Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae


1
Department of Chemical Engineering, Texas A&M University, College Station, TX 77843, USA
2
Synthetic and Systems Biology Innovation Hub, Texas A&M University, College Station, TX 77843, USA
3
Department of Plant Pathology and Microbiology, Texas A&M University, College Station, TX 77843, USA
*
Author to whom correspondence should be addressed.
Fermentation 2019, 5(1), 6; https://doi.org/10.3390/fermentation5010006
Submission received: 8 October 2018
/
Revised: 13 December 2018
/
Accepted: 24 December 2018
/
Published: 1 January 2019
(This article belongs to the Special Issue Postgenomic Microbial Physiology and Fermentation)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The use of seawater in fermentation can potentially reduce the freshwater burden in the bio-based production of chemicals and fuels. We previously developed a Saccharomyces cerevisiae carotenoids hyperproducer SM14 capable of accumulating 18 mg g−1 DCW (DCW: dry cell weight) of β-carotene in rich media (YPD). In this work, the impacts of seawater on the carotenoid production of SM14 were investigated. When using nutrient-reduced media (0.1× YNB) in freshwater the β-carotene production of SM14 was 6.51 ± 0.37 mg g−1 DCW; however in synthetic seawater, the production was increased to 8.67 ± 0.62 mg g−1 DCW. We found that this improvement was partially due to the NaCl present in the synthetic seawater, since supplementation of 0.5 M NaCl in freshwater increased β-carotene production to 11.85 ± 0.77 mg g−1 DCW. The combination of synthetic seawater with higher carbon-to-nitrogen ratio (C:N = 50) further improved the β-carotene production to 10.44 ± 0.35 mg g−1 DCW. We further showed that the carotenoid production improvement in these conditions is related with lipid content and composition. These results demonstrated the benefit of using seawater to improve the production of carotenoids in S. cerevisiae, and have the potential to expand the utilization of seawater.
1. Introduction
Carotenoids belong to a large class of pigmented compounds naturally produced by plants, algae, fungi, and bacteria [1,2,3,4]. In the native organisms, these compounds serve as natural colorants, photoprotective agents, and antioxidants [1,3,5,6]. Carotenoids also play roles as vitamin A precursors, antioxidants, and antimicrobials in animals [5,7]. Thus, these compounds have potential applications in the food, cosmetics, and health industries [1,6,8]. Currently, most of the carotenoids produced in industry are extracted from natural sources such as plants and algae, or chemically synthesized. However, the structural complexity of most carotenoids make them difficult to be synthesized chemically, and the solvent-based chemical extraction from plants and algae faces challenges due to unpredictable feedstock availability [6,9]. Thus, there have been extensive efforts to engineer microbial hosts for their production [10,11]. In our prior work, we developed a novel evolution engineering approach based on the antioxidant potential of β-carotene and successfully generated carotenoid hyper-producing strains of Saccharomyces cerevisiae [12]. Briefly, crtE, crtYB, and crtI were introduced into the S. cerevisiae strain GSY1136 [13] via the integration vector YIplac211YB/I/E* [10]. To test the hypothesis that production of β-carotene can protect the cells from oxidative stress, the catalase gene CTT1 was deleted to remove the native capacity of yeast for detoxifying hydrogen peroxide. The resulting strain YLH2 (GSY1136 YIplac211YB/I/E* ΔCTT1) was evolved by periodic challenge with hydrogen peroxide, and the evolved isolates exhibiting increased carotenoid production (SM11, SM12, SM13, SM14, SM22, SM24, etc.) were selected based on their darker red color [12]. In subsequent work, we optimized bioreactor conditions for production using freshwater [14]. However, industrial fermentation requires a large amount of water, which generates pressures on freshwater [15]. Using alternative water sources such as unpurified seawater in industrial fermentation will help to reduce freshwater usage in bio-based production of chemicals [16], especially in arid regions of the world that have easy access to saline water sources. Prior work has shown the use of seawater media to be promising for ethanol fermentation by Saccharomyces cerevisiae [17], and seawater media has also been used to improve carotenoid production in Rhodotorula glutinis [18]. Therefore, seawater may also have the potential to improve carotenoid production in engineered Saccharomyces cerevisiae. In this work, we analyzed the impact of using seawater and reduced nutrients on the carotenoid productivity of a hyper-producer (SM14) generated from our prior work [12], and showed that the carotenoid production of the engineered non-native producer can also be improved by using seawater media.
2. Results and Discussion
2.1. Impact of YNB Concentration on β-Carotene Production of SM14
Seawater contains trace elements (Na+, Mg2+, K+, Ca2+, Cl−1) and SO42− that are present in yeast nitrogen base (YNB) (see Table S1 for comparison). Thus, we hypothesized that using seawater may reduce the requirement of YNB, and compared the β-carotene production of SM14 with different concentrations of YNB (1×, 0.7×, 0.4×, 0.1×) (while keeping the concentration of other constituents the same as described in Materials and Methods) in freshwater and seawater. Results showed that in 1×, 0.7×, or 0.4× YNB, biomass formation and β-carotene concentrations were both lower in seawater than in freshwater, but β-carotene yields were not significantly different between freshwater and seawater (Figure S1). Interestingly, in 0.1× YNB, the biomass formation, β-carotene concentration, and yield in seawater were significantly higher than that in freshwater (p-value < 0.01). Furthermore, the β-carotene yield in seawater in 0.1× YNB was significantly higher than in higher concentrations of YNB (p-value < 0.01), which suggests that SM14 produced and accumulated more carotenoids in seawater under nutrient-reduced conditions. Since the difference in β-carotene production between freshwater and seawater was most obvious with lower YNB concentration, subsequent experiments were conducted with 0.1× YNB.
2.2. Impact of Water Sources on β-Carotene Production of SM14
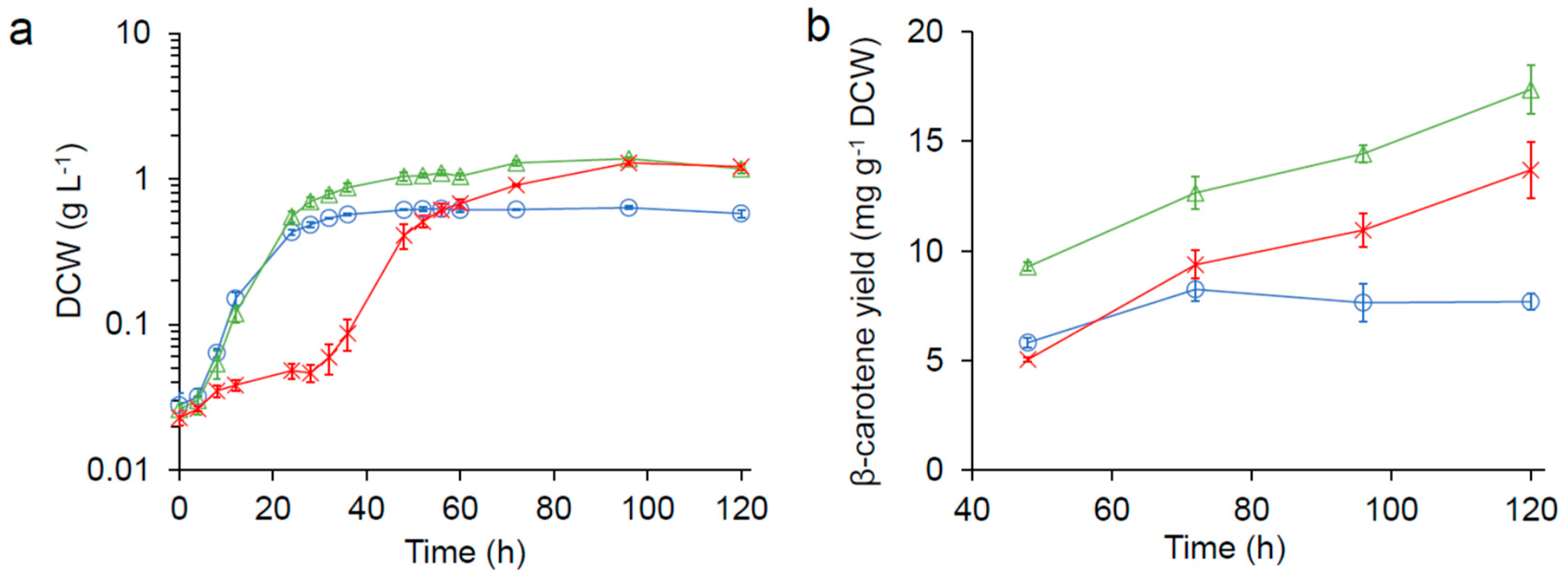
For nutrient-reduced condition, we first tested the impact of water source on the carotenoid production of SM14 in 20 mL media using either freshwater, 1/3× seawater (water: synthetic seawater = 2:1), or seawater in 125 mL flasks. As show in Figure 1a, the growth of SM14 was significantly inhibited in seawater, with a 24 h longer lag phase; however, the culture reached a higher final cell density compare with that in freshwater. Interestingly, no growth inhibition and a higher final cell density were observed in 1/3× seawater.
