
Use of Indigenous Lactic Acid Bacteria for Industrial Fermented Sausage Production: Microbiological, Chemico-Physical and Sensory Features and Biogenic Amine Content

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. LAB Strains Used as Starter Cultures
2.2. Fermented Sausage Production
2.3. Physico-Chemical Parameters
2.4. Microbial Counts
2.5. Biogenic Amine Quantification
2.6. Aroma Profile
2.7. Consumer Test
2.8. Statistical Analysis
3. Results and Discussion
3.1. Evolution of Physico-Chemical Parameters (pH, aw and Weight Loss)
3.2. Microbial Characterisation
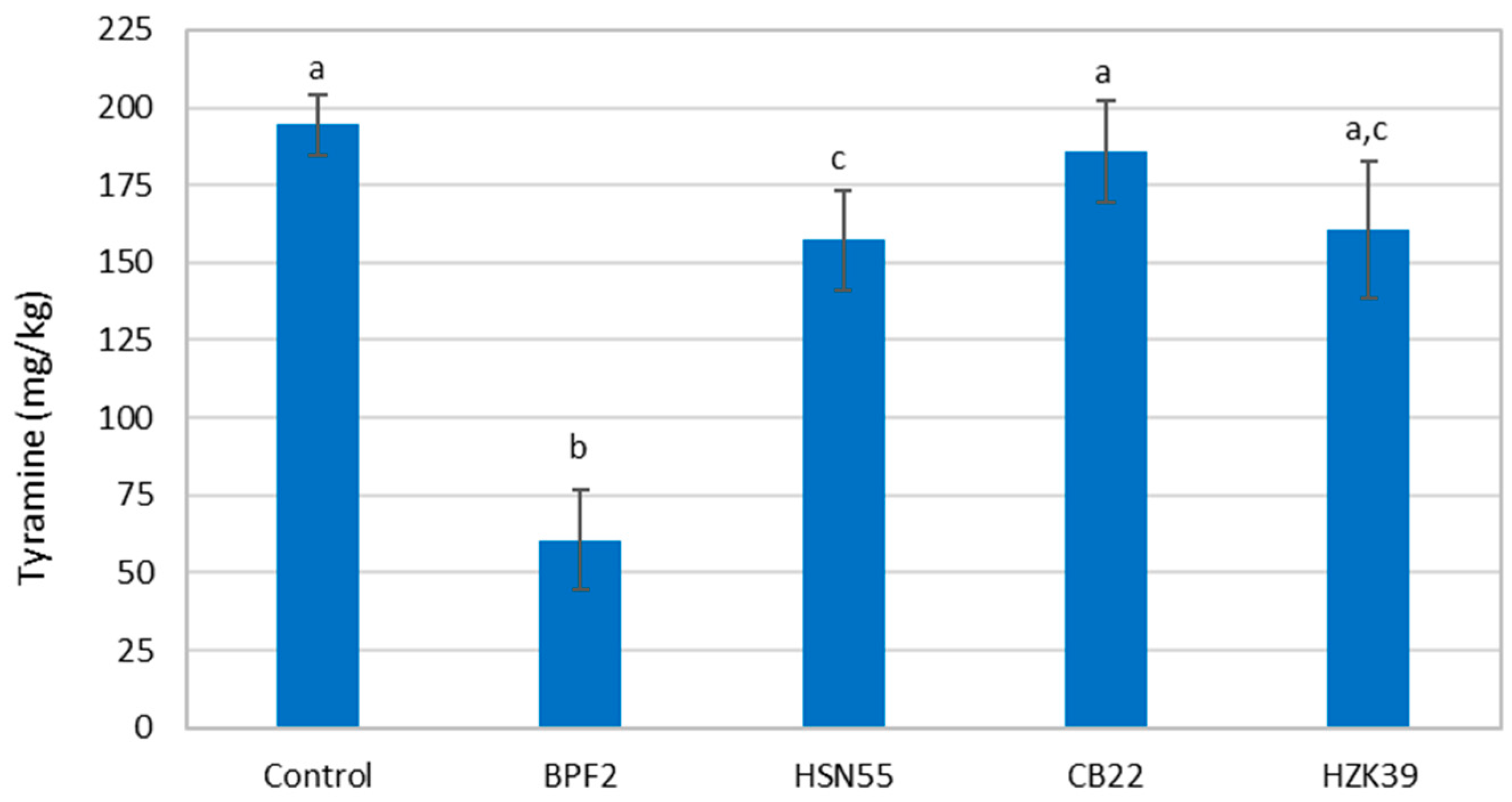
3.3. Biogenic Amine Determination
3.4. Aroma Profile of Fermented Sausages
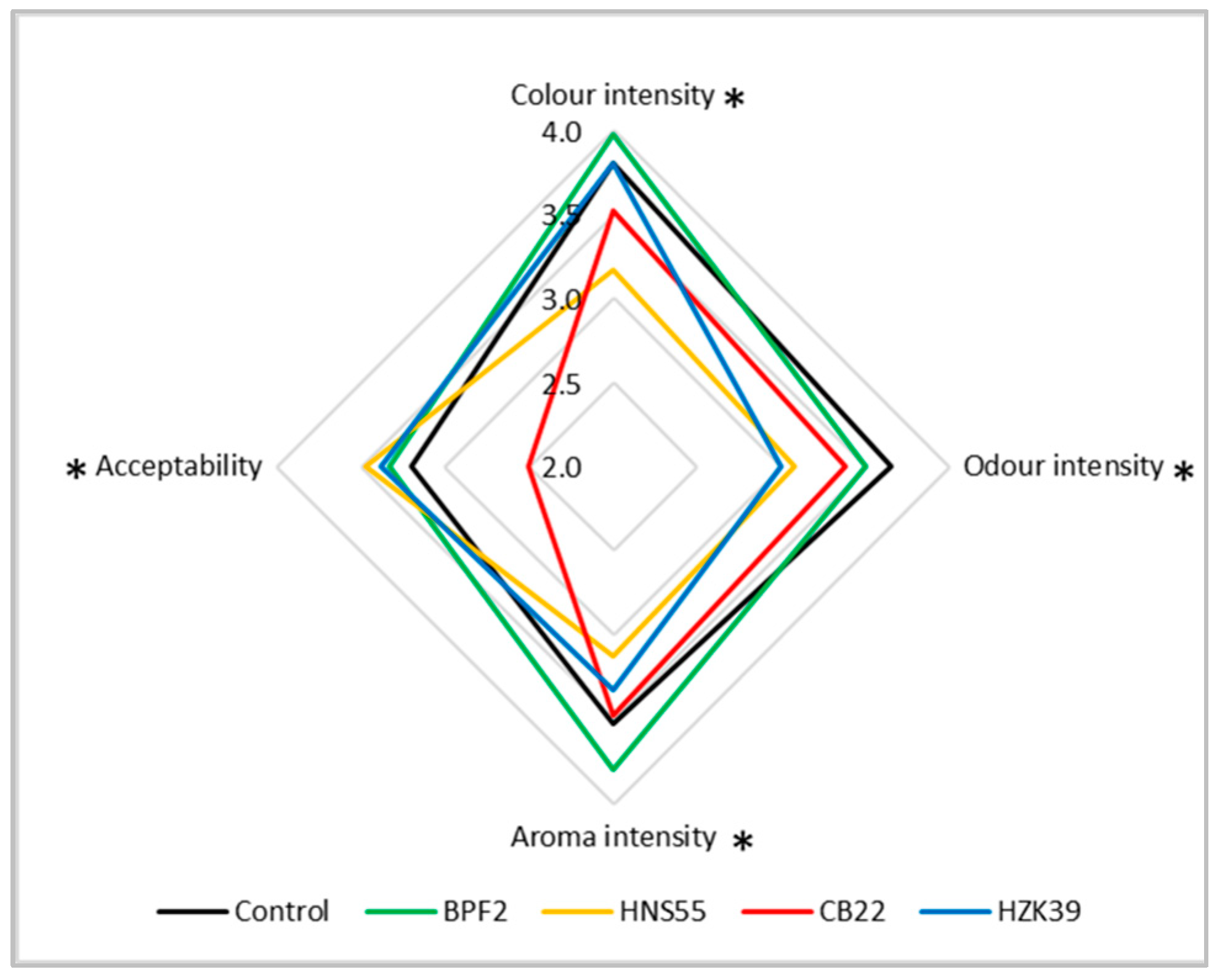
3.5. Sensory Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feiner, G. Meat Products Handbook. Practical Science and Technology, 1st ed.; Woodhead Publishing: Cambridge, UK, 2006. [Google Scholar]
- Cocconcelli, P.S.; Fontana, C. Starter cultures for meat fermentation. In Handbook of Meat Processing; Toldrá, F., Ed.; Blackwell Publishing: Ames, IA, USA, 2010; pp. 199–218. [Google Scholar]
- García-Díez, J.; Saraiva, C. Use of starter cultures in foods from animal origin to improve their safety. Int. J. Environ. Res. Publ. Health 2021, 18, 2544. [Google Scholar] [CrossRef] [PubMed]
- Cocconcelli, P.S.; Fontana, C. Bacteria. In Handbook of Fermented Meat and Poultry; Toldrá, F., Hui, Y.H., Astiasarán, I., Sebranek, J.G., Talon, R., Eds.; Wiley Blackwell: Ames, IA, USA, 2014; pp. 117–128. [Google Scholar]
- Laranjo, M.; Potes, M.E.; Elias, M. Role of starter cultures on the safety of fermented meat products. Front. Microbiol. 2019, 10, 853. [Google Scholar] [CrossRef] [PubMed]
- Seleshe, S.; Kang, S.N. Effect of Different Pediococcus pentosaceus and Lactobacillus plantarum strains on quality characteristics of dry fermented sausage after completion of ripening period. Food Sci. Anim. Resour. 2021, 41, 636–649. [Google Scholar] [CrossRef] [PubMed]
- Trząskowska, M.; Kołożyn-Krajewska, D.; Wójciak, K.; Dolatowski, Z. Microbiological quality of raw-fermented sausages with Lactobacillus casei LOCK 0900 probiotic strain. Food Control 2014, 35, 184–191. [Google Scholar] [CrossRef]
