
Additions to Dictyosporiaceae: Neoxylochrysis typhicola comb. et gen. nov., Two New Species and Four New Host Records from Medicinal Plants in Southwestern China

 , , , ,
, , , ,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimen Collection, Examination, and Single Spore Isolation
2.2. DNA Extraction, PCR Amplification and Sequencing
2.3. Sequence Alignment and Phylogenetic Analysis
3. Results
3.1. Phylogenetic Analysis
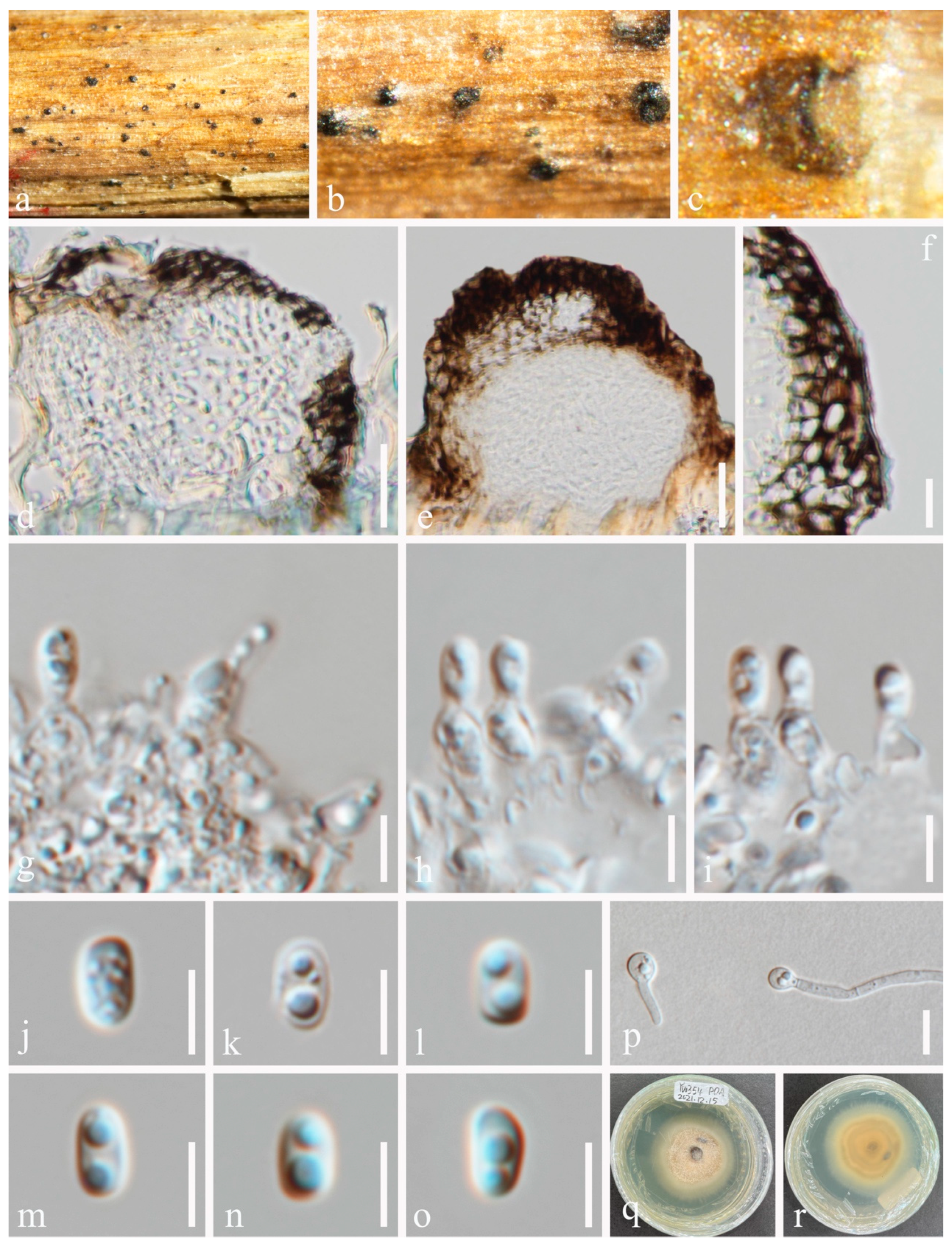
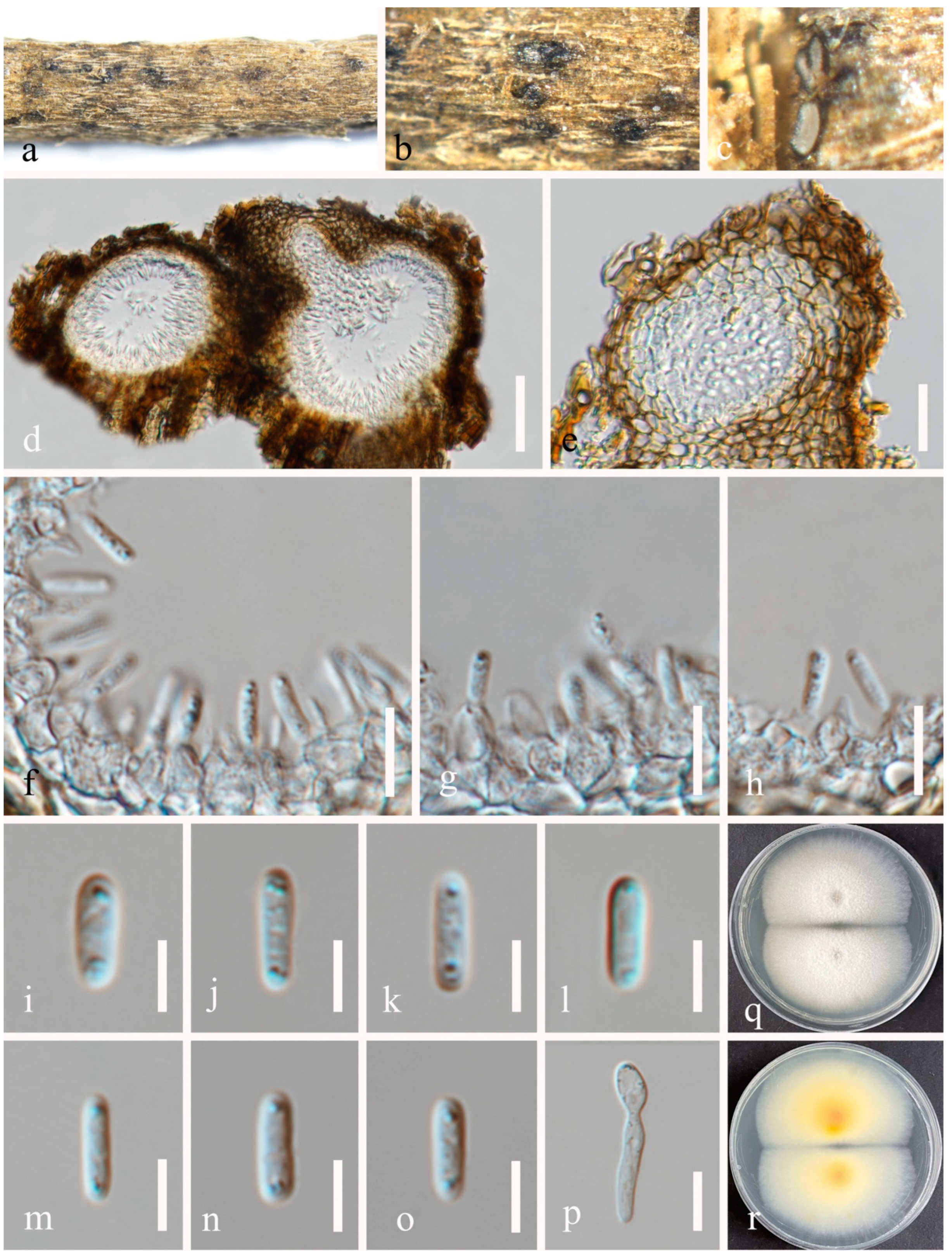
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, J.K.; Hyde, K.D.; Jones, E.B.G.; Ariyawansa, H.A.; Bhat, D.J.; Boonmee, S.; Maharachchikumbura, S.S.N.; McKenzie, E.H.C.; Phookamsak, R.; Phukhamsakda, C.; et al. Fungal diversity notes 1–110: Taxonomic and phylogenetic contributions to fungal species. Fungal Divers. 2015, 72, 1–197. [Google Scholar] [CrossRef]
- Boonmee, S.; D’souza, M.J.; Luo, Z.L.; Pinruan, U.; Tanaka, K.; Su, H.Y.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Taylor, J.E.; et al. Dictyosporiaceae fam. nov. Fungal Divers. 2016, 80, 457–482. [Google Scholar] [CrossRef]
- Kirk, P.M. New or interesting microfungi II. Dematiaceous hyphomycetes from Esher Common, Surrey. Trans. Br. Mycol. Soc. 1981, 77, 279–297. [Google Scholar] [CrossRef]
- Cai, L.; Guo, X.Y.; Hyde, K.D. Morphological and molecular characterisation of a new anamorphic genus Cheirosporium from freshwater in China. Persoonia 2008, 20, 53–58. [Google Scholar] [CrossRef]
- Li, W.L.; Luo, Z.L.; Liu, J.K.; Bhat, D.J.; Bao, D.F.; Su, H.Y.; Hyde, K.D. Lignicolous freshwater fungi from China I: Aquadictyospora lignicola gen. et sp. nov. and new record of Pseudodictyosporium wauense from northwestern Yunnan Province. Mycosphere 2017, 8, 1587–1597. [Google Scholar] [CrossRef]
- Shen, H.W.; Bao, D.F.; Wanasinghe, D.N.; Boonmee, S.; Liu, J.K.; Luo, Z.L. Novel species and records of Dictyosporiaceae from freshwater habitats in China and Thailand. J. Fungi 2022, 8, 1200. [Google Scholar] [CrossRef]
- Tian, W.H.; Chen, Y.P.; Maharachchikumbura, S.S.N. Neodigitodesmium, a novel genus of family Dictyosporiaceae from Sichuan Province, China. Phytotaxa 2022, 559, 176–184. [Google Scholar] [CrossRef]
- Barr, M.E. New taxa and combinations in the Loculoascomycetes. Mycotaxon 1987, 29, 501–505. [Google Scholar]
