Beyond Melanin: Proteomics Reveals Virulence-Related Proteins in Paracoccidioides brasiliensis and Paracoccidioides lutzii Yeast Cells Grown in the Presence of L-Dihydroxyphenylalanine

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Maintenance
2.2. Melanin Production
2.3. Protein Extraction
2.4. Sample Processing for Shotgun Proteomics
2.5. Mass Spectrometry Analysis
2.6. Protein Identification and Quantitation Following Mass Spectrometry
2.7. Bioinformatic Analyses
3. Results
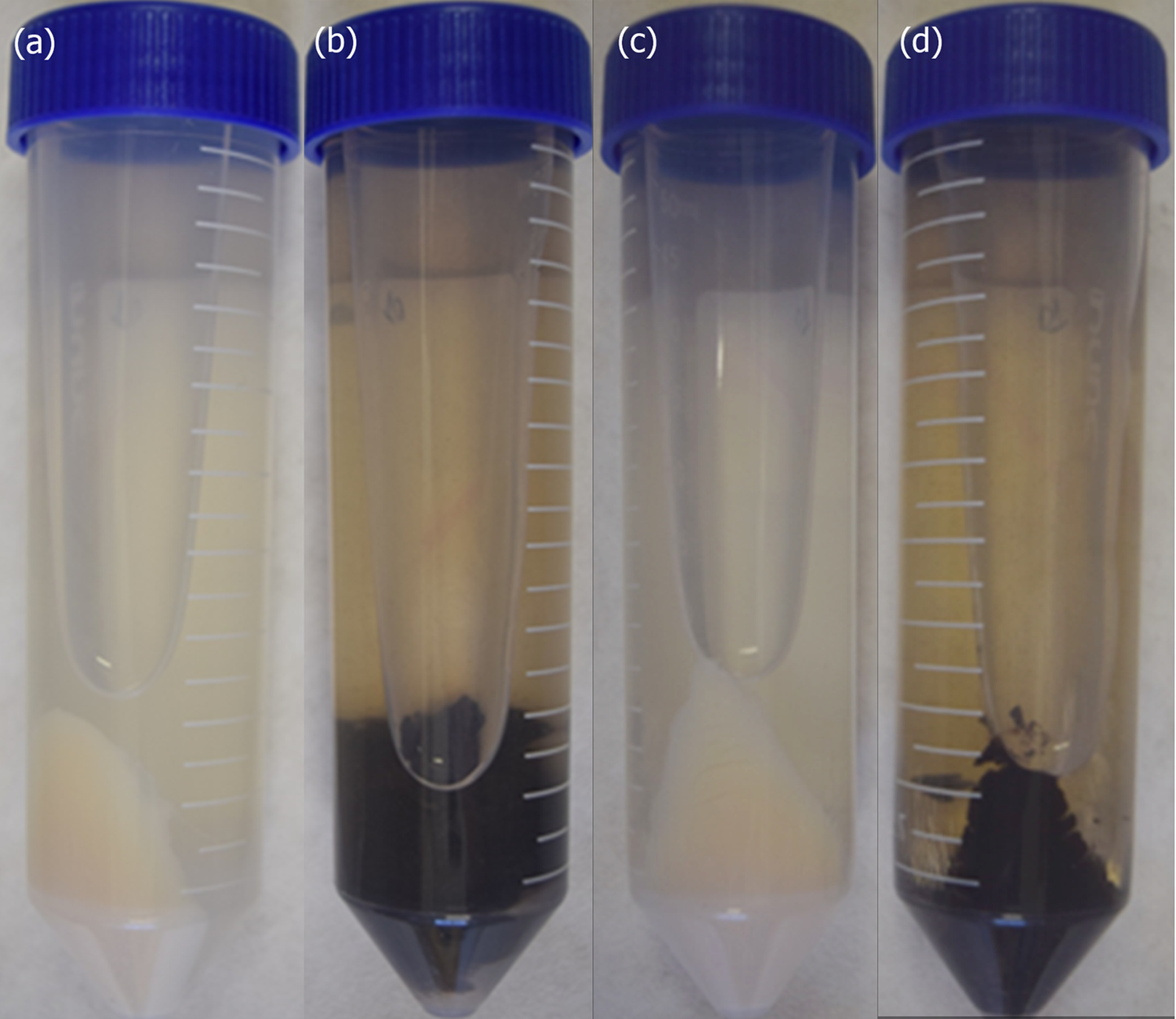
3.1. Fungal Melanization and Protein Extract Control
3.2. Proteomic Analysis of Melanized and Non-Melanized Cells
3.3. Enriched Pathways
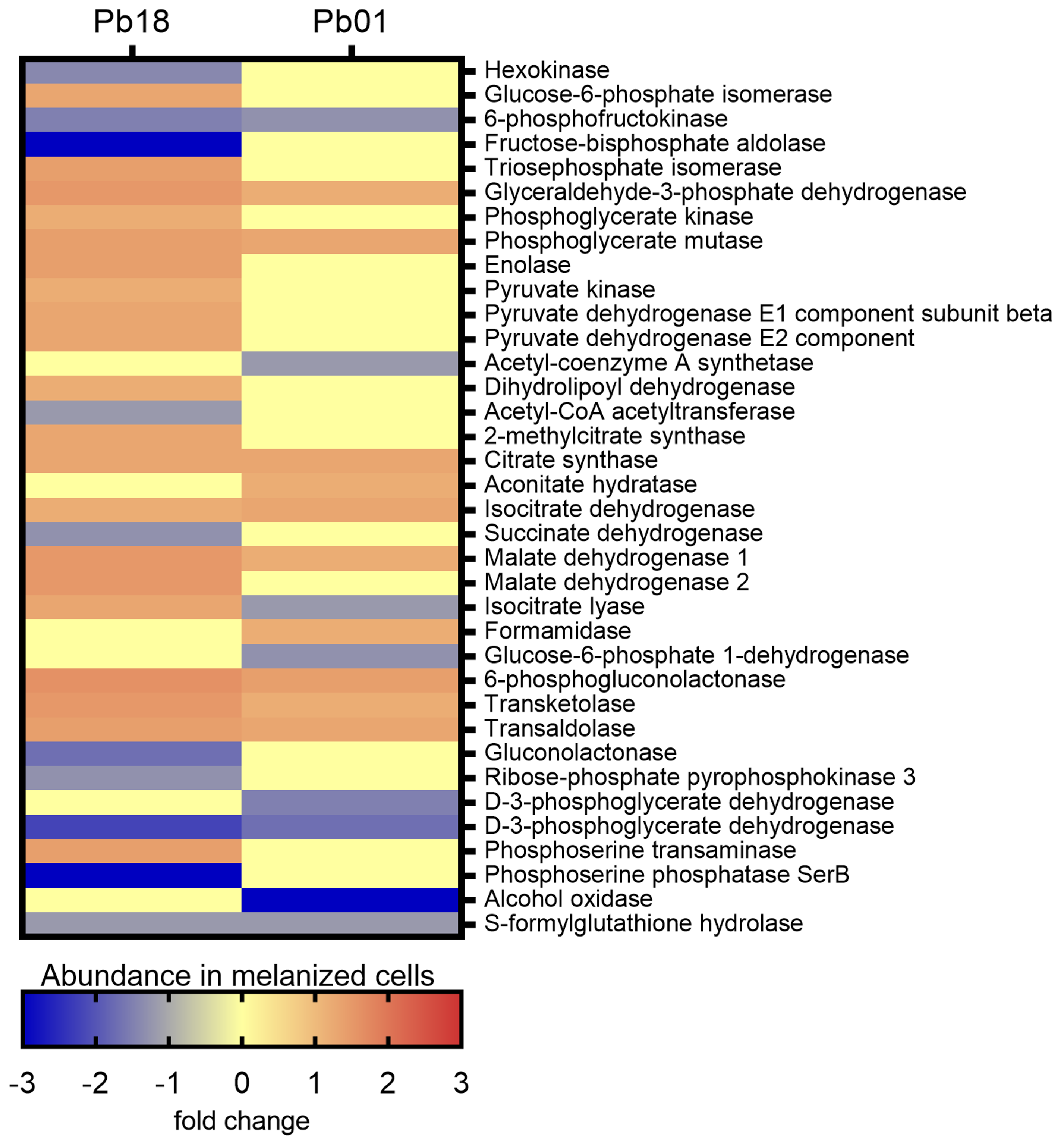
3.3.1. Carbon Metabolism
3.3.2. Nucleotide Metabolism
3.3.3. Amino Acid Metabolism
3.3.4. Other Metabolic Pathways
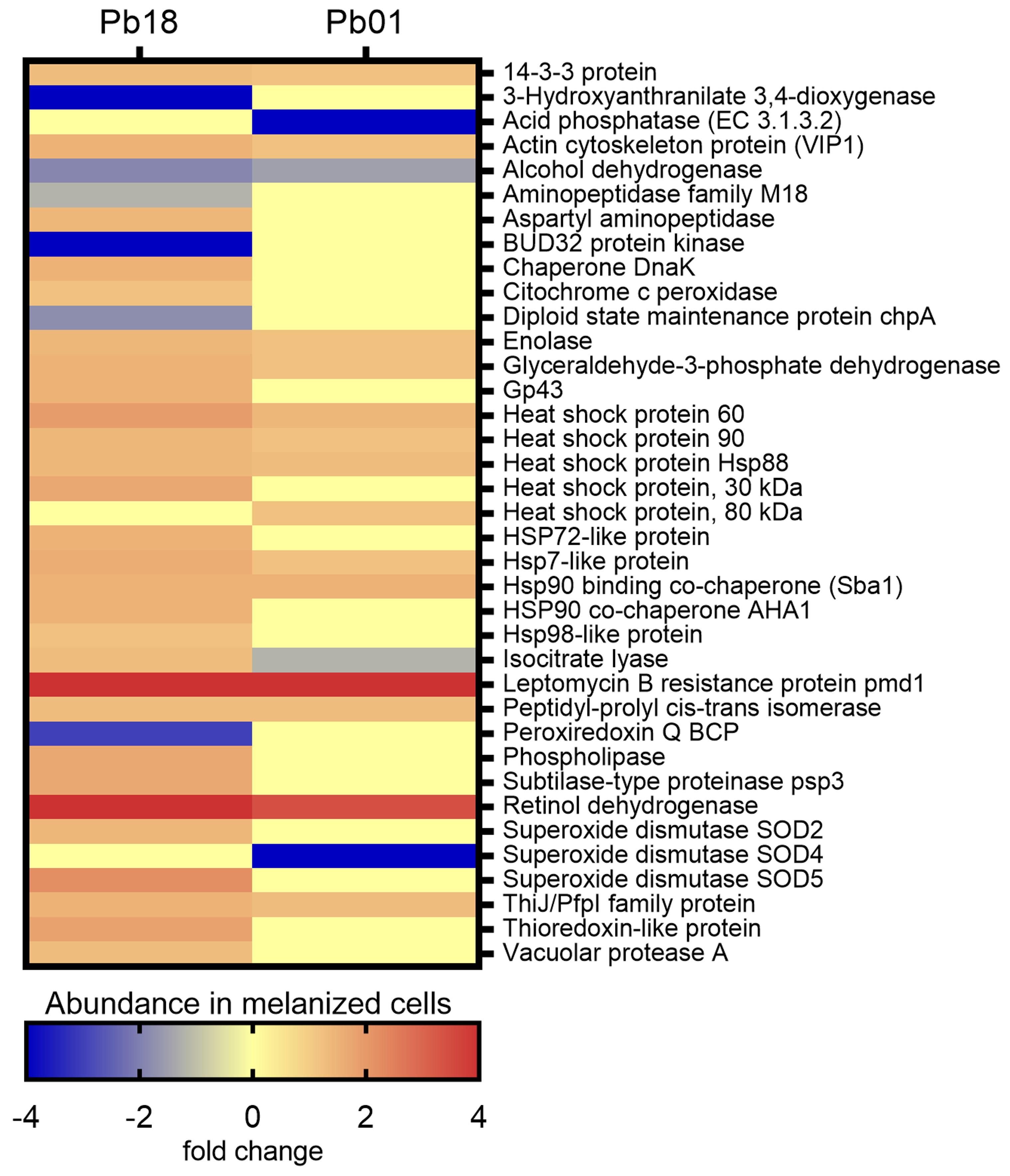
3.4. Virulence-Associated Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hrycyk, M.F.; Garcia Garces, H.; de Bosco, S.M.G.; de Oliveira, S.L.; Marques, S.A.; Bagagli, E. Ecology of Paracoccidioides brasiliensis, P. lutzii and related species: Infection in armadillos, soil occurrence and mycological aspects. Med. Mycol. 2018, 56, 950–962. [Google Scholar] [CrossRef]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian guidelines for the clinical management of paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- de Teixeira, M.M.; Theodoro, R.C.; de Oliveira, F.F.M.; Machado, G.C.; Hahn, R.C.; Bagagli, E.; San-Blas, G.; Soares Felipe, M.S. Paracoccidioides lutzii sp. nov.: Biological and clinical implications. Med. Mycol. 2014, 52, 19–28. [Google Scholar] [CrossRef]
- de Macedo, P.M.; de Teixeira, M.M.; Barker, B.M.; Zancopé-Oliveira, R.M.; Almeida-Paes, R.; do Francesconi Valle, A.C. Clinical features and genetic background of the sympatric species Paracoccidioides brasiliensis and Paracoccidioides americana. PLoS Negl. Trop. Dis. 2019, 13, e0007309. [Google Scholar] [CrossRef]
- Hahn, R.C.; Rodrigues, A.M.; Della Terra, P.P.; Nery, A.F.; Hoffmann-Santos, H.D.; Góis, H.M.; Fontes, C.J.F.; de Camargo, Z.P. Clinical and epidemiological features of paracoccidioidomycosis due to Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2019, 13, e0007437. [Google Scholar] [CrossRef]
- Pereira, E.F.; Gegembauer, G.; Chang, M.R.; de Camargo, Z.P.; Nunes, T.F.; Ribeiro, S.M.; de Carvalho, L.R.; Maldonado, B.M.; Mendes, R.P.; Paniago, A.M.M. Comparison of clinico-epidemiological and radiological features in paracoccidioidomycosis patients regarding serological classification using antigens from Paracoccidioides brasiliensis complex and Paracoccidioides lutzii. PLoS Negl. Trop. Dis. 2020, 14, e0008485. [Google Scholar] [CrossRef]
