The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor

Abstract

1. Introduction
2. The Role of Phenolic Compounds in Plants
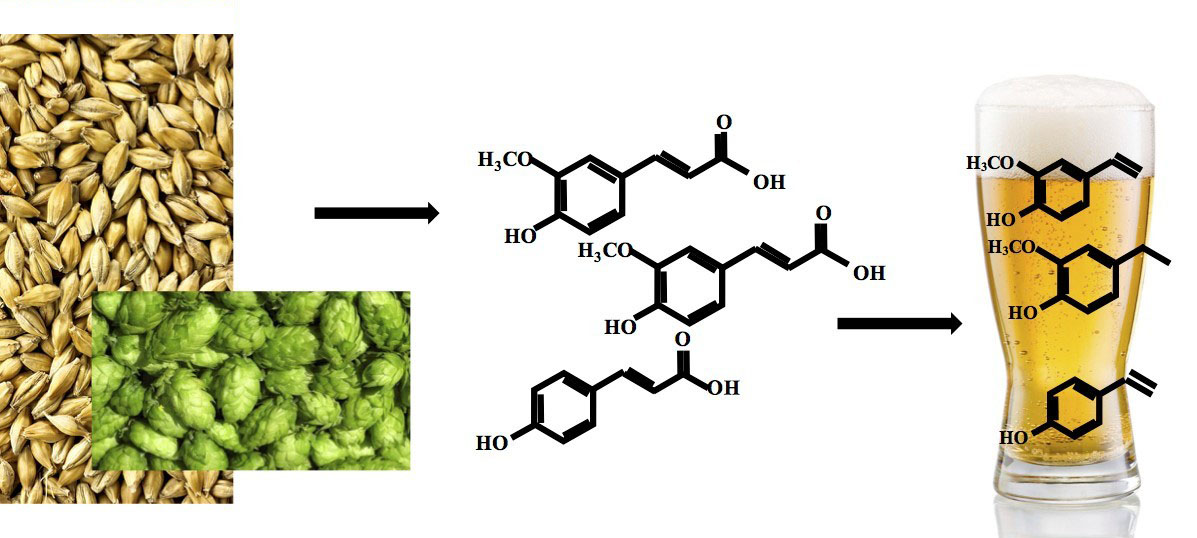
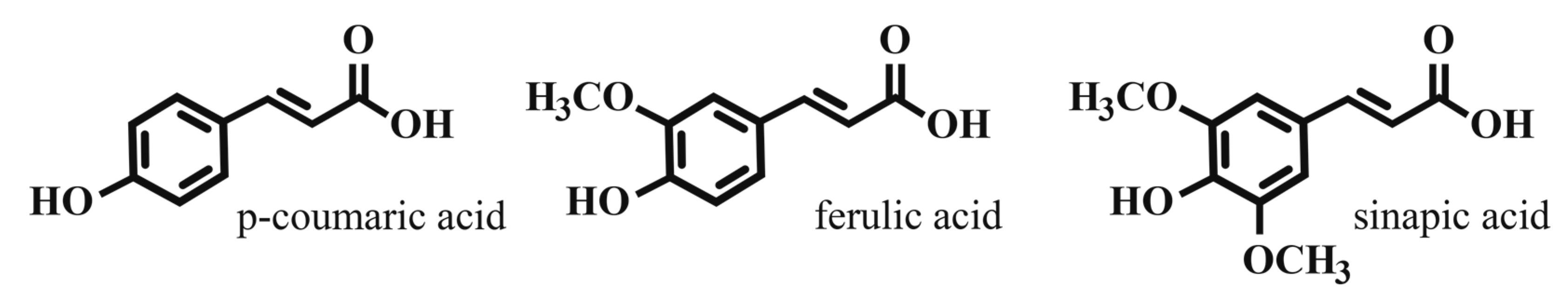
3. Extraction of Simple Phenolics into Beer Wort
3.1. Raw Materials
3.1.1. Grains and Malt
3.1.2. Hops
3.2. Wood Aging
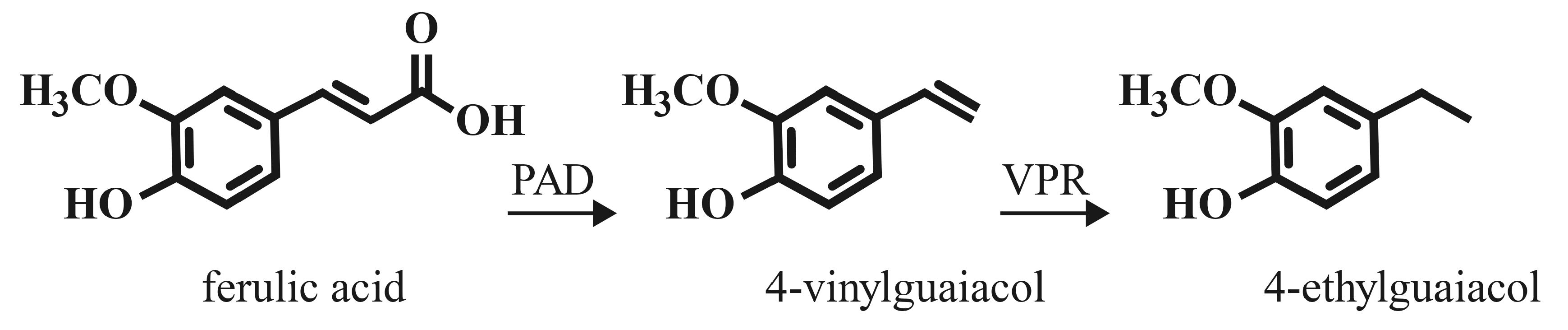
4. Transformation of Hydroxycinnamic Acids to Volatile Compounds
4.1. Thermal Decarboxylation to Vinyl Derivatives
4.2. Microbial Enzymatic Decarboxylation to Vinyl Derivatives
4.3. Vinylphenol Reductase Enzyme Activity
5. Organoleptic Properties of Volatile Phenols in Beer
5.1. Desirable Characteristics
5.2. Undesirable/Spoilage Characteristics
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- The Chemistry of Beer. Available online: https://www.newfoodmagazine.com/article/9164/the-chemistry-of-beer/ (accessed on 17 January 2018).
- Bamforth, C. Beer: A Quality Perspective; Elsevier: London, UK, 2009; pp. 12–27. [Google Scholar]
- Callemien, D.; Collin, S. Structure, organoleptic properties, quantification methods, and stability of phenolic compounds in beer—A review. Food Rev. Int. 2009, 26, 1–84. [Google Scholar] [CrossRef]
- Vanholme, R.; Demedts, B.; Morreel, K.; Ralph, J.; Boerjan, W. Lignin biosynthesis and structure. Plant Physiol. 2010, 153, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Iiyama, K.; Lam, T.B.T.; Stone, B.A. Covalent cross-links in the cell wall. Plant Physiol. 1994, 104, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Faulds, C.B.; Williamson, G. The role of hydroxycinnamates in the plant cell wall. J. Sci. Food Agric. 1999, 79, 393–395. [Google Scholar] [CrossRef]
- Campos, F.M.; Couto, J.A.; Figueiredo, A.R.; Tóth, I.V.; Rangel, A.O.S.S.; Hogg, T.A. Cell membrane damage induced by phenolic acids on wine lactic acid bacteria. Int. J. Food Microbiol. 2009, 135, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Collin, S.; Jerkovic, V.; Bröhan, M.; Callemien, D. Polyphenols and Beer Quality. In Natural Products: Phytochemistry, Botany and Metabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 2333–2359. [Google Scholar]
- McMurrough, I.; Roche, G.P.; Cleary, K.G. Phenolic acids in beers and worts. J. Inst. Brew. 1984, 90, 181–187. [Google Scholar] [CrossRef]
- Wackerbauer, K.; Kramer, P.; Siepert, J. Phenolic aromatic substances in beer. Phenolic carboxylic acid and phenols in the raw material and beer preparation. Brauwelt 1982, 122, 618–620. [Google Scholar]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Variability in the release of free and bound hydroxycinnamic acids from diverse malted barley (Hordeum vulgare L.) cultivars during wort production. J. Agric. Food Chem. 2007, 55, 11002–11010. [Google Scholar] [CrossRef] [PubMed]
- Hernanz, D.; Nuñez, V.; Sancho, A.I.; Faulds, C.B.; Williamson, G.; Bartolomé, B.; Gómez-Cordovés, C. Hydroxycinnamic acids and ferulic acid dehydrodimers in barley and processed barley. J. Agric. Food Chem. 2001, 49, 4884–4888. [Google Scholar] [CrossRef] [PubMed]
