
Glucose Catabolite Repression Participates in the Regulation of Sialidase Biosynthesis by Antarctic Strain Penicillium griseofulvum P29

 , , ,
, , ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Enzyme Activity Determination
2.3. RNA Isolation, Copy-DNA Synthesis, and Real-Time RT PCR
2.4. Analytical Methods
2.5. Statistical Evaluation of the Results
3. Results
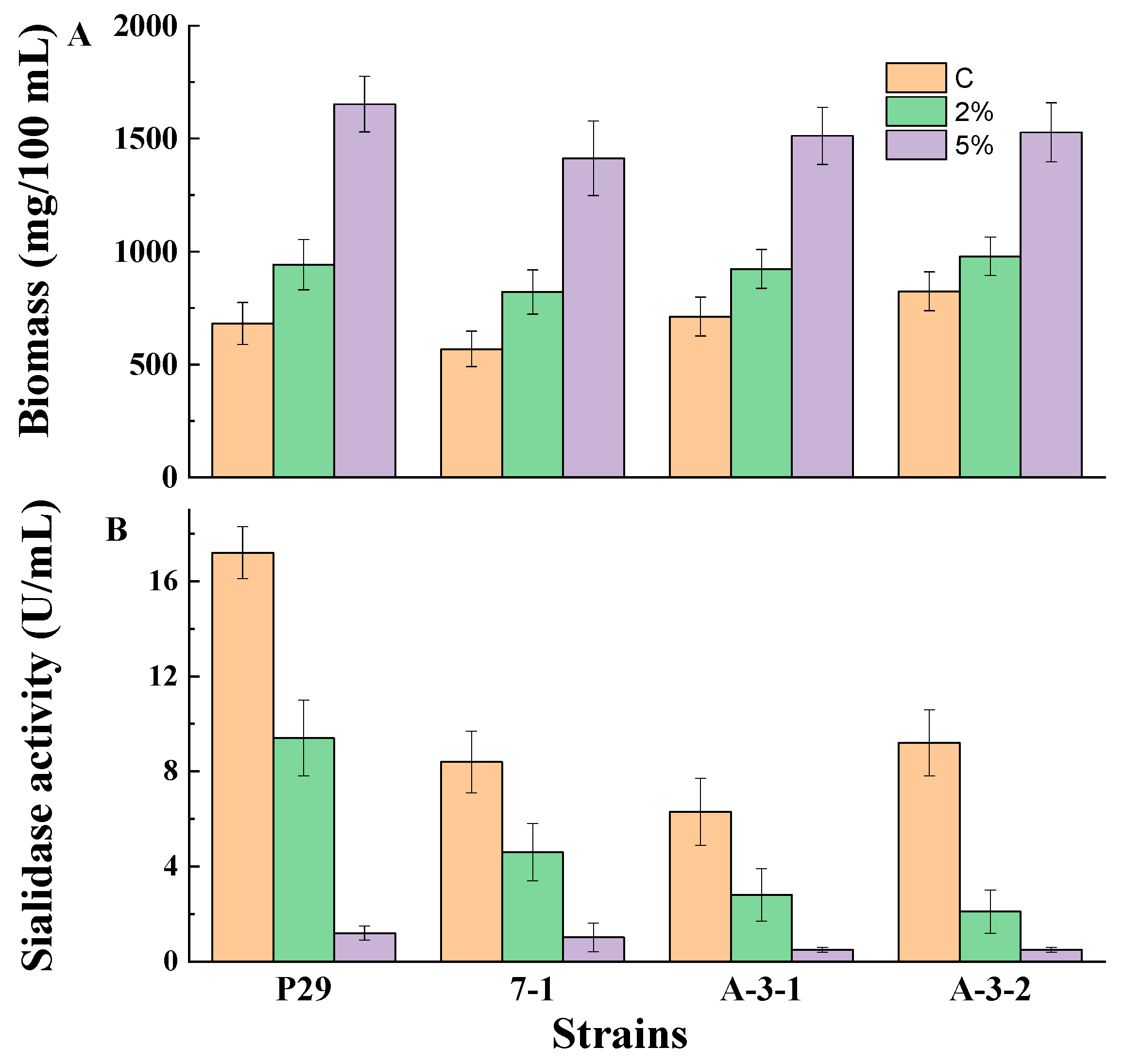
3.1. Glucose-Mediated Repression of Sialidase Production by Growing Fungal Mycelia
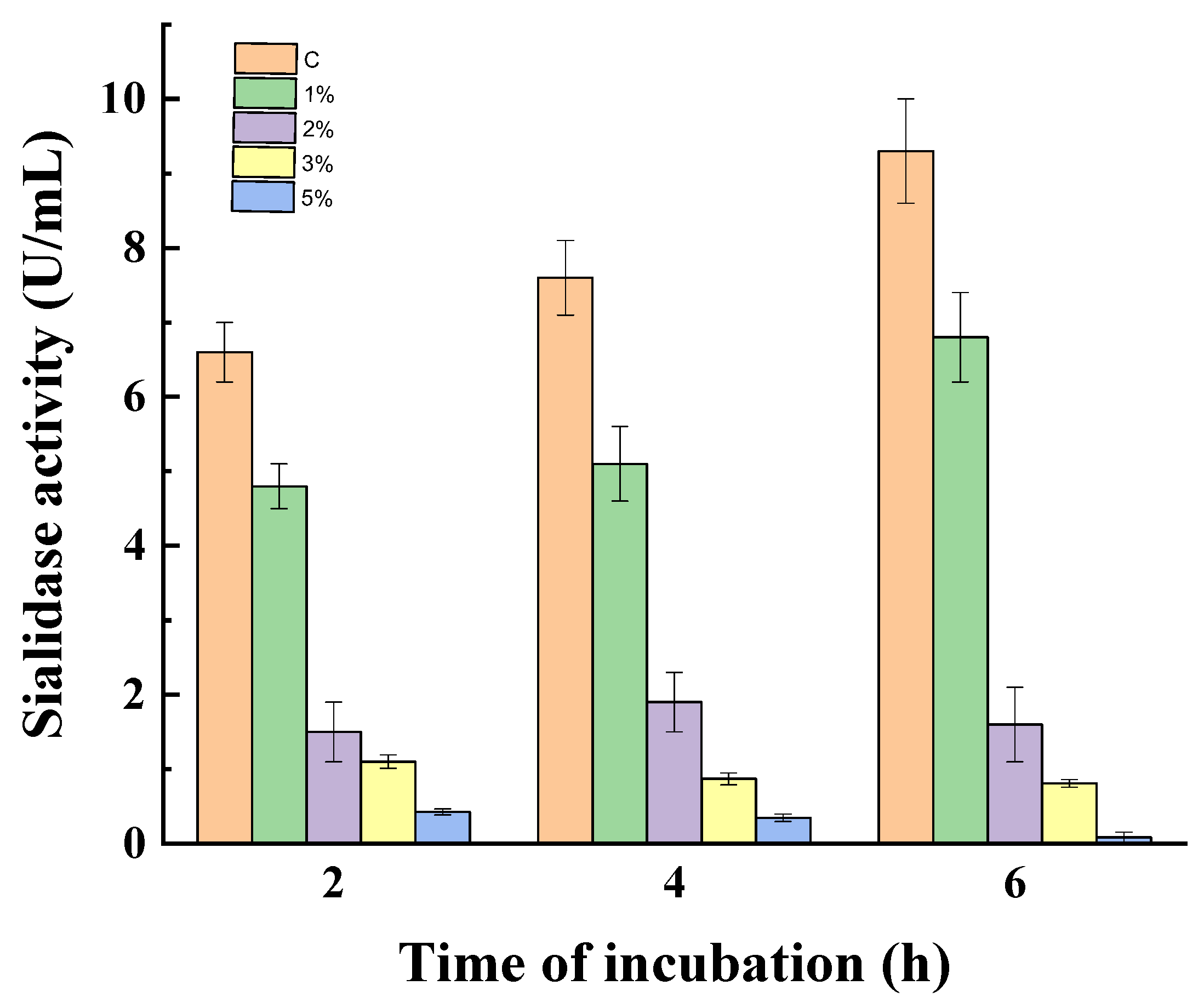
3.2. Effect of Catabolite Repression on Non-Growing Mycelia
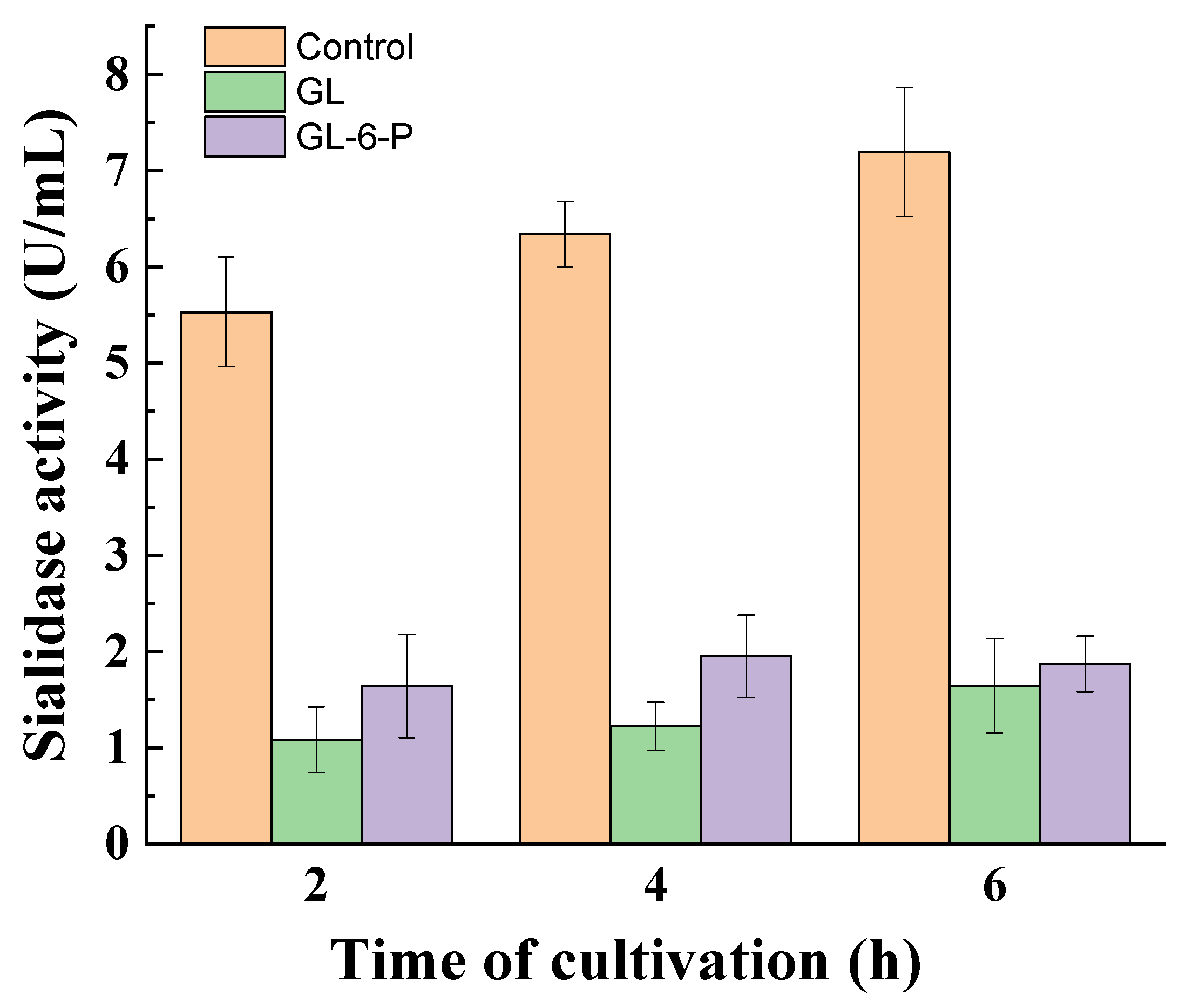
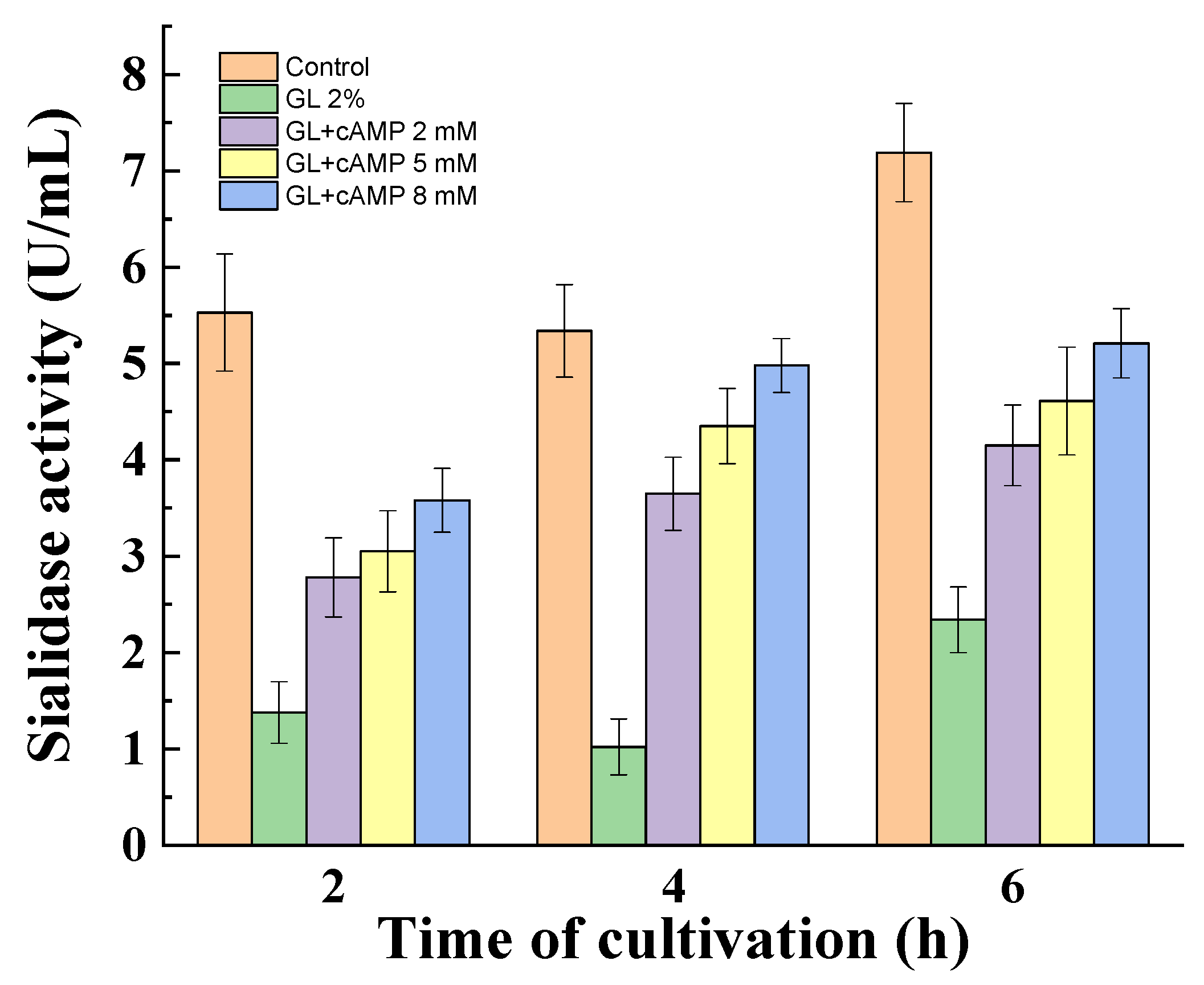
3.3. Influence of Catabolite Repression Regulators
3.4. Comparative Sialidase Gene Expression in the Presence of Glucose
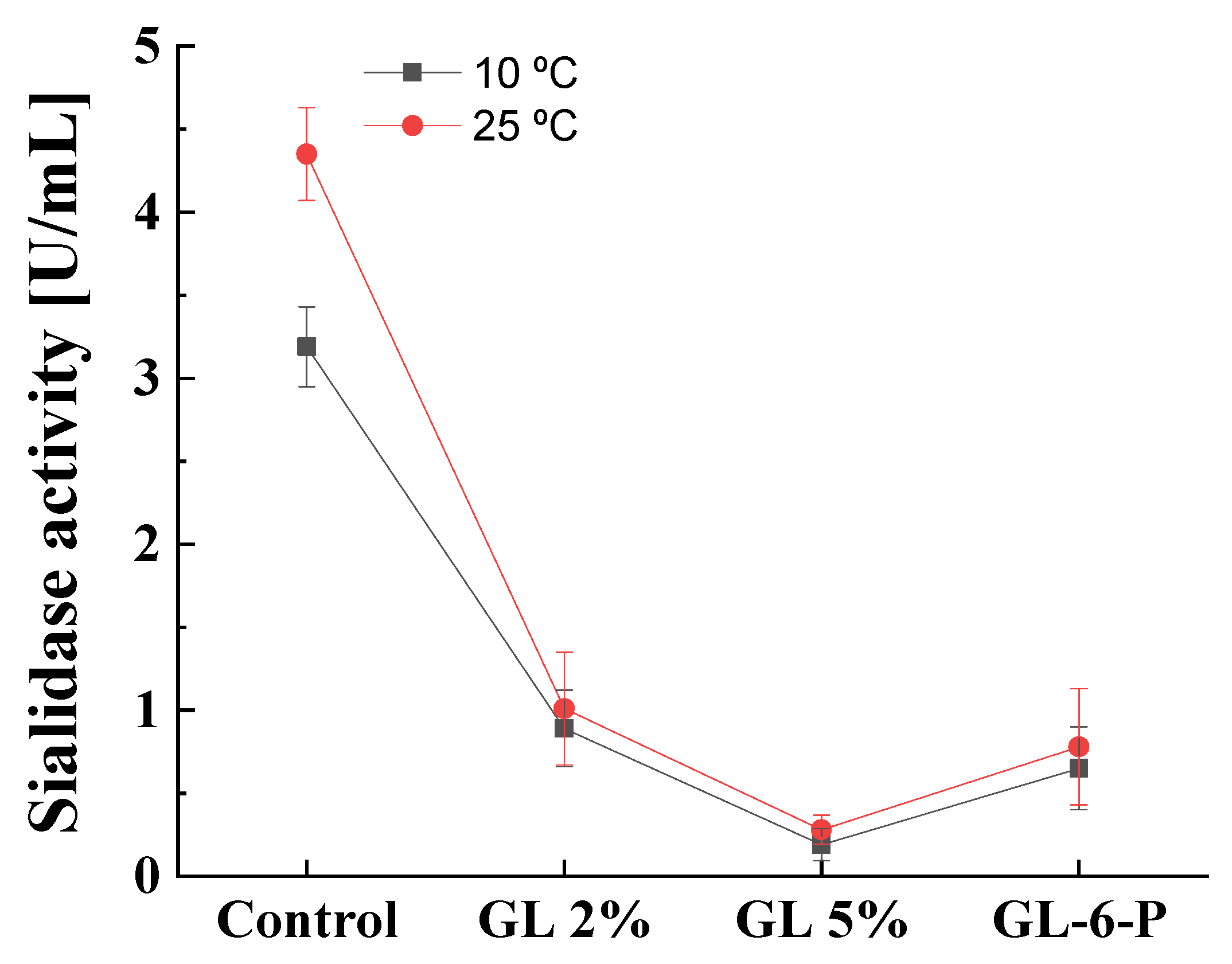
3.5. Effect of the Temperature on Catabolite Repression of Cold-Active Sialidase
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, Y. Sialidases from Clostridium perfringens and their Inhibitors. Front. Cell. Infect. Microbiol. 2020, 9, 462. [Google Scholar] [CrossRef]
