Effects of 3-Month Exposure to E-Cigarette Aerosols on Glutamatergic Receptors and Transporters in Mesolimbic Brain Regions of Female C57BL/6 Mice

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. JUUL Exposure
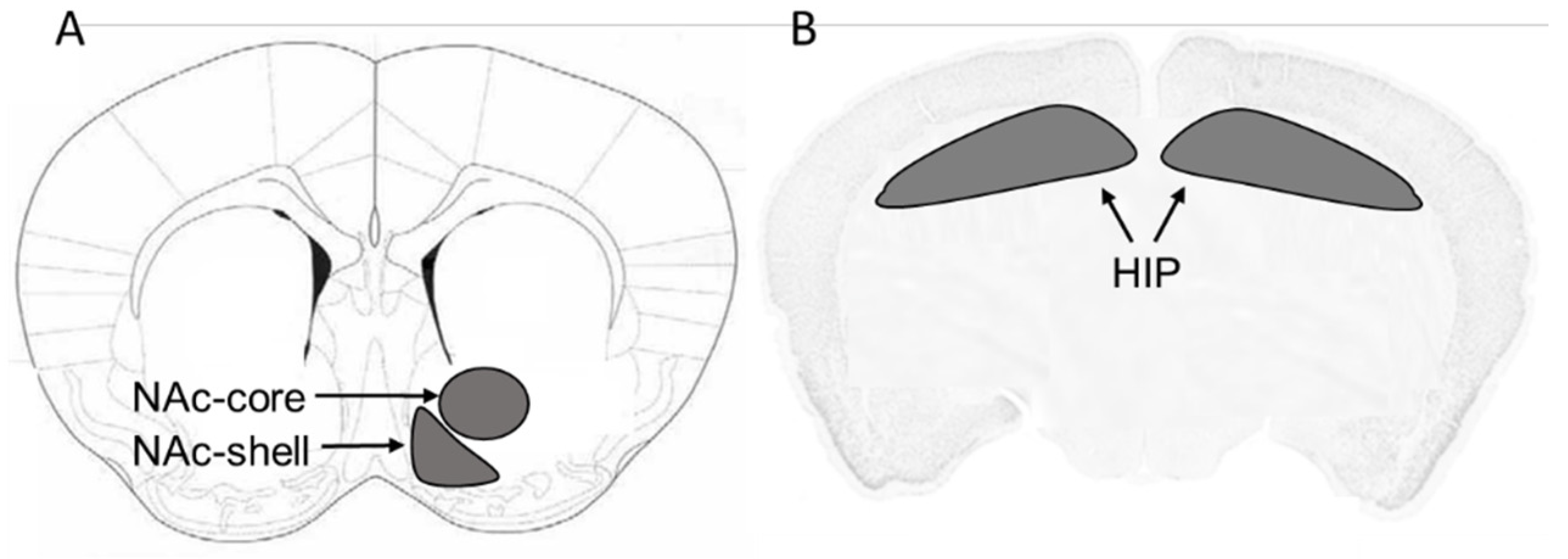
2.2. Brain Tissue Harvesting
2.3. Western Blot Technique
2.4. Statistical Analyses
3. Results
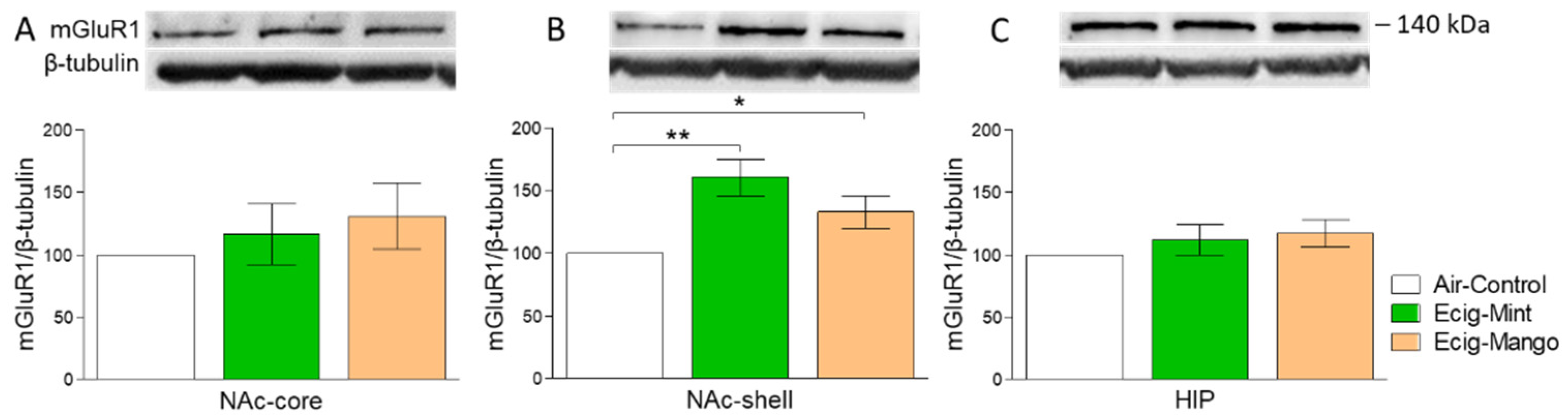
3.1. Effect of Three-Month Exposure to JUUL on mGluR1 Protein Expression
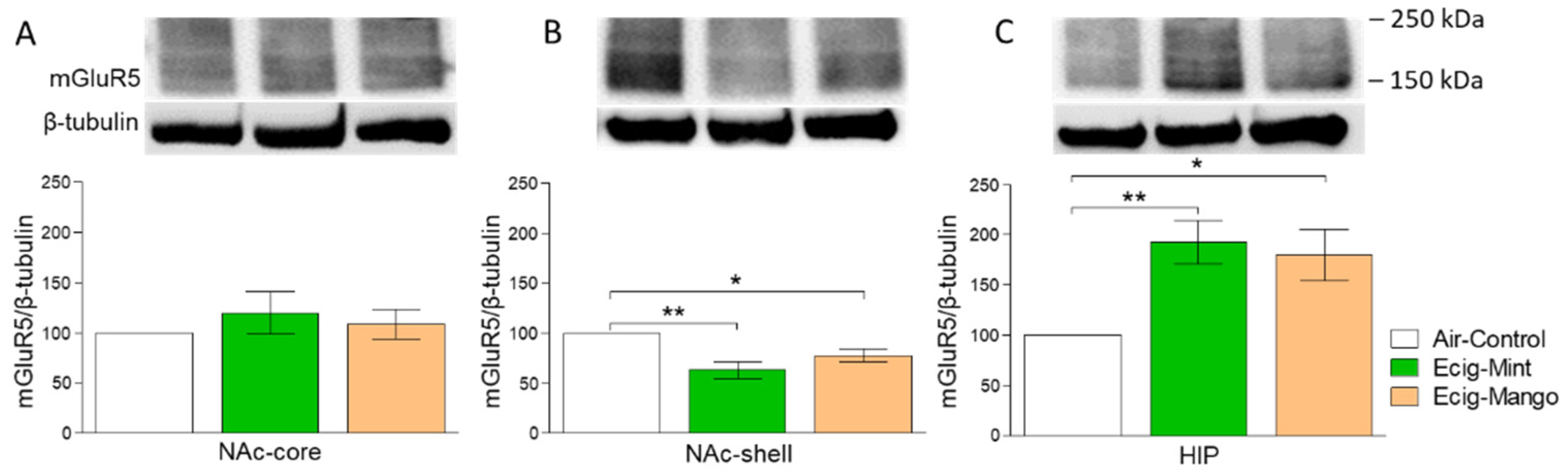
3.2. Effect of Three-Month Exposure to JUUL on mGluR5 Protein Expression
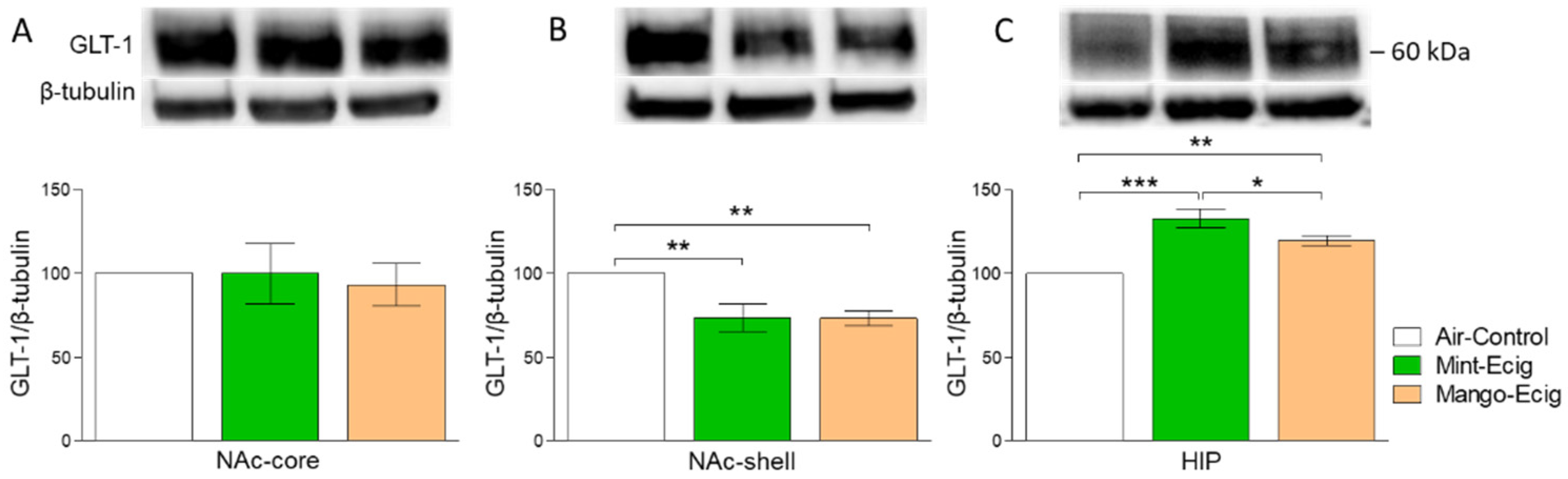
3.3. Effect of Three-Month Exposure to JUUL on GLT-1 Protein Expression
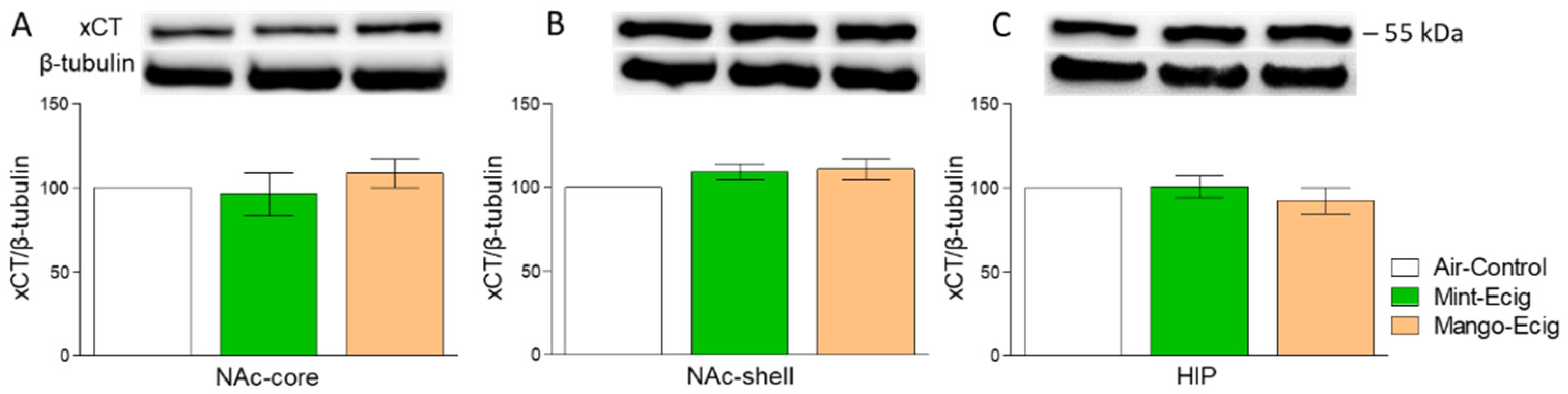
3.4. Effect of Three-Month Exposure to JUUL Pods on xCT Protein Expression
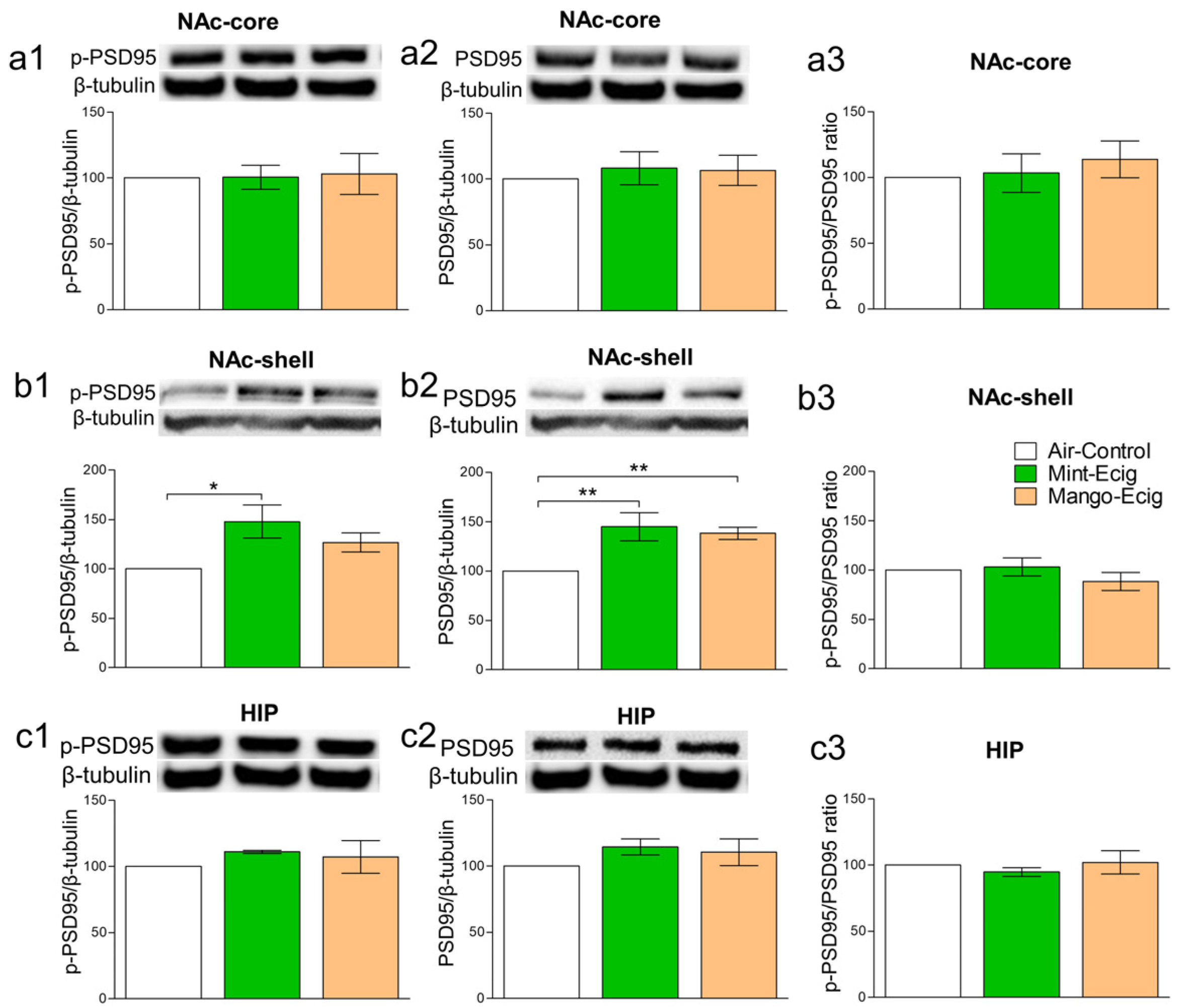
3.5. Effect of Three-Month Exposure to JUUL Pods on p-PSD95 and PSD95 Protein Expression
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cullen, K.A.; Ambrose, B.K.; Gentzke, A.S.; Apelberg, B.J.; Jamal, A.; King, B.A. Notes from the field: Use of electronic cigarettes and any tobacco product among middle and high school students—United States, 2011–2018. Mortal. Wkly. Rep. 2018, 67, 1276. [Google Scholar] [CrossRef]
- Alexander, L.E.C.; Vyas, A.; Schraufnagel, D.E.; Malhotra, A. Electronic cigarettes: The new face of nicotine delivery and addiction. J. Thorac. Dis. 2015, 7, E248. [Google Scholar]