The β-carotene production of strain SM14 in both seawater and 1/3× seawater were significantly higher compared to that in freshwater (Figure 1b). The β-carotene yield observed in freshwater peaked at 8.25 ± 0.51 mg g−1 DCW (DCW: dry cell weight) after 72 h. On the other hand, the β-carotene production in seawater and 1/3× seawater continued to increase and reached 13.70 ± 1.29 mg g−1 DCW and 17.37 ± 1.12 mg g−1 DCW, respectively, after 120 h.
2.3. Impact of NaCl on β-Carotene Production of SM14
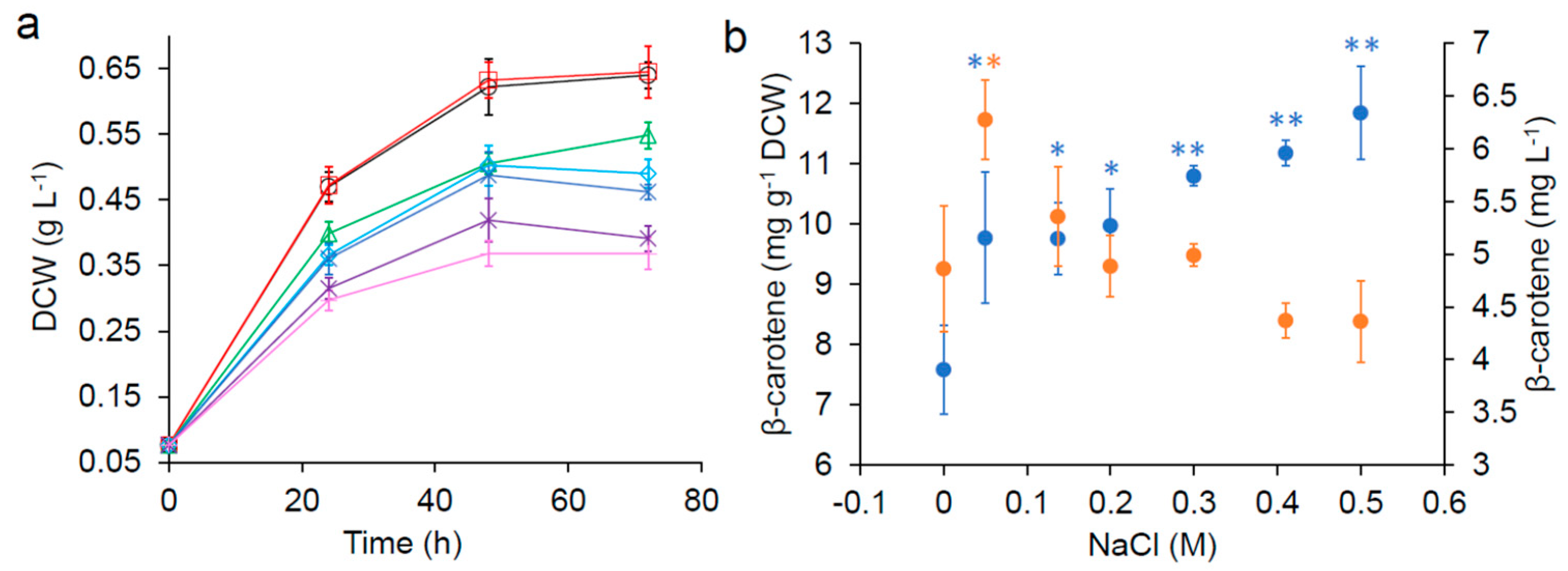
It has been reported that the addition of NaCl increased carotenoid production in some native producers [19,20,21,22,23]. Sodium chloride is the main component in seawater (the synthetic seawater contains ~0.41 M NaCl), thus the impact of excess NaCl on cell growth and β-carotene production was investigated in freshwater supplemented with varying concentrations of additional NaCl (Figure 2). All concentrations of NaCl tested significantly improved β-carotene production of SM14 in the nutrient-reduced media after 72 h (Figure 2b). However, the growth rate and final biomass decreased as the concentration of NaCl increased (Figure 2a), suggesting that while NaCl had a detrimental impact on cell growth, it had a positive impact on β-carotene production.
As the carotenoid hyperproducers were selected in the presence of hydrogen peroxide, it was not surprising that genes regulated by the major regulators involved in oxidative stress response in yeast (i.e., Yap1 [24,25,26], Skn7 [26], Mga2 [27], and Msn2/4 [28]) were differentially expressed [12]. Prior studies have shown that exposure to osmotic stress confers tolerance to oxidative stress in yeast [29]; suggesting that the same pathways involved in osmotic and oxidative stress resistance overlap. Indeed, the regulators Msn2, Msn4, Pdr3, and Hsf1 have been found to be common between oxidative and osmotic stress [30]. However, it is unclear whether increased oxidative stress tolerance enhances carotenoid production. Thus, it is not known at this point whether the commonalities between the osmotic and oxidative stress tolerance regulon are directly connected with the carotenoid hyper-producing phenotype in SM14.
2.4. Impact of pH on β-Carotene Production of SM14
The pH of the nutrient-reduced media differed depending on whether seawater or freshwater is used, with values of ~5.2, 6.5, and 7.0 for 0.1× YNB media in freshwater, 1/3× seawater, and seawater, respectively. Prior work has shown that differences in pH have potential impacts on carotenoid production on microbial producers [14,22]. Thus, β-carotene production in freshwater with varying pHs (~5.2, 6.5, and 7.0) was quantified. The results showed that the initial pH of the media did not appear to have any significant impact on β-carotene production (Figure S2a) or growth (Figure S2b), and the pH decreased to ~4.0 in all conditions before reaching a stationary phase (Figure S2b) when the carotenoids started to accumulate in cells.
