Monoclonal Antibodies as Tools to Combat Fungal Infections



Abstract
1. Introduction
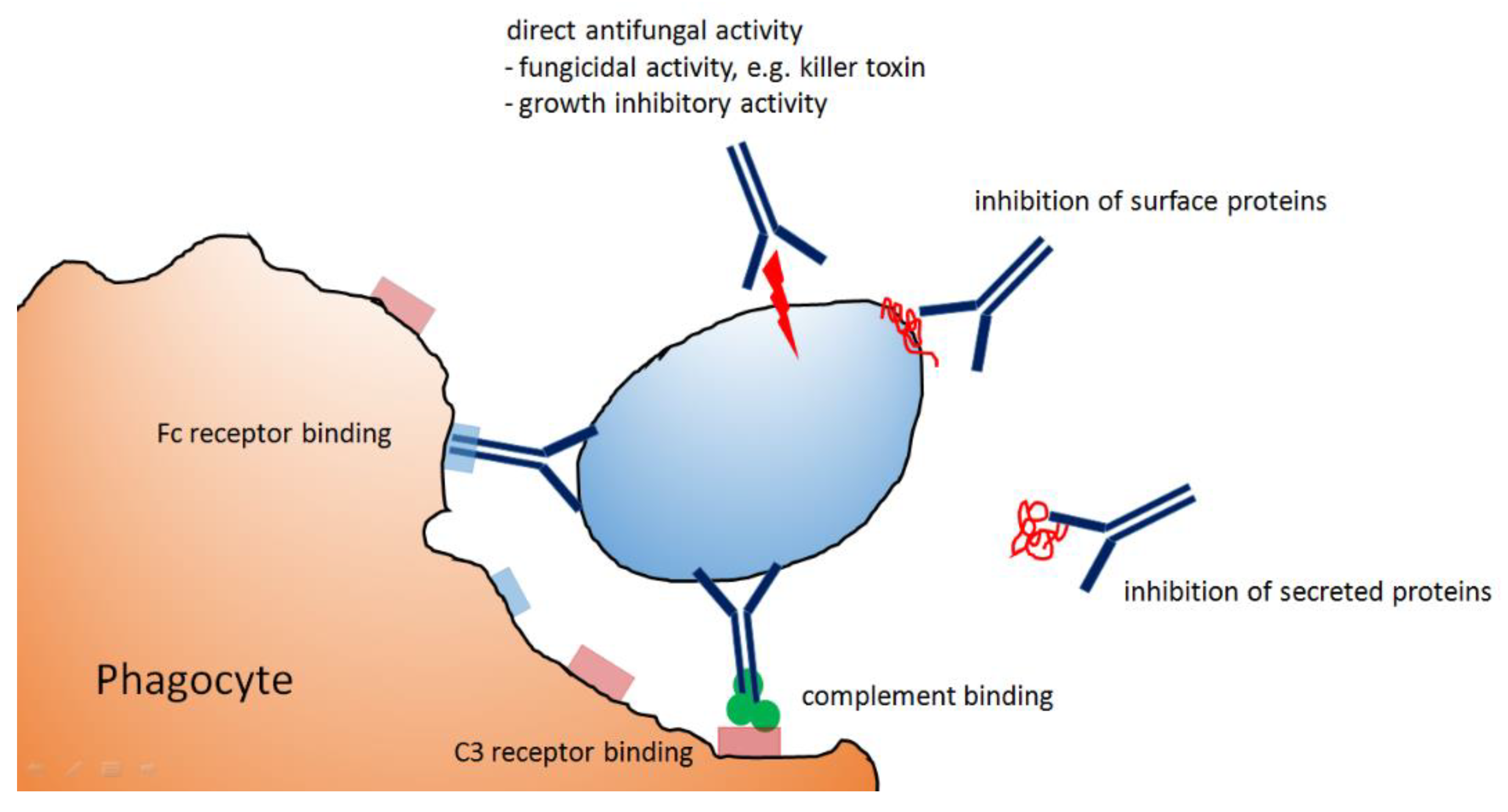
2. Elimination of Microbial Pathogens by Antibody-Dependent Mechanisms
3. The Cell Wall as Primary Target Structure for Antifungal Antibodies
4. Protective Antibodies against Cryptococcus neoformans
5. Protective Antibodies against Candida albicans
6. Protective Antibodies against Dimorphic Fungi
7. Protective Antibodies against Molds
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Brown, G.D.; Denning, D.W.; Gow, N.A.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef] [PubMed]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef] [PubMed]
- Bowen, A.; Wear, M.P.; Cordero, R.J.; Oscarson, S.; Casadevall, A. A Monoclonal Antibody to Cryptococcus neoformans Glucuronoxylomannan Manifests Hydrolytic Activity for Both Peptides and Polysaccharides. J. Biol. Chem. 2017, 292, 417–434. [Google Scholar] [CrossRef] [PubMed]
- Dromer, F.; Charreire, J.; Contrepois, A.; Carbon, C.; Yeni, P. Protection of mice against experimental cryptococcosis by anti-Cryptococcus neoformans monoclonal antibody. Infect. Immun. 1987, 55, 749–752. [Google Scholar] [CrossRef] [PubMed]
- Fleuridor, R.; Zhong, Z.; Pirofski, L. A human IgM monoclonal antibody prolongs survival of mice with lethal cryptococcosis. J. Infect. Dis. 1998, 178, 1213–1216. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yuan, R.; Clynes, R.; Oh, J.; Ravetch, J.V.; Scharff, M.D. Antibody-mediated modulation of Cryptococcus neoformans infection is dependent on distinct Fc receptor functions and IgG subclasses. J. Exp. Med. 1998, 187, 641–648. [Google Scholar] [CrossRef]
- Mukherjee, J.; Scharff, M.D.; Casadevall, A. Protective murine monoclonal antibodies to Cryptococcus neoformans. Infect. Immun. 1992, 60, 4534–4541. [Google Scholar] [CrossRef]
- Saylor, C.A.; Dadachova, E.; Casadevall, A. Murine IgG1 and IgG3 isotype switch variants promote phagocytosis of Cryptococcus neoformans through different receptors1. J. Immunol. 2010, 184, 336–343. [Google Scholar] [CrossRef]
- Yuan, R.R.; Casadevall, A.; Oh, J.; Scharff, M.D. T cells cooperate with passive antibody to modify Cryptococcus neoformans infection in mice. Proc. Natl. Acad. Sci. USA 1997, 94, 2483–2488. [Google Scholar] [CrossRef]
- Yuan, R.; Casadevall, A.; Spira, G.; Scharff, M.D. Isotype switching from IgG3 to IgG1 converts a nonprotective murine antibody to Cryptococcus neoformans into a protective antibody. J. Immunol. 1995, 154, 1810–1816. [Google Scholar]
- Rivera, J.; Casadevall, A. Mouse genetic background is a major determinant of isotype-related differences for antibody-mediated protective efficacy against Cryptococcus neoformans. J. Immunol. 2005, 174, 8017–8026. [Google Scholar] [CrossRef] [PubMed]
- Taborda, C.P.; Casadevall, A. Immunoglobulin M efficacy against Cryptococcus neoformans: Mechanism, dose dependence, and prozone-like effects in passive protection experiments. J. Immunol. 2001, 166, 2100–2107. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, J.; Nussbaum, G.; Scharff, M.D.; Casadevall, A. Protective and nonprotective monoclonal antibodies to Cryptococcus neoformans originating from one B cell. J. Exp. Med. 1995, 181, 405–409. [Google Scholar] [CrossRef]
- Maitta, R.W.; Datta, K.; Chang, Q.; Luo, R.X.; Witover, B.; Subramaniam, K.; Pirofski, L.A. Protective and nonprotective human immunoglobulin M monoclonal antibodies to Cryptococcus neoformans glucuronoxylomannan manifest different specificities and gene use profiles. Infect. Immun. 2004, 72, 4810–4818. [Google Scholar] [CrossRef]
- Nakouzi, A.; Zhang, T.; Oscarson, S.; Casadevall, A. The common Cryptococcus neoformans glucuronoxylomannan M2 motif elicits non-protective antibodies. Vaccine 2009, 27, 3513–3518. [Google Scholar] [CrossRef]
- Shapiro, S.; Beenhouwer, D.O.; Feldmesser, M.; Taborda, C.; Carroll, M.C.; Casadevall, A. Immunoglobulin G monoclonal antibodies to Cryptococcus neoformans protect mice deficient in complement component C3. Infect. Immun. 2002, 70, 2598–2604. [Google Scholar] [CrossRef]