- De Silva, J.F.; de Oliveira, H.C.; Marcos, C.M.; Assato, P.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Advances and challenges in paracoccidioidomycosis serology caused by Paracoccidioides species complex: An update. Diagn. Microbiol. Infect. Dis. 2016, 84, 87–94. [Google Scholar] [CrossRef]
- Siqueira, I.M.; Fraga, C.L.F.; Amaral, A.C.; Souza, A.C.O.; Jerônimo, M.S.; Correa, J.R.; Magalhães, K.G.; Inácio, C.A.; Ribeiro, A.M.; Burguel, P.H.; et al. Distinct patterns of yeast cell morphology and host responses induced by representative strains of Paracoccidioides brasiliensis (Pb18) and Paracoccidioides lutzii (Pb01). Med. Mycol. 2016, 54, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.A.; Grisolia, J.C.; Burger, E.; de Araujo Paula, F.B.; Dias, A.L.T.; Malaquias, L.C.C. Virulence factors of Paracoccidioides brasiliensis as therapeutic targets: A review. Antonie Van Leeuwenhoek 2020, 113, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Casadevall, A. The contribution of melanin to microbial pathogenesis. Cell. Microbiol. 2003, 5, 203–223. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Greer, E.M.; McGrail, C.W. The role of melanins in melanotic fungi for pathogenesis and environmental survival. Appl. Microbiol. Biotechnol. 2020, 104, 4247–4257. [Google Scholar] [CrossRef] [PubMed]
- Plonka, P.M.; Grabacka, M. Melanin synthesis in microorganisms—Biotechnological and medical aspects. Acta Biochim. Pol. 2006, 53, 429–443. [Google Scholar] [CrossRef] [PubMed]
- Gómez, B.L.; Nosanchuk, J.D.; Díez, S.; Youngchim, S.; Aisen, P.; Cano, L.E.; Restrepo, A.; Casadevall, A.; Hamilton, A.J. Detection of melanin-like pigments in the dimorphic fungal pathogen Paracoccidioides brasiliensis in vitro and during infection. Infect. Immun. 2001, 69, 5760–5767. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.B.; Marques, A.F.; Nosanchuk, J.D.; Casadevall, A.; Travassos, L.R.; Taborda, C.P. Melanin in the dimorphic fungal pathogen Paracoccidioides brasiliensis: Effects on phagocytosis, intracellular resistance and drug susceptibility. Microbes Infect. 2006, 8, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Chaves, E.G.A.; Parente-Rocha, J.A.; Baeza, L.C.; Araújo, D.S.; Borges, C.L.; de Oliveira, M.A.P.; de Soares, C.M.A. Proteomic Analysis of Paracoccidioides brasiliensis During Infection of Alveolar Macrophages Primed or Not by Interferon-Gamma. Front. Microbiol. 2019, 10, 96. [Google Scholar] [CrossRef]