2.5. Impact of Carbon-to-Nitrogen Ratio (C:N) on β-Carotene Production of SM14
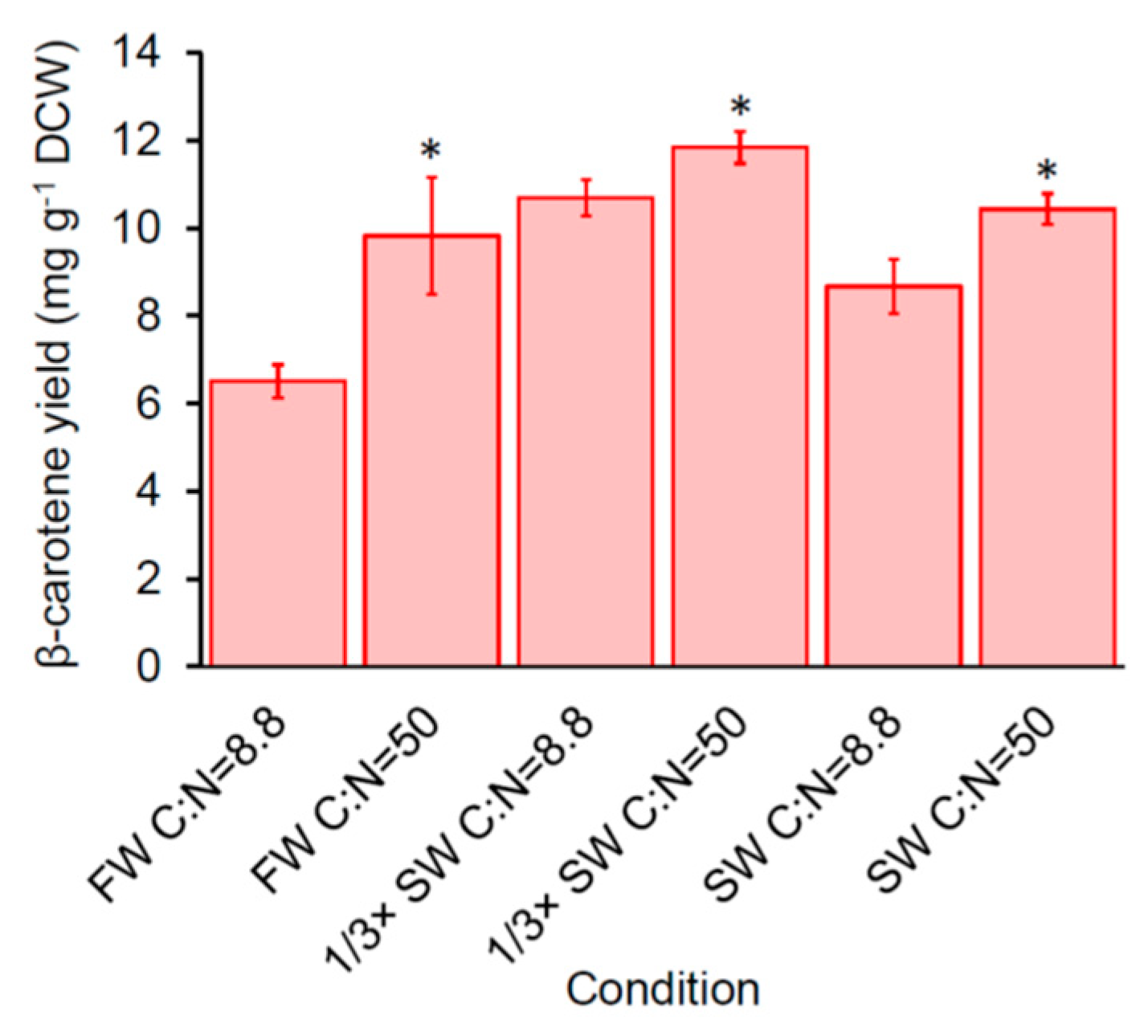
We previously demonstrated that increasing carbon-to-nitrogen (C:N) ratio from 8.8 to 50 improved carotenoid production of engineered yeast (including SM14) in benchtop bioreactors [14]. To determine if increasing the C:N ratio also increases carotenoid production in nutrient-reduced conditions, we compared the β-carotene production of SM14 in 0.1× YNB media with the normal C:N ratio (8.8) (use 5 g ammonium sulfate L−1 in the media as described in Materials and Methods) and high C:N ratio (50) (use 0.88 g ammonium sulfate L−1 in the media while keeping the concentration of other constituents the same as described in Materials and Methods) in freshwater, 1/3× seawater, and seawater (Figure 3). The results showed that C:N = 50 significantly increased β-carotene production of SM14 comparing with C:N = 8.8, from 6.51 ± 0.37 mg g−1 DCW to 9.83 ± 1.34 mg g−1 DCW in freshwater, from 10.70 ± 0.41 mg g−1 DCW to 11.84 ± 0.36 mg g−1 DCW in 1/3× seawater, and from 8.67 ± 0.62 mg g−1 DCW to 10.44 ± 0.35 mg g−1 DCW in seawater after 72 h, which indicates that increasing the C:N ratio can further improve carotenoid production of SM14 in seawater.
2.6. Lipid Production and Composition
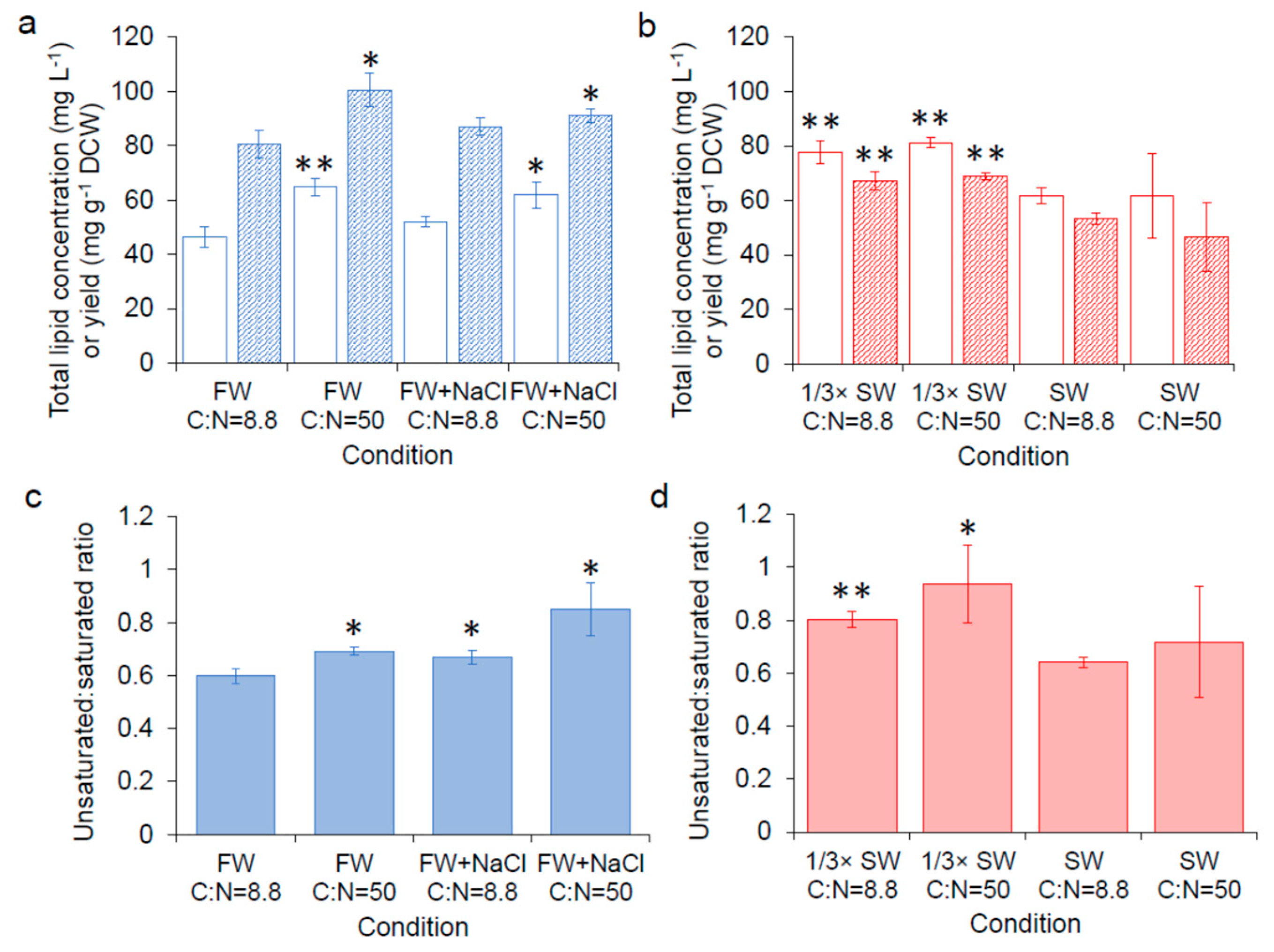
Based on transcriptome results comparing the global gene expression of carotenoid hyper-producers to the parental strains from our previous work, we found that genes related with lipid biosynthesis (e.g., CYB5, ERG27, INO1) were differently expressed in the hyper-producers [12]; the direction of gene expression changes pointed to the possibility that the hyper-producers may have a higher lipid content. Thus, in our follow-up work, we applied prior knowledge from oleaginous yeasts that showed increased lipid content when cells are grown in nitrogen-limited conditions [31] to our strains, and found an increased carotenoid production when the C:N ratio was increased from 8.8 to 50 [14]. Using Raman spectroscopy, we confirmed that the improved carotenoid production by increasing the C:N ratio to 50 was correlated with increased total lipid [14]. In this work, lipid analysis was carried out with SM14 cultured in different conditions to identify potential impacts of seawater and varying C:N ratio on lipid content and composition of the cell. Samples in freshwater media were collected from cultures after 72 h and samples in seawater media were collected after 120 h when β-carotene production reached the highest level (based on Figure 1). As show in Figure 4a,c and Figure S3a, supplementing freshwater with 0.05 M NaCl or increasing the C:N ratio from 8.8 to 50 in freshwater significantly increased lipid concentration, yield or ratio of unsaturated to saturated fatty acids compared with C:N = 8.8 in freshwater. This result suggests that the higher carotenoid production in these conditions may be related with the higher production of total lipids or the increased unsaturation of fatty acids observed with the addition of NaCl or increasing the C:N ratio. In contrast, increasing the C:N ratio in seawater did not impact lipid production or fatty acid saturation (Figure 4b,d, and Figure S3b). Corresponding with β-carotene production (Figure 3), the total lipid yield and concentration are also higher in 1/3× seawater than in seawater, which provides further support that β-carotene production is related with lipid content. It has been reported that salt stress affects fatty acid composition in yeast, and an increase in total sterol and squalene biosynthesis was observed in S. cerevisiae under osmotic stress [32]. Squalene is produced via the mevalonate (MVA) pathway in yeast, and the precursor, farnesyl diphosphate (FPP) is also the precursor for carotenoid biosynthesis [10]. Therefore, an upregulation in the MVA pathway in response to osmotic stress may contribute to the increased production of carotenoids in seawater and in freshwater supplemented with additional NaCl.