- Taborda, C.P.; Rivera, J.; Zaragoza, O.; Casadevall, A. More is not necessarily better: Prozone-like effects in passive immunization with IgG. J. Immunol. 2003, 170, 3621–3630. [Google Scholar] [CrossRef]
- Savoy, A.C.; Lupan, D.M.; Manalo, P.B.; Roberts, J.S.; Schlageter, A.M.; Weinhold, L.C.; Kozel, T.R. Acute lethal toxicity following passive immunization for treatment of murine cryptococcosis. Infect. Immun. 1997, 65, 1800–1807. [Google Scholar] [CrossRef]
- Lendvai, N.; Casadevall, A. Monoclonal antibody-mediated toxicity in Cryptococcus neoformans infection: Mechanism and relationship to antibody isotype. J. Infect. Dis. 1999, 180, 791–801. [Google Scholar] [CrossRef][Green Version]
- Westin Kwon, K.; Lendvai, N.; Morrison, S.; Trinh, K.R.; Casadevall, A. Biological activity of a mouse-human chimeric immunoglobulin G2 antibody to Cryptococcus neoformans polysaccharide. Clin. Diagn. Lab. Immunol. 2002, 9, 201–204. [Google Scholar] [CrossRef]
- Matthews, R.C.; Burnie, J.P.; Tabaqchali, S. Isolation of immunodominant antigens from sera of patients with systemic candidiasis and characterization of serological response to Candida albicans. J. Clin. Microbiol. 1987, 25, 230–237. [Google Scholar] [CrossRef]
- Matthews, R.; Burnie, J. Cloning of a DNA sequence encoding a major fragment of the 47 kilodalton stress protein homologue of Candida albicans. FEMS Microbiol. Lett. 1989, 51, 25–30. [Google Scholar] [CrossRef]
- O’Meara, T.R.; Robbins, N.; Cowen, L.E. The Hsp90 Chaperone Network Modulates Candida Virulence Traits. Trends Microbiol. 2017, 25, 809–819. [Google Scholar] [CrossRef]
- Matthews, R.C.; Burnie, J.P.; Howat, D.; Rowland, T.; Walton, F. Autoantibody to heat-shock protein 90 can mediate protection against systemic candidosis. Immunology 1991, 74, 20–24. [Google Scholar]
- Matthews, R.C.; Rigg, G.; Hodgetts, S.; Carter, T.; Chapman, C.; Gregory, C.; Illidge, C.; Burnie, J. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. Antimicrob. Agents Chemother. 2003, 47, 2208–2216. [Google Scholar] [CrossRef]
- Burt, E.T.; Daly, R.; Hoganson, D.; Tsirulnikov, Y.; Essmann, M.; Larsen, B. Isolation and partial characterization of Hsp90 from Candida albicans. Ann. Clin. Lab. Sci. 2003, 33, 86–93. [Google Scholar]
- Pachl, J.; Svoboda, P.; Jacobs, F.; Vandewoude, K.; van der Hoven, B.; Spronk, P.; Masterson, G.; Malbrain, M.; Aoun, M.; Garbino, J.; et al. A randomized, blinded, multicenter trial of lipid-associated amphotericin B alone versus in combination with an antibody-based inhibitor of heat shock protein 90 in patients with invasive candidiasis. Clin. Infect. Dis. 2006, 42, 1404–1413. [Google Scholar] [CrossRef]
- Torosantucci, A.; Bromuro, C.; Chiani, P.; De Bernardis, F.; Berti, F.; Galli, C.; Norelli, F.; Bellucci, C.; Polonelli, L.; Costantino, P.; et al. A novel glyco-conjugate vaccine against fungal pathogens. J. Exp. Med. 2005, 202, 597–606. [Google Scholar] [CrossRef]
- Rachini, A.; Pietrella, D.; Lupo, P.; Torosantucci, A.; Chiani, P.; Bromuro, C.; Proietti, C.; Bistoni, F.; Cassone, A.; Vecchiarelli, A. An anti-beta-glucan monoclonal antibody inhibits growth and capsule formation of Cryptococcus neoformans in vitro and exerts therapeutic, anticryptococcal activity in vivo. Infect. Immun. 2007, 75, 5085–5094. [Google Scholar] [CrossRef]
- Torosantucci, A.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Palma, A.S.; Liu, Y.; Mignogna, G.; Maras, B.; Colone, M.; Stringaro, A.; et al. Protection by anti-beta-glucan antibodies is associated with restricted beta-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS ONE 2009, 4, e5392. [Google Scholar] [CrossRef]
- Capodicasa, C.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Catellani, M.; Palma, A.S.; Liu, Y.; Feizi, T.; Cassone, A.; Benvenuto, E.; et al. Plant production of anti-β-glucan antibodies for immunotherapy of fungal infections in humans. Plant. Biotechnol. J. 2011, 9, 776–787. [Google Scholar] [CrossRef]
- Matveev, A.L.; Krylov, V.B.; Khlusevich, Y.A.; Baykov, I.K.; Yashunsky, D.V.; Emelyanova, L.A.; Tsvetkov, Y.E.; Karelin, A.A.; Bardashova, A.V.; Wong, S.S.W.; et al. Novel mouse monoclonal antibodies specifically recognizing β-(1→3)-D-glucan antigen. PLoS ONE 2019, 14, e0215535. [Google Scholar] [CrossRef]
