CD147 and Cyclooxygenase Expression in Feline Oral Squamous Cell Carcinoma

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Immunohistochemical Detection of COX Enzymes and CD147 in Feline OSCC Biopsies
2.1.1. Patient Signalment and Biopsy Information
2.1.2. Primary Antibodies and Matching Isotype Control Antibodies
2.1.3. Immunohistochemistry Protocol
2.1.4. Immunohistochemistry Grading System
2.2. Quantitative Reverse Transcriptase Polymerase Chain Reaction (RT-qPCR) for Determination of Relative Gene Expression of COX Enzymes and CD147 in Feline OSCC Cell Lines
2.2.1. Cell Lines and Culture Reagents
2.2.2. Experimental Conditions, RNA Extraction, and cDNA Synthesis
2.2.3. Primer Design and RT-qPCR Optimization
2.3. Enzyme-Linked Immunosorbent Assay (ELISA) Detection of PGE2 in Feline OSCC Cell Lines
2.4. Statistical Analysis
3. Results
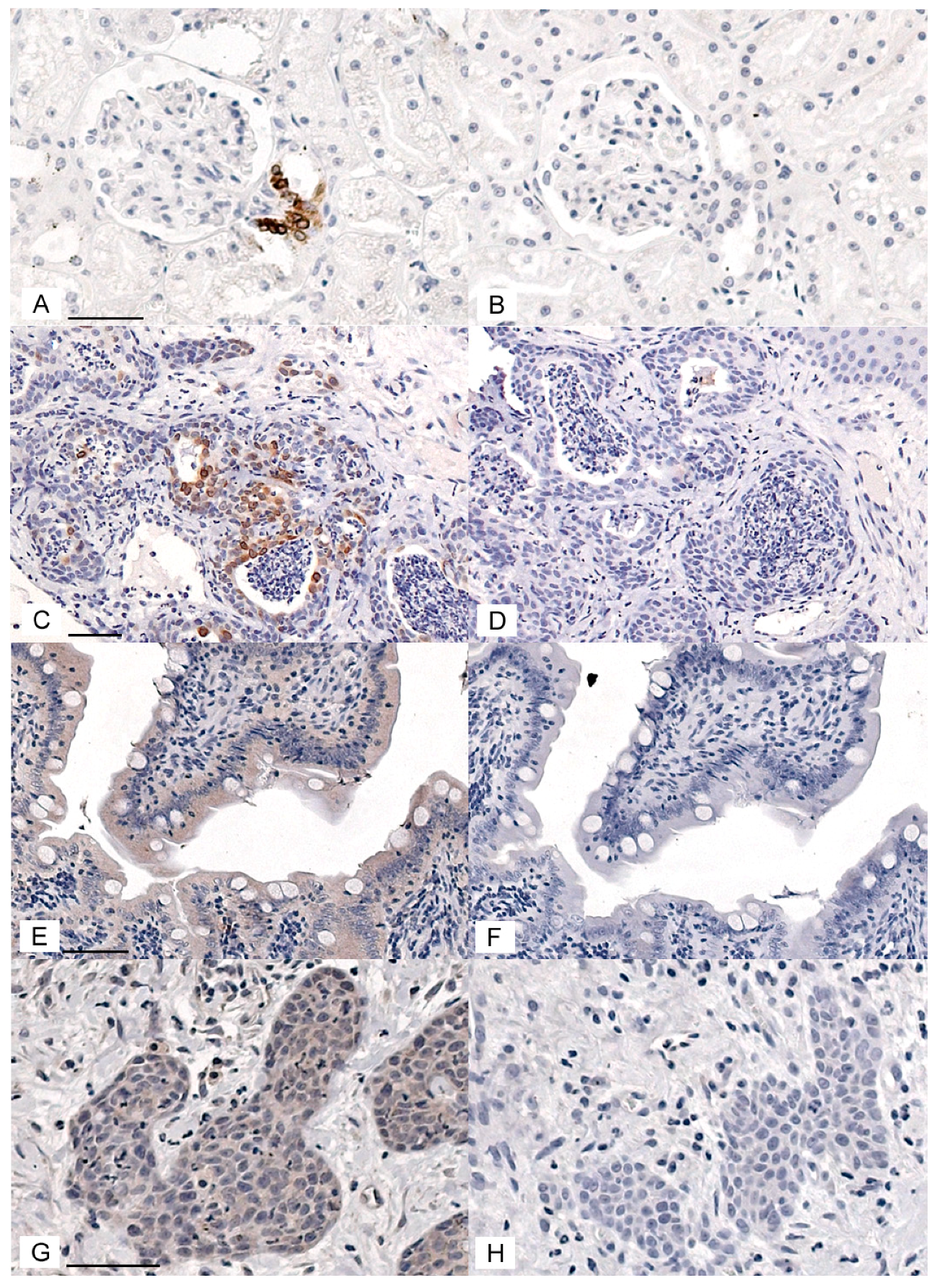
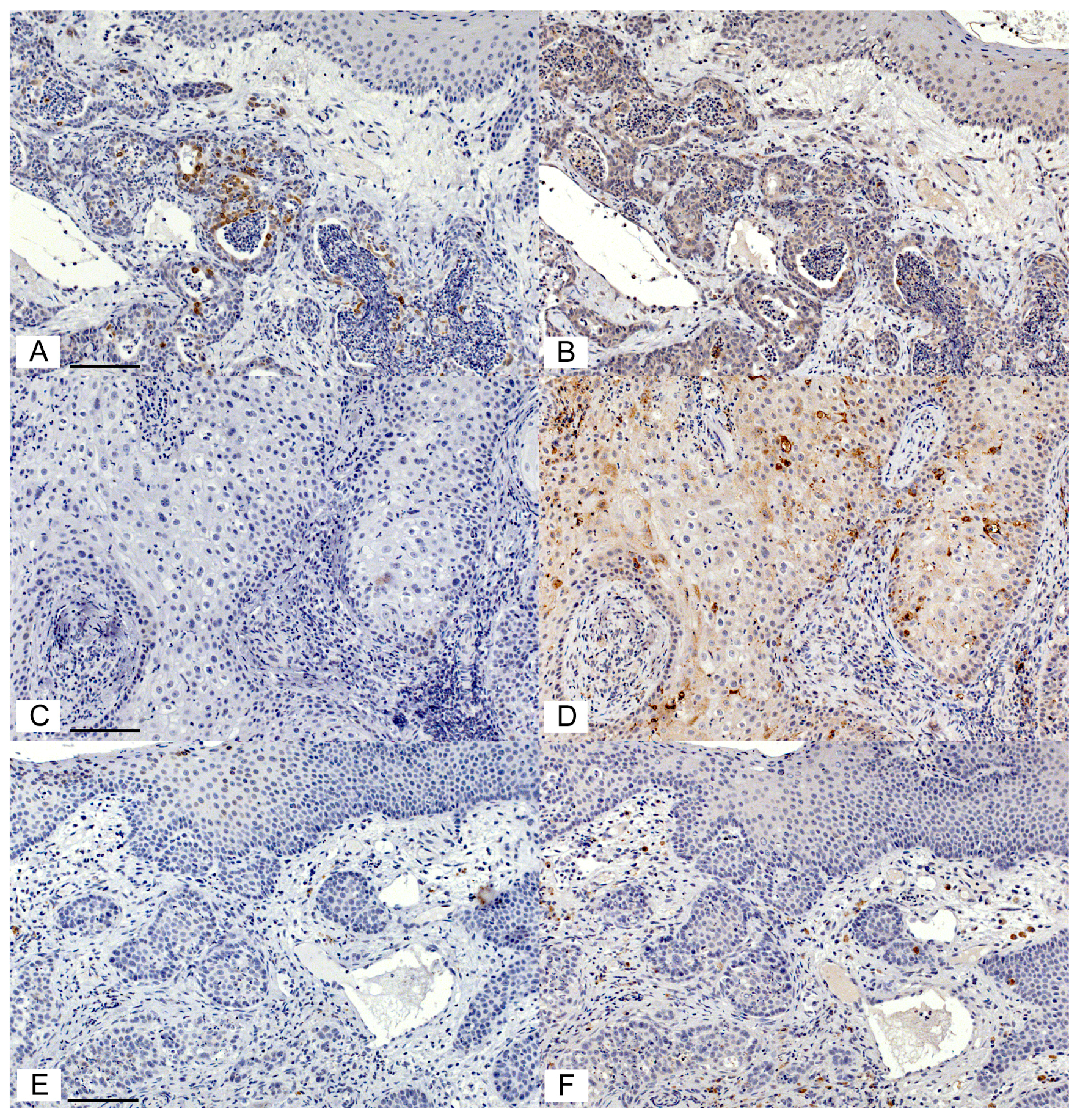
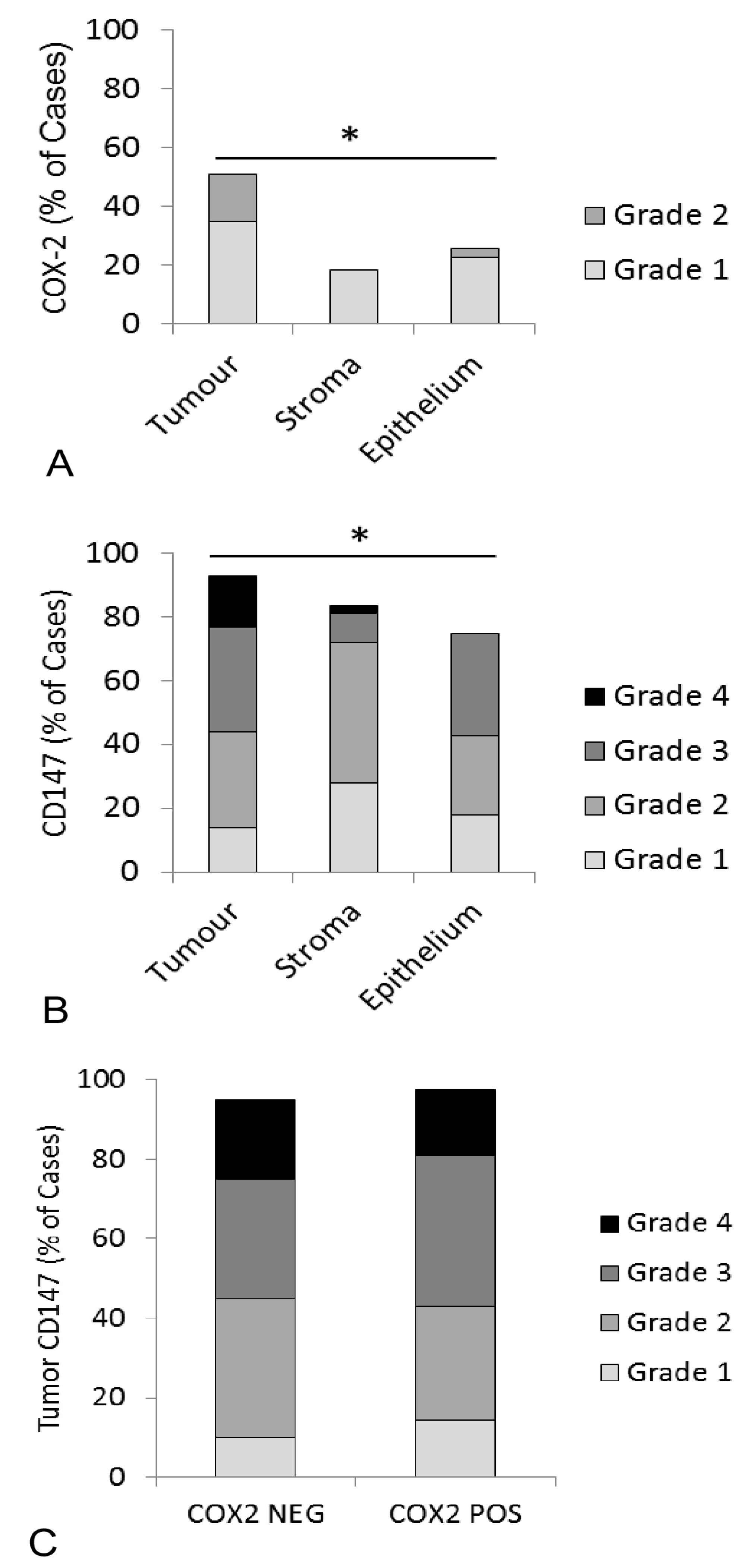
3.1. Immunohistochemical Detection of COX Enzymes and CD147 in Feline OSCC Biopsies
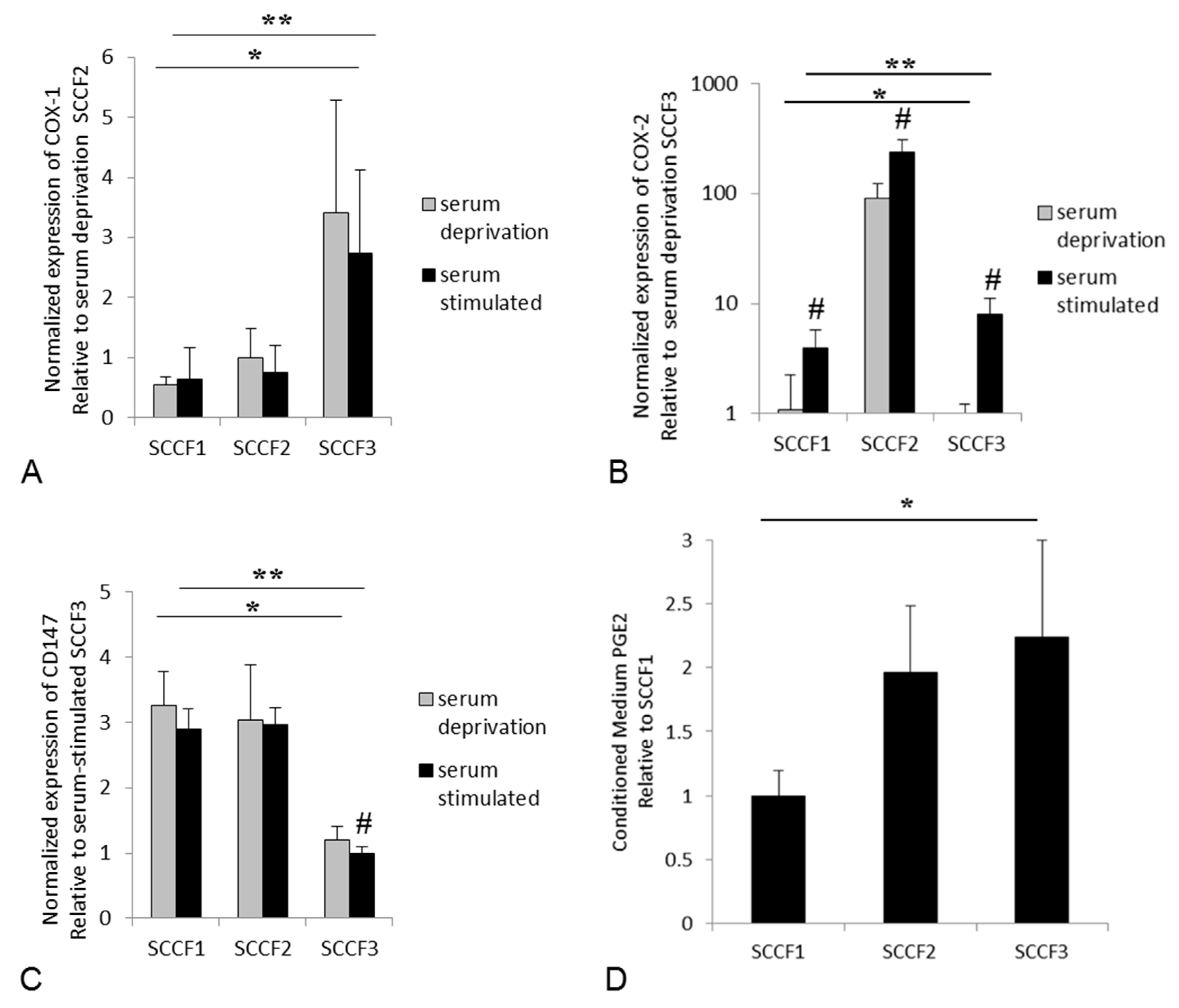
3.2. Expression of COX-1, COX-2, and CD147, and Secretion of PGE2 in Feline OSCC Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stebbins, K.E.; Morse, C.C.; Goldschmidt, M.H. Feline oral neoplasia: A ten-year survey. Vet. Pathol. 1989, 26, 121–128. [Google Scholar] [CrossRef] [PubMed]
- DiBernardi, L.; Dore, M.; Davis, J.A.; Owens, J.G.; Mohammed, S.I.; Guptill, C.F.; Knapp, D.W. Study of feline oral squamous cell carcinoma: Potential target for cyclooxygenase inhibitor treatment. Prostaglandins Leukot. Essent. Fatty Acids 2007, 76, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Marretta, J.J.; Garrett, L.D.; Marretta, S.M. Feline oral squamous cell carcinoma: An overview. Vet. Med. 2007, 102, 392–406. [Google Scholar]