- Arabyan, N.; Weis, A.M.; Huang, B.C.; Weimer, B.C. Implication of Sialidases in Salmonella infection: Genome Release of Sialidase Knockout Strains from Salmonella enterica serovar Typhimurium LT2. Genome Announc. 2017, 5, e00341-17. [Google Scholar] [CrossRef]
- Nugroho, C.M.H.; Kurnia, R.S.; Tarigan, S.; Silaen, O.S.M.; Triwidyaningtyas, S.; Wibawan, I.W.T.; Natalia, L.; Takdir, A.K.; Soebandrio, A. Screening and Purification of NanB Sialidase from Pasteurella multocida with Activity in Hydrolyzing Sialic Acid Neu5Acα(2-6)Gal and Neu5Acα(2-3)Gal. Sci. Rep. 2022, 12, 9425. [Google Scholar] [CrossRef]
- Lai, J.C.C.; Karunarathna, H.M.T.; Wong, H.H.; Peiris, J.S.M.; Nicholls, J.M. Neuraminidase Activity and Specificity of Influenza A Virus are Influenced Byhemagglutinin-Receptor Binding. Emerg. Microbes Infect. 2019, 8, 327–338. [Google Scholar] [CrossRef]
- Warwas, M.L.; Watson, J.N.; Bennet, A.J.; Moore, M.M. Structure and Role of Sialic Acids on the Surface of Aspergillus fumigatus Conidiospores. Glycobiology 2007, 17, 401–410. [Google Scholar] [CrossRef]
