58, 6334–6340. [Google Scholar] [CrossRef] [PubMed]
- Welke, J.E.; Zanus, M.; Lazzarotto, M.; Zini, C.A. Quantitative analysis of headspace volatile compounds using comprehensive two-dimensional gas chromatography and their contribution to the aroma of Chardonnay wine. Food Res. Int. 2014, 59, 85–99. [Google Scholar] [CrossRef]
- Tristezza, M.; Tufariello, M.; Capozzi, V.; Spano, G.; Mita, G.; Grieco, F. The oenological potential of Hanseniaspora uvarum in simultaneous and sequential co-fermentation with Saccharomyces cerevisiae for industrial wine production. Front. Microbiol. 2016, 7, 670. [Google Scholar] [CrossRef] [PubMed]
- Fleet, G.H.; Heard, G.M. Yeasts-growth during fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Harwood Academic, Eds.; CRC Press: Chur, Switzerland, 1993; pp. 27–54. [Google Scholar]
- Mendoza, L.M.; de Nadra, M.C.M.; Farías, M.E. Kinetics and metabolic behavior of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 2007, 29, 1057–1063. [Google Scholar] [CrossRef] [PubMed]
- Domizio, P.; Romani, C.; Lencioni, L.; Comitini, F.; Gobbi, M.; Mannazzu, I.; Ciani, M. Outlining a future for non-Saccharomyces yeasts: Selection of putative spoilage wine strains to be used in association with Saccharomyces cerevisiae for grape juice fermentation. Int. J. Food Microbiol. 2011, 147, 170–180. [Google Scholar] [CrossRef]
- Suzzi, G.; Arfelli, G.; Schirone, M.; Corsetti, A.; Perpetuini, G.; Tofalo, R. Effect of grape indigenous Saccharomyces cerevisiae strains on Montepulciano d’Abruzzo red wine quality. Food Res. Int. 2012, 46, 22–29. [Google Scholar] [CrossRef]
- Bisson, L.F. Stuck and sluggish fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar]
- Milanovic, V.; Ciani, M.; Oro, L.; Comitini, F. Starmerella bombicola influences the metabolism of Saccharomyces cerevisiae at pyruvate decarboxylase and alcohol dehydrogenase level during mixed wine fermentation. Microb. Cell Factories 2012, 11, 18. [Google Scholar] [CrossRef] [PubMed]
- Petitgonnet, C.; Klein, G.L.; Roullier-Gall, C.; Schmitt-Kopplin, P.; Quintanilla-Casas, B.; Vichi, S.; Diane, J.D.; Alexandre, H. Influence of cell-cell contact between L. thermotolerans and S. cerevisiae on yeast interactions and the exo-metabolome. Food Microbiol. 2019, 83, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Mas, A.; Esteve-Zarzoso, B. Interaction between Hanseniaspora uvarum and Saccharomyces cerevisiae during alcoholic fermentation. Int. J. Food Microb. 2015, 206, 67–74. [Google Scholar] [CrossRef] [PubMed]
- Erten, H.; Campbell, I. The production of low-alcohol wines by aerobic yeasts. J. Inst. Brew. 1953, 59, 207–215. [Google Scholar] [CrossRef]