- Carballo, J. The role of fermentation reactions in the generation of flavor and aroma of foods. In Fermentation, Effects on Food Properties; Mehta, B.M., Kamal-Eldin, A., Iwanski, R.Z., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 51–83. [Google Scholar]
- Franciosa, L.; Alessandria, V.; Dolci, P.; Rantsiou, K.; Cocolin, L. Sausage fermentation and starter cultures in the era of molecular biology methods. Int. J. Food Microbiol. 2018, 279, 26–32. [Google Scholar] [CrossRef]
- Barbieri, F.; Tabanelli, G.; Montanari, C.; Dall’Osso, N.; Šimat, V.; Smole Možina, S.; Baños, A.; Özogul, F.; Bassi, D.; Fontana, C.; et al. Mediterranean spontaneously fermented sausages: Spotlight on microbiological and quality features to exploit their bacterial biodiversity. Foods 2021, 10, 2691. [Google Scholar] [CrossRef]
- dos Santos Cruxen, C.E.; Funck, G.D.; Haubert, L.; da Silva Dannenberg, G.; de Lima Marques, J.; Chaves, F.C.; da Silva, W.P.; Fiorentini, Â.M. Selection of native bacterial starter culture in the production of fermented meat sausages: Application potential, safety aspects, and emerging technologies. Food Res. Int. 2019, 122, 371–382. [Google Scholar] [CrossRef]
- Franciosa, I.; Ferrocino, I.; Corvaglia, M.R.; Giordano, M.; Coton, M.; Mounier, J.; Rantsiou, K.; Cocolin, L. Autochthonous starter culture selection for Salame Piemonte PGI production. Food Res. Int. 2022, 162, 112007. [Google Scholar] [CrossRef]
- Talon, R.; Leroy, S.; Lebert, I.; Giammarinaro, P.; Chacornac, J.P.; Latorre-Moratalla, M.; Vidal-Carou, C.; Zanardi, E.; Conter, M.; Lebecque, A. Safety improvement and preservation of typical sensory qualities of traditional dry fermented sausages using autochthonous starter cultures. Int. J. Food Microbiol. 2008, 126, 227–234. [Google Scholar] [CrossRef]
- Chikindas, M.L.; Weeks, R.; Drider, D.; Chistyakov, V.A.; Dicks, L.M. Functions and emerging applications of bacteriocins. Curr. Opin. Biotechnol. 2018, 49, 23–28. [Google Scholar] [CrossRef]
- Oliveira, M.; Ferreira, V.; Magalhães, R.; Paula Teixeira, P. Biocontrol strategies for Mediterranean-style fermented sausages. Food Res. Int. 2018, 103, 438–449. [Google Scholar] [CrossRef] [PubMed]