2.7. The Impact of Seawater on β-Carotene Production of Other Evolved Hyper-Producers
Due to potential differences in regulation and metabolism, the same factors may not have the same impact in different strain backgrounds. Indeed, some reports showed that addition of moderate concentrations of NaCl increased carotenoid production [19,20,21,22,23], while others showed decreased production [2]. Thus, whether the observed positive impact of seawater on carotenoid production in strain SM14 also applies to other evolved hyper-producers and the parental strain (YLH2) was investigated. The carotenoid production of YLH2 and some evolved hyper-producers (SM11, SM12, SM13, SM22, SM24) were quantified in nutrient-reduced condition in either freshwater or 1/3× seawater. As show in Figure S4, consistent with the observation with SM14, the use of 1/3× seawater improved β-carotene production of all tested evolved hyper-producers as well as the ancestor strain YLH2 compared with that in freshwater.
3. Conclusions
In conclusion, this work demonstrated that using seawater improved carotenoid production in engineered carotenoids producing S. cerevisiae under nutrient-reduced conditions. This improvement is partially due to the slight osmotic stress imposed by the NaCl content in seawater. Based on our prior transcriptomic results, carotenoid hyper-producers exhibited differential expression in lipid biosynthesis related genes [12]; suggesting a positive correlation between lipid content and β-carotene production. This phenomenon was also observed in this work, as a positive correlation between increased β-carotene production while an increase in total lipid content and unsaturation were observed. Increasing the C:N ratio further improved carotenoid production in both freshwater and seawater, and also was shown to be correlated with lipid content and composition. In comparing the highest carotene production found in this work with previously reported data (see Table S3), the carotene yield we obtained in this work was higher than that reported in the native producer Rhodotorula glutinis [18]. Though the highest carotene production in this work is still lower than our previous work using rich media (YPD) [12] or normal YNB media [14], we showed that using seawater is another potential way to improve carotenoid production, especially under nutrient limited conditions. These findings have the potential to expand the utilization of seawater in microbial fermentation and improve the production of carotenoids.
4. Materials and Methods
4.1. Strains and Growth Conditions
Saccharomyces cerevisiae strains SM14, SM11, SM12, SM13, SM22, SM24, and YLH2 [12] were used in this study (Table S2). YLH2 is a S288c strain engineered to produce β-carotene and has the catalase gene ctt1 deleted; it is the parental strain for SM11, SM12, SM13, SM14, SM22, and SM24, which were evolved under oxidative stress to improve β-carotene production [12]. Unless otherwise noted, stock cultures were kept at −80 °C, strains were streaked out on YNB plates from frozen stocks and single colonies were picked and cultured aerobically in test tubes containing 3 mL yeast nitrogen base (YNB, 1.7 g L−1, AMRESCO) or 0.1× yeast nitrogen base (0.1× YNB, 0.17 g L−1) media supplemented with 5 g ammonium sulfate L−1 and 20 g glucose L−1 made with either freshwater or synthetic seawater (RICCA) at 30 °C. Inoculum of all experiments was collected from overnight cultures in YNB and washed twice with sterile water before inoculation to avoid the impacts of remaining nutrients in the inoculum. The initial OD600 in all experiments was normalized to about 0.1. All experiments were performed in triplicates and statistical tests for significance were determined via 2-tailed Student’s t-test.
4.2. Carotenoid Quantification
β-Carotene was quantified by measuring absorbance at 454 nm with a microplate reader (TECAN Infinites M200) [12,14] or by using high-performance liquid chromatography (HPLC, Agilent Technologies, 1260 Infinity, Santa Clara, CA, USA) and Agilent ZOBAX Eclipse Plus C18 column (4.6 × 100 mm, 3.5-Micron) as previously described [33]. Briefly, acetonitrile–methanol–isopropanol (50:30:20 v/v) was used as mobile phase at a flow rate of 1 mL min−1, the column temperature was set at 40 °C, and β-carotene was detected by UV detector (VWD signal) with wavelength of 450 nm. The quantification by HPLC and absorbance were compared and were found to correlate well (Table S4). The simpler spectrophotometric assay was used for carotenoid quantification in this work.
4.3. Lipid Quantification
The total lipids of S. cerevisiae were extracted in the form of fatty acid methyl ester (FAME) using the sulfuric acid–methanol method as previously described [34], details can be found in Supplementary method. The lipid composition was analyzed by GC–MS using a SHIMADZU-QP2010 SE GC-MS (SHIMADZU CORPORATION, Japan) equipped with a ZB-5MSi column (thickness 0.25 μm; length 30 m; diameter 0.25 mm) as previously described [34].