- Tanaka, K.; Hirayama, K.; Yonezawa, H.; Sato, G.; Toriyabe, A.; Kudo, H.; Hashimoto, A.; Matsumura, M.; Harada, Y.; Kurihara, Y.; et al. Revision of the Massarineae (Pleosporales, Dothideomycetes). Stud. Mycol. 2015, 82, 75–136. [Google Scholar] [CrossRef] [PubMed]
- Crous, P.W.; Schumache, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfeld, M.J.; Summerel, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. 2. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Piątek, M.; Rodriguez-Flakus, P.; Domic, A.; Palabral-Aguilera, A.N.; Gómez, M.I.; Flakus, A. Phylogenetic placement of Leptosphaeria polylepidis, a pathogen of Andean endemic Polylepis tarapacana, and its newly discovered mycoparasite Sajamaea mycophila gen. et sp. nov. Mycol. Prog. 2020, 19, 1–14. [Google Scholar] [CrossRef]
- Atienza, V.; Hawksworth, D.L.; Pérez-Ortega, S. Verrucoccum (Dothideomycetes, Dictyosporiaceae), a new genus of lichenicolous fungi on Lobaria s. lat. for the Dothidea hymeniicola species complex. Mycologia 2021, 113, 1233–1252. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.B.; Jeewon, R.; Karunarathna, S.C.; Phukhamsakda, C.; Doilom, M.; Kakumyan, P.; Suwannarach, N.; Phookamsak, R.; Lumyong, S. Reappraisal of Immotthia in Dictyosporiaceae, Pleosporales: Introducing Immotthia bambusae sp. nov. and Pseudocyclothyriella clematidis comb. et gen. nov. based on morphology and phylogeny. Front. Microbiol. 2021, 12, 656235. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.F.; Liu, J.K.; Hyde, K.D.; Chen, Y.Y.; Ran, H.Y.; Liu, Z.Y. Ascomycetes from karst landscapes of Guizhou Province, China. Fungal Divers. 2023, 122, 1–160. [Google Scholar] [CrossRef]
- Jayasiri, S.C.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Jeewon, R.; Phillips, A.J.L.; Bhat, D.J.; Wanasinghe, D.N.; Liu, J.K.; Lu, Y.Z.; et al. Diversity, morphology and molecular phylogeny of Dothideomycetes on decaying wild seed pods and fruits. Mycosphere 2019, 10, 1–186. [Google Scholar] [CrossRef]
- Lu, L.; Karunarathna, S.C.; Dai, D.Q.; Xiong, Y.R.; Suwannarach, N.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; Jayawardena, R.S.; Tibpromma, S. Description of four novel species in Pleosporales associated with coffee in Yunnan, China. J. Fungi 2022, 8, 1113. [Google Scholar] [CrossRef] [PubMed]
- Boerema, G.H.; de Gruyter, J.; Noordeloos, M.E.; Hamers, M.E.C. Phoma Identification Manual: Differentiation of Specific and Infra-specific Taxa in Culture; CABI: Wallingford, UK, 2004; p. 470. [Google Scholar]