- Martin, C.K.; Tannehill-Gregg, S.H.; Wolfe, T.D.; Rosol, T.J. Bone-invasive oral squamous cell carcinoma in cats: Pathology and expression of parathyroid hormone-related protein. Vet. Pathol. 2011, 48, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Bilgic, O.; Duda, L.; Sanchez, M.D.; Lewis, J.R. Feline Oral Squamous Cell Carcinoma: Clinical Manifestations and Literature Review. J. Vet. Dent. 2015, 32, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Hutson, C.A.; Willauer, C.C.; Walder, E.J.; Stone, J.L.; Klein, M.K. Treatment of mandibular squamous cell carcinoma in cats by use of mandibulectomy and radiotherapy: Seven cases (1987–1989). J. Am. Vet. Med. Assoc. 1992, 201, 777–781. [Google Scholar] [PubMed]
- Supsavhad, W.; Dirksen, W.P.; Martin, C.K.; Rosol, T.J. Animal models of head and neck squamous cell carcinoma. Vet. J. 2016, 210, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bertone, E.R.; Snyder, L.A.; Moore, A.S. Environmental and lifestyle risk factors for oral squamous cell carcinoma in domestic cats. J. Vet. Intern. Med. 2003, 17, 557–562. [Google Scholar] [CrossRef] [PubMed]
- De Camargo Cancela, M.; Voti, L.; Guerra-Yi, M.; Chapuis, F.; Mazuir, M.; Curado, M.P. Oral cavity cancer in developed and in developing countries: Population-based incidence. Head. Neck. 2010, 32, 357–367. [Google Scholar] [CrossRef] [PubMed]
- Strati, K.; Lambert, P.F. Human Papillomavirus Association with Head and Neck Cancers: Understanding Virus Biology and using it in the Development of Cancer Diagnostics. Expert. Opin. Med. Diagn. 2008, 2, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Munday, J.S.; French, A.F. Felis catus papillomavirus types 1 and 4 are rarely present in neoplastic and inflammatory oral lesions of cats. Res. Vet. Sci. 2015, 100, 220–222. [Google Scholar] [CrossRef] [PubMed]
- Supsavhad, W.; Dirksen, W.P.; Hildreth, B.E.; Rosol, T.J. p16, pRb, and p53 in Feline Oral Squamous Cell Carcinoma. Vet. Sci. 2016, 3. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, Y.; Hatori, M.; Ando, Y.; Ito, D.; Toyoshima, T.; Tanaka, M.; Shintani, S. Inhibition of cyclooxygenase-2 suppresses the invasiveness of oral squamous cell carcinoma cell lines via down-regulation of matrix metalloproteinase-2 production and activation. Clin. Exp. Metastasis 2009, 26, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Vendramini-Costa, D.B.; Carvalho, J.E. Molecular link mechanisms between inflammation and cancer. Curr. Pharm. Des. 2012, 18, 3831–3852. [Google Scholar] [CrossRef] [PubMed]