- De Lima, P.S.; Chung, D.; Bailão, A.M.; Cramer, R.A.; de Soares, C.M.A. Characterization of the Paracoccidioides Hypoxia Response Reveals New Insights into Pathogenesis Mechanisms of This Important Human Pathogenic Fungus. PLoS Negl. Trop. Dis. 2015, 9, e0004282. [Google Scholar] [CrossRef]
- De Lima, P.S.; Casaletti, L.; Bailão, A.M.; de Vasconcelos, A.T.R.; da Fernandes, G.R.; de Soares, C.M.A. Transcriptional and proteomic responses to carbon starvation in Paracoccidioides. PLoS Negl. Trop. Dis. 2014, 8, e2855. [Google Scholar] [CrossRef]
- Teixeira, H.C.; Calich, V.L.; Singer-Vermes, L.M.; D’Imperio-Lima, M.R.; Russo, M. Experimental paracoccidioidomycosis: Early immunosuppression occurs in susceptible mice after infection with pathogenic fungi. Braz. J. Med. Biol. Res. Rev. Bras. Pesqui. Med. E Biol. 1987, 20, 587–589. [Google Scholar]
- Teixeira, M.M.; Theodoro, R.C.; de Carvalho, M.J.A.; Fernandes, L.; Paes, H.C.; Hahn, R.C.; Mendoza, L.; Bagagli, E.; San-Blas, G.; Felipe, M.S.S. Phylogenetic analysis reveals a high level of speciation in the Paracoccidioides genus. Mol. Phylogenet. Evol. 2009, 52, 273–283. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.C.; Lima, D.B.; Leprevost, F.V.; Santos, M.D.M.; Fischer, J.S.G.; Aquino, P.F.; Moresco, J.J.; Yates, J.R.; Barbosa, V.C. Integrated analysis of shotgun proteomic data with PatternLab for proteomics 4.0. Nat. Protoc. 2016, 11, 102–117. [Google Scholar] [CrossRef] [PubMed]
- Eng, J.K.; Jahan, T.A.; Hoopmann, M.R. Comet: An open-source MS/MS sequence database search tool. Proteomics 2013, 13, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, P.C.; Fischer, J.S.G.; Xu, T.; Cociorva, D.; Balbuena, T.S.; Valente, R.H.; Perales, J.; Yates, J.R.; Barbosa, V.C. Search engine processor: Filtering and organizing peptide spectrum matches. Proteomics 2012, 12, 944–949. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef]
- Elias Moreira, A.L.; Milhomem Cruz-Leite, V.R.; O’Hara Souza Silva, L.; Alves Parente, A.F.; Bailão, A.M.; Maria de Almeida Soares, C.; Parente-Rocha, J.A.; Ruiz, O.H.; Borges, C.L. Proteome characterization of Paracoccidioides lutzii conidia by using nanoUPLC-MSE. Fungal Biol. 2020, 124, 766–780. [Google Scholar] [CrossRef]
- Araújo, F.S.; Coelho, L.M.; do Silva, L.C.; da Silva Neto, B.R.; Parente-Rocha, J.A.; Bailão, A.M.; de Oliveira, C.M.A.; da Fernandes, G.R.; Hernández, O.; Ochoa, J.G.M.; et al. Effects of Argentilactone on the Transcriptional Profile, Cell Wall and Oxidative Stress of Paracoccidioides spp. PLoS Negl. Trop. Dis. 2016, 10, e0004309. [Google Scholar] [CrossRef]