- Tennakoon, D.S.; Bhat, D.J.; Kuo, C.H.; Hyde, K.D. Leaf litter saprobic Dictyosporiaceae (Pleosporales, Dothideomycetes): Pseudocoleophoma zingiberacearum sp. nov. from Hedychium coronarium. Kavaka 2019, 53, 1–7. [Google Scholar] [CrossRef]
- Li, W.J.; McKenzie, E.H.C.; Liu, J.K.; Bhat, D.J.; Dai, D.Q.; Camporesi, E.; Tian, Q.; Maharachchikumbura, S.S.N.; Luo, Z.L.; Shang, Q.J.; et al. Taxonomy and phylogeny of hyaline-spored coelomycetes. Fungal Divers. 2020, 100, 279–801. [Google Scholar] [CrossRef]
- Kularathnage, N.D.; Wanasinghe, D.N.; Senanayake, I.C.; Yang, Y.H.; Manawasinghe, I.S.; Phillips, A.J.L.; Hyde, K.D.; Dong, W.; Song, J.G. Microfungi associated with ornamental palms: Byssosphaeria phoenicis sp. nov. (Melanommataceae) and Pseudocoleophoma rhapidis sp. nov. (Dictyosporiaceae) from south China. Phytotaxa 2022, 568, 149–169. [Google Scholar] [CrossRef]
- Phukhamsakda, C.; McKenzie, E.H.C.; Phillips, A.J.L.; Gareth Jones, E.B.; Jayarama Bhat, D.; Stadler, M.; Bhunjun, C.S.; Wanasinghe, D.N.; Thongbai, B.; Camporesi, E.; et al. Microfungi associated with Clematis (Ranunculaceae) with an integrated approach to delimiting species boundaries. Fungal Divers. 2020, 102, 1–203. [Google Scholar] [CrossRef]
- Ranojevic, N. Dritter Beitrag zur Pilzflora Serbiens. Ann. Mycol. 1914, 12, 393–421. [Google Scholar]
- Ellis, M.B. Dematiaceous Hyphomycetes; CABI: Wallingford, UK, 1971. [Google Scholar]
- Crous, P.W.; Shivas, R.G.; Quaedvlieg, W.; van der Bank, M.; Zhang, Y.; Summerell, B.A.; Guarro, J.; Wingfield, M.J.; Wood, A.R.; Alfenas, A.C.; et al. Fungal Planet description sheets: 214–280. Persoonia 2014, 32, 184–306. [Google Scholar] [CrossRef]
- Prasher, I.B.; Verma, R.K. Two new species of Dictyosporium from India. Phytotaxa 2015, 204, 193–202. [Google Scholar] [CrossRef]
- Silva, C.R.; Gusmão, L.F.P.; Castaneda-Ruiz, R.F. Dictyosporium amoenum sp. nov. from Chapada Diamantina, Bahia, Brazil. Mycotaxon 2016, 130, 1125–1133. [Google Scholar] [CrossRef]
- Tennakoon, D.S.; de Silva, N.I.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Kumla, J.; Suwannarach, N.; Lumyong, S. Exploring more on Dictyosporiaceae: The species geographical distribution and intriguing novel additions from plant litter. Diversity 2023, 15, 410. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Rathnayaka, A.R.; Marasinghe, D.S.; Calabon, M.S.; Gentekaki, E.; Lee, H.B.; Hurdeal, V.G.; Pem, D.; Dissanayake, L.S.; Wijesinghe, S.N.; et al. Morphological approaches in studying fungi: Collection, examination, isolation, sporulation and preservation. Mycosphere 2020, 11, 2678–2754. [Google Scholar] [CrossRef]