- Chan, G.; Boyle, J.O.; Yang, E.K.; Zhang, F.; Sacks, P.G.; Shah, J.P.; Edelstein, D.; Soslow, R.A.; Koki, A.T.; Woerner, B.M.; et al. Cyclooxygenase-2 expression is up-regulated in squamous cell carcinoma of the head and neck. Cancer. Res. 1999, 59, 991–994. [Google Scholar] [PubMed]
- Hayes, A.; Scase, T.; Miller, J.; Murphy, S.; Sparkes, A.; Adams, V. COX-1 and COX-2 Expression in Feline Oral Squamous Cell Carcinoma. J. Comp. Pathol. 2006, 135, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Rathore, K.; Alexander, M.; Cekanova, M. Piroxicam inhibits Masitinib-induced cyclooxygenase 2 expression in oral squamous cell carcinoma cells in vitro. Transl. Res. 2014, 164, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.E.; Bear, M.D.; Rosol, T.J.; Premanandan, C.; Kisseberth, W.C.; London, C.A. Characterization of STAT3 expression, signaling and inhibition in feline oral squamous cell carcinoma. BMC Vet. Res. 2015, 11, 206–219. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.K.; Dirksen, W.P.; Carlton, M.M.; Lanigan, L.G.; Pillai, S.P.; Werbeck, J.L.; Simmons, J.K.; Hildreth, B.E., 3rd; London, C.A.; Toribio, R.E.; et al. Combined zoledronic acid and meloxicam reduced bone loss and tumour growth in an orthotopic mouse model of bone-invasive oral squamous cell carcinoma. Vet. Comp. Oncol. 2015, 13, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; McLean, M.K. Toxicology of frequently encountered nonsteroidal anti-inflammatory drugs in dogs and cats. Vet. Clin. North Am. Small Anim. Pract. 2012, 42, 289–306. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, L.S.; Delgado, M.L.; Ricardo, S.; Garcez, F.; do Amaral, B.; Pacheco, J.J.; Lopes, C.; Bousbaa, H. EMMPRIN expression in oral squamous cell carcinomas: Correlation with tumor proliferation and patient survival. Biomed. Res. Int. 2014, 2014, 905680. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Huang, H.; Li, H.; Chen, W.; Pan, C. EMMPRIN expression in tongue squamous cell carcinoma. J. Oral. Pathol. Med. 2009, 38, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Tumor angiogenesis: MMP-mediated induction of intravasation- and metastasis-sustaining neovasculature. Matrix. Biol. 2015, 44–46, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Deryugina, E.I.; Quigley, J.P. Matrix metalloproteinases and tumor metastasis. Cancer Metastasis Rev. 2006, 25, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Papadimitropoulou, A.; Mamalaki, A. The glycosylated IgII extracellular domain of EMMPRIN is implicated in the induction of MMP-2. Mol. Cell. Biochem. 2013, 379, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, Y.; Huang, Y.; Huang, D.; Li, Y.; Wu, J.; Duan, M. Expression of COX-2, CD44v6 and CD147 and relationship with invasion and lymph node metastasis in hypopharyngeal squamous cell carcinoma. PLoS ONE 2013, 8, e71048. [Google Scholar] [CrossRef] [PubMed]
- Sweeny, L.; Hartman, Y.E.; Zinn, K.R.; Prudent, J.R.; Marshall, D.J.; Shekhani, M.S.; Rosenthal, E.L. A novel extracellular drug conjugate significantly inhibits head and neck squamous cell carcinoma. Oral. Oncol. 2013, 49, 991–997. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Ishikawa, K. Combined inhibition of EMMPRIN and epidermal growth factor receptor prevents the growth and migration of head and neck squamous cell carcinoma cells. Int. J. Oncol. 2014, 44, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Amagase, K.; Ebara, S.; Akiba, Y.; Takeuchi, K. Cyclooxygenase (COX)-1 and COX-2 both play an important role in the protection of the duodenal mucosa in cats. J. Pharmacol. Exp. Ther. 2013, 344, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Pelligand, L.; Suemanotham, N.; King, J.N.; Seewald, W.; Syme, H.; Smith, K.; Lees, P.; Elliott, J. Effect of Cyclooxygenase(COX)-1 and COX-2 inhibition on furosemide-induced renal responses and isoform immunolocalization in the healthy cat kidney. BMC. Vet. Res. 2015, 11, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Mykkanen, A.K.; Niku, M.; Ilves, M.; Koho, N.M. Expression