- Borba, J.V.V.B.; Tauhata, S.B.F.; de Oliveira, C.M.A.; Ferreira Marques, M.; Bailão, A.M.; de Soares, C.M.A.; Pereira, M. Chemoproteomic identification of molecular targets of antifungal prototypes, thiosemicarbazide and a camphene derivative of thiosemicarbazide, in Paracoccidioides brasiliensis. PLoS ONE 2018, 13, e0201948. [Google Scholar] [CrossRef]
- Araújo, D.S.; Pereira, M.; Portis, I.G.; de Dos Santos Junior, A.C.M.; Fontes, W.; de Sousa, M.V.; do Assunção, L.P.; Baeza, L.C.; Bailão, A.M.; Ricart, C.A.O.; et al. Metabolic Peculiarities of Paracoccidioides brasiliensis Dimorphism as Demonstrated by iTRAQ Labeling Proteomics. Front. Microbiol. 2019, 10, 555. [Google Scholar] [CrossRef]
- Baeza, L.C.; da Mata, F.R.; Pigosso, L.L.; Pereira, M.; de Souza, G.H.M.F.; Coelho, A.S.G.; de Almeida Soares, C.M. Differential Metabolism of a Two-Carbon Substrate by Members of the Paracoccidioides Genus. Front. Microbiol. 2017, 8, 2308. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, J.F.; Farrer, R.A.; Desjardins, C.A.; Gallo, J.E.; Sykes, S.; Sakthikumar, S.; Misas, E.; Whiston, E.A.; Bagagli, E.; Soares, C.M.A.; et al. Genome Diversity, Recombination, and Virulence across the Major Lineages of Paracoccidioides. mSphere 2016, 1, e00213-16. [Google Scholar] [CrossRef] [PubMed]
- Soares, D.A.; Oliveira, M.B.; Evangelista, A.F.; Venancio, E.J.; Andrade, R.V.; Felipe, M.S.S.; Petrofeza, S. Phospholipase gene expression during Paracoccidioides brasiliensis morphological transition and infection. Mem. Inst. Oswaldo Cruz 2013, 108, 808–811. [Google Scholar] [CrossRef] [PubMed]
- Portis, I.G.; de Sousa Lima, P.; Paes, R.A.; Oliveira, L.N.; Pereira, C.A.; Parente-Rocha, J.A.; Pereira, M.; Nosanchuk, J.D.; de Almeida Soares, C.M. Copper overload in Paracoccidioides lutzii results in the accumulation of ergosterol and melanin. Microbiol. Res. 2020, 239, 126524. [Google Scholar] [CrossRef] [PubMed]
- Petito, G.; de Curcio, J.S.; Pereira, M.; Bailão, A.M.; Paccez, J.D.; Tristão, G.B.; de Morais, C.O.B.; de Souza, M.V.; de Castro Moreira Santos, A.; Fontes, W.; et al. Metabolic Adaptation of Paracoccidioides brasiliensis in Response to in vitro Copper Deprivation. Front. Microbiol. 2020, 11, 1834. [Google Scholar] [CrossRef] [PubMed]
- da Rodrigues, L.N.S.; de Brito, W.A.; Parente, A.F.A.; Weber, S.S.; Bailão, A.M.; Casaletti, L.; Borges, C.L.; de Soares, C.M.A. Osmotic stress adaptation of Paracoccidioides lutzii, Pb01, monitored by proteomics. Fungal Genet. Biol. 2016, 95, 13–23. [Google Scholar] [CrossRef]