- Chomnunti, P.; Hongsanan, S.; Hudson, B.A.; Tian, Q.; Peršoh, D.; Dhami, M.K.; Alias, A.S.; Xu, J.C.; Liu, X.Z.; Stadler, M.; et al. The sooty moulds. Fungal Divers. 2014, 66, 1–36. [Google Scholar] [CrossRef]
- Rayner, R.W. A Mycological Colour Chart; Commonwealth Mycological Institute and British Mycological Society: Kew, UK, 1970. [Google Scholar]
- Jayasiri, S.C.; Hyde, K.D.; Ariyawansa, H.A.; Bhat, J.; Buyck, B.; Cai, L.; Dai, Y.C.; Abd-Elsalam, K.A.; Ertz, D.; Hidayat, I.; et al. The Faces of Fungi database: Fungal names linked with morphology, phylogeny and human impacts. Fungal Divers. 2015, 74, 3–18. [Google Scholar] [CrossRef]
- Wu, Z.H.; Wang, T.H.; Huang, W.; Qu, Y.B. A simplified method for chromosome DNA preparation from filamentous fungi. Mycosystema 2001, 20, 575–577. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- Rehner, S.A.; Buckley, E. A Beauveria phylogeny inferred from nuclear ITS and EF1-alpha sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- GenBank. Available online: http://www.ncbi.nlm.nih.gov (accessed on 12 July 2023).
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Dissanayake, A.J.; Bhunjun, C.S.; Maharachchikumbura, S.S.N.; Liu, J.K. Applied aspects of methods to infer phylogenetic relationships amongst fungi. Mycosphere 2020, 11, 2653–2677. [Google Scholar] [CrossRef]
- Nylander, J. MrModeltest (Version 2.2); Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Silvestro, D.; Michalak, I. raxmlGUI: A graphical front-end for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree 1.4.4. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree (accessed on 11 July 2024).
- Figshare. Available online: https://figshare.com/account/home#/data (accessed on 23 June 2024).
- Reisinger, O. Remarques sur les genres Dendryphiella et Dendryphion. Bull. De La Société Mycol. De Fr. 1968, 84, 27–51. [Google Scholar]
- Dela Cruz, T.; Edison, E. Marine Dendryphiella Species from Different Geographical Locations: An Integrated, Polyphasic Approach to Its Taxonomy and Physioecology. 2006. Available online: https://nbn-resolving.org/urn:nbn:de:gbv:084-10363 (accessed on 14 October 2020).