- Bassi, D.; Milani, G.; Belloso Daza, M.V.; Barbieri, F.; Montanari, C.; Lorenzini, S.; Šimat, V.; Gardini, F.; Tabanelli, G. Taxonomical identification and safety characterization of Lactobacillaceae from Mediterranean natural fermented sausages. Foods 2022, 11, 2776. [Google Scholar] [CrossRef] [PubMed]
- García-López, J.D.; Teso-Pérez, C.; Martín-Platero, A.M.; Peralta-Sánchez, J.M.; Fonollá- Joya, J.; Martínez-Bueno, M.; Baños, A. Lactiplantibacillus paraplantarum BPF2 and Pediococcus acidilactici ST6, two bacteriocinogenic isolated strains from Andalusian spontaneous fermented sausages. Foods 2023, 12, 2445. [Google Scholar] [CrossRef] [PubMed]
- Tabanelli, G.; Barbieri, F.; Baños, A.; Garcia Madero, J.M.; Belloso Daza, M.V.; Cortimiglia, C.; Milani, G.; Bassi, D.; Gardini, F.; Montanari, C. Companilactobacillus alimentarius: An extensive characterization of strains isolated from spontaneous fermented sausages. Int. J. Food Microbiol. 2024, 410, 110489. [Google Scholar] [CrossRef]
- Milani, G.; Tabanelli, G.; Barbieri, F.; Montanari, C.; Gardini, F.; Belloso Daza, M.V.; Castellone, V.; Bozzetti, M.; Cocconcelli, P.S.; Bassi, D. Technological traits and mitigation activity of autochthonous lactic acid bacteria from mediterranean fermented meat-products. LWT 2024, 196, 115861. [Google Scholar] [CrossRef]
- García-López, J.D.; Barbieri, F.; Baños, A.; Garcia Madero, J.M.; Gardini, F.; Montanari, C.; Tabanelli, G. Use of two autochthonous bacteriocinogenic strains as starter cultures in the production of salchichónes, a type of Spanish fermented sausages. Curr. Res. Food Sci. 2023, 7, 100615. [Google Scholar] [CrossRef]
- Montanari, C.; Barbieri, F.; Gardini, F.; Tabanelli, G. Competition between starter cultures and wild microbial population in sausage fermentation: A case study regarding a typical Italian salami (Ventricina). Foods 2021, 10, 2138. [Google Scholar] [CrossRef]
- Martuscelli, M.; Crudele, M.A.; Gardini, F.; Suzzi, G. Biogenic amine formation and oxidation by Staphylococcus xylosus strains from artisanal fermented sausages. Lett. Appl. Microbiol. 2000, 31, 228–232. [Google Scholar] [CrossRef]
- Montanari, C.; Bargossi, E.; Gardini, A.; Lanciotti, R.; Magnani, R.; Gardini, F.; Tabanelli, G. Correlation between volatile profiles of Italian fermented sausages and their size and starter culture. Food Chem. 2016, 192, 736–744. [Google Scholar] [CrossRef]