- Eisenman, H.C.; Chow, S.-K.; Tsé, K.K.; McClelland, E.E.; Casadevall, A. The effect of L-DOPA on Cryptococcus neoformans growth and gene expression. Virulence 2011, 2, 329–336. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Gómez, B.L.; Youngchim, S.; Díez, S.; Aisen, P.; Zancopé-Oliveira, R.M.; Restrepo, A.; Casadevall, A.; Hamilton, A.J. Histoplasma capsulatum synthesizes melanin-like pigments in vitro and during mammalian infection. Infect. Immun. 2002, 70, 5124–5131. [Google Scholar] [CrossRef]
- Figueiredo-Carvalho, M.H.G.; dos Santos, F.B.; Nosanchuk, J.D.; Zancope-Oliveira, R.M.; Almeida-Paes, R. L-Dihydroxyphenylalanine induces melanin production by members of the genus Trichosporon. FEMS Yeast Res. 2014, 14, 988–991. [Google Scholar] [CrossRef]
- Van de Sande, W.W.J.; de Kat, J.; Coppens, J.; Ahmed, A.O.A.; Fahal, A.; Verbrugh, H.; van Belkum, A. Melanin biosynthesis in Madurella mycetomatis and its effect on susceptibility to itraconazole and ketoconazole. Microbes Infect. 2007, 9, 1114–1123. [Google Scholar] [CrossRef]
- Teixeira, P.A.C.; De Castro, R.A.; Ferreira, F.R.L.; Cunha, M.M.L.; Torres, A.P.; Penha, C.V.L.Y.; Rozental, S.; Lopes-Bezerra, L.M. L-DOPA accessibility in culture medium increases melanin expression and virulence of Sporothrix schenckii yeast cells. Med. Mycol. 2010, 48, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Eisenhofer, G.; Tian, H.; Holmes, C.; Matsunaga, J.; Roffler-Tarlov, S.; Hearing, V.J. Tyrosinase: A developmentally specific major determinant of peripheral dopamine. FASEB J. 2003, 17, 1248–1255. [Google Scholar] [CrossRef] [PubMed]
- Peres da Silva, R.; Matsumoto, M.T.; Braz, J.D.; Voltan, A.R.; de Oliveira, H.C.; Soares, C.P.; Mendes Giannini, M.J.S. Differential gene expression analysis of Paracoccidioides brasiliensis during keratinocyte infection. J. Med. Microbiol. 2011, 60, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, M.C.; Nakayasu, E.S.; Matsuo, A.L.; Sobreira, T.J.P.; Longo, L.V.G.; Ganiko, L.; Almeida, I.C.; Puccia, R. Vesicle and vesicle-free extracellular proteome of Paracoccidioides brasiliensis: Comparative analysis with other pathogenic fungi. J. Proteome Res. 2012, 11, 1676–1685. [Google Scholar] [CrossRef]
- Herrero, E.; Ros, J.; Bellí, G.; Cabiscol, E. Redox control and oxidative stress in yeast cells. Biochim. Biophys. Acta 2008, 1780, 1217–1235. [Google Scholar] [CrossRef]
- Silva-Bailão, M.G.; de Lima, P.S.; Oliveira, M.M.E.; Oliveira, L.C.; Almeida-Paes, R.; Borges, C.L.; Bailão, A.M.; Coelho, A.S.G.; de Soares, C.M.A.; Zancopé-Oliveira, R.M. Comparative proteomics in the three major human pathogenic species of the genus Sporothrix. Microbes Infect. 2020. [CrossRef]