- Telford, J.C.; Yeung, J.H.F.; Xu, G.; Kiefel, M.J.; Watts, A.G.; Hader, S.; Chan, J.; Bennet, A.J.; Moore, M.M.; Taylor, G.L. The Aspergillus fumigatus Sialidase is a 3-deoxy-D-glycero-D-galacto-2-nonulosonic Acid Hydrolase (KDNase): Structural and Mechanistic Insights. J. Biol. Chem. 2011, 286, 10783–19792. [Google Scholar] [CrossRef] [PubMed]
- Nesbitt, J.R.; Steves, E.Y.; Schonhofer, C.R.; Cait, A.; Manku, S.S.; Yeung, J.H.F.; Bennet, A.J.; McNagny, K.M.; Choy, J.C.; Hughes, M.R.; et al. The Aspergillus fumigatus Sialidase (Kdnase) Contributes to Cell Wall Integrity and Virulence in Amphotericin B-Treated Mice. Front. Microbiol. 2018, 8, 2706. [Google Scholar] [CrossRef] [PubMed]
- Royal, G.C., Jr.; Nandedkar, A.K.N.; Sampson, C.C.; Faggett, T. Neuraminidase Production by Candida albicans. J. Nat. Med. Assoc. 1984, 76, 143–145. [Google Scholar]
- Kajihara, Y.; Okamoto, R.; Yamamoto, N.; Izumi, M. Synthesis of glycopeptides. In Methods Enzymol; Fukuda, M., Ed.; Academic Press: San Diego, CA, USA, 2010; Volume 478, pp. 503–519. [Google Scholar] [CrossRef]