- Han, Y.; Kanbe, T.; Cherniak, R.; Cutler, J.E. Biochemical characterization of Candida albicans epitopes that can elicit protective and nonprotective antibodies. Infect. Immun. 1997, 65, 4100–4107. [Google Scholar] [CrossRef]
- Han, Y.; Morrison, R.P.; Cutler, J.E. A vaccine and monoclonal antibodies that enhance mouse resistance to Candida albicans vaginal infection. Infect. Immun. 1998, 66, 5771–5776. [Google Scholar] [CrossRef]
- Han, Y.; Cutler, J.E. Antibody response that protects against disseminated candidiasis. Infect. Immun. 1995, 63, 2714–2719. [Google Scholar] [CrossRef]
- Han, Y.; Riesselman, M.H.; Cutler, J.E. Protection against candidiasis by an immunoglobulin G3 (IgG3) monoclonal antibody specific for the same mannotriose as an IgM protective antibody. Infect. Immun. 2000, 68, 1649–1654. [Google Scholar] [CrossRef]
- Zhang, M.X.; Bohlman, M.C.; Itatani, C.; Burton, D.R.; Parren, P.W.; St Jeor, S.C.; Kozel, T.R. Human recombinant antimannan immunoglobulin G1 antibody confers resistance to hematogenously disseminated candidiasis in mice. Infect. Immun. 2006, 74, 362–369. [Google Scholar] [CrossRef]
- Nishiya, C.T.; Boxx, G.M.; Robison, K.; Itatani, C.; Kozel, T.R.; Zhang, M.X. Influence of IgG Subclass on Human Antimannan Antibody-Mediated Resistance to Hematogenously Disseminated Candidiasis in Mice. Infect. Immun. 2015, 84, 386–394. [Google Scholar] [CrossRef]
- Cywes-Bentley, C.; Skurnik, D.; Zaidi, T.; Roux, D.; Deoliveira, R.B.; Garrett, W.S.; Lu, X.; O’Malley, J.; Kinzel, K.; Zaidi, T.; et al. Antibody to a conserved antigenic target is protective against diverse prokaryotic and eukaryotic pathogens. Proc. Natl. Acad. Sci. USA 2013, 110, E2209–E2218. [Google Scholar] [CrossRef]
- Kavishwar, A.; Shukla, P.K. Candidacidal activity of a monoclonal antibody that binds with glycosyl moieties of proteins of Candida albicans. Med. Mycol. 2006, 44, 159–167. [Google Scholar] [CrossRef][Green Version]
- De Bernardis, F.; Boccanera, M.; Adriani, D.; Spreghini, E.; Santoni, G.; Cassone, A. Protective role of antimannan and anti-aspartyl proteinase antibodies in an experimental model of Candida albicans vaginitis in rats. Infect. Immun. 1997, 65, 3399–3405. [Google Scholar] [CrossRef] [PubMed]
- Viudes, A.; Lazzell, A.; Perea, S.; Kirkpatrick, W.R.; Peman, J.; Patterson, T.F.; Martinez, J.P.; López-Ribot, J.L. The C-terminal antibody binding domain of Candida albicans mp58 represents a protective epitope during candidiasis. FEMS Microbiol. Lett. 2004, 232, 133–138. [Google Scholar] [CrossRef]
- Sevilla, M.J.; Robledo, B.; Rementeria, A.; Moragues, M.D.; Pontón, J. A fungicidal monoclonal antibody protects against murine invasive candidiasis. Infect. Immun. 2006, 74, 3042–3045. [Google Scholar] [CrossRef]
- Xin, H. Active immunizations with peptide-DC vaccines and passive transfer with antibodies protect neutropenic mice against disseminated candidiasis. Vaccine 2016, 34, 245–251. [Google Scholar] [CrossRef]
- Liu, Y.; Filler, S.G. Candida albicans, Als3., a multifunctional adhesin and invasin. Eukaryot. Cell 2011, 10, 168–173. [Google Scholar] [CrossRef]
- Moragues, M.D.; Omaetxebarria, M.J.; Elguezabal, N.; Sevilla, M.J.; Conti, S.; Polonelli, L.; Pontón, J. A monoclonal antibody directed against a Candida albicans cell wall mannoprotein exerts three anti-C. albicans activities. Infect. Immun. 2003, 71, 5273–5279. [Google Scholar] [CrossRef]
- Brena, S.; Cabezas-Olcoz, J.; Moragues, M.D.; Fernández de Larrinoa, I.; Domínguez, A.; Quindós, G.; Pontón, J. Fungicidal monoclonal antibody C7 interferes with iron acquisition in Candida albicans. Antimicrob. Agents Chemother. 2011, 55, 3156–3163. [Google Scholar] [CrossRef]
- Luo, G.; Ibrahim, A.S.; French, S.W.; Edwards, J.E., Jr.; Fu, Y. Active and passive immunization with rHyr1p-N protects mice against hematogenously disseminated candidiasis. PLoS ONE 2011, 6, e25909. [Google Scholar] [CrossRef]