- Vanbeneden, N.; Van Roey, T.; Willems, F.; Delvaux, F.; Delvaux, F.R. Release of phenolic flavour precursors during wort production: Influence of process parameters and grist composition on ferulic acid release during brewing. Food Chem. 2008, 111, 83–91. [Google Scholar] [CrossRef]
- McMurrough, I.; Madigan, D.; Donnelly, D.; Hurley, J.; Doyle, A.-H.; Hennigan, G.; McNulty, N.; Smyth, M.R. Control of ferulic acid and 4-vinylguaiacol in brewing. J. Inst. Brew. 1996, 102, 327–332. [Google Scholar] [CrossRef]
- Jamar, C.; du Jardin, P.; Fauconnier, M.-L. Cell wall polysaccharides hydrolysis of malting barley (Hordeum vulgare L.): A review. Biotechnol. Agron. Soc. Environ. 2011, 15, 301–313. [Google Scholar]
- Samaras, T.S.; Camburn, P.A.; Chandra, S.X.; Gordon, M.H.; Ames, J.M. Antioxidant properties of kilned and roasted malts. J. Agric. Food Chem. 2005, 53, 8068–8074. [Google Scholar] [CrossRef] [PubMed]
- Sancho, A.I.; Faulds, C.B.; Bartolomé, B.; Williamson, G. Characterisation of feruloyl esterase activity in barley. J. Sci. Food Agric. 1999, 79, 447–449. [Google Scholar] [CrossRef]
- Coghe, S.; Benoot, K.; Delvaux, F.; Vanderhaegen, B.; Delvaux, F.R. Ferulic acid release and 4-vinylguaiacol formation during brewing and fermentation: Indications for feruloyl esterase activity in Saccharomyces cerevisiae. J. Agric. Food Chem. 2004, 52, 602–608. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Han, Z.; Huang, Y.; Chen, Z.-H.; Zhang, G.; Dai, F. Genetic diversity of individual phenolic acids in barley and their correlation with barley malt quality. J. Agric. Food Chem. 2015, 63, 7051–7057. [Google Scholar] [CrossRef] [PubMed]
- Dvořáková, M.; Douanier, M.; Jurková, M.; Kellner, V.; Dostálek, P. Comparison of antioxidant activity of barley (Hordeum vulgare L.) and malt extracts with the content of free phenolic compounds measured by high performance liquid chromatography coupled with coularray detector. J. Inst. Brew. 2008, 114, 150–159. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Gonçalves, L.M.; Guido, L.F. Overall antioxidant properties of malt and how they are influenced by the individual constituents of barley and the malting process. Compr. Rev. Food Sci. Food Saf. 2016, 15, 927–943. [Google Scholar] [CrossRef]
- Nardini, M.; Ghiselli, A. Determination of free and bound phenolic acids in beer. Food Chem. 2004, 84, 137–143. [Google Scholar] [CrossRef]
- Shewry, P.R. Improving the protein content and composition of cereal grain. J. Cereal Sci. 2007, 46, 239–250. [Google Scholar] [CrossRef]
- Mpofu, A.; Sapirstein, H.D.; Beta, T. Genotype and environmental variation in phenolic content, phenolic acid composition, and antioxidant activity of hard spring wheat. J. Agric. Food Chem. 2006, 54, 1265–1270. [Google Scholar] [CrossRef] [PubMed]
- Dinelli, G.; Segura-Carretero, A.; Di Silvestro, R.; Marotti, I.; Arráez-Román, D.; Benedettelli, S.; Ghiselli, L.; Fernadez-Gutierrez, A. Profiles of phenolic compounds in modern and old common wheat varieties determined by liquid chromatography coupled with time-of-flight mass spectrometry. J. Chromatogr. A 2011, 1218, 7670–7681. [Google Scholar] [CrossRef] [PubMed]