- Yu, V.; Rahimy, M.; Korrapati, A.; Xuan, Y.; Zou, A.E.; Krishnan, A.R.; Tsui, T.; Aguilera, J.A.; Advani, S.; Alexander, L.E.C. Electronic cigarettes induce DNA strand breaks and cell death independently of nicotine in cell lines. Oral Oncol. 2016, 52, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Lyes, M.; Sladewski, K.; Enany, S.; McEachern, E.; Mathew, D.P.; Das, S.; Moshensky, A.; Bapat, S.; Pride, D.T. Electronic cigarette inhalation alters innate immunity and airway cytokines while increasing the virulence of colonizing bacteria. J. Mol. Med. 2016, 94, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Goniewicz, M.L.; Boykan, R.; Messina, C.R.; Eliscu, A.; Tolentino, J. High exposure to nicotine among adolescents who use Juul and other vape pod systems (‘pods’). Tob. Control 2019, 28, 676–677. [Google Scholar] [CrossRef] [PubMed]
- Willett, J.G.; Bennett, M.; Hair, E.C.; Xiao, H.; Greenberg, M.S.; Harvey, E.; Cantrell, J.; Vallone, D. Recognition, use and perceptions of JUUL among youth and young adults. Tob. Control 2019, 28, 115–116. [Google Scholar] [CrossRef]
- Jackler, R.K.; Ramamurthi, D. Nicotine arms race: JUUL and the high-nicotine product market. Tob. Control 2019, 28, 623–628. [Google Scholar] [CrossRef]
- US Food and Drug Administration. Statement from FDA Commissioner Scott Gottlieb, MD, on New Steps to Address Epidemic of Youth E-Cigarette Use; FDA: Montgomery, MD, USA, 2018.