- Rudkin, F.M.; Raziunaite, I.; Workman, H.; Essono, S.; Belmonte, R.; MacCallum, D.M.; Johnson, E.M.; Silva, L.M.; Palma, A.S.; Feizi, T.; et al. Single human B cell-derived monoclonal anti-Candida antibodies enhance phagocytosis and protect against disseminated candidiasis. Nat. Commun. 2018, 9, 5288. [Google Scholar] [CrossRef]
- Schaller, M.; Bein, M.; Korting, H.C.; Baur, S.; Hamm, G.; Monod, M.; Beinhauer, S.; Hube, B. The secreted aspartyl proteinases Sap1 and Sap2 cause tissue damage in an in vitro model of vaginal candidiasis based on reconstituted human vaginal epithelium. Infect. Immun. 2003, 71, 3227–3234. [Google Scholar] [CrossRef]
- Schmitt, M.J.; Breinig, F. Yeast viral killer toxins: Lethality and self-protection. Nature Rev. Microbiol. 2006, 4, 212–221. [Google Scholar] [CrossRef]
- Magliani, W.; Conti, S.; de Bernardis, F.; Gerloni, M.; Bertolotti, D.; Mozzoni, P.; Cassone, A.; Polonelli, L. Therapeutic potential of antiidiotypic single chain antibodies with yeast killer toxin activity. Nat. Biotechnol. 1997, 15, 155–158. [Google Scholar] [CrossRef]
- Sanford, J.E.; Lupan, D.M.; Schlageter, A.M.; Kozel, T.R. Passive immunization against Cryptococcus neoformans with an isotype-switch family of monoclonal antibodies reactive with cryptococcal polysaccharide. Infect. Immun. 1990, 58, 1919–1923. [Google Scholar] [CrossRef]
- Zebedee, S.L.; Koduri, R.K.; Mukherjee, J.; Mukherjee, S.; Lee, S.; Sauer, D.F.; Scharff, M.D.; Casadevall, A. Mouse-human immunoglobulin G1 chimeric antibodies with activities against Cryptococcus neoformans. Antimicrob. Agents Chemother. 1994, 38, 1507–1514. [Google Scholar] [CrossRef]
- Rosas, A.L.; Nosanchuk, J.D.; Casadevall, A. Passive immunization with melanin-binding monoclonal antibodies prolongs survival of mice with lethal Cryptococcus neoformans infection. Infect. Immun. 2001, 69, 3410–3412. [Google Scholar] [CrossRef] [PubMed]
- Beenhouwer, D.O.; Yoo, E.M.; Lai, C.W.; Rocha, M.A.; Morrison, S.L. Human immunoglobulin G2 (IgG2) and IgG4, but not IgG1 or IgG3, protect mice against Cryptococcus neoformans infection. Infect. Immun. 2007, 75, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.L.; Shi, L.; Barreto-Bergter, E.; Nimrichter, L.; Farias, S.E.; Rodrigues, E.G.; Travassos, L.R.; Nosanchuk, J.D. Monoclonal antibody to fungal glucosylceramide protects mice against lethal Cryptococcus neoformans infection. Clin. Vaccine Immunol. 2007, 14, 1372–1376. [Google Scholar] [CrossRef]
- Xin, H.; Cutler, J.E. Vaccine and monoclonal antibody that enhance mouse resistance to candidiasis. Clin. Vaccine Immunol. 2011, 18, 1656–1667. [Google Scholar] [CrossRef]
- Nosanchuk, J.D.; Steenbergen, J.N.; Shi, L.; Deepe, G.S., Jr.; Casadevall, A. Antibodies to a cell surface histone-like protein protect against Histoplasma capsulatum. J. Clin. Investig. 2003, 112, 1164–1175. [Google Scholar] [CrossRef]
- Guimarães, A.J.; Frases, S.; Gomez, F.J.; Zancopé-Oliveira, R.M.; Nosanchuk, J.D. Monoclonal antibodies to heat shock protein 60 alter the pathogenesis of Histoplasma capsulatum. Infect. Immun. 2009, 77, 1357–1367. [Google Scholar] [CrossRef]
- Lopes, L.C.; Guimarães, A.J.; de Cerqueira, M.D.; Gómez, B.L.; Nosanchuk, J.D. A histoplasma capsulatum-specific IgG1 isotype monoclonal antibody, H1C, to a 70-kilodalton cell surface protein is not protective in murine histoplasmosis. Clin. Vaccine Immunol. 2010, 17, 1155–1158. [Google Scholar] [CrossRef] [PubMed]
- Nosanchuk, J.D.; Zancopé-Oliveira, R.M.; Hamilton, A.J.; Guimarães, A.J. Antibody therapy for histoplasmosis. Front. Microbiol. 2012, 3, 21. [Google Scholar] [CrossRef] [PubMed]
- De Mattos Grosso, D.; de Almeida, S.R.; Mariano, M.; Lopes, J.D. Characterization of gp70 and anti-gp70 monoclonal antibodies in Paracoccidioides brasiliensis pathogenesis. Infect. Immun. 2003, 71, 6534–6542. [Google Scholar] [CrossRef] [PubMed]
- Xander, P.; Vigna, A.F.; Feitosa, L.; dos, S.; Pugliese, L.; Bailao, A.M.; Soares, C.M.; Mortara, R.A.; Mariano, M.; Lopes, J.D. A surface 75-kDa protein with acid phosphatase activity recognized by monoclonal antibodies that inhibit Paracoccidioides brasiliensis growth. Microbes Infect. 2007, 9, 1484–1492. [Google Scholar] [CrossRef] [PubMed]