- Giudici, P.; Romano, P.; Zambonelli, C. A biometric study of higher alcohol production in Saccharomyces cerevisiae. Can. J. Microbiol. 1990, 36, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Mateos, J.R.; Pérez-Nevado, F.; Fernández, M.R. Influence of Saccharomyces cerevisiae yeast strain on the major volatile compounds of wine. Enzym. Microb. Technol. 2006, 40, 151–157. [Google Scholar] [CrossRef]
- Guth, H.; Sies, A. Flavour of wines: Towards an understanding by reconstitution experiments and an analysis of ethanol’s effect on odour activity of key compounds. In Proceedings of the 11th Australian Wine Industry Technical Conference, Glen Osmond, SA, Australia, 7–11 October 2002; pp. 128–139. [Google Scholar]
- Conner, J.M.; Birkmyre, L.; Paterson, A.; Piggott, J.R. Headspace concentrations of ethyl esters at different alcoholic strengths. J. Sci. Food Agric. 1998, 77, 121–126. [Google Scholar] [CrossRef]
- Tilloy, V.; Ortiz-Julien, A.; Dequin, S. Reduction of ethanol yield and improvement of glycerol formation by adaptive evolution of the wine yeast Saccharomyces cerevisiae under hyperosmotic conditions. Appl. Environ. Microbiol. 2014, 80, 2623–2632. [Google Scholar] [CrossRef]
- Escudero, A.; Gogorza, B.; Melus, M.A.; Ortin, N.; Cacho, J.; Ferreira, V. Characterization of the aroma of a wine from Maccabeo. Key role played by compounds with low odor activity values. J. Agric. Food Chem. 2004, 52, 3516–3524. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Compounds | T1 | TC |
|---|---|---|
| Ethanol (%v/v) | 12.63 ± 0.05 | 13.5 ± 0.28 (*) |
| Acetic acid (g/L) | 0.56 ± 0.02 | 0.49 ± 0.04 |
| Lactic acid (g/L) | 0.5 ± 0.01 | 0.47 ± 0.02 |
| Malic acid (g/L) | 2.85 ± 0.21 | 2.85 ± 0.07 |
| Tartaric acid (g/L) | 1.31 ± 0.08 | 1 ± 0.01 (*) |
| Glycerol (g/L) | 10 ± 0.71 | 9.3 ± 0.57 |
| pH | 3.43 ± 0.00 | 3.49 ± 0.01 (*) |
| Total acidity (g/L) | 5.95 ± 0.07 | 5.80 ± 0.14 |
| Residual sugar (g/L) | 1.55 ± 0.64 | 1.70 ± 0.28 |
| Compounds (µg/L) | Treatments | Descriptor | Threshold Perception (µg/L) | Ref | OAV T1 | OAV TC | |||
|---|---|---|---|---|---|---|---|---|---|
| T1 | TC | p < 0.05 | Aromatic Serie | ||||||
| Ethyl esters | |||||||||
| Ethyl hexanoate | 600 ± 15 | 200 ± 83 | * | fruity, apple | 14 | 1 | 42.85 | 14.28 | 5 |
| Ethyl octanoate | 872 ± 58 | 369 ± 187 | * | pineapple, pear | 5 | 1 | 174.4 | 73.8 | 5 |
| Ethyl-3-hydroxybutanoate | 212 ± 53 | 174 ± 14 | grape, caramel | 67,000 | 2 | 0.003 | 0.002 | 2 | |
| Ethyl decanoate | 1140 ± 40 | 590 ± 81 | * | floral | 200 | 1 | 5.7 | 2.95 | 4 |
| Ethyl dodecanoate | 672 ± 52 | nd | leaf, fruity | 1500 | 1 | 0.448 | - | 5 | |
| Ethyl tetradecanoate | 31 ± 1 | 121 ± 22 | * | waxy | 2000 | 1 | 0.015 | 0.060 | 6 |
| Ethyl palmitate | 345 ± 23 | 224 ± 14 | * | waxy | 1500 | 1 | 0.23 | 0.149 | 6 |