- Pigosso, L.L.; Parente, A.F.A.; Coelho, A.S.G.; Silva, L.P.; Borges, C.L.; Bailão, A.M.; de Soares, C.M.A. Comparative proteomics in the genus Paracoccidioides. Fungal Genet. Biol. 2013, 60, 87–100. [Google Scholar] [CrossRef]
- Chew, S.Y.; Ho, K.L.; Cheah, Y.K.; Ng, T.S.; Sandai, D.; Brown, A.J.P.; Than, L.T.L. Glyoxylate cycle gene ICL1 is essential for the metabolic flexibility and virulence of Candida glabrata. Sci. Rep. 2019, 9, 2843. [Google Scholar] [CrossRef]
- Ting, S.Y.; Ishola, O.A.; Ahmed, M.A.; Tabana, Y.M.; Dahham, S.; Agha, M.T.; Musa, S.F.; Muhammed, R.; Than, L.T.L.; Sandai, D. Metabolic adaptation via regulated enzyme degradation in the pathogenic yeast Candida albicans. J. Mycol. Med. 2017, 27, 98–108. [Google Scholar] [CrossRef]
- Marcos, C.M.; de Oliveira, H.C.; de Silva, J.F.; Assato, P.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. The multifaceted roles of metabolic enzymes in the Paracoccidioides species complex. Front. Microbiol. 2014, 5, 719. [Google Scholar] [CrossRef]
- Lacerda Pigosso, L.; Baeza, L.C.; Vieira Tomazett, M.; Batista Rodrigues Faleiro, M.; Brianezi Dignani de Moura, V.M.; Melo Bailão, A.; Borges, C.L.; Alves Parente Rocha, J.; Rocha Fernandes, G.; Gauthier, G.M.; et al. Paracoccidioides brasiliensis presents metabolic reprogramming and secretes a serine proteinase during murine infection. Virulence 2017, 8, 1417–1434. [Google Scholar] [CrossRef] [PubMed]
- Parente-Rocha, J.A.; Parente, A.F.A.; Baeza, L.C.; Bonfim, S.M.R.C.; Hernandez, O.; McEwen, J.G.; Bailão, A.M.; Taborda, C.P.; Borges, C.L.; de Soares, C.M.A. Macrophage Interaction with Paracoccidioides brasiliensis Yeast Cells Modulates Fungal Metabolism and Generates a Response to Oxidative Stress. PLoS ONE 2015, 10, e0137619. [Google Scholar] [CrossRef] [PubMed]
- Silva, P.F.F.; Novaes, E.; Pereira, M.; Soares, C.M.A.; Borges, C.L.; Salem-Izacc, S.M. In silico characterization of hypothetical proteins from Paracoccidioides lutzii. Genet. Mol. Res. 2015, 14, 17416–17425. [Google Scholar] [CrossRef] [PubMed]
- Navarro, M.V.; Chaves, A.F.A.; Castilho, D.G.; Casula, I.; Calado, J.C.P.; Conceição, P.M.; Iwai, L.K.; de Castro, B.F.; Batista, W.L. Effect of Nitrosative Stress on the S-Nitroso-Proteome of Paracoccidioides brasiliensis. Front. Microbiol. 2020, 11, 1184. [Google Scholar] [CrossRef]
- Cleare, L.G.; Zamith-Miranda, D.; Nosanchuk, J.D. Heat Shock Proteins in Histoplasma and Paracoccidioides. Clin. Vaccine Immunol. 2017, 24, 24. [Google Scholar] [CrossRef]