- Buissa-Filho, R.; Puccia, R.; Marques, A.F.; Pinto, F.A.; Muñoz, J.E.; Nosanchuk, J.D.; Travassos, L.R.; Taborda, C.P. The monoclonal antibody against the major diagnostic antigen of Paracoccidioides brasiliensis mediates immune protection in infected BALB/c mice challenged intratracheally with the fungus. Infect. Immun. 2008, 76, 3321–3328. [Google Scholar] [CrossRef]
- Ferreira, K.S.; Maranhão, A.Q.; Garcia, M.C.; Brígido, M.M.; Santos, S.S.; Lopes, J.D.; Almeida, S.R. Dendritic cells transfected with scFv from Mab 7.B12 mimicking original antigen gp43 induces protection against experimental Paracoccidioidomycosis. PLoS ONE 2011, 6, e15935. [Google Scholar] [CrossRef]
- Thomaz, L.; Nosanchuk, J.D.; Rossi, D.C.; Travassos, L.R.; Taborda, C.P. Monoclonal antibodies to heat shock protein 60 induce a protective immune response against experimental Paracoccidioides lutzii. Microbes Infect. 2014, 16, 788–795. [Google Scholar] [CrossRef]
- Nascimento, R.C.; Espindola, N.M.; Castro, R.A.; Teixeira, P.A.C.; Penha, C.V.L.Y.; Lopes-Bezerra, L.M.; Almeida, S.R. Passive immunization with monoclonal antibody against a 70-kDa putative adhesin of Sporothrix schenckii induces protection in murine sporotrichosis. Eur. J. Immunol. 2008, 38, 3080–3089. [Google Scholar] [CrossRef]
- De Almeida, J.R.; Kaihami, G.H.; Jannuzzi, G.P.; de Almeida, S.R. Therapeutic vaccine using a monoclonal antibody against a 70-kDa glycoprotein in mice infected with highly virulent Sporothrix schenckii and Sporothrix brasiliensis. Med. Mycol. 2015, 53, 42–50. [Google Scholar] [CrossRef]
- De Almeida, J.R.; Santiago, K.L.; Kaihami, G.H.; Maranhão, A.Q.; de Macedo Brígido, M.; de Almeida, S.R. The Efficacy of Humanized Antibody against the Sporothrix Antigen, gp70, in Promoting Phagocytosis and Reducing Disease Burden. Front. Microbiol. 2017, 8, 345. [Google Scholar] [CrossRef]
- Frosco, M.B.; Chase, T., Jr.; Macmillan, J.D. The effect of elastase-specific monoclonal and polyclonal antibodies on the virulence of Aspergillus fumigatus in immunocompromised mice. Mycopathologia 1994, 125, 65–76. [Google Scholar] [CrossRef] [PubMed]
- Cenci, E.; Mencacci, A.; Spreca, A.; Montagnoli, C.; Bacci, A.; Perruccio, K.; Velardi, A.; Magliani, W.; Conti, S.; Polonelli, L.; et al. Protection of killer antiidiotypic antibodies against early invasive aspergillosis in a murine model of allogeneic T-cell-depleted bone marrow transplantation. Infect. Immun. 2002, 70, 2375–2382. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Kavishwar, A.; Shiva Keshava, G.B.; Shukla, P.K. Monoclonal immunoglobulin G1 directed against Aspergillus fumigatus cell wall glycoprotein protects against experimental murine aspergillosis. Clin. Diagn. Lab. Immunol. 2005, 12, 1063–1068. [Google Scholar] [CrossRef] [PubMed]
- Appel, E.; Vallon-Eberhard, A.; Rabinkov, A.; Brenner, O.; Shin, I.; Sasson, K.; Shadkchan, Y.; Osherov, N.; Jung, S.; Mirelman, D. Therapy of murine pulmonary aspergillosis with antibody-alliinase conjugates and alliin. Antimicrob. Agents Chemother. 2010, 54, 898–906. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Heesemann, L.; Kotz, A.; Echtenacher, B.; Broniszewska, M.; Routier, F.; Hoffmann, P.; Ebel, F. Studies on galactofuranose-containing glycostructures of the pathogenic mold Aspergillus fumigatus. Int. J. Med. Microbiol. 2011, 321, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Wharton, R.E.; Stefanov, E.K.; King, R.G.; Kearney, J.F. Antibodies generated against Streptococci protect in a mouse model of disseminated aspergillosis. J. Immunol. 2015, 194, 4387–4396. [Google Scholar] [CrossRef]
- Chauvin, D.; Hust, M.; Schütte, M.; Chesnay, A.; Parent, C.; Moreira, G.M.S.G.; Arroyo, J.; Sanz, A.B.; Pugnière, M.; Martineau, P.; et al. Targeting Aspergillus fumigatus Crf Transglycosylases With Neutralizing Antibody Is Relevant but Not Sufficient to Erase Fungal Burden in a Neutropenic Rat Model. Front. Microbiol. 2019, 10, 600. [Google Scholar] [CrossRef]
- Yadav, R.K.; Shukla, P.K. A novel monoclonal antibody against enolase antigen of Aspergillus fumigatus protects experimental aspergillosis in mice. FEMS Microbiol. Lett. 2019, 366, fnz015. [Google Scholar] [CrossRef]