of monocarboxylate transporters I and IV and the ancillary protein CD147 in the intestinal tract of healthy horses and ponies. Am. J. Vet. Res. 2015, 76, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Ritzhaupt, A.; Wood, I.S.; Ellis, A.; Hosie, K.B.; Shirazi-Beechey, S.P. Identification and characterization of a monocarboxylate transporter (MCT1) in pig and human colon: Its potential to transport L-lactate as well as butyrate. J. Physiol. 1998, 513 (Pt 3), 719–732. [Google Scholar] [CrossRef]
- Halestrap, A.P.; Meredith, D. The SLC16 gene family-from monocarboxylate transporters (MCTs) to aromatic amino acid transporters and beyond. Pflugers. Arch. 2004, 447, 619–628. [Google Scholar] [CrossRef] [PubMed]
- Tannehill-Gregg, S.; Kergosien, E.; Rosol, T.J. Feline head and neck squamous cell carcinoma cell line: Characterization, production of parathyroid hormone-related protein, and regulation by transforming growth factor-beta. In Vitro Cell. Dev. Biol. Anim. 2001, 37, 676–683. [Google Scholar] [CrossRef]
- Martin, C.K.; Dirksen, W.P.; Shu, S.T.; Werbeck, J.L.; Thudi, N.K.; Yamaguchi, M.; Wolfe, T.D.; Heller, K.N.; Rosol, T.J. Characterization of bone resorption in novel in vitro and in vivo models of oral squamous cell carcinoma. Oral. Oncol. 2012, 48, 491–499. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, N.; Beckers, S.; Waigel, S.; Mensah, J.; Wu, J.; Mo, J.; Fleisher, K.E.; Bouquot, J.; Sacks, P.G.; Zacharias, W. Increased EMMPRIN (CD 147) expression during oral carcinogenesis. Exp. Mol. Pathol. 2006, 80, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wang, J.; Fan, L.; Guo, Y. Inhibition of CD147 expression promotes chemosensitivity in HNSCC cells by deactivating MAPK/ERK signaling pathway. Exp. Mol. Pathol. 2017, 102, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Asawakarn, S.; Asawakarn, T. Role of matrix metalloproteinases in animals. Thai J. Vet. Med. 2012, 42, 137–142. [Google Scholar]
- Mandara, M.T.; Pavone, S.; Mandrioli, L.; Bettini, G.; Falzone, C.; Baroni, M. Matrix metalloproteinase-2 and matrix metalloproteinase-9 expression in canine and feline meningioma. Vet. Pathol. 2009, 46, 836–845. [Google Scholar] [CrossRef] [PubMed]