- King, A.C.; Smith, L.J.; McNamara, P.J.; Matthews, A.K.; Fridberg, D.J. Passive exposure to electronic cigarette (e-cigarette) use increases desire for combustible and e-cigarettes in young adult smokers. Tob. Control 2015, 24, 501–504. [Google Scholar] [CrossRef]
- Vogel, E.A.; Prochaska, J.J.; Rubinstein, M.L. Measuring e-cigarette addiction among adolescents. Tob. Control 2020, 29, 258–262. [Google Scholar] [CrossRef]
- US Department of Health and Human Services. E-Cigarette Use among Youth and Young Adults; A Report of the Surgeon General; Public Health Service: Rockville, MD, USA, 2016.
- Pomerleau, C.S.; Pomerleau, O.F.; Garcia, A.W. Biobehavioral research on nicotine use in women. Br. J. Addict. 1991, 86, 527–531. [Google Scholar] [CrossRef]
- Killen, J.D.; Fortmann, S.P.; Newman, B.; Varady, A. Evaluation of a treatment approach combining nicotine gum with self-guided behavioral treatments for smoking relapse prevention. J. Consult. Clin. Psychol. 1990, 58, 85. [Google Scholar] [CrossRef] [PubMed]
- Fortmann, S.P.; Killen, J.D. Who shall quit? Comparison of volunteer and population-based recruitment in two minimal-contact smoking cessation studies. Am. J. Epidemiol. 1994, 140, 39–51. [Google Scholar] [CrossRef]
- Pang, R.D.; Goldenson, N.I.; Kirkpatrick, M.; Barrington-Trimis, J.L.; Cho, J.; Leventhal, A.M. Sex differences in the appeal of flavored e-cigarettes among young adult e-cigarette users. Psychol. Addict. Behav. 2020, 34, 303. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.-H.; Gamst, A.; Lee, M.; Cummins, S.; Yin, L.; Zoref, L. The use and perception of electronic cigarettes and snus among the US population. PLoS ONE 2013, 8, e79332. [Google Scholar]
- Alasmari, F.; Alexander, L.E.C.; Nelson, J.A.; Schiefer, I.T.; Breen, E.; Drummond, C.A.; Sari, Y. Effects of chronic inhalation of electronic cigarettes containing nicotine on glial glutamate transporters and α-7 nicotinic acetylcholine receptor in female CD-1 mice. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 77, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Alasmari, F.; Crotty Alexander, L.; Hammad, A.; Bojanowski, C.M.; Moshensky, A.; Sari, Y. Effects of chronic inhalation of electronic cigarette vapor containing nicotine on neurotransmitters in the frontal cortex and striatum of C57BL/6 mice. Front. Pharmacol. 2019, 10, 885. [Google Scholar] [CrossRef]
- Tanaka, K.; Watase, K.; Manabe, T.; Yamada, K.; Watanabe, M.; Takahashi, K.; Iwama, H.; Nishikawa, T.; Ichihara, N.; Kikuchi, T. Epilepsy and exacerbation of brain injury in mice lacking the glutamate transporter GLT-1. Science 1997, 276, 1699–1702. [Google Scholar] [CrossRef]
- Danbolt, N.C. Glutamate uptake. Prog. Neurobiol. 2001, 65, 1–105. [Google Scholar] [CrossRef]
- Althobaiti, Y.S.; Alshehri, F.S.; Almalki, A.H.; Sari, Y. Effects of ceftriaxone on glial glutamate transporters in Wistar rats administered sequential ethanol and methamphetamine. Front. Neurosci. 2016, 10, 427. [Google Scholar] [CrossRef]