- NIST. NIST/NIH/EPA mass spectral library. In Standard Reference Database 1, NIST 11. Standard Reference Data Program; National Institute of Standards and Technology: Gaithersburg, MD, USA, 2011. [Google Scholar]
- Vignolo, G.; Fontana, C.; Fadda, S. Semidry and dry fermented sausages. In Handbook of Meat Processing; Toldrá, F., Ed.; Blackwell Publishing: Ames, IA, USA, 2010; Chapter 22; pp. 379–398. [Google Scholar]
- Montanari, C.; Barbieri, F.; Gardini, G.; Magnani, R.; Gottardi, D.; Gardini, F.; Tabanelli, G. Effects of starter cultures and type of casings on the microbial features and volatile profile of fermented sausages. Fermentation 2022, 8, 683. [Google Scholar] [CrossRef]
- Pellegrini, M.; Barbieri, F.; Montanari, C.; Iacumin, L.; Bernardi, C.; Gardini, F.; Comi, G. Microbial spoilage of traditional goose sausages produced in a Northern Region of Italy. Microorganisms 2023, 11, 1942. [Google Scholar] [CrossRef] [PubMed]
- Comi, G.; Urso, R.; Iacumin, L.; Rantsiou, K.; Cattaneo, P.; Cantoni, C.; Cocolin, L. Characterisation of naturally fermented sausages produced in the North East of Italy. Meat Sci. 2005, 69, 381–392. [Google Scholar] [CrossRef] [PubMed]
- Barbieri, F.; Montanari, C.; Gardini, F.; Tabanelli, G. Biogenic amine production by lactic acid bacteria: A review. Foods 2019, 8, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Fernández, M.; Calles-Enríquez, M.; Sánchez-Llana, E.; Cañedo, E.; Martin, M.C.; Alvarez, M.A. Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 2012, 30, 132–138. [Google Scholar] [CrossRef] [PubMed]
- García Fontán, M.C.; Lorenzo, J.M.; Parada, A.; Franco, I.; Carballo, J. Microbiological characteristics of Androlla, a Spanish traditional pork sausage. Food Microbiol. 2007, 24, 52–58. [Google Scholar] [CrossRef]
- García Fontán, M.C.; Lorenzo, J.M.; Martínez, S.; Franco, I.; Carballo, J. Microbiological characteristics of Botillo, a Spanish traditional pork sausage. LWT 2007, 40, 1610–1622. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2248–2393. [Google Scholar]
- Smid, E.J.; Kleerebezem, M. Production of aroma compounds in lactic fermentations. Annu. Rev. Food Sci. Technol. 2014, 5, 313–326. [Google Scholar] [CrossRef]