- Pisamai, S.; Rungsipipat, A.; Kunnasut, N.; Suriyaphol, G. Immunohistochemical Expression Profiles of Cell Adhesion Molecules, Matrix Metalloproteinases and their Tissue Inhibitors in Central and Peripheral Neoplastic Foci of Feline Mammary Carcinoma. J. Comp. Pathol. 2017, 157, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Tamamoto, T.; Ohno, K.; Takahashi, M.; Fukushima, K.; Kanemoto, H.; Fujino, Y.; Tsujimoto, H. Plasma matrix metalloproteinase-9 activity in cats with lymphoma. Vet. Comp. Oncol. 2017, 15, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Beam, S.L.; Rassnick, K.M.; Moore, A.S.; McDonough, S.P. An immunohistochemical study of cyclooxygenase-2 expression in various feline neoplasms. Vet. Pathol. 2003, 40, 496–500. [Google Scholar] [CrossRef] [PubMed]
- Sayasith, K.; Sirois, J.; Dore, M. Molecular characterization of feline COX-2 and expression in feline mammary carcinomas. Vet. Pathol. 2009, 46, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Cho, H.; Lee, T.B.; Choi, C.H.; Min, Y.D.; Kim, S.S.; Kim, K.J. Impacts of cytosolic phospholipase A2, 15-prostaglandin dehydrogenase, and cyclooxygenase-2 expressions on tumor progression in colorectal cancer. Yonsei Med. J. 2010, 51, 692–699. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, S.I.; Khan, K.N.; Sellers, R.S.; Hayek, M.G.; DeNicola, D.B.; Wu, L.; Bonney, P.L.; Knapp, D.W. Expression of cyclooxygenase-1 and 2 in naturally-occurring canine cancer. Prostaglandins Leukot. Essent. Fatty Acids 2004, 70, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Millanta, F.; Asproni, P.; Canale, A.; Citi, S.; Poli, A. COX-2, mPGES-1 and EP2 receptor immunohistochemical expression in canine and feline malignant mammary tumours. Vet. Comp. Oncol. 2014, 14, 270–280. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.M.; Saeki, K.; Tanaka, Y.; Nishimura, R.; Nakagawa, T. Effects of etoposide alone and in combination with piroxicam on canine osteosarcoma cell lines. Vet. J. 2016, 218, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Boutemmine, D.; Bouchard, N.; Boerboom, D.; Jones, H.E.; Goff, A.K.; Dore, M.; Sirois, J. Molecular characterization of canine prostaglandin G/H synthase-2 and regulation in prostatic adenocarcinoma cells in vitro. Endocrinology 2002, 143, 1134–1143. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Hase, S.; Miyazawa, T.; Takeuchi, K. Up-regulation of cyclooxygenase-2 by inhibition of cyclooxygenase-1: A key to nonsteroidal anti-inflammatory drug-induced intestinal damage. J. Pharmacol Exp. Ther. 2002, 300, 754–761. [Google Scholar] [CrossRef] [PubMed]
- Newman, S.J.; Mrkonjich, L. Cyclooxygenase-2 expression in feline pancreatic adenocarcinomas. J. Vet. Diagn. Investig. 2006, 18, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, T.; Sakamoto, H.; Horiuchi, T.; Yamamoto, S.; Suematsu, A.; Oda, H.; Koshihara, Y. Involvement of prostaglandin E(2) in interleukin-1α-induced parathyroid hormone-related peptide production in synovial fibroblasts of patients with rheumatoid arthritis. J. Clin. Endocrinol. Metab. 2001, 86, 3272–3278. [Google Scholar] [CrossRef] [PubMed]
- Remillard, T.C.; Bratslavsky, G.; Jensen-Taubman, S.; Stetler-Stevenson, W.G.; Bourboulia, D. Molecular mechanisms of tissue inhibitor of metalloproteinase 2 in the tumor microenvironment. Mol. Cell. Ther. 2014, 2, 17–24. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gender | (N) | Breed | (N) | Age (Years) | (N) | Tumor Location | (N) |
|---|---|---|---|---|---|---|---|
| Male | 24 | DSH | 16 | 5–7 | 3 | Gingiva | 8 |
| Female | 19 | DMH | 2 | 8–10 | 10 | Sublingual | 14 |
| DLH | 9 | 11–13 | 16 | Tongue | 5 | ||
| Persian | 1 | 14–17 | 15 | Buccal Mucosa | 5 | ||
| Burmese | 1 | Palate | 1 | ||||
| Oral cavity | 7 | ||||||
| Undisclosed | 2 | Undisclosed | 16 | Undisclosed | 1 | Undisclosed | 5 |
| Grade | Descriptions |
|---|---|
| Negative | less than 1% positive cells |
| Grade 1 | 1–9% positive cells (light, moderate or heavy staining) |
| Grade 2 | 10–50% light cells with scattered moderate or heavy staining |
| Grade 3 | mostly light, <50% moderate with scattered heavy staining |