- Hammad, A.M.; Alasmari, F.; Althobaiti, Y.S.; Sari, Y. Modulatory effects of Ampicillin/Sulbactam on glial glutamate transporters and metabotropic glutamate receptor 1 as well as reinstatement to cocaine-seeking behavior. Behav. Brain Res. 2017, 332, 288–298. [Google Scholar] [CrossRef]
- Alhaddad, H.; Das, S.C.; Sari, Y. Effects of ceftriaxone on ethanol intake: A possible role for xCT and GLT-1 isoforms modulation of glutamate levels in P rats. Psychopharmacology 2014, 231, 4049–4057. [Google Scholar] [CrossRef]
- Alasmari, F.; Bell, R.L.; Rao, P.; Hammad, A.M.; Sari, Y. Peri-adolescent drinking of ethanol and/or nicotine modulates astroglial glutamate transporters and metabotropic glutamate receptor-1 in female alcohol-preferring rats. Pharmacol. Biochem. Behav. 2018, 170, 44–55. [Google Scholar] [CrossRef]
- Alshehri, F.S.; Hakami, A.Y.; Althobaiti, Y.S.; Sari, Y. Effects of ceftriaxone on hydrocodone seeking behavior and glial glutamate transporters in P rats. Behav. Brain Res. 2018, 347, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Kenny, P.J.; Markou, A. The ups and downs of addiction: Role of metabotropic glutamate receptors. Trends Pharmacol. Sci. 2004, 25, 265–272. [Google Scholar] [CrossRef]
- Goodwani, S.; Saternos, H.; Alasmari, F.; Sari, Y. Metabotropic and ionotropic glutamate receptors as potential targets for the treatment of alcohol use disorder. Neurosci. Biobehav. Rev. 2017, 77, 14–31. [Google Scholar] [CrossRef]