- Montanari, C.; Gatto, V.; Torriani, S.; Barbieri, F.; Bargossi, E.; Lanciotti, R.; Grazia, L.; Magnani, R.; Tabanelli, G.; Gardini, F. Effects of the diameter on physico-chemical, microbiological and volatile profile in dry fermented sausages produced with two different starter cultures. Food Biosci. 2018, 22, 9–18. [Google Scholar] [CrossRef]
- Olivares, A.; Navarro, J.L.; Flores, M. Effect of fat content on aroma generation during processing of dry fermented sausages. Meat Sci. 2011, 87, 264–273. [Google Scholar] [CrossRef]
- Flores, M. Understanding the implications of current health trends on the aroma of wet and dry cured meat products. Meat Sci. 2018, 144, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, J.A.; Hierro, E.M.; Bruna, J.M.; de la Hoz, L. Changes in the components of dry-fermented sausages during ripening. Crit. Rev. Food Sci. Nutr. 1999, 39, 329–367. [Google Scholar] [CrossRef] [PubMed]
- Speranza, G.; Corti, S.; Fontana, G.; Manitto, P. Conversion of meso-2,3-butanediol into 2-butanol by Lactobacilli. Stereochemical and enzymatic aspects. J. Agric. Food Chem. 1997, 45, 3476–3480. [Google Scholar] [CrossRef]
- Gänzle, M.G. Lactic metabolism revisited: Metabolism of lactic acid bacteria in food fermentations and food spoilage. Curr. Opin. Food Sci. 2015, 2, 106–117. [Google Scholar] [CrossRef]
- von Wright, A.; Axelsson, L. Lactic acid bacteria: An introduction. In Lactic Acid Bacteria: Microbiological and Functional Aspects, 4th ed.; Lahtinen, S., Ouwehand, A.C., Salminen, S., von Wright, A., Eds.; CRC Press: Boca Raton, FL, USA, 2011; pp. 1–16. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | LAB Strain | Origin |
|---|---|---|
| Control | Latilactobacillus sakei TMX | Commercial starter |
| BPF2 | Lactiplantibacillus paraplantarum BPF2 | Spanish fermented sausage |
| HNS55 | Latilactobacillus curvatus HNS55 | Croatian fermented sausage |
| CB22 | Companilactobacillus alimentarius CB22 | Spanish fermented sausage |
| HZK39 | Latilactobacillus sakei HZK39 | Croatian fermented sausage |
| Microbial Group | Time (Days) | Control | BPF2 | HNS55 | CB22 | HZK39 |
|---|---|---|---|---|---|---|
| Lactobacilli | Meat batter | 4.23 ± 0.63 | ||||
| 0 | 7.79 a | 7.72 a | 7.60 a | 7.41 a | 7.67 a | |
| 5 | 9.10 a | 8.12 bc | 9.07 a | 8.37 b | 8.01 c | |