- Zügel, U.; Kaufmann, S.H. Role of heat shock proteins in protection from and pathogenesis of infectious diseases. Clin. Microbiol. Rev. 1999, 12, 19–39. [Google Scholar] [CrossRef]
- Carvalho, K.C.; Ganiko, L.; Batista, W.L.; Morais, F.V.; Marques, E.R.; Goldman, G.H.; Franco, M.F.; Puccia, R. Virulence of Paracoccidioides brasiliensis and gp43 expression in isolates bearing known PbGP43 genotype. Microbes Infect. 2005, 7, 55–65. [Google Scholar] [CrossRef]
- Leitão, N.P.; Vallejo, M.C.; Conceição, P.M.; Camargo, Z.P.; Hahn, R.; Puccia, R. Paracoccidioides lutzii Plp43 is an active glucanase with partial antigenic identity with P. brasiliensis gp43. PLoS Negl. Trop. Dis. 2014, 8, e3111. [Google Scholar] [CrossRef]
- Tamayo, D.; Muñoz, J.F.; Lopez, Á.; Urán, M.; Herrera, J.; Borges, C.L.; Restrepo, Á.; Soares, C.M.; Taborda, C.P.; Almeida, A.J.; et al. Identification and Analysis of the Role of Superoxide Dismutases Isoforms in the Pathogenesis of Paracoccidioides spp. PLoS Negl. Trop. Dis. 2016, 10, e0004481. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | Pb18 | Pb01 | ||
|---|---|---|---|---|
| Count in Gene Set | False Discovery Rate | Count in Gene Set | False Discovery Rate | |
| Carbon metabolism | ||||
| Glycolysis/Gluconeogenesis | 13/34 | 1.41 × 10−18 | 4/33 | 7.21 × 10−6 |
| Oxidative phosphorylation | 13/71 | 4.64 × 10−15 | Not enriched | |
| Citrate cycle | 9/27 | 1.66 × 10−12 | 4/26 | 3.70 × 10−6 |
| Methane metabolism | 8/20 | 1.03 × 10−11 | 7/19 | 1.21 × 10−12 |
| Pentose phosphate pathway | 8/23 | 2.33 × 10−11 | 5/23 | 3.75 × 10−8 |
| Glyoxylate and dicarboxylate metabolism | 7/33 | 8.81 × 10−9 | 5/21 | 1.24 × 10−7 |
| Fatty Acid metabolism | 3/29 | 0.0018 | Not enriched | |
| Nucleotide metabolism | ||||
| Purine metabolism | 27/86 | 6.77 × 10−43 | 11/80 | 7.10 × 10−18 |
| Pyrimidine metabolism | 19/61 | 4.25 × 10−29 | 11/56 | 2.94 × 10−19 |
| Amino acid metabolism | ||||
| Val, Leu, and Ile degradation | 13/28 | 1.19 × 10−17 | 6/26 | 7.04 × 10−8 |
| Tyr metabolism | 12/30 | 1.01 × 10−15 | 10/27 | 2.73 × 10−14 |
| Ala, Asp, and Glu metabolism | 11/30 | 3.49 × 10−14 | 7/30 | 4.21 × 10−9 |
| Phe, Tyr, and Trp biosynthesis | 7/17 | 1.32 × 10−9 | 5/16 | 2.82 × 10−7 |
| Gly, Ser, and Thr metabolism | 8/40 | 8.04 × 10−9 | 9/36 | 1.09 × 10−11 |
| Phe metabolism | 7/25 | 1.07 × 10−8 | 7/23 | 9.43 × 10−10 |
| Val, Leu, and Ile biosynthesis | 5/14 | 7.10 × 10−7 | 3/12 | 0.00017 |
| Beta-Ala metabolism | 5/19 | 2.39 × 10−6 | 2/16 | 0.0066 |
| Lys degradation | 4/19 | 5.97 × 10−5 | 4/18 | 1.84 × 10−5 |
| Arg biosynthesis | 4/19 | 5.97 × 10−5 | 3/19 | 0.00048 |
| Other pathways | ||||