- Gebremariam, T.; Alkhazraji, S.; Soliman, S.S.M.; Gu, Y.; Jeon, H.H.; Zhang, L.; French, S.W.; Stevens, D.A.; Edwards, J.E., Jr.; Filler, S.G.; et al. Anti-CotH3 antibodies protect mice from mucormycosis by prevention of invasion and augmenting opsonophagocytosis. Sci. Adv. 2019, 5, eaaw1327. [Google Scholar] [CrossRef]
- Lopes, L.C.; Rollin-Pinheiro, R.; Guimarães, A.J.; Bittencourt, V.C.; Martinez, L.R.; Koba, W.; Farias, S.E.; Nosanchuk, J.D.; Barreto-Bergter, E. Monoclonal antibodies against peptidorhamnomannans of Scedosporium apiospermum enhance the pathogenicity of the fungus. PLoS Negl. Trop. Dis. 2010, 4, e853. [Google Scholar] [CrossRef]
- Liedke, S.C.; Miranda, D.Z.; Gomes, K.X.; Gonçalves, J.L.S.; Frases, S.; Nosanchuk, J.D.; Rodrigues, M.L.; Nimrichter, L.; Peralta, J.M.; Guimarães, A.J. Characterization of the antifungal functions of a WGA-Fc (IgG2a) fusion protein binding to cell wall chitin oligomers. Sci. Rep. 2017, 7, 12187. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Albuquerque, P.C.; Lazar-Molnar, E.; Wang, X.; Santambrogio, L.; Gácser, A.; Nosanchuk, J.D. A monoclonal antibody to Histoplasma capsulatum alters the intracellular fate of the fungus in murine macrophages. Eukaryot. Cell 2008, 7, 1109–1117. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, A.K.; Kumar, R.; Kumar, A.; Shukla, P.K. A monoclonal IgM directed against immunodominant catalase B of cell wall of Aspergillus fumigatus exerts anti-A. fumigatus activities. Mycoses 2009, 52, 524–533. [Google Scholar] [CrossRef] [PubMed]
- Gebremariam, T.; Liu, M.; Luo, G.; Bruno, V.; Phan, Q.T.; Waring, A.J.; Edwards, J.E.; Filler, S.G.; Yeaman, M.R.; Ibrahim, A.S. CotH3 mediates fungal invasion of host cells during mucormycosis. J. Clin. Investig. 2014, 124, 237–250. [Google Scholar] [CrossRef] [PubMed]
- Jannuzzi, G.P.; Souza, N.A.; Françoso, K.S.; Pereira, R.H.; Santos, R.P.; Kaihami, G.H.; Almeida, J.R.F.; Batista, W.L.; Amaral, A.C.; Maranhão, A.Q.; et al. Therapeutic treatment with scFv-PLGA nanoparticles decreases pulmonary fungal load in a murine model of paracoccidioidomycosis. Microbes Infect. 2018, 20, 48–56. [Google Scholar] [CrossRef]
- Gazendam, R.P.; van Hamme, J.L.; Tool, A.T.; Hoogenboezem, M.; van den Berg, J.M.; Prins, J.M.; Vitkov, L.; van de Veerdonk, F.L.; van den Berg, T.K.; Roos, D.; et al. Human Neutrophils Use Different Mechanisms To Kill Aspergillus fumigatus Conidia and Hyphae: Evidence from Phagocyte Defects. J. Immunol. 2016, 196, 1272–1283. [Google Scholar] [CrossRef]
- Braem, S.G.; Rooijakkers, S.H.; van Kessel, K.P.; de Cock, H.; Wösten, H.A.; van Strijp, J.A.; Haas, P.J. Effective Neutrophil Phagocytosis of Aspergillus fumigatus Is Mediated by Classical Pathway Complement Activation. J. Innate Immun. 2015, 7, 364–374. [Google Scholar] [CrossRef]
- Stuehler, C.; Khanna, N.; Bozza, S.; Zelante, T.; Moretti, S.; Kruhm, M.; Lurati, S.; Conrad, B.; Worschech, E.; Stevanović, S.; et al. Cross-protective TH1 immunity against Aspergillus fumigatus and Candida albicans. Blood 2011, 117, 5881–5891. [Google Scholar] [CrossRef]
- Farmakiotis, D.; Kontoyiannis, D.P. Mucormycoses. Infect. Dis. Clin. N. Am. 2016, 30, 143–163. [Google Scholar] [CrossRef]


{kind=link}
| Antigen | Infection Model | Application of mAbs # | Protective | Non-Protective | Reference |
|---|---|---|---|---|---|
| GXM | m., i.v. | i.p./24 h/10–100 µg | IgG1 | [4] | |
| capsular polysaccharide | m., i.v. | i.p./−24 h/1 mg i.p./48 h +96 h/500 µg | IgG1 *, IgG2a, IgG2b * IgG1 *, IgG2a, IgG2b * | [53] | |
| GXM | m., i.p. | i.p./0 h/1 mg | IgA, IgG1 > IgM > IgG3 | [7] | |
| GXM | m., i.v. | i.p./−4 h/1 mg | chIgG1 | [54] | |
| GXM | m., i.p. | i.p./−15 min/1 mg | two IgM | one IgM | [13] |
| GXM | m., i.v. | i.p./−24 h/1 mg | IgG1, IgG2b, IgG2a > IgG3 | [6] | |
| GXM | m., i.v. | preincubation of yeasts with mAb | hIgM | [5] | |
| melanin | m., i.v. | i.v./−30 min/1 mg | IgM | [55] | |
| GXM | m., i.v. m., i.p. m., i.t. | i.p./−5, −30 min/0.1, 0.5 or 1 mg i.p./−5, −30 min/0.1, 0.5 or 1 mg i.p./−5, −30 min/0.1, 0.5 or 1 mg | IgM 12A1, dose-dependent | IgMs 12A1 and 13F1 IgM 13F1 IgMs 12A1 and 13F1 | [12] |