Supplementary Materials
The following are available online at https://www.mdpi.com/2311-5637/5/1/6/s1, Supplementary Method: Lipid extraction; Table S1: Concentration of common components in normal YNB and synthetic seawater; Table S2: List of strains used; Table S3: Highest β-carotene production in different conditions; Table S4: Comparison of β-carotene measurement between spectrophotometric assay and HPLC; Figure S1: Effect of YNB concentration on biomass formation and β-carotene production of SM14 in freshwater ( ![Fermentation 05 00006 i012]() ) and seawater (
) and seawater ( ![Fermentation 05 00006 i015]() ) after 72 h incubation. (a) Biomass (DCW: dry cell weight). (b) β-carotene concentration. (c) β-carotene yield. Error bars are standard deviations; Figure S2: β-carotene production after 120 h (a), cell growth (solid lines) (DCW: dry cell weight) and pH change (dashed lines) (b) of SM14 in freshwater with different initial pH, (
) after 72 h incubation. (a) Biomass (DCW: dry cell weight). (b) β-carotene concentration. (c) β-carotene yield. Error bars are standard deviations; Figure S2: β-carotene production after 120 h (a), cell growth (solid lines) (DCW: dry cell weight) and pH change (dashed lines) (b) of SM14 in freshwater with different initial pH, ( ![Fermentation 05 00006 i001]() ) initial pH 5.2, (
) initial pH 5.2, ( ![Fermentation 05 00006 i005]() ) initial pH 6.5, (
) initial pH 6.5, ( ![Fermentation 05 00006 i002]() ) initial pH 6.9. Error bars are standard deviations; Figure S3 Individual fatty acid yield. (a) (
) initial pH 6.9. Error bars are standard deviations; Figure S3 Individual fatty acid yield. (a) ( ![Fermentation 05 00006 i012]() ) C:N = 8.8 in freshwater, (
) C:N = 8.8 in freshwater, ( ![Fermentation 05 00006 i013]() ) C:N = 50 in freshwater, (
) C:N = 50 in freshwater, ( ![Fermentation 05 00006 i014]() ) C:N = 8.8 in freshwater with 0.05 M NaCl, (
) C:N = 8.8 in freshwater with 0.05 M NaCl, ( ![Fermentation 05 00006 i015]() ) C:N = 50 in freshwater with 0.05 M NaCl; (b) (
) C:N = 50 in freshwater with 0.05 M NaCl; (b) ( ![Fermentation 05 00006 i012]() ) C:N = 8.8 in 1/3× seawater, (
) C:N = 8.8 in 1/3× seawater, ( ![Fermentation 05 00006 i013]() ) C:N = 50 in 1/3× seawater, (
) C:N = 50 in 1/3× seawater, ( ![Fermentation 05 00006 i014]() ) C:N = 8.8 in seawater, (
) C:N = 8.8 in seawater, ( ![Fermentation 05 00006 i015]() ) C:N = 50 in seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a) or seawater (b) (* p-value <0.05, ** p-value <0.01). MT: Methyl tetradecanoate; 9-HA: 9-Hexadecenoic acid; HA: Hexadecanoic acid; 11-OA: 11-Octadecenoic acid; MS: Methyl stearate; Figure S4: β-carotene production of carotenogenic strains in 0.1× YNB with freshwater (
) C:N = 50 in seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a) or seawater (b) (* p-value <0.05, ** p-value <0.01). MT: Methyl tetradecanoate; 9-HA: 9-Hexadecenoic acid; HA: Hexadecanoic acid; 11-OA: 11-Octadecenoic acid; MS: Methyl stearate; Figure S4: β-carotene production of carotenogenic strains in 0.1× YNB with freshwater ( ![Fermentation 05 00006 i001]() ) or 1/3× seawater (
) or 1/3× seawater ( ![Fermentation 05 00006 i005]() ). (a) SM11. (b) SM12. (c) SM13. (d) SM22. (e) SM24. (f) YLH2. Error bars are standard deviations; Supplementary data: Fatty acid composition
). (a) SM11. (b) SM12. (c) SM13. (d) SM22. (e) SM24. (f) YLH2. Error bars are standard deviations; Supplementary data: Fatty acid composition
 ) and seawater (
) and seawater (  ) after 72 h incubation. (a) Biomass (DCW: dry cell weight). (b) β-carotene concentration. (c) β-carotene yield. Error bars are standard deviations; Figure S2: β-carotene production after 120 h (a), cell growth (solid lines) (DCW: dry cell weight) and pH change (dashed lines) (b) of SM14 in freshwater with different initial pH, (
) after 72 h incubation. (a) Biomass (DCW: dry cell weight). (b) β-carotene concentration. (c) β-carotene yield. Error bars are standard deviations; Figure S2: β-carotene production after 120 h (a), cell growth (solid lines) (DCW: dry cell weight) and pH change (dashed lines) (b) of SM14 in freshwater with different initial pH, (  ) initial pH 5.2, (
) initial pH 5.2, (  ) initial pH 6.5, (
) initial pH 6.5, (  ) initial pH 6.9. Error bars are standard deviations; Figure S3 Individual fatty acid yield. (a) ( ) C:N = 8.8 in freshwater, (
) initial pH 6.9. Error bars are standard deviations; Figure S3 Individual fatty acid yield. (a) ( ) C:N = 8.8 in freshwater, (  ) C:N = 50 in freshwater, (
) C:N = 50 in freshwater, (  ) C:N = 8.8 in freshwater with 0.05 M NaCl, ( ) C:N = 50 in freshwater with 0.05 M NaCl; (b) ( ) C:N = 8.8 in 1/3× seawater, ( ) C:N = 50 in 1/3× seawater, ( ) C:N = 8.8 in seawater, ( ) C:N = 50 in seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a) or seawater (b) (* p-value <0.05, ** p-value <0.01). MT: Methyl tetradecanoate; 9-HA: 9-Hexadecenoic acid; HA: Hexadecanoic acid; 11-OA: 11-Octadecenoic acid; MS: Methyl stearate; Figure S4: β-carotene production of carotenogenic strains in 0.1× YNB with freshwater ( ) or 1/3× seawater ( ). (a) SM11. (b) SM12. (c) SM13. (d) SM22. (e) SM24. (f) YLH2. Error bars are standard deviations; Supplementary data: Fatty acid composition
) C:N = 8.8 in freshwater with 0.05 M NaCl, ( ) C:N = 50 in freshwater with 0.05 M NaCl; (b) ( ) C:N = 8.8 in 1/3× seawater, ( ) C:N = 50 in 1/3× seawater, ( ) C:N = 8.8 in seawater, ( ) C:N = 50 in seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a) or seawater (b) (* p-value <0.05, ** p-value <0.01). MT: Methyl tetradecanoate; 9-HA: 9-Hexadecenoic acid; HA: Hexadecanoic acid; 11-OA: 11-Octadecenoic acid; MS: Methyl stearate; Figure S4: β-carotene production of carotenogenic strains in 0.1× YNB with freshwater ( ) or 1/3× seawater ( ). (a) SM11. (b) SM12. (c) SM13. (d) SM22. (e) SM24. (f) YLH2. Error bars are standard deviations; Supplementary data: Fatty acid compositionAuthor Contributions
Y.G. performed the experiments, analyzed the data and drafted the manuscript. S.X. quantified lipid composition. K.C.K., J.S.Y., and S.X. revised the manuscript. All authors read and approved the final manuscript.
Funding
The work was partially supported by the National Science Foundation (MCB-1054276) and Texas A&M Engineering Experiment Station (TEES).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Fong, N.J.; Burgess, M.L.; Barrow, K.D.; Glenn, D.R. Carotenoid accumulation in the psychrotrophic bacterium Arthrobacter agilis in response to thermal and salt stress. Appl. Microbiol. Biotechnol. 2001, 56, 750–756. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Distributions, biosyntheses and functions of carotenoids in algae. Agro Food Ind. Hi Tech 2013, 24, 55–58. [Google Scholar]
- Avalos, J.; Limon, M.C. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Kirti, K.; Amita, S.; Priti, S.; Mukesh Kumar, A.; Jyoti, S. Colorful world of microbes: Carotenoids and their applications. Adv. Biol. 2014, 2014, 13. [Google Scholar] [CrossRef]
- Nigam, P.S.; Luke, J.S. Food additives: Production of microbial pigments and their antioxidant properties. Curr. Opin. Food Sci. 2016, 7, 93–100. [Google Scholar] [CrossRef]