- Chethana, K.W.T.; Niranjan, M.; Dong, W.; Samarakoon, M.C.; Bao, D.F.; Calabon, M.S.; Chaiwan, N.; Chuankid, B.; Dayarathne, M.C.; de Silva, N.I.; et al. AJOM new records and collections of fungi: 101–150. Asian J. Mycol. 2021, 4, 113–260. [Google Scholar] [CrossRef]
- Hyde, K.D.; Hongsanan, S.; Jeewon, R.; Bhat, D.J.; McKenzie, E.H.C.; Jones, E.B.G.; Phookamsak, R.; Ariyawansa, H.A.; Boonmee, S.; Zhao, Q.; et al. Fungal diversity notes 367–490: Taxonomic and phylogenetic contributions to fungal taxa. Fungal Divers. 2016, 80, 1–270. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, M.; Bussmann, W.R.; Liu, H.M.; Liu, Y.Y.; Peng, Y.D.; Zu, K.L.; Zhao, Y.M.; Liu, Z.B.; Yu, S.X. Species richness and conservation gap analysis of karst areas: A case study of vascular plants from Guizhou, China. Glob. Ecol. Conserv. 2018, 16, e00460. [Google Scholar] [CrossRef]
- Shan, Z.J.; Ye, J.F.; Hao, D.C.; Xiao, P.G.; Chen, Z.D.; Lu, A.M. Distribution patterns and industry planning of commonly used traditional Chinese medicinal plants in China. Plant Divers. 2022, 44, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Z.; Huang, X.L.; Tong, B.L.; Wang, D.; Liu, J.M.; Liao, X.F.; Sun, Q.W. Effects of rhizosphere fungi on the chemical composition of fruits of the medicinal plant Cinnamomum migao endemic to southwestern China. BMC Microbiol. 2021, 21, 206. [Google Scholar] [CrossRef]
- Du, T.Y.; Karunarathna, S.C.; Zhang, X.; Dai, D.Q.; Mapook, A.; Suwannarach, N.; Xu, J.C.; Stephenson, S.L.; Elgorban, A.M.; Al-Rejaie, S.; et al. Endophytic fungi associated with Aquilaria sinensis (Agarwood) from China show antagonism against bacterial and fungal pathogens. J. Fungi 2022, 8, 1197. [Google Scholar] [CrossRef] [PubMed]
- Niu, X.P.; Gao, H.; Qi, J.M.; Chen, M.C.; Tao, A.; Xu, J.T.; Dai, Z.G.; Su, J.G. Colletotrichum species associated with jute (Corchorus capsularis L.) anthracnose in southeastern China. Sci. Rep. 2016, 6, 25179. [Google Scholar] [CrossRef] [PubMed]
- Petrasch, S.; Knapp, S.J.; van Kan, J.A.L.; Blanco-Ulate, B. Grey mould of strawberry, a devastating disease caused by the ubiquitous necrotrophic fungal pathogen Botrytis cinerea. Mol. Plant Pathol. 2019, 20, 877–892. [Google Scholar] [CrossRef]
- Yang, H.Z.; He, R.Y.; Cui, Y.; Li, Y.; Ge, X. Saprophytic Bacillus accelerates the release of effective components in agarwood by degrading cellulose. Molecules 2022, 27, 1428. [Google Scholar] [CrossRef]
- Du, H.Z.; Lu, Y.H.; Cheewangkoon, R.; Liu, J.K. Morpho-phylogenetic evidence reveals novel species and new records of Nigrograna(Nigrogranaceae) associated with medicinal plants in Southwestern China. MycoKeys 2024, 110, 1–33. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Strain/Specimen Number | GenBank Accession Number | |||
|---|---|---|---|---|---|
| ITS | LSU | SSU | TEF1-α | ||
| Aquadictyospora clematidis | MFLUCC 17-2080 T | MT310592 | MT214545 | MT226664 | MT394727 |
| Aquadictyospora lignicola | MFLUCC 17-1318 T | MF948621 | MF948629 | N/A | MF953164 |
| Aquadictyospora lignicola | UESTCC 23.0213 | PP925610 | PP925664 | PP925653 | PP926517 |
| Aquaticheirospora lignicola | HKUCC10304 T | AY864770 | AY736378 | AY736377 | N/A |
| Cheirosporium triseriale | HMAS 180703 T | EU413953 | EU413954 | N/A | N/A |
| Dendryphiella eucalyptorum | CBS 137987 T | KJ869139 | KJ869196 | N/A | N/A |
| Dendryphiella eucalyptorum | UESTCC 23.0214 | PP925612 | PP925666 | PP925655 | PP926520 |
| Dendryphiella eucalyptorum | UESTCC 23.0215 | PP925613 | PP925667 | PP925656 | PP926518 |
| Dendryphiella eucalyptorum | UESTCC 23.0216 | PP925614 | PP925668 | PP925657 | N/A |