| GXM | m., preincubation of yeasts | i.p./−24 h/1 mg | IgG1 *, IgG2a *, IgG2b * | IgG3 * | [16] |
| GXM | m., i.p. | i.p./−1 h/0.5, 5, 50, 100, 1000 µg | one hIgM (at 100 µg) | two hIgM | [14] |
| GXM | m., i.v. | i.v./10 d/500 µg | chIgG2 | [20] | |
| GXM | m., i.v. | i.p./−18 h/0.1–1 mg | recomb. h-IgG2 *, h-IgG4 * | recomb. h-IgG1 *, h-IgG3 * | [56] |
| glucosylceramide | m., i.t. | i.p./−24 h/100, 250, 500 µg | IgG2b at 500 µg | IgG2b at 100 and 250 µg | [57] |
| β-glucan | m., i.v. | i.p./−2 h, +1 d/200 µg | IgG2 | [29] | |
| GXM M2 motif | m., i.v. | i.p./−30 min/500 µg | IgA, IgM | [15] |
| Antigen | Infection Model | Application of mAbs # | Protective | Non-Protective | Reference |
|---|---|---|---|---|---|
| Hsp90 | m., i.v. | i.v./−1 h/740 µg | IgG | [24] | |
| β-1,2-linked mannotriose | m., i.v. | i.p./−4 h, 20 h/125 µg | IgM | [35] [35] | |
| polysaccharide mannoprotein SAP2 | r., i.vg. r., i.vg. r., i.vg. | i.v./30 min/100 µg/mL i.v./30 min/100 µg/mL i.v./30 min/100 µg/mL | IgM IgG1 | IgG1 | [41] |
| β-1,2-linked mannotriose | m., i.vg. | i.p./−4 h, 24 h /35 µg, 10 µg i.vg./−4 h, 24 h/35 µg, 10 µg | IgM IgM | [34] | |
| antiidiotypic KT antibody | r., i.v. | i.v./0 h/10 µg | single chain antibody | [52] | |
| β-1,2-linked mannotriose | m., i.v. m., i.vg. | i.p./−4 h/125 µg i.vg./−4 h/10 µg | IgM, IgG3 | [36] | |
| Hsp90 | m., i.v. | i.v./2 h/2 mg/kg | recomb. h-IgG | [25] | |
| PRA1 (mannoprotein 58) | m., i.v. | i.p./−2 h/1.8 mg | IgG1 | [42] | |
| β-1,3-glucan | m., i.v. | i.p./−2 h/250 µg | IgG2 | [28] | |
| ALS3 mannoprotein | m., i.v. | i.p./−4 h, 1 d, 2 d/200, 100, 100 µg | IgM | [43] | |
| mannan | m., i.v. | i.p./−4 h/63 µg–4 mg | h-IgG1 | [37] | |
| cell wall carbohydrate | m., i.v. | i.v./−2 h/100 µg | IgA | [40] | |
| β-1,3-glucan | m., i.v. | i.p./−2 h/100 µg | IgG2 * | IgM * | [30] |
| Fba peptide | m., i.v. | i.p./−4 h/8 µg | IgM | [58] | |
| β-1,3-glucan | m., i.v. r., i.v. | i.p./−2 h/100 µg i.v./1 h + 24 h + 48 h/50 µg | scFv-Fc | [31] | |
| PNAG | m., keratitis model | i.p./24 h/200 µg | hIgG1 | [39] | |
| Fba peptide Met6 peptide | m., i.v. m., i.v. | i.p./−4 h, every day/100 µg i.p./−4 h. every day/250 µg | IgM IgG3 | [44] | |
| mannan | m., | i.p./−4 h/1 mg | hIgG1, hIgG3, hIgG4 | hIgG2 | [38] |
| Unknown surface antigen HYR1 protein | m., i.v. m., i.v. | i.p./−4 h/1 mg i.p./−4 h/1 mg | h-IgG1 h-IgG1 | [49] | |
| β-1,3-glucan | m., i.v. | i.p./−2 h/150 µg | IgG1, IgG3 | [32] |
| Antigen | Infection Model | Application of mAbs # | Protective | Non-Protective | Reference |
|---|---|---|---|---|---|
| Histoplasma capsulatum | |||||
| histone 2b-like protein | m., i.n. | i.p./−2 h/100 µg | IgM | [59] | |
| Hsp60 | m., i.n. | i.p./−2 h/500 µg | IgG1 **, IgG2a | IgG2b ** | [60] |
| 70 kDa surface protein | m., i.n. | i.p./−2 h/100–500 µg | IgG1 | [61] | |
| chitin | m., i.n. | i.p./−2 h/10 µg | WGA-Fc (IgG2a) | [62] | |
| Paracoccidioides brasiliensis | |||||
| glycoprotein of 70 kDa (gp70) | m., i.t. | i.v./−3 d, 3 d, 6 d, 9 d, 42 d/100 µg each | combination of two IgG1 | [63] | |
| 75 kDa secreted phosphatase | m., i.t. | i.v./−3 d/100 µg | IgG, IgM | [64] | |
| glycoprotein of 43 kDa (gp43) | m., i.t. | i.p./30 d/1 mg | IgG2b | [65] | |
| gp43 | m., i.t. | i.m./14 d, 21 d/DCs expressing the scFv s | scFv | [66] | |
| m., i.t. | |||||
| Paracoccidioides lutzii | |||||
| Heat shock protein 60 | m., i.t. | not sp./−24 h/1 mg | IgG2a, IgG2b | [67] | |
| Sporothrix schenckii | |||||
| 70 kDa glycoprotein | m., i.p. | i.p./−24 h, 3 d, 6 d, 42 d/100 µg | IgG1 | [68] | |
| 70 kDa glycoprotein | m., i.p. | i.p./3 d, 10 d/100 µg | IgG1 | [69] | |
| 70 kDa glycoprotein | m., i.p. | not sp./3 d/100 µg | hIgG1 | [70] |
| Antigen | Infection Model | Application of mAbs # | Protective | Non-Protective | Reference |
|---|---|---|---|---|---|
| Aspergillus fumigatus | |||||
| elastase | m., i.n. | i.p./4 h/50 µg | isotype not sp. | [71] | |
| antiidiotypic KT antibody | m., i.n. | i.n./each day/2 × 1 µg | rat IgM | [72] | |
| cell wall glycoprotein | m., i.v. | i.v./−2 h/50 µg | IgG1 | [73] | |
| unknown cell wall antigen | m., i.n. | i.t./1 h/50 nmol | IgM + alliinase | [74] | |