| 11 | 9.17 a | 8.39 b | 9.10 a | 8.60 c | 8.21 b | |
| 20 | 8.73 a | 8.43 b | 8.97 a | 8.45 b | 8.06 c | |
| 45 | 7.62 a | 8.50 b | 9.06 c | 9.02 c | 7.69 a | |
| Staphylococci | Meat batter | 3.57 ± 0.15 | ||||
| 0 | 5.57 a | 4.90 b | 5.24 a | 4.80 b | 5.07 b | |
| 5 | 5.50 a | 4.70 b | 5.20 c | 4.95 c | 5.01 c | |
| 11 | 6.30 a | 5.17 b | 6.15 a | 5.39 b | 5.19 b | |
| 20 | 6.00 a | 5.82 a | 6.21 a | 5.48 b | 5.27 b | |
| 45 | 6.30 a | 5.90 b | 6.85 c | 6.02 b | 5.25 d | |
| Enterobacteria | Meat batter | 2.12 ± 0.07 | ||||
| 0 | 2.01 a | 2.04 a | 1.98 a | 2.01 a | 2.00 a | |
| 5 | 2.24 a | 2.20 a | 2.19 a | 2.03 a | 1.70 a | |
| 11 | 1.92 a | 1.87 a | 1.87 a | 2.02 a | 1.72 a | |
| 20 | 1.69 a | 1.50 a | 1.84 a | 1.65 a | 1.75 a | |
| 45 | 2.02 a | 1.24 b | 2.10 a | 2.42 c | 1.59 d | |
| Enterococci | Meat batter | 2.09 ± 0.94 | ||||
| 0 | 1.96 a | 2.01 a | 2.00 a | 1.98 a | 1.94 a | |
| 5 | 2.47 a | 2.14 b | 2.69 a | 4.44 c | 2.82 a | |
| 11 | 2.43 a | 2.10 a | 3.14 b | 4.84 c | 2.84 b | |
| 20 | 2.29 a | 1.93 b | 4.33 c | 5.56 d | 3.51 e | |
| 45 | 2.96 a | 3.00 a | 4.14 b | 6.07 c | 3.17 a | |
| Yeasts | Meat batter | 1.69 ± 0.30 | ||||
| 0 | 2.10 a | 2.05 a | 2.03 a | 2.09 a | 2.10 a | |
| 5 | 1.80 a | 2.22 a | 1.79 a | 2.33 a | 1.73 a | |
| 11 | 3.22 a | 3.12 a | 3.30 a | 2.38 b | 2.91 a | |
| 20 | 2.56 ab | 2.90 a | 3.06 a | 2.39 b | 3.00 a | |
| 45 | 2.87 a | 2.83 a | 3.04 a | 3.22 a | 2.62 a | |
| Moulds | Meat batter | <1 * | ||||
| 0 | <1 a | <1 a | <1 a | <1 a | <1 a | |
| 5 | 1.58 a | 1.46 a | 0.49 b | 1.78 ac | 2.14 c | |
| 11 | 5.14 a | 5.18 a | 5.02 a | 4.70 b | 5.12 a | |
| 20 | 4.44 a | 5.11 b | 4.70 a | 4.62 a | 4.61 a | |
| 45 | 3.09 a | 4.19 b | 3.67 c | 4.19 b | 4.20 b | |
| Compound (%) | 20 Days | 45 Days | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | BPF2 | HNS55 | CB22 | HZK39 | Control | BPF2 | HNS55 | CB22 | HZK39 | |
| Nonanal | 0.68 a | 0.36 b | 0.40 b | 0.39 b | 0.32 b | 0.93 a | 0.74 b | 0.87 a | 1.07 a | 1.10 a |
| Benzaldehyde | 1.76 a | 1.12 b | 0.80 c | 0.68 c | 0.75 c | 2.85 a | 2.52 a | 0.83 b | 1.04 b | 0.91 b |
| Benzeneacetaldehyde | 10.40 a | 3.84 b | 1.45 c | 0.92 d | 1.04 cd | 18.60 a | 17.01 a | 3.58 b | 2.25 c | 6.68 d |
| Aldehydes | 12.83 a | 5.31 b | 2.65 c | 1.99 c | 2.11 c | 22.38 a | 20.27 a | 5.28 b | 4.37 b | 8.70 c |