- Zuchowski, J.; Jonczyk, K.; Pecio, L.; Oleszek, W. Phenolic acid concentrations in organically and conventionally cultivated spring and winter wheat. J. Sci. Food Agric. 2011, 91, 1089–1095. [Google Scholar] [CrossRef] [PubMed]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-H.; Tsao, R.; Yang, R.; Cui, S.W. Phenolic acid profiles and antioxidant activities of wheat bran extracts and the effect of hydrolysis conditions. Food Chem. 2006, 95, 466–473. [Google Scholar] [CrossRef]
- Back, W.; Diener, C.; Sacher, B. Hefeweizenbier—Taste spectrum and technology. Brauwelt Int. 2000, 18, 112–119. [Google Scholar]
- Kowalczyk, D.; Świeca, M.; Cichocka, J.; Gawlik-Dziki, U. The phenolic content and antioxidant activity of the aqueous and hydroalcoholic extracts of hops and their pellets. J. Inst. Brew. 2013, 119, 103–110. [Google Scholar] [CrossRef]
- Kellner, V.; Jurková, M.; Čulík, J.; Horák, T.; Čejka, P. Some phenolic compounds in Czech hops and beer of Pilsner type. Brew. Sci. 2007, 60, 32–37. [Google Scholar]
- Pascoe, H.M.; Ames, J.M.; Chandra, S. Critical stages of the brewing process for changes in antioxidant activity and levels of phenolic compounds in ale. J. Am. Soc. Brew. Chem. 2003, 61, 203–209. [Google Scholar]
- Proestos, C.; Komaitis, M. Antioxidant capacity of hops. Beer Health Dis. Prev. 2009, 467–475. [Google Scholar]
- Pasteur, L.; Robb, D.C.; Faulkner, F. Studies on Fermentation; the Diseases of Beer, Their Causes, and the Means of Preventing Them; A translation, Made with the Author’s Sanction, of Études sur la bière; with Notes, Index, and Original Illustrations; MacMillan: Basingstoke, UK, 1879. [Google Scholar]
- Flanders Red Ales. Available online: http://www.milkthefunk.com/wiki/Flanders_Red_Ale (accessed on 18 January 2018).
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of an industrially produced lambic beer shares members of a traditionally produced one and reveals a core microbiota for lambic beer fermentation. Food Microbiol. 2015, 49, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Snauwaert, I.; Roels, S.P.; Van Nieuwerburg, F.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. Microbial diversity and metabolite composition of Belgian red-brown acidic ales. Int. J. Food Microbiol. 2016, 221, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Martens, H.; Iserentant, D.; Verachtert, H. Microbiological aspects of a mixed yeast-bacterial fermentation in the production of a special Belgian acidic ale. J. Inst. Brew. 1997, 103, 85–91. [Google Scholar] [CrossRef]
- Van Oevelen, D.; Spaepen, M.; Timmermans, P.; Verachtert, H. Microbial aspects of spontaneous fermentation in production of lambic and gueuze. J. Inst. Brew. 1977, 83, 356–360. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Bamforth, C.W.; Mills, D.A. Brewhouse-resident microbiota are responsible for multi-stage fermentation of American coolship ale. PLoS ONE 2012, 7, e35507. [Google Scholar] [CrossRef] [PubMed]