- Manimala, M.; Murugesan, R. In vitro antioxidant and antimicrobial activity of carotenoid pigment extracted from Sporobolomyces sp. Isolated from natural source. J. Appl. Nat. Sci. 2014, 6, 649–653. [Google Scholar] [CrossRef]
- Anunciato, T.P.; da Rocha, P.A. Carotenoids and polyphenols in nutricosmetics, nutraceuticals, and cosmeceuticals. J. Cosmet. Dermatol. 2012, 11, 51–54. [Google Scholar] [CrossRef]
- Vachali, P.; Bhosale, P.; Bernstein, P.S. Microbial carotenoids. In Microbial Carotenoids from Fungi: Methods and Protocols; Barredo, J.-L., Ed.; Humana Press: Totowa, NJ, USA, 2012; pp. 41–59. [Google Scholar]
- Verwaal, R.; Wang, J.; Meijnen, J.P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef]
- Zhao, J.; Li, Q.; Sun, T.; Zhu, X.; Xu, H.; Tang, J.; Zhang, X.; Ma, Y. Engineering central metabolic modules of Escherichia coli for improving beta-carotene production. Metab. Eng. 2013, 17, 42–50. [Google Scholar] [CrossRef]
- Reyes, L.H.; Gomez, J.M.; Kao, K.C. Improving carotenoids production in yeast via adaptive laboratory evolution. Metab. Eng. 2014, 21, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Kao, K.C.; Sherlock, G. Molecular characterization of clonal interference during adaptive evolution in asexual populations of Saccharomyces cerevisiae. Nat. Genet. 2008, 40, 1499–1504. [Google Scholar] [CrossRef] [PubMed]
- Olson, M.L.; Johnson, J.; Carswell, W.F.; Reyes, L.H.; Senger, R.S.; Kao, K.C. Characterization of an evolved carotenoids hyper-producer of Saccharomyces cerevisiae through bioreactor parameter optimization and raman spectroscopy. J. Ind. Microbiol. Biotechnol. 2016, 43, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Gerbens-Leenes, P.W.; Lienden, A.R.V.; Hoekstra, A.Y.; van der Meer, T.H. Biofuel scenarios in a water perspective: The global blue and green water footprint of road transport in 2030. Glob. Environ. Chang. 2012, 22, 764–775. [Google Scholar] [CrossRef] [Green Version]
- Zaky, A.S. Marine fermentation, the sustainable approach for bioethanol production. EC Microbiol. 2017, ECO.01, 25–27. [Google Scholar]
- Goncalves, F.A.; dos Santos, E.S.; de Macedo, G.R. Alcoholic fermentation of Saccharomyces cerevisiae, Pichia stipitis and Zymomonas mobilis in the presence of inhibitory compounds and seawater. J. Basic Microbiol. 2015, 55, 695–708. [Google Scholar] [CrossRef] [PubMed]
- Bhosale, P.; Gadre, R.V. Production of beta-carotene by a Rhodotorula glutinis mutant in sea water medium. Bioresour. Technol. 2001, 76, 53–55. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Breierova, E.; Koci, R. Production of carotenoid-/ergosterol-supplemented biomass by red yeast Rhodotorula glutinis grown under external stress. Food Technol. Biotechnol. 2010, 48, 56–61. [Google Scholar]
- Farhat, N.; Rabhi, M.; Falleh, H.; Jouini, J.; Abdelly, C.; Smaoui, A. Optimization of salt concentrations for a higher carotenoid production in Dunaliella salina (chlorophyceae). J. Phycol. 2011, 47, 1072–1077. [Google Scholar] [CrossRef]
- Marova, I.; Carnecka, M.; Halienova, A.; Certik, M.; Dvorakova, T.; Haronikova, A. Use of several waste substrates for carotenoid-rich yeast biomass production. J. Environ. Manag. 2012, 95, S338–S342. [Google Scholar] [CrossRef]
- Kanzy, H.M.; Nasr, N.; El-Shazly, H.A.; Barakat, O.S. Optimization of carotenoids production by yeast strains of Rhodotorula using salted cheese whey. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 456–469. [Google Scholar]
- Ben Abdallah, S.; Aung, B.; Amyot, L.; Lalin, I.; Lachaal, M.; Karray-Bouraoui, N.; Hannoufa, A. Salt stress (NaCl) affects plant growth and branch pathways of carotenoid and flavonoid biosyntheses in Solanum nigrum. Acta Physiol. Plant. 2016, 38, 72. [Google Scholar] [CrossRef]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The response to heat shock and oxidative stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef]
- Kuge, S.; Jones, N. YAP1 dependent activation of TRX2 is essential for the response of Saccharomyces cerevisiae to oxidative stress by hydroperoxides. Embo J. 1994, 13, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Godon, C.; Lagniel, G.; Spector, D.; Garin, J.; Labarre, J.; Toledano, M.B. Yap1 and skn7 control two specialized oxidative stress response regulons in yeast. J. Biol. Chem. 1999, 274, 16040–16046. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.; Ideker, T. Genome-wide fitness and expression profiling implicate Mga2 in adaptation to hydrogen peroxide. PLoS Genet. 2009, 5, e1000488. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.; Leroy, C.; Isnard, A.D.; Labarre, J.; Boy-Marcotte, E.; Toledano, M.B. The control of the yeast H2O2 response by the Msn2/4 transcription factors. Mol. Microbiol. 2002, 45, 233–241. [Google Scholar] [CrossRef]
- Berry, D.B.; Gasch, A.P. Stress-activated genomic expression changes serve a preparative role for impending stress in yeast. Mol. Biol. Cell 2008, 19, 4580–4587. [Google Scholar] [CrossRef]
- Święciło, A. Cross-stress resistance in Saccharomyces cerevisiae yeast—New insight into an old phenomenon. Cell Stress Chaperones 2016, 21, 1–14. [Google Scholar] [CrossRef]
- Sitepu, I.R.; Sestric, R.; Ignatia, L.; Levin, D.; German, J.B.; Gillies, L.A.; Almada, L.A.; Boundy-Mills, K.L. Manipulation of culture conditions alters lipid content and fatty acid profiles of a wide variety of known and new oleaginous yeast species. Bioresour. Technol. 2013, 144, 360–369. [Google Scholar] [CrossRef] [Green Version]
- Turk, M.; Mejanelle, L.; Sentjurc, M.; Grimalt, J.O.; Gunde-Cimerman, N.; Plemenitas, A. Salt-induced changes in lipid composition and membrane fluidity of halophilic yeast-like melanized fungi. Extremophiles 2004, 8, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Lv, X.; Ye, L.; Zhou, P.; Yu, H. Construction of lycopene-overproducing Saccharomyces cerevisiae by combining directed evolution and metabolic engineering. Metab. Eng. 2015, 30, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Qin, X.; Cheng, Y.; Laskar, D.; Qiao, W.; Sun, S.; Reyes, L.H.; Wang, X.; Dai, S.Y.; Sattler, S.E.; et al. Simultaneous conversion of all cell wall components by an oleaginous fungus without chemi-physical pretreatment. Green Chem. 2015, 17, 1657–1667. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Growth kinetics of SM14 in 0.1× yeast nitrogen base (YNB) media (20 mL) made with freshwater ( ![Fermentation 05 00006 i001]() ), 1/3× seawater (
), 1/3× seawater ( ![Fermentation 05 00006 i002]() ), and seawater (
), and seawater ( ![Fermentation 05 00006 i003]() ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
), 1/3× seawater ( ), and seawater (  ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
Figure 1.
Growth kinetics of SM14 in 0.1× yeast nitrogen base (YNB) media (20 mL) made with freshwater ( ![Fermentation 05 00006 i001]() ), 1/3× seawater (
), 1/3× seawater ( ![Fermentation 05 00006 i002]() ), and seawater (
), and seawater ( ![Fermentation 05 00006 i003]() ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.
), 1/3× seawater ( ), and seawater ( ) in 125 mL flasks. (a) growth kinetics (DCW: dry cell weight). (b) β-carotene production.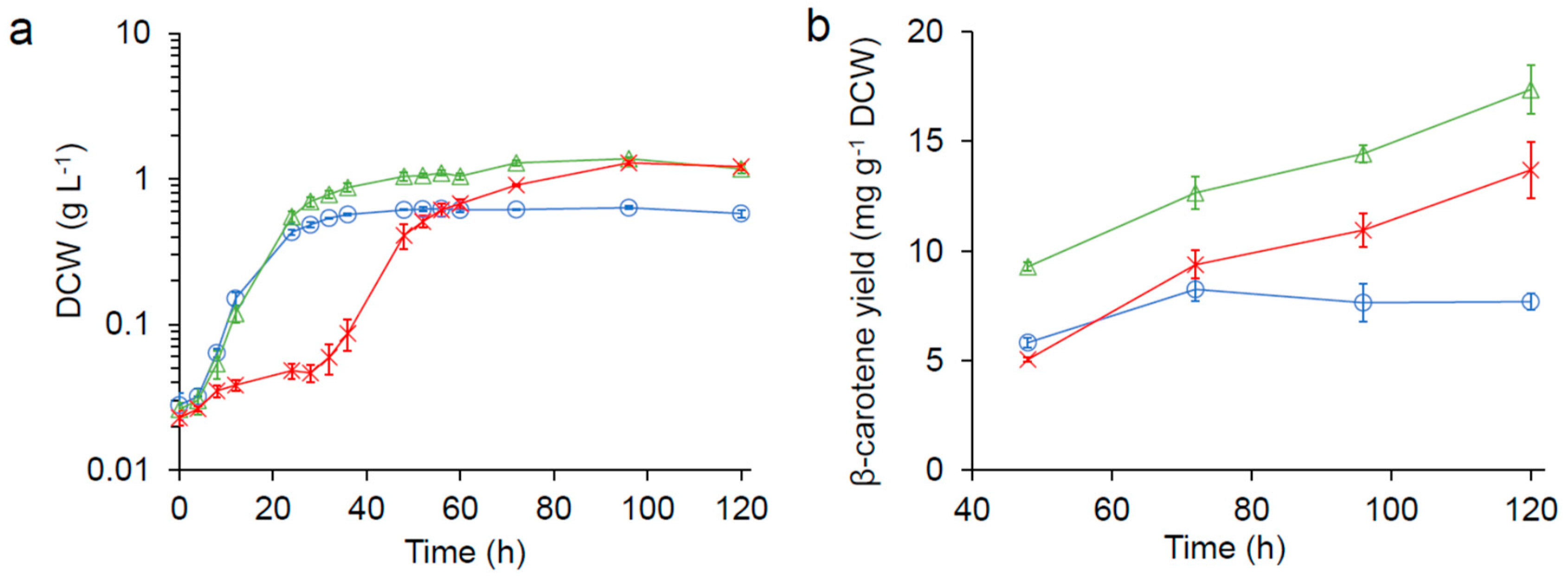
Figure 2.