| Grade 4Grade 5 | mostly moderate, <50% heavy staining>50% heavy staining |
| Primer Name | Primer Sequence | Amplicon Length | Intron-Spanning |
|---|---|---|---|
| * fPTGS1-S | GTCCTTCAACAGGGACTGGAC | 129 | no |
| fPTGS1-4-AS | ATGCTGGTTACTTATCTCGCTCC | ||
| * fPTGS2-S | GGCGTGAACCACGAGAAGTA | 136 | yes |
| fPTGS2-AS | GATGGCATGGACTGTGGTCA | ||
| * fCD147-S | ATTGACCCCACCGGTACCTA | 95 | yes |
| fCD147-AS | CCCAAAAGGACCTGAGCGAA | ||
| fTUBB-S | CAAGCGGTTTGCTGCTGTTA | 159 | yes |
| fTUBB-AS | GCTAGTCGGGACTGCTCTTC | ||
| fTBP-S | TCTATGAGAAGCGACGGAAGC | 117 | yes |
| fTBP-AS | GCCTTTGTCGTTGATGTGCC | ||
| ** fRPS18-S | TTGCCCAACACTTCACCCAT | 92 | yes |
| ** fRPS18-AS | AGGCGCAGTTTATGCTGTCT | ||
| ** fHPRT1-S | AATTGGGGCCCCCTTTTCTC | 158 | no |
| ** fHPRT1-AS | CCACCAAATGTGCTTGGCTT | ||
| fGAPDH-S | TGTCCATCCTTCGTCCCTCA | 144 | yes |
| fGAPDH-AS | CGGACACTCCAGAACCGTAG | ||
| ** fB2M-S | GTTCTCTCTTCCACAGGAGGC | 146 | yes |
| ** fB2M-AS | ACTCGCAAAATGTGCTGGAAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nasry, W.H.S.; Wang, H.; Jones, K.; Dirksen, W.P.; Rosol, T.J.; Rodriguez-Lecompte, J.C.; Martin, C.K. CD147 and Cyclooxygenase Expression in Feline Oral Squamous Cell Carcinoma. Vet. Sci. 2018, 5, 72. https://doi.org/10.3390/vetsci5030072
Nasry WHS, Wang H, Jones K, Dirksen WP, Rosol TJ, Rodriguez-Lecompte JC, Martin CK. CD147 and Cyclooxygenase Expression in Feline Oral Squamous Cell Carcinoma. Veterinary Sciences. 2018; 5(3):72. https://doi.org/10.3390/vetsci5030072
Chicago/Turabian StyleNasry, Walaa Hamed Shaker, Haili Wang, Kathleen Jones, Wessel P. Dirksen, Thomas J. Rosol, Juan Carlos Rodriguez-Lecompte, and Chelsea K. Martin. 2018. "CD147 and Cyclooxygenase Expression in Feline Oral Squamous Cell Carcinoma" Veterinary Sciences 5, no. 3: 72. https://doi.org/10.3390/vetsci5030072
APA StyleNasry, W. H. S., Wang, H., Jones, K., Dirksen, W. P., Rosol, T. J., Rodriguez-Lecompte, J. C., & Martin, C. K. (2018). CD147 and Cyclooxygenase Expression in Feline Oral Squamous Cell Carcinoma. Veterinary Sciences, 5(3), 72. https://doi.org/10.3390/vetsci5030072