- Kim, S.; Oh, D.B.; Kwon, O.; Kang, H.A. Identification and Functional Characterization of the NanH Extracellular Sialidase from Corynebacterium diphtheriae. J. Biochem. 2010, 147, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Xia, G.; Yu, Z.; Zhao, Y.; Wang, Y.; Wang, S.; He, M.; Wang, J.; Xue, C. Sialoglycoproteins Isolated from the Eggs of Carassius auratus Prevents Osteoporosis by Suppressing the Activation of Osteoclastogenesis Related NF-κB and MAPK Pathways. J. Funct. Foods 2015, 17, 491–503. [Google Scholar] [CrossRef]
- Giurgea, L.T.; Morens, D.M.; Taubenberger, J.K.; Memoli, M.J. Influenza Neuraminidase: A Neglected Protein and Its Potential for a Better Influenza. Vaccine 2020, 8, 409. [Google Scholar] [CrossRef] [PubMed]
- Worrall, E.E.; Sudarisman, P.A. Sialivac: An Intranasal Homologous Inactivated Split Virus Vaccine Containing Bacterial Sialidase for the Control of Avian Influenza in Poultry. Vaccine 2009, 27, 4161–4168. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, A.A.; Cyplenkov, N.A.; Dekker, P.J.T.; Efimova, Y.M. Sialidases. U.S. Patent 8,012,733B2, 6 September 2011. Available online: https://patents.google.com/patent/US8012733B2/en (accessed on 20 March 2024).