Effect of NaCl supplementation on the growth kinetics and β-carotene production of SM14 in 0.1× YNB media in freshwater. (a) growth kinetics (DCW: dry cell weight) with 0 M ( ![Fermentation 05 00006 i004]() ), 0.05 M (
), 0.05 M ( ![Fermentation 05 00006 i005]() ), 0.137 M (
), 0.137 M ( ![Fermentation 05 00006 i002]() ), 0.2 M (
), 0.2 M ( ![Fermentation 05 00006 i006]() ), 0.3 M (
), 0.3 M ( ![Fermentation 05 00006 i007]() ), 0.41 M (
), 0.41 M ( ![Fermentation 05 00006 i008]() ) and 0.5 M (
) and 0.5 M ( ![Fermentation 05 00006 i009]() ) NaCl. (b) β-carotene yield (
) NaCl. (b) β-carotene yield ( ![Fermentation 05 00006 i010]() ) and concentration (
) and concentration ( ![Fermentation 05 00006 i011]() ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
 ), 0.05 M ( ), 0.137 M ( ), 0.2 M (
), 0.05 M ( ), 0.137 M ( ), 0.2 M (  ), 0.3 M (
), 0.3 M (  ), 0.41 M (
), 0.41 M (  ) and 0.5 M (
) and 0.5 M (  ) NaCl. (b) β-carotene yield (
) NaCl. (b) β-carotene yield (  ) and concentration (
) and concentration (  ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
Figure 2.
Effect of NaCl supplementation on the growth kinetics and β-carotene production of SM14 in 0.1× YNB media in freshwater. (a) growth kinetics (DCW: dry cell weight) with 0 M ( ![Fermentation 05 00006 i004]() ), 0.05 M (
), 0.05 M ( ![Fermentation 05 00006 i005]() ), 0.137 M (
), 0.137 M ( ![Fermentation 05 00006 i002]() ), 0.2 M (
), 0.2 M ( ![Fermentation 05 00006 i006]() ), 0.3 M (
), 0.3 M ( ![Fermentation 05 00006 i007]() ), 0.41 M (
), 0.41 M ( ![Fermentation 05 00006 i008]() ) and 0.5 M (
) and 0.5 M ( ![Fermentation 05 00006 i009]() ) NaCl. (b) β-carotene yield (
) NaCl. (b) β-carotene yield ( ![Fermentation 05 00006 i010]() ) and concentration (
) and concentration ( ![Fermentation 05 00006 i011]() ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).
), 0.05 M ( ), 0.137 M ( ), 0.2 M ( ), 0.3 M ( ), 0.41 M ( ) and 0.5 M ( ) NaCl. (b) β-carotene yield ( ) and concentration ( ) after 72 h. Error bars are standard deviations. *: Statistically significantly different from the control (no NaCl) (* p-value < 0.05, ** p-value < 0.01).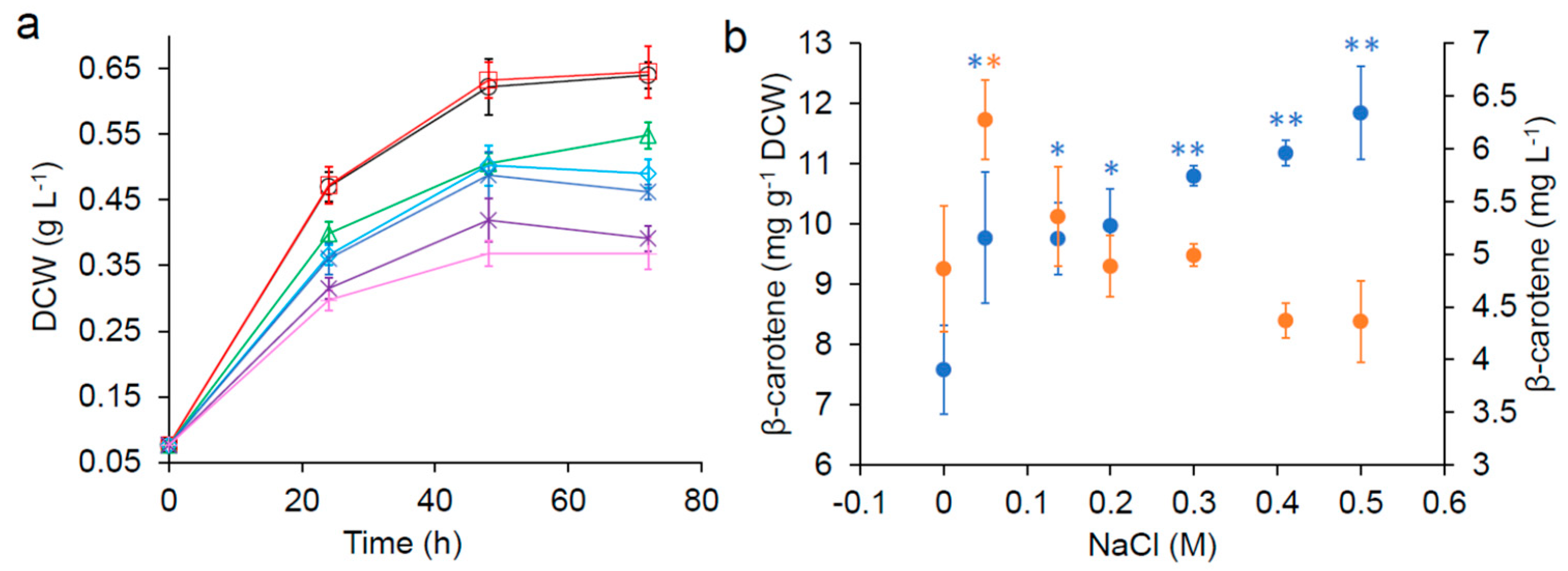
Figure 3.
Effect of carbon-to-nitrogen ratio (C:N) on β-carotene production of SM14 after 72 h. FW: fresh water; SW: seawater.. Error bars are standard deviations. *: Statistically significantly different between condition A and B of the same water sources (* p-value < 0.05).
Figure 3.
Effect of carbon-to-nitrogen ratio (C:N) on β-carotene production of SM14 after 72 h. FW: fresh water; SW: seawater.. Error bars are standard deviations. *: Statistically significantly different between condition A and B of the same water sources (* p-value < 0.05).