| galactomannan | m., i.v. | i.p./−15 min/200 µg | IgM | [75] | |
| sialylated oligosaccharides | m., i.v. m., i.t. | i.v/0 min/200 µg i.t./0 min/50 µg | IgM | [76] | |
| Crf1 protein | r., i.t. | i.t./4 mg/kg/0 h + 32 h | h-IgG1 | [77] | |
| enolase | m., i.v. | i.v./2 h/50 µg | IgM | [78] | |
| Rhizopus delemar | |||||
| CotH3 protein | m., i.t. | i.p./48 h/30 µg | IgG1 | [79] | |
| Scedosporium apiospermum | |||||
| peptidorhamnomannan | m., i. t. | i.p./−2 h/250 µg | IgG1 | [80] |
| Antibody | Antigen | Subclass | Fungus | Antifungal Activity | Mode of Action | Reference |
|---|---|---|---|---|---|---|
| Mycograb | Hsp90 | rec. mAb | Candida albicans | stress resistance ↓ | inhibition of Hsp90 | [25] |
| C7 | ALS3 mannoprotein | IgM | Candida albicans Candia lusitaniae Cryptococcus neoformans Aspergillus fumigatus Scedosporium proliferans | growth inhibition, adhesion to HEp2 cells ↓ | reduced iron acquisition | [46] [47] |
| 2G8 | β-glucan | IgG2b | Candida albicans | growth inhibition | unknown | [28,30] |
| G5 | cell wall carbohydrate | IgA | Candida albicans | growth inhibition | unknown | [40] |
| 5H5 | β-1,3-glucan | IgG3 | Candida albicans Aspergillus fumigatus | growth inhibition, phagocytosis ↑ | unknown, osponization | [32] |
| M1g1 | mannan | h-IgG1 | Candida albicans | phagocytosis ↑, killing ↑ | complement binding ↑ | [37] |
| 2G8 scFv-Fc | β-glucan | scFv-Fc | Candida albicans | neutrophil mediated killing ↑ | osponization | [31] |
| 6D2, 11B11 | melanin | IgM | Cryptococcus neoformans | growth inhibition | unknown | [55] |
| 12A1 | glucuronoxylomannan | IgM | Cryptococcus neoformans | phagocytosis ↑ | opzonisation | [12] |
| recomb. 3E5 | GXM | IgG1, IgG3 | Cryptococcus neoformans | phagocytosis ↑ | opzonisation | [56] |
| recomb. 3E5 | GXM | IgG1, IgG3 | Cryptococcus neoformans | phagocytosis ↑ | opzonisation | [8] |
| 2G8 | β-glucan | IgG2b | Cryptococcus neoformans | growth inhibition, phagocytosis ↑ | unknown, | [29] |
| WGA-Fc | chitin | (IgG2a) | Cryptococcus neoformans | growth inhibition, phagocytosis ↑ | unknown, opsonization | [81] |
| 4E12 | Hsp60 | IgG2a | Histoplasma capsulatum | phagocytosis ↑ | opsonization | [60] |
| 9C7 | histone 2b-like protein | IgM | Histoplasma capsulatum | phagocytosis ↑, phagosomal maturation ↑ | opsonization | [59] [82] |
| MS112-IIB1 | Crf1, glycosylhydrolase | hum. IgG1 | Aspergillus fumigatus | growth inhibition | inhibition of enzymatic activity | [77] |
| R-5 | enolase | IgM | Aspergillus fumigatus | growth inhibition | unknown | [78] |
| 7 | catalase B | IgM | Aspergillus fumigatus | growth inhibition | unknown | [83] |
| 2G8 | β-1,3-glucan | IgG2b | Aspergillus fumigatus | hyphal growth ↓, adherence to epithelial cell ↓ | unknown | [28,30] |
| 3G11 | β-1,3-glucan | IgG1 | Aspergillus fumigatus | inhibition of germination, phagocytosis ↑ | unknown, opsonization | [32] |
| C1, C2, C3 | CotH3 protein | Rhizopus delemar | phagocytosis ↑, cytokine response ↑ | opsonization | [84] | |
| 3E | Gp43 | IgG2b | Paracoccidioides brasiliensis | phagocytosis ↑, NO ↑, IFNγ ↑ | opsonization | [65] |
| 1G6, 5E7C | 75 kDa phosphatase | IgG, IgM | Paracoccidioides brasiliensis | phagocytosis ↑, growth inhibition | opsonization | [64] |
| 7B6, 4E12 | Hsp60 | IgG2a, IgG2b | Paracoccidioides lutzii | phagocytosis ↑ | opsonization | [67] |
| P6E7 | Gp70 | Sporothrix spp. | phagocytosis ↑ | opsonization | [70] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ulrich, S.; Ebel, F. Monoclonal Antibodies as Tools to Combat Fungal Infections. J. Fungi 2020, 6, 22. https://doi.org/10.3390/jof6010022
Ulrich S, Ebel F. Monoclonal Antibodies as Tools to Combat Fungal Infections. Journal of Fungi. 2020; 6(1):22. https://doi.org/10.3390/jof6010022
Chicago/Turabian StyleUlrich, Sebastian, and Frank Ebel. 2020. "Monoclonal Antibodies as Tools to Combat Fungal Infections" Journal of Fungi 6, no. 1: 22. https://doi.org/10.3390/jof6010022
APA StyleUlrich, S., & Ebel, F. (2020). Monoclonal Antibodies as Tools to Combat Fungal Infections. Journal of Fungi, 6(1), 22. https://doi.org/10.3390/jof6010022