- Bule, P.; Chuzel, L.; Blagova, E.; Wu, L.; Gray, M.A.; Henrissat, B.; Rapp, E.; Bertozzi, C.R.; Taron, C.H.; Davies, G.J. Inverting Family GH156 Sialidases Define an Unusual Catalytic Motif for Glycosidase Action. Nat. Commun. 2019, 10, 4816. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Schafer, D.; Miller, C.A.; Tanenbaum, S.W.; Flashner, M. Induction and Regulation of Neuraminidase Synthesis in Arthrobacter sialophilus. J. Bacteriol. 1978, 136, 3874–3879. [Google Scholar] [CrossRef] [PubMed]
- Gualdi, L.; Hayre, J.K.; Gerlini, A.; Bidossi, A.; Colomba, L.; Trappetti, C.; Pozzi, G.; Docquier, J.-D.; Andrew, P.; Ricci, S.; et al. Regulation of Neuraminidase Expression in Streptococcus pneumoniae. BMC Microbiol. 2012, 12, 200. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; McClane, B.A. The Sialidases of Clostridium perfringens type D Strain CN3718 Differ in their Properties and Sensitivities to Inhibitors. Appl. Environ. Microbiol. 2014, 80, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Evans, D.R.; Freedman, J.C.; McClane, B.A. NanR regulates NanI Sialidase Expression by Clostridium perfringens F4969, a Human Enteropathogenic Strain. Infect. Immun. 2017, 85, e00241-17. [Google Scholar] [CrossRef]
- Blanchette, K.A.; Shenoy, A.T.; Milner, J., 2nd; Gilley, R.P.; McClure, E.; Hinojosa, C.A.; Kumar, N.; Daugherty, S.C.; Tallon, L.J.; Ott, S.; et al. Neuraminidase A-Exposed Galactose Promotes Streptococcus pneumoniae Biofilm Formation during Colonization. Infect. Immun. 2016, 84, 2922–2932. [Google Scholar] [CrossRef]
- Zhao, S.; Xiang, B.; Yang, L.; Chen, J.; Zhu, C.; Chen, Y.; Cui, J.; Hu, S.; Hu, Y. Genetic Modifications of Critical Regulators Provide New Insights into Regulation Modes of Raw-Starch-Digesting Enzyme Expression in Penicillium. Biotechnol. Biofuels 2022, 15, 62. [Google Scholar] [CrossRef]
- Wang, L.; Wang, A.; Wang, D.; Hong, J. The Novel Properties of Kluyveromyces marxianus Glucose Sensor/Receptor Repressor Pathway and the Construction of Glucose Repression-Released Strains. Microb. Cell Fact. 2023, 22, 123. [Google Scholar] [CrossRef]
- Kunitake, E.; Uchida, R.; Asano, K.; Kanamaru, K.; Kimura, M.; Kimura, T.; Kobayashi, T. cAMP Signaling Factors Regulate Carbon Catabolite Repression of Hemicellulase Genes in Aspergillus nidulans. AMB Expr. 2022, 12, 126. [Google Scholar] [CrossRef]
- Gauna, A.; Larran, A.S.; Feldman, S.R.; Permingeat, H.R.; Perotti, V.E. Secretome characterization of the lignocellulose-degrading fungi Pycnoporus sanguineus and Ganoderma resinaceum growing on Panicum prionitis biomass. Mycologia 2021, 113, 877–890. [Google Scholar] [CrossRef]
- Lima, J.O.; Pereira, J.F.; de Araújo, E.F.; de Queiroz, M.V. Pectin Lyase Overproduction by Penicillium griseoroseum Mutants Resistant to Catabolite Repression. Braz. J. Microbiol. 2017, 48, 602–606. [Google Scholar] [CrossRef] [PubMed]
- Abrashev, R.; Krumova, E.; Petrova, P.; Eneva, R.; Kostadinova, N.; Miteva-Staleva, J.; Engibarov, S.; Stoyancheva, G.; Gocheva, Y.; Kolyovska, V.; et al. Distribution of a Novel Enzyme of Sialidase Family among Native Filamentous Fungi. Fungal Biol. 2021, 125, 412–425. [Google Scholar] [CrossRef] [PubMed]
- Dolashki, A.; Abrashev, R.; Kaynarov, D.; Krumova, E.; Velkova, L.; Eneva, R.; Engibarov, S.; Gocheva, Y.; Miteva-Staleva, J.; Dishliyska, V.; et al. Structural and Functional Characterization of Cold-Active Sialidase Isolated from Antarctic Fungus Penicillium griseofulvum P29. Biochem. Biophys. Rep. 2024, 37, 101610. [Google Scholar] [CrossRef]
- Kim, D.; Choi, K.Y.; Yoo, M.; Zylstra, G.J.; Kim, E. Biotechnological Potential of Rhodococcus Biodegradative Pathways. J. Microbiol. Biotechnol. 2018, 28, 1037–1051. [Google Scholar] [CrossRef] [PubMed]
- Uchida, Y.; Tsukada, Y.; Sugimori, T. Distribution of Neuraminidase in Arthrobacter and its Purification by Affinity Chromatography. J. Biochem. 1977, 82, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Abrashev, I.; Velcheva, P.; Nikolov, P.; Kourteva, J. Substrate for Colorimetric Determination of Enzyme Activity. Bulgaria Patent N 47647/IIR, 1980. Available online: https://cir.nii.ac.jp/crid/1572543025073467520 (accessed on 21 March 2024).