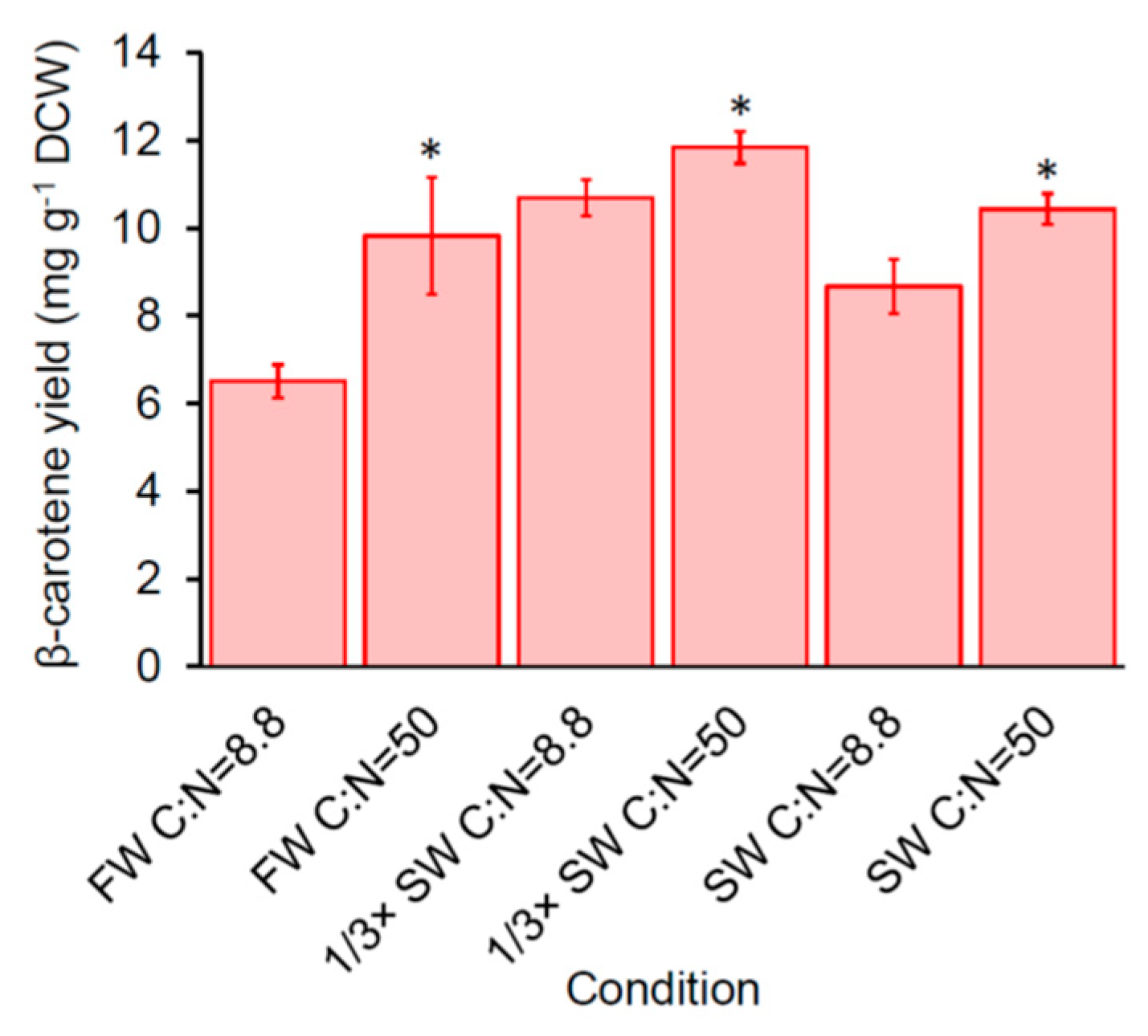
Figure 4.
Lipid content. (a) Total lipid concentration ( ![Fermentation 05 00006 i012]() ) and yield (
) and yield ( ![Fermentation 05 00006 i013]() ) of SM14 in freshwater, (b) total lipid concentration (
) of SM14 in freshwater, (b) total lipid concentration ( ![Fermentation 05 00006 i014]() ) and yield (
) and yield ( ![Fermentation 05 00006 i015]() ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) and yield ( ) of SM14 in freshwater, (b) total lipid concentration ( ) and yield ( ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
Figure 4.
Lipid content. (a) Total lipid concentration ( ![Fermentation 05 00006 i012]() ) and yield (
) and yield ( ![Fermentation 05 00006 i013]() ) of SM14 in freshwater, (b) total lipid concentration (
) of SM14 in freshwater, (b) total lipid concentration ( ![Fermentation 05 00006 i014]() ) and yield (
) and yield ( ![Fermentation 05 00006 i015]() ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).
) and yield ( ) of SM14 in freshwater, (b) total lipid concentration ( ) and yield ( ) of SM14 in seawater, (c) the ratio of unsaturated fatty acids to saturated fatty acids in freshwater, (d) the ratio of unsaturated fatty acids to saturated fatty acids in seawater. FW: fresh water; FW + NaCl: fresh water with 0.05 M NaCl; SW: seawater. Error bars are standard deviations. *: Statistically significantly different from C:N = 8.8 in freshwater (a,c) or seawater (b,d) (* p-value < 0.05, ** p-value < 0.01).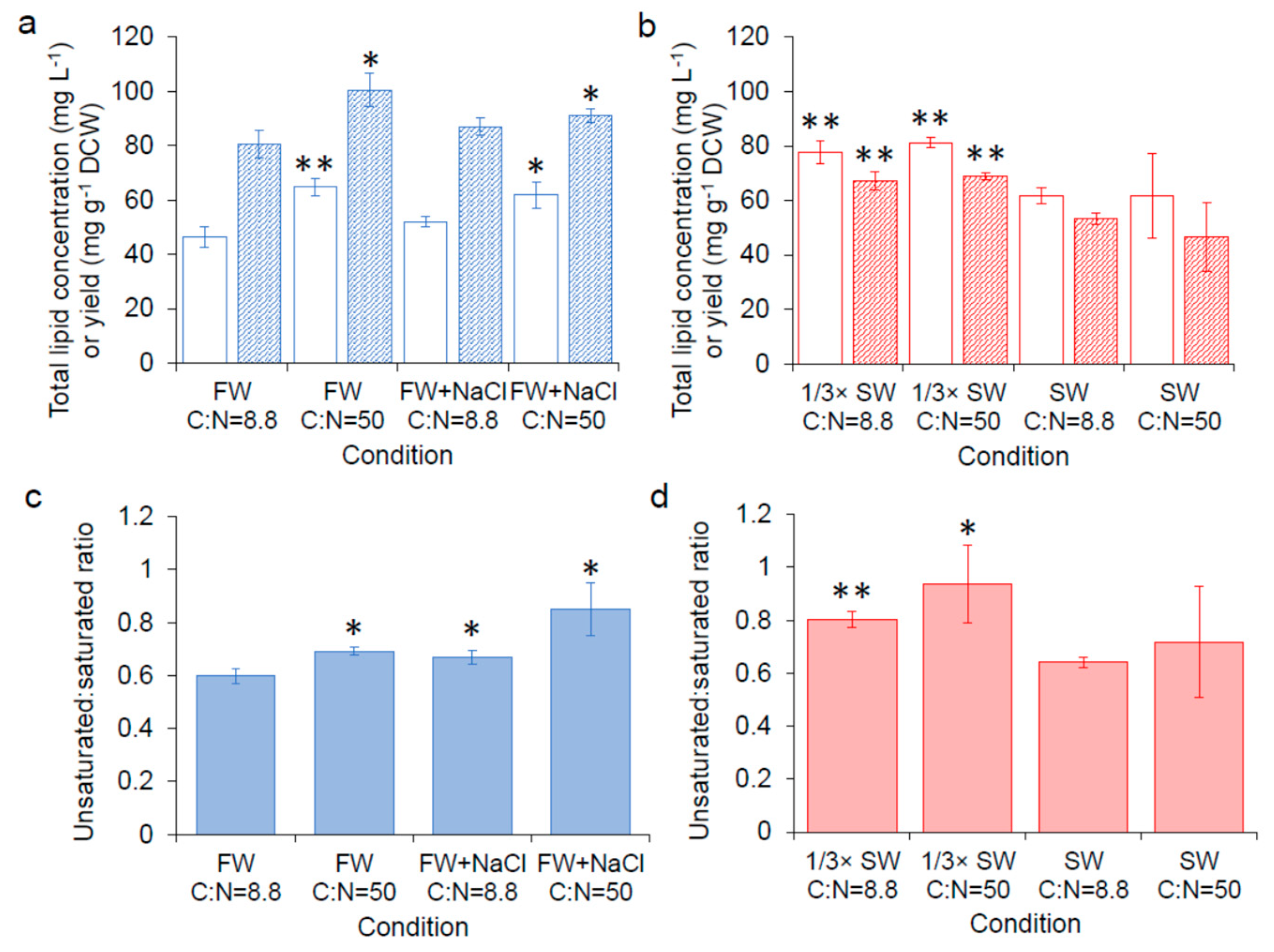
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Guo, Y.; Xie, S.; Yuan, J.S.; Kao, K.C. Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae. Fermentation 2019, 5, 6. https://doi.org/10.3390/fermentation5010006
AMA Style
Guo Y, Xie S, Yuan JS, Kao KC. Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae. Fermentation. 2019; 5(1):6. https://doi.org/10.3390/fermentation5010006
Chicago/Turabian StyleGuo, Yuqi, Shangxian Xie, Joshua S. Yuan, and Katy C. Kao. 2019. "Effects of Seawater on Carotenoid Production and Lipid Content of Engineered Saccharomyces cerevisiae" Fermentation 5, no. 1: 6. https://doi.org/10.3390/fermentation5010006
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.