- Spitaels, F.; Wieme, A.D.; Janssens, M.; Aerts, M.; Daniel, H.-M.; Van Landschoot, A.; De Vuyst, L.; Vandamme, P. The microbial diversity of traditional spontaneously fermented lambic beer. PLoS ONE 2014, 9, e95384. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Xu, Y.; Yu, A. Influence of oak chips geographical origin, toast level, dosage and aging time on volatile compounds of apple cider. J. Inst. Brew. 2006, 112, 255–263. [Google Scholar] [CrossRef]
- Wyler, P.; Angeloni, L.H.P.; Alcarde, A.R.; da Cruz, S.H. Effect of oak wood on the quality of beer. J. Inst. Brew. 2015, 121, 62–69. [Google Scholar] [CrossRef]
- Cadahía, E.; Fernández de Simón, B.; Jalocha, J. Volatile compounds in Spanish, French, and American oak woods after natural seasoning and toasting. J. Agric. Food Chem. 2003, 51, 5923–5932. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.P.; Howell, G.S.; Michaelis, C.S.; Dickmann, D.I. The content of phenolic acid and aldehyde flavor components of white oak as affected by site and species. Am. J. Enol. Vitic. 1992, 43, 333–338. [Google Scholar]
- Sara, C.; Concei, L.M.; Isabel, S.M.; Pedro, B.A. Influence of botanical species and geographical origin on the content of low molecular weight phenolic compounds of woods used in Portuguese cooperage. Holzforschung 2005, 54, 255–261. [Google Scholar]
- Garcia, R.; Soares, B.; Dias, C.B.; Freitas, A.M.C.; Cabrita, M.J. Phenolic and furanic compounds of Portuguese chestnut and French, American and Portuguese oak wood chips. Eur. Food Res. Technol. 2012, 235, 457–467. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Flavor chemistry of beer: Part II: Flavour and threshold of 239 aroma volatiles. MBAA Tech. Q. 1975, 3, 151–168. [Google Scholar]
- Vanbeneden, N.; Gils, F.; Delvaux, F.; Delvaux, F.R. Formation of 4-vinyl and 4-ethyl derivatives from hydroxycinnamic acids: Occurrence of volatile phenolic flavour compounds in beer and distribution of Pad1-activity among brewing yeasts. Food Chem. 2008, 107, 221–230. [Google Scholar] [CrossRef]
- Spitaels, F.; Kerrebroeck, S.V.; Wieme, A.D.; Snauwaert, I.; Aerts, M.; Landschoot, A.V.; De Vuyst, L.; Vandamme, P. Microbiota and metabolites of aged bottled gueuze beers converge to the same composition. Food Microbiol. 2015, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Barthelmebs, L.; Divies, C.; Cavin, J.M. Knockout of the p-coumarate decarboxylase gene from Lactobacillus plantarum reveals the existence of two other inducible enzymatic activities involved in phenolic acid metabolism. Appl. Environ. Microbiol. 2000, 66, 3368–3375. [Google Scholar] [CrossRef] [PubMed]
- Bhuiya, M.W.; Lee, S.G.; Jez, J.M.; Yu, O. Structure and mechanism of ferulic acid decarboxylase (FDC1) from Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2015, 81, 4216–4223. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Ferguson, K.L.; Boyer, D.R.; Lin, X.N.; Marsh, E.N.G. Isofunctional enzymes PAD1 and UbiX catalyze formation of a novel cofactor required by ferulic acid decarboxylase and 4-hydroxy-3-polyprenylbenzoic acid decarboxylase. ACS Chem. Biol. 2015, 10, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Lind, M.E.S.; Himo, F. Theoretical study of the reaction mechanism of phenolic acid decarboxylase. FEBS J. 2015, 282, 4703–4713. [Google Scholar] [CrossRef] [PubMed]