| Dendryphiella phitsanulokensis | MFLUCC 17-2513 T | MG754400 | MG754401 | MG754402 | N/A |
| Dendryphiella trisepta | COAD 2388 T | MK278898 | MK277357 | N/A | N/A |
| Dendryphiella vinosa | MFLU 20-0444 | MT907477 | MT907480 | N/A | N/A |
| Dendryphiella vinosa | UESTCC 23.0217 | PP925611 | PP925665 | PP925654 | PP926519 |
| Dictyocheirospora bannica | KH 332 T | NR_154039 | NG_059061 | NG_064841 | AB808489 |
| Dictyocheirospora cheirospora | KUMCC 17-0035 T | MF177035 | MF177036 | MF928073 | N/A |
| Dictyocheirospora gigantica | BCC 11346 | DQ018095 | N/A | N/A | N/A |
| Dictyocheirospora alangii | CGMCC 3.25622 | PP925607 | PP925662 | PP925652 | PP926516 |
| Dictyocheirospora multiappendiculata | KUNCC 22-10734 T | OP526632 | OP526642 | OP526623 | OP542234 |
| Dictyocheirospora rotunda | MFLUCC 14-0293 T | KU179099 | KU179100 | KU179101 | N/A |
| Dictyocheirospora rotunda | MFLU 18-1041 | MH381764 | MH381773 | MH381758 | MH388818 |
| Dictyocheirospora rotunda | UESTCC 23.0219 | PP925608 | PP925663 | N/A | N/A |
| Dictyocheirospora rotunda | UESTCC 24.0183 | PP925609 | PP925661 | PP925651 | PP926515 |
| Dictyocheirospora suae | KUNCC 22-12424 T | OP526631 | OP526641 | OP526622 | OP542233 |
| Dictyocheirospora subramanianii | BCC 3503 | DQ018094 | N/A | N/A | N/A |
| Dictyocheirospora xishuangbannaensis | KUMCC 17-0181 T | MH388342 | MH376714 | MH388310 | MH388377 |
| Dictyosporium elegans | NBRC 32502 T | DQ018087 | DQ018100 | DQ018079 | N/A |
| Dictyosporium olivaceosporum | KH 375 T | LC014542 | AB807514 | AB797224 | AB808490 |
| Digitodesmium bambusicola | CBS 110279 T | DQ018091 | DQ018103 | N/A | N/A |
| Gregarithecium curvisporum | KT 922 T | AB809644 | AB807547 | AB797257 | AB808523 |
| Immotthia bambusae | KUN-HKAS 112012AI T | MW489455 | MW489450 | MW489461 | MW504646 |
| Jalapriya pulchra | MFLUCC 17-1683 | MF948628 | MF948636 | N/A | MF953171 |
| Jalapriya toruloides | CBS 209.65 | DQ018093 | DQ018104 | DQ018081 | N/A |
| Neodendryphiella mali | FMR 16561 T | LT906655 | LT906657 | N/A | N/A |
| Neodigitodesmium cheirosporum | UESTCC 22.0020 T | ON595714 | ON595713 | ON595712 | ON595700 |
| Neoxylochrysis typhicola | MFLUCC 16-0123 T | KX576655 | KX576656 | N/A | N/A |
| Neoxylochrysis typhicola | CGMCC 3.25688 | PP925617 | PP925671 | PP925660 | PP926523 |
| Periconia igniaria | CBS 845.96 | LC014586 | AB807567 | AB797277 | AB808543 |
| Periconia igniaria | CBS 379.86 | LC014585 | AB807566 | AB797276 | AB808542 |
| Pseudocoleophoma bauhiniae | MFLUCC 17-2586 T | MK347736 | MK347953 | MK347844 | MK360076 |
| Pseudocoleophoma bauhiniae | MFLUCC 17-2280 | MK347735 | MK347952 | MK347843 | MK360075 |
| Pseudocoleophoma calamagrostidis | KT 3284 T | LC014592 | LC014609 | LC014604 | LC014614 |
| Pseudocoleophoma flavescens | CBS 178.93 | N/A | GU238075 | GU238216 | N/A |
| Pseudocoleophoma polygonicola | KT 731 T | AB809634 | AB807546 | AB797256 | AB808522 |
| Pseudocoleophoma puerensis | ZHKUCC 22-0204 T | OP297799 | OP297769 | OP297783 | OP321568 |
| Pseudocoleophoma puerensis | ZHKUCC 22-0205 | OP297800 | OP297770 | OP297784 | OP321569 |
| Pseudocoleophoma rhapidis | ZHKUCC 21-0124 T | ON244664 | ON244661 | ON244667 | ON243581 |
| Pseudocoleophoma rhapidis | ZHKUCC 22-0004 | ON244665 | ON244662 | ON244668 | ON243582 |
| Pseudocoleophoma rosae | CGMCC 3.25623 T | PP925615 | PP925669 | PP925658 | PP926521 |
| Pseudocoleophoma rosae | UESTCC 24.0184 | PP925616 | PP925670 | PP925659 | PP926522 |
| Pseudocoleophoma rusci | MFLUCC 16-1444 T | MT185549 | MT183514 | MT214983 | N/A |
| Pseudocoleophoma yunnanensis | ZHKUCC 22-0200 T | OP297795 | OP297765 | OP297779 | OP321564 |