| Ethyl succinate | 127 ± 0.011 | 145 ± 6 | * | ripe melon | 1,000,000 | 1 | 0.0001 | 0.0001 | 5 |
| Ethyl lactate | 603 ± 21 | 409 ± 63 | * | strawberrry | 14,000 | 1 | 0.043 | 0.029 | 5 |
| ∑ Ethyl esters | 4600 | 2232 | |||||||
| Acetate esters | |||||||||
| Isoamyl acetate | 3210 ± 18 | 2892 ± 191 | * | banana | 30 | 2 | 107 | 96.4 | 5 |
| Hexyl acetate | 24 ± 24 | 297 ± 11 | * | red fruit | 1500 | 1 | 0.03 | 0.443 | 5 |
| ∑ acetate esters | 3234 | 3189 | |||||||
| TOTAL ESTERS | 7491 (1.77%) | 5421 (1.03%) | |||||||
| Higher Alcohols | |||||||||
| 2-Methyl-1-propanol | 18,480 ± 1620 | 12,990 ± 299 | * | solvent | 7000 | 3 | 2.64 | 1.85 | 1 |
| 1-Butanol | 444 ± 1 | 692 ± 72 | * | solvent | 9000 | 3 | 0.049 | 0.076 | 1 |
| 3-Methyl-1-butanol | 318,400 ± 12,300 | 371,000 ± 16,140 | * | burned, alcohol | 30,000 | 3 | 10.61 | 12.36 | 1 |
| 1-Pentanol | 40 ± 2 | nd | fruity, balsmic | 4000 | 3 | 0.01 | - | 5 | |
| 4-Methyl-1-pentanol | 59 ± 8 | 60 ± 18 | almond | 50,000 | 3 | 0.001 | 0.001 | 2 | |
| 3-Methyl-1-pentanol | 214 ± 18 | 220 ± 10 | herbaceous | 50,000 | 3 | 0.004 | 0.044 | 3 | |
| 1-Hexanol | 1080 ± 142 | 852 ± 82 | grass, green leaf | 2500 | 4 | 0.432 | 0.340 | 3 | |
| trans-3-Hexenol | 40 ± 0 | nd | herbaceous, land | 400 | 1 | 0.1 | - | 3 | |
| 3-Ethoxy-1-propanol | 150 ± 11 | 65 ± 3 | * | ripe pear | 100 | 1 | 1.5 | 0.65 | 5 |
| cis-3-Hexenol | 54 ± 8 | 57 ± 8 | cutted grass | 400 | 1 | 0.135 | 0.014 | 6 | |
| 2-Ethyl hexanol | 256 ± 12 | 255 ± 17 | rose, citrus | 8000 | 1 | 0.032 | 0.031 | 4 | |
| 2,3-Butanediol | 233 ± 15 | 340 ± 13 | * | butter | 120,000 | 1 | 0.001 | 0.002 | 6 |
| Furfurol | 102 ± 19 | 190 ± 20 | * | floral | 5000 | 4 | 0.02 | 0.038 | 4 |
| 3-(Methylthio)-1-propanol | 819 ± 41 | 1450 ± 38 | * | cooked vegetal | 1000 | 5 | 0.819 | 1.45 | 3 |
| Benzyl alcohol | 610 ± 50 | 11 ± 4 | * | caramelo, cítrico | 10,000 | 4 | 0.061 | 0.0001 | 2 |
| 2-Phenylethyl alcohol | 53,885 ± 3012 | 86,072 ± 731 | * | rose | 14,000 | 4 | 3.848 | 6.148 | 2 |
| Tyrosol | 17,110 ± 895 | 23,071 ± 3245 | * | honey | - | - | 2 | ||
| Tryptophol | 1910 ± 98 | 1780 ± 98 | * | - | - | ||||
| ∑ Higher alcohols | 414,357 (98.14%) | 515,754 (98.45%) | |||||||
| Fatty acids | |||||||||
| Acetic acid | 39 ± 6 | nd | vinegar | 200 | 1 | 0.195 | - | 6 | |
| Isobutanoic acid | 181 ± 21 | 396 ± 9 | * | butter, cheese | 8100 | 3 | 0.022 | 0.048 | 6 |
| Butanoic acid | 56 ± 3 | 295 ± 29 | * | fatty, rancid | 1000 | 3 | 0.056 | 0.295 | 6 |
| Hexanoic acid | 176 ± 53 | 231 ± 42 | * | cheese, sudor | 3000 | 3 | 0.058 | 0.077 | 6 |
| Octanoic acid | 235 ± 50 | 824 ± 9 | * | rancid butter | 3000 | 3 | 0.078 | 0.276 | 6 |
| Decanoic acid | 94 ± 9 | 773 ± 310 | * | fatty, rancid | 10,000 | 3 | 0.009 | 0.007 | 6 |