- Mukai, N.; Masaki, K.; Fujii, T.; Kawamukai, M.; Iefuji, H. PAD1 and FDC1 are essential for the decarboxylation of phenylacrylic acids in Saccharomyces cerevisiae. J. Biosci. Bioeng. 2010, 109, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Gallone, B.; Steensels, J.; Prahl, T.; Soriaga, L.; Saels, V.; Herrera-Malaver, B.; Merlevede, A.; Roncoroni, M.; Voordeckers, K.; Miraglia, L.; et al. Domestication and divergence of Saccharomyces cerevisiae beer yeasts. Cell 2016, 166, 1397–1410. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, H.; Angulo, I.; de las Rivas, B.; Campillo, N.; Páez, J.A.; Muñoz, R.; Mancheño, J.M. p-Coumaric acid decarboxylase from Lactobacillus plantarum: Structural insights into the active site and decarboxylation catalytic mechanism. Proteins: Struct. Funct. Bioinform. 2010, 78, 1662–1676. [Google Scholar] [CrossRef]
- Barthelmebs, L.; Diviès, C.; Cavin, J.F. Expression in Escherichia coli of native and chimeric phenolic acid decarboxylases with modified enzymatic activities and method for screening recombinant E. coli strains expressing these enzymes. Appl. Environ. Microbiol. 2001, 67, 1063–1069. [Google Scholar] [CrossRef] [PubMed]
- Landete, J.M.; Rodríguez, H.; Curiel, J.A.; de las Rivas, B.; Mancheño, J.M.; Muñoz, R. Gene cloning, expression, and characterization of phenolic acid decarboxylase from Lactobacillus brevis RM84. J. Ind. Microbiol. Biotechnol. 2010, 37, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Annemüller, G. Die Berliner Weiße, 1st ed.; VLB: Berlin, Germany, 2008. [Google Scholar]
- Van Oevelen, D.; Delescaille, F.; Verachtert, H. Synthesis of aroma compounds during spontaneous fermentation of lambic and gueuze. J. Inst. Brew. 1976, 82, 322–326. [Google Scholar] [CrossRef]
- Spaepen, M.; Van Oevelen, D.; Verachtert, H. Fatty acids and esters produced during spontaneous fermentation of lambic and gueuze. J. Inst. Brew. 1978, 84, 278–282. [Google Scholar] [CrossRef]
- Crauwels, S.; Steensels, J.; Aerts, G.; Willems, K.; Verstrepen, K.; Lievens, B. Brettanomyces bruxellensis, essential contributor in spontaneous beer fermentations providing novel opportunities for the brewing industry. Brew. Sci. 2015, 68, 110–121. [Google Scholar]
- Conterno, L.; Joseph, C.M.L.; Arvik, T.J.; Henick-Kling, T.; Bisson, L.F. Genetic and physiological characterization of Brettanomyces bruxellensis strains isolated from wines. Am. J. Enol. Vitic. 2006, 57, 139–147. [Google Scholar]
- Lentz, M.; Harris, C. Analysis of growth inhibition and metabolism of hydroxycinnamic acids by brewing and spoilage strains of Brettanomyces yeast. Foods 2015, 4, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Zhu, B.; Steensels, J.; Busschaert, P.; De Samblanx, G.; Marchal, K.; Willems, K.A.; Verstrepen, K.J.; Lievens, B. Assessing genetic diversity among Brettanomyces yeasts by DNA fingerprinting and whole-genome sequencing. Appl. Environ. Biol. 2014, 80, 4398–4413. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Van Opstaele, F.; Jaskula-Goiris, B.; Steensels, J.; Verreth, C.; Bosmans, L.; Paulussen, C.; Herrera-Malaver, B.; de Jonge, R.; De Clippeleer, J.; et al. Fermentation assays reveal differences in sugar and (off-)flavor metabolism across different Brettanomyces bruxellensis strains. FEMS Yeast Res. 2017, 17, fow105. [Google Scholar] [CrossRef] [PubMed]