- Lichtensteiger, C.A.; Vimr, E.R. Neuraminidase (sialidase) activity of Haemophilus parasuis. FEMS Microbiol. Lett. 1997, 152, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Coulouris, G.; Zaretskaya, I.; Cutcutache, I.; Rozen, S.; Madden, T. Primer-BLAST: A Tool to Design Target-Specific Primers for Polymerase Chain Reaction. BMC Bioinformat. 2012, 13, 134. [Google Scholar] [CrossRef] [PubMed]
- Løvdal, T.; Lillo, C. Reference Gene Selection for Quantitative Real-Time PCR Normalization in Tomato Subjected to Nitrogen, Cold, and Light Stress. Anal. Biochem. 2009, 387, 238–242. [Google Scholar] [CrossRef]
- Somogy, M. Notes on sugar determination. J. Biol. Chem. 1952, 195, 19–23. [Google Scholar] [CrossRef]
- Engibarov, S.; Eneva, R.; Abrashev, I. Neuraminidase (sialidase) from Aeromonas sp. Strain 40/02—Isolation and Partial Purification. Ann. Microbiol. 2015, 65, 1515–1523. [Google Scholar] [CrossRef]
- Martin, K.; McDougall, B.M.; McIlroy, S.; Chen, J.; Seviour, R.J. Biochemistry and Molecular Biology of Exocellular Fungal beta-(1,3)- and beta-(1,6)-Glucanases. FEMS Microbiol. Rev. 2007, 31, 168–192. [Google Scholar] [CrossRef]
- Kislitsin, V.Y.; Chulkin, A.M.; Dotsenko, A.S.; Sinelnikov, I.G.; Sinitsyn, A.P.; Rozhkova, A.M. The Role of Intracellular b-Glucosidase in Cellulolytic Response Induction in Filamentous Fungus Penicillium verruculosum. Res. Microbiol. 2023, 30, 104178. [Google Scholar] [CrossRef]
- Feng, B.; Friedlin, E.; Marzluf, G.A. A Reporter Gene Analysis of Penicillin Biosynthesis Gene Expression in Penicillium chrysogenum and its Regulation by Nitrogen and Glucose Catabolite Repression. Appl. Environ. Microbiol. 1994, 60, 4432–4439. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Han, C.; Shimizu, H.; Wong, B. The Effect of Fructose, Galactose, and Glucose on the Induction of β-Galactosidase in Escherichia coli. J. Exp. Microbiol. Immunol. 2002, 2, 1–5. [Google Scholar]
- Dynesen, J.; Smits, H.P.; Olsson, L.; Nielsen, J. Carbon Catabolite Repression of Invertase during Batch Cultivations of Saccharomyces cerevisiae: The Role of Glucose, Fructose, and Mannose. Appl. Microbiol. Biotechnol. 1998, 50, 579–582. [Google Scholar] [CrossRef]
- Jankovic, I.; Brückner, R. Carbon Catabolite Repression of Sucrose Utilization in Staphylococcus xylosus: Catabolite Control Protein CcpA Ensures Glucose Preference and Autoregulatory Limitation of Sucrose Utilization. J. Mol. Microbiol. Biotechnol. 2007, 12, 114–120. [Google Scholar] [CrossRef]
- Chan, V.; Dreolini, L.F.; Flintoff, K.A.; Lloyd, S.J.; Mattenley, A.A. The Effects of Glycerol, Glucose, Galactose, Lactose and Glucose with Galactose on the Induction of β-Galactosidase in Escherichia coli. J. Exp. Microbiol. Immunol. 2002, 2, 130–137. [Google Scholar]
- Gancedo, J.M. Yeast Carbon Catabolite Repression. Microbiol. Mol. Biol. Rev. 1998, 62, 334–361. [Google Scholar] [CrossRef]
- Runquistd, D.; Parachin, N.S.; Hahn-Hägerdal, B. Challenges in Co-Fermentation of Lignocellulose Derived Sugars using Baker’s Yeast. In Woodhead Publishing Series in Energy, Bioalcohol Production; Waldron, K., Ed.; Woodhead Publishing: Cambridge, UK, 2010; pp. 224–245. [Google Scholar] [CrossRef]
- Lagree, K.; Woolford, C.A.; Huang, M.Y.; May, G.; McManus, C.J.; Solis, N.V.; Filler, S.G.; Mitchell, A.P. Roles of Candida albicans Mig1 and Mig2 in Glucose Repression, Pathogenicity Traits, and SNF1 Essentiality. PLoS Genet. 2020, 16, e1008582. [Google Scholar] [CrossRef]
- Adnan, M.; Zheng, W.; Islam, W.; Arif, M.; Abubakar, Y.S.; Wang, Z.; Lu, G. Carbon Catabolite Repression in Filamentous Fungi. Int. J. Mol. Sci. 2018, 19, 48. [Google Scholar] [CrossRef]