| Acetone | 7.69 a | 6.06 b | 12.70 c | 7.96 a | 8.46 a | 12.38 a | 9.84 b | 14.86 c | 11.96 a | 11.14 a |
| 2-butanone | 4.90 a | 1.45 b | 2.94 c | 2.42 c | 1.79 b | 8.45 a | 8.06 a | 9.79 b | 25.16 | 19.81 c |
| 2-pentanone | 2.23 a | 2.50 a | 3.58 b | 4.16 c | 2.63 a | 2.77 a | 4.85 b | 3.55 c | 3.83 c | 2.48 a |
| Methyl isobutyl ketone | 3.95 a | 4.25 a | 4.74 b | 5.74 c | 4.61 b | 3.39 a | 2.15 b | 2.67 b | 1.94 b | 2.19 b |
| Acetoin | 7.25 a | 14.23 b | 3.53 c | 6.91 a | 4.80 c | 7.27 a | 6.54 a | 5.87 b | 6.40 a | 5.19 b |
| 2-nonanone | 0.93 a | 0.04 b | 0.19 b | 0.27 b | 0.03 b | 1.04 a | 0.00 b | 0.32 c | 0.54 c | 0.40 c |
| Ketones | 26.96 a | 28.53 a | 27.68 a | 27.46 a | 22.33 b | 35.31 a | 31.45 b | 37.06 a | 49.84 c | 41.21 d |
| Isopropyl alcohol | 6.82 a | 0.58 b | 0.73 b | 0.72 b | 0.65 b | 0.92 | 1.12 | 1.19 | 1.02 | 1.21 |
| Ethanol | 8.03 a | 5.21 b | 4.49 c | 3.97 c | 2.98 d | 1.77 a | 3.86 b | 3.42 b | 2.21 c | 2.86 c |
| 2-hexanol | 2.80 a | 2.82 a | 3.01 a | 4.00 b | 3.17 a | 2.25 a | 2.37 a | 0.96 b | 0.60 b | 0.95 b |
| 1-pentanol | 0.67 a | 0.29 b | 0.42 a | 0.55 a | 0.57 a | 0.35 a | 1.52 b | 0.59 a | 0.92 c | 0.53 a |
| 3-buten-1-ol, 3-methyl- | 0.41 a | 0.62 a | 0.66 a | 0.75 a | 0.55 a | 0.30 a | 0.26 a | 0.39 a | 0.26 a | 0.30 a |
| 2-buten-1-ol, 3-methyl- | 0.71 a | 0.54 a | 0.89 a | 0.77 a | 0.56 a | 0.48 a | 0.37 a | 0.42 a | 0.26 a | 0.24 a |
| 1-octen-3-ol | 0.71 a | 0.35 b | 0.37 b | 0.62 a | 0.37 a | 0.28 a | 0.74 b | 0.40 a | 0.78 b | 0.40 a |
| Alcohols | 20.15 a | 10.40 b | 10.56 b | 11.37 b | 8.85 c | 6.35 a | 10.25 b | 7.38 a | 6.04 a | 6.49 a |
| Acetic acid | 34.12 a | 48.81 b | 50.90 b | 52.06 b | 57.94 c | 29.34 a | 35.00 a | 44.77 b | 36.83 a | 39.92 ab |
| Propanoic acid | 0.31 a | 0.41 a | 0.77 b | 0.52 ab | 0.64 b | 0.51 | 0.28 | 0.55 | 0.30 | 0.39 |
| Butanoic acid | 3.22 a | 5.10 b | 5.68 b | 4.79 c | 6.25 d | 3.94 a | 1.91 b | 3.21 a | 1.36 c | 1.99 b |
| Hexanoic acid | 0.69 a | 0.96 b | 1.16 b | 1.06 b | 1.11 b | 0.69 a | 0.35 b | 0.81 a | 0.60 a | 0.58 a |
| Octanoic acid | 1.72 a | 0.48 b | 0.60 b | 0.75 b | 0.77 b | 1.49 a | 0.49 b | 0.94 c | 0.65 bc | 0.72 bc |
| Acids | 40.06 a | 55.76 b | 59.11 b | 59.18 b | 66.71 c | 35.97 a | 38.03 a | 50.28 b | 39.75 c | 43.61 d |