- Crauwels, S.; Van Assche, A.; de Jonge, R.; Borneman, A.R.; Verreth, C.; Troels, P.; De Samblanx, G.; Marchal, K.; Van, d.P.Y.; Willems, K.A.; et al. Comparative phenomics and targeted use of genomics reveals variation in carbon and nitrogen assimilation among different Brettanomyces bruxellensis strains. Appl. Microbiol. Biotechnol. 2015, 99, 9123–9134. [Google Scholar] [CrossRef] [PubMed]
- Godoy, L.; Garrido, D.; Martinez, C.; Saaverda, J.; Combina, M.; Ganga, M.A. Study of the coumarate decarboxylase and vinylphenol reductase activities of Dekkera bruxellensis (anamorph Brettanomyces bruxellensis) isolates. Lett. Appl. Microbiol. 2009, 48, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Vigentini, I.; Romano, A.; Compagno, C.; Merico, A.; Molinari, F.; Tirelli, A.; Foschino, R.; Volonterio, G. Physiological and oenological traits of different Dekkera/Brettanomyces bruxellensis strains under wine-model conditions. FEMS Yeast Res. 2008, 8, 1087–1096. [Google Scholar] [CrossRef] [PubMed]
- Chatonnet, P.; Dubourdieu, D.; Boidron, J.N.; Pons, M. The origin of ethylphenols in wines. J. Sci. Food Agric. 1992, 60, 165–178. [Google Scholar] [CrossRef]
- Romano, D.; Valdetara, F.; Zambelli, P.; Galafassi, S.; De Vitis, V.; Molinari, F.; Compagno, C.; Foschino, R.; Vigentini, I. Cloning the putative gene of vinyl phenol reductase of Dekkera bruxellensis in Saccharomyces cerevisiae. Food Microbiol. 2017, 63, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Dias, L.; Dias, S.; Sancho, T.; Stender, H.; Querol, A.; Malfeito-Ferreira, M.; Loureiro, V. Identification of yeasts isolated from wine-related environments and capable of producing 4-ethylphenol. Food Microbiol. 2003, 20, 567–574. [Google Scholar] [CrossRef]
- Sangorrín, M.P.; García, V.; Lopes, C.A.; Sáez, J.S.; Martínez, C.; Ganga, M.A. Molecular and physiological comparison of spoilage wine yeasts. J. Appl. Microbiol. 2013, 114, 1066–1074. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, A.; Forte, M.; Nardini, M. Characterization of phenolics content and antioxidant activity of different beer types. J. Agric. Food Chem. 2010, 58, 10677–10683. [Google Scholar] [CrossRef] [PubMed]
- Achilli, G.; Piero Cellerino, G.; Gamache, P.H.; Vico Melzi d’Eril, G. Identification and determination of phenolic constituents in natural beverages and plant extracts by means of a coulometric electrode array system. J. Chromatogr. A 1993, 632, 111–117. [Google Scholar] [CrossRef]
- Floridi, S.; Montanari, L.; Marconi, O.; Fantozzi, P. Determination of free phenolic acids in wort and beer by coulometric array detection. J. Agric. Food Chem. 2003, 51, 1548–1554. [Google Scholar] [CrossRef] [PubMed]