- Chen, Y.; Dong, L.; Alam, M.A.; Pardeshi, L.; Miao, Z.; Wang, F.; Tan, K.; Hynes, M.J.; Kelly, J.M.; Wong, K.H. Carbon Catabolite Repression Governs Diverse Physiological Processes and Development in Aspergillus nidulans. mBio 2022, 13, e03734-21. [Google Scholar] [CrossRef]
- Chulkin, A.M.; Vavilova, E.A.; Benevolenskij, S.V. Transcriptional regulator of carbon catabolite repression CreA of filamentous fungus Penicillium canescens. Mol. Biol. 2010, 44, 677–687. [Google Scholar] [CrossRef]
- Peng, M.; Khosravi, C.; Lubbers, R.J.M.; Kun, R.S.; Pontes, M.V.A.; Battaglia, E.; Chen, C.; Dalhuijsen, S.; Daly, P.; Lipzen, A.; et al. CreA-mediated repression of gene expression occurs at low monosaccharide levels during fungal plant biomass conversion in a time and substrate dependent manner. Cell Surf. 2021, 7, 100050. [Google Scholar] [CrossRef]
- Banani, H.; Marcet-Houben, M.; Ballester, A.R.; Abbruscato, P.; González-Candelas, L.; Gabaldón, T.; Spadaro, D. Genome sequencing and secondary metabolism of the postharvest pathogen Penicillium griseofulvum. BMC Genom. 2016, 17, 19. [Google Scholar] [CrossRef]
- Knudsen, S. Promoter 2.0: For the recognition of PolII promoter sequences. Bioinformatics 1999, 15, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Wang, Y.; Huang, S.; Zhu, P.; Huang, W.E.; Ling, J.; Xu, J. Metabolic-Activity-Based Assessment of Antimicrobial Effects by D2O-Labeled Single-Cell Raman Microspectroscopy. Anal. Chem. 2017, 89, 4108–4115. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ren, L.; Chen, R.; Sun, X.; Xu, J.; Zhu, P.; Yang, F. Assessing Efficacy of Clinical Disinfectants for Pathogenic Fungi by Single-Cell Raman Microspectroscopy. Front. Cell. Infect. Microbiol. 2022, 12, 772378. [Google Scholar] [CrossRef]
- Ilyés, H.; Fekete, E.; Karaffa, L.; Fekete, É.; Sándor, E.; Szentirmai, A.; Kubicek, C.P. CreA-Mediated Carbon Catabolite Repression of b-Galactosidase Formation in Aspergillus nidulans is Growth Rate Dependent. FEMS Microbiol. Lett. 2004, 235, 147–151. [Google Scholar] [CrossRef]
- Flipphi, M.; van de Vondervoort, P.J.I.; Ruijter, G.J.G.; Visser, J.; Arst, H.N., Jr.; Felenbok, B. Onset of Carbon Catabolite Repression in Aspergillus nidulans Parallel Involvement of Hexokinase and Glucokinase in Sugar Signaling. J. Biol. Chem. 2003, 278, 11849–11857. [Google Scholar] [CrossRef]
- de Assis, L.J.; Silva, L.P.; Bayram, O.; Dowling, P.; Kniemeye, O.; Krüger, T.; Brakhage, A.A.; Chen, Y.; Dong, L.; Tan, K.; et al. Carbon Catabolite Repression in Filamentous Fungi Is Regulated by Phosphorylation of the Transcription Factor CreA. Mol. Biol. Physiol. 2021, 12, e03146-20. [Google Scholar] [CrossRef] [PubMed]
- Hoyer, L.L.; Hamilton, A.C.; Steenbergen, S.M.; Vimr, E.R. Cloning, Sequencing and Distribution of the Salmonella typhimurium LT2 Sialidase Gene, NanH, Provides Evidence for Interspecies Gene Transfer. Mol. Microbiol. 1992, 6, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Sarma, S.J. The Impact of Carbon and Nitrogen Catabolite Repression in Microorganisms. Microbiol. Res. 2021, 251, 126831. [Google Scholar] [CrossRef] [PubMed]
- Franzino, T.; Boubakri, H.; Cernava, T.; Abrouk, D.; Achouak, W.; Reverchon, S.; Nasser, W.; Haichar, Z.F. Implications of Carbon Catabolite Repression for Plant–Microbe Interactions. Plant Commun. 2022, 3, 100272. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Fujita, N.; Yamamoto, K.; Ishihama, A. Novel Roles of cAMP Receptor Protein (CRP) in Regulation of Transport and Metabolism of Carbon Sources. PLoS ONE 2011, 6, e20081. [Google Scholar] [CrossRef] [PubMed]