| Pseudocoleophoma yunnanensis | ZHKUCC 22-0201 | OP297796 | OP297766 | OP297780 | OP321565 |
| Pseudocoleophoma zingiberacearum | NCYUCC 19-0052 T | MN615939 | MN616753 | N/A | MN629281 |
| Pseudocoleophoma zingiberacearum | NCYUCC 19-0053 | MN615940 | MN616754 | N/A | MN629282 |
| Pseudocoleophoma zingiberacearum | NCYUCC 19-0054 | MN615941 | MN616755 | N/A | MN629283 |
| Pseudoconiothyrium broussonetiae | CBS 145036 T | MK442618 | MK442554 | N/A | N/A |
| Pseudocyclothyriella clematidis | MFLUCC 17-2177A T | MT310595 | MT214548 | MT226667 | MT394730 |
| Pseudocyclothyriella clematidis | MFLU 16-0280 | MT310596 | MT214549 | N/A | N/A |
| Pseudodictyosporium elegans | CBS 688.93 T | DQ018099 | DQ018106 | DQ018084 | N/A |
| Pseudodictyosporium thailandica | MFLUCC 16-0029 T | KX259520 | KX259522 | KX259524 | KX259526 |
| Sajamaea mycophila | APA-2999 T | MK795715 | MK795718 | N/A | N/A |
| Verrucoccum spribillei | SPO2343 T | MT918780 | MT918765 | MT918773 | N/A |
| Vikalpa australiense | HKUCC 8797 | DQ018092 | N/A | N/A | N/A |
| Species | Morphology of Conidia | Conidiomata | Conidiogenous Cells | Conidia | References |
|---|---|---|---|---|---|
| P. bauhiniae |  | 130–150 × 90–115 μm, immersed to superficial, subglobose, dark brown, multiloculate | 2.5–5.5 × 2–3 μm, phialidic, hyaline, aseptate, doliiform to lageniform | 7.5–11 × 2–3 μm, hyaline, oblong to ellipsoidal, aseptate, with guttules | [15] |
| P. calamagrostidis |  | (150–)250–500 × 220–300 μm, immersed to erumpent, depressed globose, glabrous | 5–9 × 2–4 μm, hyaline, doliiform to subglobose | 6–10 × 2–2.5 μm, hyaline, cylindrical, aseptate, with guttules at each end | [9] |
| P. flavescens | 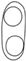 | 20–140 μm diam., globose, glabrous or covered by hyphae, solitary or confluent | 4–6 × 3–6 μm, globose to doliiform | 4–7 × 2–3.5 μm, hyaline, ellipsoidal, aseptate, with 2 very large polar guttules | [17,19] |
| P. polygonicola |  | 170–250 μm diam., superficial, ampulliform, glabrous, uniloculate | 7–17 × 3.5–5 μm, hyaline, aseptate, doliiform to lageniform | (9–)11.5–18(–21.5) × 3–4.5 μm, hyaline, cylindrical, aseptate | [9] |
| P. rhapidis |  | 150–225 × 225–300 μm, immersed, subglobose, black, uniloculate | 7–10 × 13–17 μm, phialidic, hyaline, doliiform | 20–25 × 10–15 μm, hyaline, oblong to obovoid, aseptate | [20] |
| P. rosae |  | 100–184 × 98–215 μm, semi-immersed, solitary to gregarious, globose to subglobose, pyriform or irregular, unilocular | 3–5 × 4–7 μm, phialidic, aseptate, smooth-walled, hyaline | 6–9 × 2–4 μm, hyaline, aseptate, oblong to cylindrical, thin-walled, guttules concentrated at the ends | This study |
| P. rusci |  | 130–200 × 250–330 μm, deeply immersed, globose, subglobose or ovoid, brown, glabrous, unilocular | 4–9 × 3–7 μm, phialidic, enteroblastic, hyaline, doliiform, ampulliform to subcylindrical | 8–14 × 3–6 μm, hyaline, cylindrical or fusiform, with a rounded apex and a slightly narrow truncate base | [19] |
| P. zingiberacearum |  | 200–220 × 110–150 μm, immersed, multi-loculate, depressed globose, glabrous, non-ostiolate | 1.5–2.5 × 1–1.5 μm, phialidic, doliiform to lageniform, hyaline, aseptate | 12–14 × 2–3 μm, solitary, hyaline, aseptate, oblong to ellipsoidal, with rounded to obtuse ends, with guttules | [18] |
| Neoxylochrysis typhicola (P. typhicola) |  | 60–100 × 140–150 μm, semi-erumpent, uniloculate solitary to scattered, subglobose | 2–5 × 2–5 μm, enteroblastic, smooth-walled, hyaline | 9–11 × 2–3 μm, hyaline, oblong to cylindrical, with rounded or obtuse ends, septate, guttulate | [50], this study |