- Montanari, L.; Perretti, G.; Natella, F.; Guidi, A.; Fantozzi, P. Organic and phenolic acids in beer. LWT—Food Sci. Technol. 1999, 32, 535–539. [Google Scholar] [CrossRef]
- Heresztyn, T. Metabolism of volatile phenolic compounds from hydroxycinnamic acids by Brettanomyces yeast. Arch. Microbiol. 1986, 146, 96–98. [Google Scholar] [CrossRef]
- Oelofse, A.; Lonvaud-Funel, A.; du Toit, M. Molecular identification of Brettanomyces bruxellensis strains isolated from red wines and volatile phenol production. Food Microbiol. 2009, 26, 377–385. [Google Scholar] [CrossRef] [PubMed]
- The Good Scents Company. Available online: http://www.thegoodscentscompany.com/ (accessed on 5 December 2017).
- Flavor and Extract Manufacturers Association of the United States. Available online: https://www.femaflavor.org/about (accessed on 5 December 2017).
- Wasserman, A.E. Organoleptic evaluation of three phenols present in wood smoke. J. Food Sci. 1966, 31, 1005–1010. [Google Scholar] [CrossRef]
- Vanbeneden, N.; Saison, D.; Delvaux, F.; Delvaux, F.R. Decrease of 4-vinylguaiacol during beer aging and formation of apocynol and vanillin in beer. J. Agric. Food Chem. 2008, 56, 11983–11988. [Google Scholar] [CrossRef] [PubMed]
- Wackerbauer, K.; Kramer, P. Bavarian wheat beer—An alternative. Production and composition. Brauwelt 1982, 122, 758–760. [Google Scholar]
- Langos, D.; Granvogl, M.; Schieberle, P. Characterization of the key aroma compounds in two Bavarian wheat beers by means of the sensomics approach. J. Agric. Food Chem. 2013, 61, 11303–11311. [Google Scholar] [CrossRef] [PubMed]
- Langos, D.; Granvogl, M. Studies on the simultaneous formation of aroma-active and toxicologically relevant vinyl aromatics from free phenolic acids during wheat beer brewing. J. Agric. Food Chem. 2016, 64, 2325–2332. [Google Scholar] [CrossRef] [PubMed]
- Narziss, L.; Miedaner, H.; Neizsche, F. The formation of 4-vinylguaiacol during the manufacture of Bavarian wheat beers (weizenbier). Monatsschrift Fuer Brauwissenschaft 1990, 43, 96–100. [Google Scholar]
- Kieninger, H.; Narziss, L.; Miedaner, H.; Hecht, S. Concerning the changes of quality determining molecular groups in the preparation of Bavarian wheat beers. Brauwissenschaft 1984, 37, 9–16. [Google Scholar]
- Schwarz, K.J.; Boitz, L.I.; Methner, F.-J. Release of phenolic acids and amino acids during mashing dependent on temperature, pH, time, and raw materials. J. Am. Soc. Brew. Chem. 2012, 70, 290–295. [Google Scholar] [CrossRef]
- Schwarz, K.J.; Stübner, R.; Methner, F.-J. Formation of styrene dependent on fermentation management during wheat beer production. Food Chem. 2012, 134, 2121–2125. [Google Scholar] [CrossRef] [PubMed]
- White, C.; Zainasheff, J. Yeast: The Practical Guide to Beer Fermentation; Brewers: Boulder, CO, USA, 2010. [Google Scholar]
- Mertens, S.; Steensels, J.; Gallone, B.; Souffriau, B.; Malcorps, P.; Verstrepen, K.J. Rapid screening method for phenolic off-flavor (POF) production in yeast. Am. Soc. Brew. Chem. 2017, 75, 318–323. [Google Scholar] [CrossRef]
- 2015 Beer Style Guidelines. Available online: https://www.bjcp.org/docs/2015_Guidelines_Beer.pdf (accessed on 22 January 2018).