| Spliceosome | 21/83 | 2.60 × 10−17 | Not enriched | |
| RNA degradation | 17/48 | 3.07 × 10−16 | 8/41 | 1.28 × 10−10 |
| RNA transport | 19/86 | 4.00 × 10−15 | 12/83 | 1.70 × 10−14 |
| Protein processing in endoplasmic reticulum | 18/74 | 4.45 × 10−15 | Not enriched | |
| Phagosome | 14/35 | 2.51 × 10−14 | Not enriched | |
| Aminoacyl t-RNA biosynthesis | 13/38 | 1.10 × 10−12 | Not enriched | |
| Proteasome | 11/35 | 1.59 × 10−10 | Not enriched | |
| Basal transcription factors | 10/29 | 5.35 × 10−10 | Not enriched | |
| SNARE interactions in vesicular transport | 3/15 | 0.0080 | Not enriched | |
| Ubiquitin mediated proteolysis | 4/52 | 0.0299 | Not enriched | |
| mRNA surveillance | Not enriched | 8/44 | 1.41 × 10−10 | |
| Peroxisome | Not enriched | 8/51 | 3.06 × 10−10 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida-Paes, R.; Almeida, M.A.; Baeza, L.C.; Marmello, L.A.M.; Trugilho, M.R.d.O.; Nosanchuk, J.D.; Soares, C.M.d.A.; Valente, R.H.; Zancopé-Oliveira, R.M. Beyond Melanin: Proteomics Reveals Virulence-Related Proteins in Paracoccidioides brasiliensis and Paracoccidioides lutzii Yeast Cells Grown in the Presence of L-Dihydroxyphenylalanine. J. Fungi 2020, 6, 328. https://doi.org/10.3390/jof6040328
Almeida-Paes R, Almeida MA, Baeza LC, Marmello LAM, Trugilho MRdO, Nosanchuk JD, Soares CMdA, Valente RH, Zancopé-Oliveira RM. Beyond Melanin: Proteomics Reveals Virulence-Related Proteins in Paracoccidioides brasiliensis and Paracoccidioides lutzii Yeast Cells Grown in the Presence of L-Dihydroxyphenylalanine. Journal of Fungi. 2020; 6(4):328. https://doi.org/10.3390/jof6040328
Chicago/Turabian StyleAlmeida-Paes, Rodrigo, Marcos Abreu Almeida, Lilian Cristiane Baeza, Leticia Andrade Mendes Marmello, Monique Ramos de Oliveira Trugilho, Joshua Daniel Nosanchuk, Celia Maria de Almeida Soares, Richard Hemmi Valente, and Rosely Maria Zancopé-Oliveira. 2020. "Beyond Melanin: Proteomics Reveals Virulence-Related Proteins in Paracoccidioides brasiliensis and Paracoccidioides lutzii Yeast Cells Grown in the Presence of L-Dihydroxyphenylalanine" Journal of Fungi 6, no. 4: 328. https://doi.org/10.3390/jof6040328
APA StyleAlmeida-Paes, R., Almeida, M. A., Baeza, L. C., Marmello, L. A. M., Trugilho, M. R. d. O., Nosanchuk, J. D., Soares, C. M. d. A., Valente, R. H., & Zancopé-Oliveira, R. M. (2020). Beyond Melanin: Proteomics Reveals Virulence-Related Proteins in Paracoccidioides brasiliensis and Paracoccidioides lutzii Yeast Cells Grown in the Presence of L-Dihydroxyphenylalanine. Journal of Fungi, 6(4), 328. https://doi.org/10.3390/jof6040328