| Species | Morphology of Ascospores | Ascomata | Asci | Ascospores | Reference |
|---|---|---|---|---|---|
| P. bauhiniae |  | 100–120 × 125–145 μm, immersed, subglobose to obpyriform, dark brown, coriaceous | 65–80 × 5–8 μm, 8-spored, clavate to cylindric-clavate, short-pedicellate with an ocular chamber | 17–20 × 3.5–4.5 μm, hyaline, cylindric-fusiform, 1–3-septate, 3–4-guttulate, without appendages and sheath | [15] |
| P. calamagrostidis |  | 160–220 × 140–200 μm, immersed, globose to depressed globose | 62.5–80 × 7.5–10 μm, 8-spored, cylindrical, rounded at the apex, short-stalked | (14.5–)16–19(–21) × 3–4.5 μm, hyaline, fusiform, 1-septate, 3–4-guttulate, with an entire sheath | [9] |
| P. polygonicola |  | 280–350 × 230–310 μm, immersed to erumpent | (67–)74–90(–100) × 9–12.5 μm, cylindrical to clavate | (17.5–)19–23(–25) × 4–6 μm, hyaline, fusiform, 1-septate, 4–6-guttulate, surrounded by a sheath | [9] |
| P. puerensis |  | 150–300 × 170–220 μm, immersed to erumpent, brown to black, globose or subglobose | 50–70 × 7–11 μm, 8-spored, cylindrical, long-stalked with club-like pedicel | 10–15 × 3.5–6 μm, hyaline to brown, narrowly ellipsoid or oblong, 1–3 thick and dark eusepta, normally 4-guttulate, without a sheath | [16] |
| P. rosae |  | 170–220 × 170–220 μm, semi-immersed or immersed, dark brown, globose to subglobose, unilocular, glabrous, thick-walled, thickened at the apex, visible as black dots or papilla on the host | 70–88 × 8–12 μm, 8-spored, cylindrical to clavate, some slightly curved, with an ocular chamber, short-stalked with club-shaped pedicel | 12–21 × 3–6 μm, hyaline, smooth-walled, fusiform with acute ends, 1-septate, slightly constricted at the septum, and occasionally 2–4-guttulate when young, surrounded by a mucilaginous sheath | This study |
| P. yunnanensis |  | 160–280 × 200–280 μm, semi-immersed to erumpent, dark brown to black, subglobose to obpyriform, solitary or scattered, coriaceous | 65–90 × 8–11 μm, 8-spored, clavate to cylindrical, short-stalked with club-shaped pedicel | 16–26 × 4–8 μm, hyaline, fusiform, 1-septate, 4-guttulate, with a distinct sheath | [16] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, N.; Du, H.-Z.; Chethana, K.W.T.; Khongphinitbunjong, K.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Liu, J.-K. Additions to Dictyosporiaceae: Neoxylochrysis typhicola comb. et gen. nov., Two New Species and Four New Host Records from Medicinal Plants in Southwestern China. J. Fungi 2024, 10, 872. https://doi.org/10.3390/jof10120872
Wu N, Du H-Z, Chethana KWT, Khongphinitbunjong K, Maharachchikumbura SSN, Hyde KD, Liu J-K. Additions to Dictyosporiaceae: Neoxylochrysis typhicola comb. et gen. nov., Two New Species and Four New Host Records from Medicinal Plants in Southwestern China. Journal of Fungi. 2024; 10(12):872. https://doi.org/10.3390/jof10120872
Chicago/Turabian StyleWu, Na, Hong-Zhi Du, Kandawatte Wedaralalage Thilini Chethana, Kitiphong Khongphinitbunjong, Sajeewa S. N. Maharachchikumbura, Kevin D. Hyde, and Jian-Kui Liu. 2024. "Additions to Dictyosporiaceae: Neoxylochrysis typhicola comb. et gen. nov., Two New Species and Four New Host Records from Medicinal Plants in Southwestern China" Journal of Fungi 10, no. 12: 872. https://doi.org/10.3390/jof10120872
APA StyleWu, N., Du, H.-Z., Chethana, K. W. T., Khongphinitbunjong, K., Maharachchikumbura, S. S. N., Hyde, K. D., & Liu, J.-K. (2024). Additions to Dictyosporiaceae: Neoxylochrysis typhicola comb. et gen. nov., Two New Species and Four New Host Records from Medicinal Plants in Southwestern China. Journal of Fungi, 10(12), 872. https://doi.org/10.3390/jof10120872