| Dodecanoic acid | 49 ± 1 | 43 ± 1 | fatty, rancid | 10,000 | 3 | 0.005 | 0.004 | 6 | |
| ∑ Acids | 83 (0.019%) | 2562 (0.49%) | |||||||
| Lactones | |||||||||
| gamma-Valerolactone | 39 ± 1 | 49 ± 3 | * | sweet, cocconut | 10 | 7 | 3.9 | 4.9 | 2 |
| gamma-Butyrolactone | 236 ± 8 | 163 ± 26 | * | caramel | 35 | 7 | 6.74 | 4.65 | 2 |
| ∑ Lactones | 275 (0.06%) | 212 (0.04%) | |||||||
| ∑ compounds (µg/L) | 422,206 | 523,858 |
| T1 | TC | ||||
|---|---|---|---|---|---|
| Compounds | OAV | ROC (%) | OAV | ROC (%) | Aromatic Serie |
| Ethyl hexanoate | 42.85 | 12.12 | 14.28 | 6.60 | 5 Fruity |
| Ethyl octanoate | 174.40 | 49.35 | 73.8 | 34.09 | 5 Fruity |
| Ethyl decano ate | 1.03 | 0.29 | 0 | 0.00 | 4 Floral |
| Isoamyl acetate | 107.00 | 30.28 | 96.40 | 44.53 | 5 Fruity |
| 2-Methyl-1-propanol | 2.64 | 3.72 | 1.85 | 6.51 | 1 Solvent |
| 3-Methyl-1-butanol | 10.61 | 3.00 | 12.36 | 5.71 | 1 Solvent |
| 3-Ethoxy-1-propanol | 1.50 | 0.42 | <1 | - | 5 Fruity |
| 3-(Methylthio)-1-propanol | <1 | - | 1.45 | 0.67 | 3 Herbaceous |
| 2-Phenylethanol | 5.38 | 1.52 | 8.61 | 3.98 | 4 Floral |
| gamma-Valerolactone | 3.90 | 1.10 | 4.90 | 2.26 | 2 Sweet |
| gamma-Butirolactone | 6.742 | 1.91 | 4.66 | 2.15 | 2 Sweet |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mestre, M.V.; Maturano, Y.P.; Gallardo, C.; Combina, M.; Mercado, L.; Toro, M.E.; Carrau, F.; Vazquez, F.; Dellacassa, E. Impact on Sensory and Aromatic Profile of Low Ethanol Malbec Wines Fermented by Sequential Culture of Hanseniaspora uvarum and Saccharomyces cerevisiae Native Yeasts. Fermentation 2019, 5, 65. https://doi.org/10.3390/fermentation5030065
Mestre MV, Maturano YP, Gallardo C, Combina M, Mercado L, Toro ME, Carrau F, Vazquez F, Dellacassa E. Impact on Sensory and Aromatic Profile of Low Ethanol Malbec Wines Fermented by Sequential Culture of Hanseniaspora uvarum and Saccharomyces cerevisiae Native Yeasts. Fermentation. 2019; 5(3):65. https://doi.org/10.3390/fermentation5030065
Chicago/Turabian StyleMestre, María Victoria, Yolanda Paola Maturano, Candelaria Gallardo, Mariana Combina, Laura Mercado, María Eugenia Toro, Francisco Carrau, Fabio Vazquez, and Eduardo Dellacassa. 2019. "Impact on Sensory and Aromatic Profile of Low Ethanol Malbec Wines Fermented by Sequential Culture of Hanseniaspora uvarum and Saccharomyces cerevisiae Native Yeasts" Fermentation 5, no. 3: 65. https://doi.org/10.3390/fermentation5030065
APA StyleMestre, M. V., Maturano, Y. P., Gallardo, C., Combina, M., Mercado, L., Toro, M. E., Carrau, F., Vazquez, F., & Dellacassa, E. (2019). Impact on Sensory and Aromatic Profile of Low Ethanol Malbec Wines Fermented by Sequential Culture of Hanseniaspora uvarum and Saccharomyces cerevisiae Native Yeasts. Fermentation, 5(3), 65. https://doi.org/10.3390/fermentation5030065