- Collier, L.A.; Ghosh, A.; Borkovich, K.A. Heterotrimeric G-Protein Signaling is required for Cellulose Degradation in Neurospora crassa. mBio 2020, 11, e02419-20. [Google Scholar] [CrossRef] [PubMed]
- St. Leger, R.J.; Durrands, P.K.; Cooper, R.M.; Charnley, A.K. Regulation of Production of Proteolytic Enzymes by the Entomopathogenic Fungus Metarhizium anisopliae. Arch. Microbiol. 1988, 150, 413–416. [Google Scholar] [CrossRef]
- Yu, N.-N.; Ketya, W.; Park, G. Intracellular Nitric Oxide and cAMP are Involved in Cellulolytic Enzyme Production in Neurospora crassa. Int. J. Mol. Sci. 2023, 24, 4503. [Google Scholar] [CrossRef]
- Joshi, S.; Satyanarayana, T. Biotechnology of Cold-Active Proteases. Biology 2013, 2, 755–783. [Google Scholar] [CrossRef]
- Carrasco, M.; Rozas, J.M.; Cifuentes, J.A.V.; Baeza, M. Pectinase Secreted by Psychrotolerant Fungi: Identifcation, Molecular Characterization and Heterologous Expression of a Cold-Active Polygalacturonase from Tetracladium sp. Microb. Cell Fact. 2019, 18, 45. [Google Scholar] [CrossRef] [PubMed]
- Han, S.J.; Park, H.; Lee, S.G.; Lee, H.K.; Yim, J.H. Optimization of Cold-Active Chitinase Production from the Antarctic Bacterium, Sanguibacter antarcticus KOPRI 21702. Appl. Microbiol. Biotechnol. 2011, 89, 613–621. [Google Scholar] [CrossRef] [PubMed]


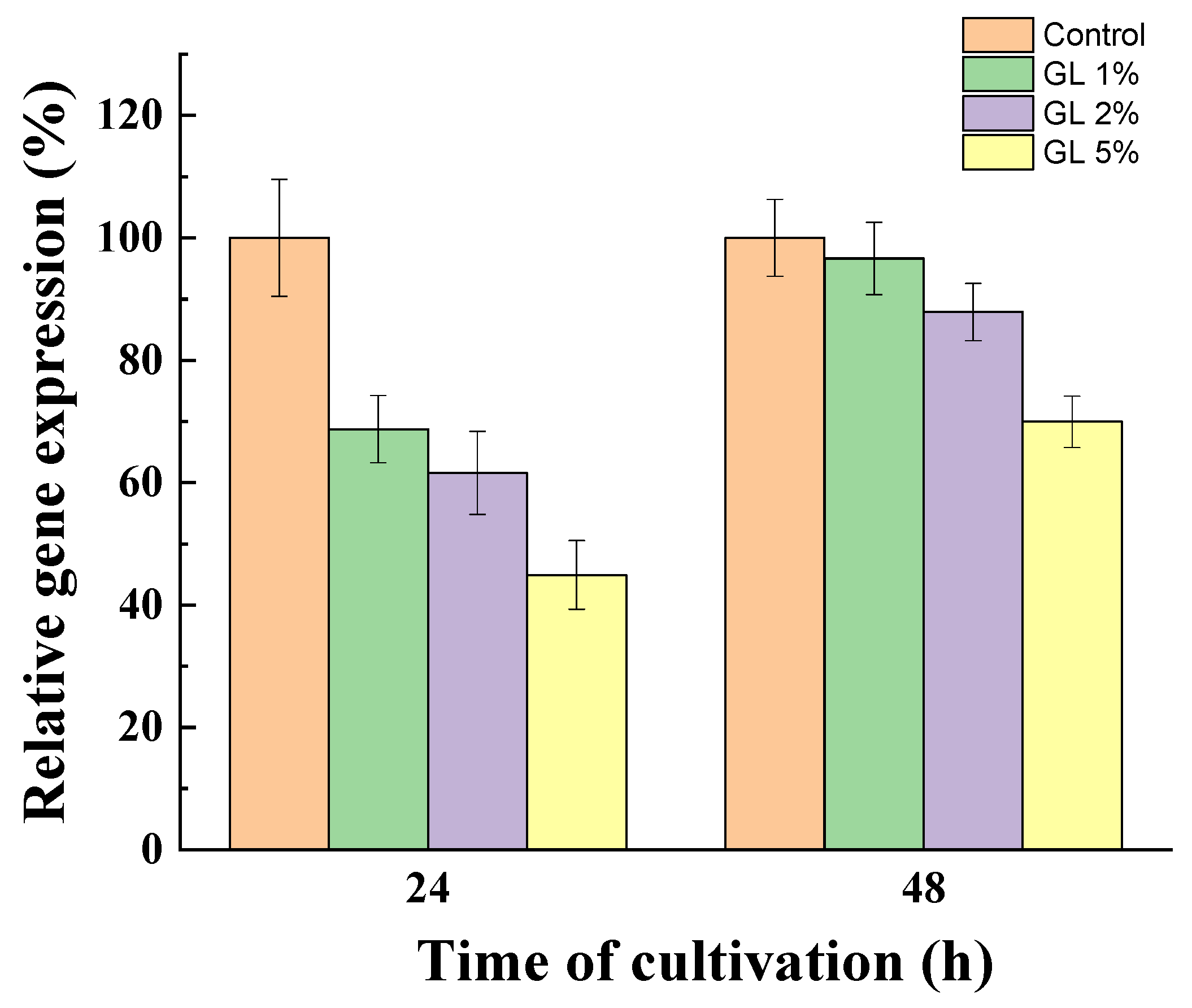

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | PCR Product (bp) | Reference |
|---|---|---|---|
| PenRTF | CAGAACTCTTCCGTTCGGCT | 100 | This study |
| PenRTR | TCACATAGGCTGCAAGGACG | ||
| GADPH_F | CTGCTCTCTCATAGCCAACAC | 157 | [33] |
| GADPH_R | CTTCCTCCAATAGCAGAGGTTT |
| Carbon Source | Biomass [g/100 mL] | Sialidase Activity [U/mL] | ||||
|---|---|---|---|---|---|---|
| 24 h | 48 h | 72 h | 24 h | 48 h | 72 h | |
| Whey | 0.2902 | 0.6438 | 0.7123 | 10.5 | 8.4 | 8.2 |
| Wh + Glucose | 0.8541 | 1.3481 | 1.7254 | 3.1 | 5.9 | 7.4 |
| Wh + Maltose | 0.6292 | 0.9245 | 0.9932 | 3.9 | 4.2 | 5.1 |
| Wh + Sucrose | 0.7998 | 1.1895 | 1.5932 | 2.5 | 6.7 | 4.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abrashev, R.; Krumova, E.; Petrova, P.; Eneva, R.; Dishliyska, V.; Gocheva, Y.; Engibarov, S.; Miteva-Staleva, J.; Spasova, B.; Kolyovska, V.; et al. Glucose Catabolite Repression Participates in the Regulation of Sialidase Biosynthesis by Antarctic Strain Penicillium griseofulvum P29. J. Fungi 2024, 10, 241. https://doi.org/10.3390/jof10040241
Abrashev R, Krumova E, Petrova P, Eneva R, Dishliyska V, Gocheva Y, Engibarov S, Miteva-Staleva J, Spasova B, Kolyovska V, et al. Glucose Catabolite Repression Participates in the Regulation of Sialidase Biosynthesis by Antarctic Strain Penicillium griseofulvum P29. Journal of Fungi. 2024; 10(4):241. https://doi.org/10.3390/jof10040241
Chicago/Turabian StyleAbrashev, Radoslav, Ekaterina Krumova, Penka Petrova, Rumyana Eneva, Vladislava Dishliyska, Yana Gocheva, Stefan Engibarov, Jeny Miteva-Staleva, Boryana Spasova, Vera Kolyovska, and et al. 2024. "Glucose Catabolite Repression Participates in the Regulation of Sialidase Biosynthesis by Antarctic Strain Penicillium griseofulvum P29" Journal of Fungi 10, no. 4: 241. https://doi.org/10.3390/jof10040241
APA StyleAbrashev, R., Krumova, E., Petrova, P., Eneva, R., Dishliyska, V., Gocheva, Y., Engibarov, S., Miteva-Staleva, J., Spasova, B., Kolyovska, V., & Angelova, M. (2024). Glucose Catabolite Repression Participates in the Regulation of Sialidase Biosynthesis by Antarctic Strain Penicillium griseofulvum P29. Journal of Fungi, 10(4), 241. https://doi.org/10.3390/jof10040241