| Compound | Factor Coordinates of the Variables, Based on Correlations | |
|---|---|---|
| Factor 1 | Factor 2 | |
| Nonanal | 0.1709 | 0.0474 |
| Benzaldehyde | 0.1108 | −0.3169 |
| Benzeneacetaldehyde | 0.6580 | −0.0887 |
| Acetone | 1.5160 | −0.2415 |
| 2-butanone | 4.2191 | −0.1024 |
| 2-pentanone | 0.4112 | −0.0984 |
| Methyl isobutyl ketone | −0.1700 | −0.0860 |
| Acetoin | 0.3177 | 0.0267 |
| 2-nonanone | 0.0333 | −0.0016 |
| Isopropyl alcohol | −0.0828 | −0.0317 |
| Ethanol | −0.0913 | 1.7874 |
| 2-hexanol | −0.2204 | 0.0148 |
| 1-pentanol | 0.1322 | 0.1523 |
| 3-buten-1-ol, 3-methyl- | −0.0161 | 0.0509 |
| 2-buten-1-ol, 3-methyl- | −0.0330 | −0.1103 |
| 1-octen-3-ol | 0.0750 | 0.0474 |
| Acetic acid | 2.3249 | −0.3169 |
| Propanoic acid | 0.0093 | −0.0887 |
| Butanoic acid | −0.2668 | −0.2415 |
| Hexanoic acid | −0.0017 | −0.1024 |
| Octanoic acid | 0.0137 | −0.0984 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbieri, F.; Montanari, C.; Angelucci, C.; Gardini, F.; Tabanelli, G. Use of Indigenous Lactic Acid Bacteria for Industrial Fermented Sausage Production: Microbiological, Chemico-Physical and Sensory Features and Biogenic Amine Content. Fermentation 2024, 10, 507. https://doi.org/10.3390/fermentation10100507
Barbieri F, Montanari C, Angelucci C, Gardini F, Tabanelli G. Use of Indigenous Lactic Acid Bacteria for Industrial Fermented Sausage Production: Microbiological, Chemico-Physical and Sensory Features and Biogenic Amine Content. Fermentation. 2024; 10(10):507. https://doi.org/10.3390/fermentation10100507
Chicago/Turabian StyleBarbieri, Federica, Chiara Montanari, Chiara Angelucci, Fausto Gardini, and Giulia Tabanelli. 2024. "Use of Indigenous Lactic Acid Bacteria for Industrial Fermented Sausage Production: Microbiological, Chemico-Physical and Sensory Features and Biogenic Amine Content" Fermentation 10, no. 10: 507. https://doi.org/10.3390/fermentation10100507
APA StyleBarbieri, F., Montanari, C., Angelucci, C., Gardini, F., & Tabanelli, G. (2024). Use of Indigenous Lactic Acid Bacteria for Industrial Fermented Sausage Production: Microbiological, Chemico-Physical and Sensory Features and Biogenic Amine Content. Fermentation, 10(10), 507. https://doi.org/10.3390/fermentation10100507