- Competition Style List, Descriptions, and Specifications. 2017. Available online: https://www.greatamericanbeerfestival.com/wp-content/uploads/2017/05/17_GABF_Beer_Style_Guidelines_Final.pdf (accessed on 22 January 2018).
- Competition Style List, Descriptions, and Specifications. 2018. Available online: https://www.worldbeercup.org/wp-content/uploads/2013/09/18_WBC_Style_Guidelines_Final.pdf (accessed on 22 January 2018).
- Yeast Bank. Available online: https://www.whitelabs.com/yeast-bank (accessed on 22 January 2018).
- Yeast Strains for Beer. Available online: http://www.wyeastlab.com/beer-strains (accessed on 22 January 2018).
- Yeast Descriptions Guide. Available online: http://www.brewingscience.com/PDF/prodlist/BSI_Yeast_Descriptions_Guide.pdf (accessed on 18 January 2018).
- Jespersen, L.; Jakobsen, M. Specific spoilage organisms in breweries and laboratory media for their detection. Int. J. Food Microbiol. 1996, 33, 139–155. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Lawrence, D.R. Spoilage Organisms in Beer. In Developments in Food Microbiology; Robinson, R.K., Ed.; Elsevier: London, UK, 1988; pp. 1–48. [Google Scholar]
- Shinohara, T.; Kubodera, S.; Yanagida, F. Distribution of phenolic yeasts and production of phenolic off-flavors in wine fermentation. J. Biosci. Bioeng. 2000, 90, 90–97. [Google Scholar] [CrossRef]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 411–430. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Compound | Descriptors | Sources | References |
|---|---|---|---|
| 4-vinylguaiacol | Clove, curry, spice, smoky, bacon | Thermal or enzymatic decarboxylation of ferulic acid | [3,18,81] |
| 4-vinylphenol | Phenolic, medicinal, spicy | Thermal or enzymatic decarboxylation of p-coumaric acid | [18,81] |
| 4-ethylguaiacol | Clove, phenol, spice, woody, smoky, vanilla | Enzymatic reduction of 4-vinyl guaiacol | [3,71,81,82] |
| 4-ethylphenol | Leather, phenol, spice, stable, smoke, creosote | Enzymatic reduction of 4-vinyl phenol | [3,71,81,82] |
| Guaiacol | Smoke, bacon | Lignin pyrolysis | [3,83] |
| Vanillin | Sweet, vanilla | Lignin pyrolysis, wood aging, degradation of 4-vinyl guaiacol | [3,84] |
| 4-vinyl syringol | Stale, “old beer” | Degradation of synapic acid glycosides | [3,8] |
© 2018 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lentz, M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation 2018, 4, 20. https://doi.org/10.3390/fermentation4010020
Lentz M. The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation. 2018; 4(1):20. https://doi.org/10.3390/fermentation4010020
Chicago/Turabian StyleLentz, Michael. 2018. "The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor" Fermentation 4, no. 1: 20. https://doi.org/10.3390/fermentation4010020
APA StyleLentz, M. (2018). The Impact of Simple Phenolic Compounds on Beer Aroma and Flavor. Fermentation, 4(1), 20. https://doi.org/10.3390/fermentation4010020