- Gil-Sanz, C.; Delgado-García, J.M.; Fairén, A.; Gruart, A. Involvement of the mGluR1 receptor in hippocampal synaptic plasticity and associative learning in behaving mice. Cereb. Cortex 2008, 18, 1653–1663. [Google Scholar] [CrossRef] [PubMed]
- Rosenbrock, H.; Kramer, G.; Hobson, S.; Koros, E.; Grundl, M.; Grauert, M.; Reymann, K.G.; Schröder, U.H. Functional interaction of metabotropic glutamate receptor 5 and NMDA-receptor by a metabotropic glutamate receptor 5 positive allosteric modulator. Eur. J. Pharmacol. 2010, 639, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Tominaga-Yoshino, K.; Urakubo, T.; Okada, M.; Matsuda, H.; Ogura, A. Repetitive induction of late-phase LTP produces long-lasting synaptic enhancement accompanied by synaptogenesis in cultured hippocampal slices. Hippocampus 2008, 18, 281–293. [Google Scholar] [CrossRef]
- Romano, C.; Sesma, M.A.; McDonald, C.T.; O’malley, K.; van den Pol, A.N.; Olney, J.W. Distribution of metabotropic glutamate receptor mGluR5 immunoreactivity in rat brain. J. Comp. Neurol. 1995, 355, 455–469. [Google Scholar] [CrossRef]
- Harrison, A.A.; Gasparini, F.; Markou, A. Nicotine potentiation of brain stimulation reward reversed by DHβE and SCH 23390, but not by eticlopride, LY 314582 or MPEP in rats. Psychopharmacology 2002, 160, 56–66. [Google Scholar] [CrossRef]
- Paterson, N.E.; Markou, A. The metabotropic glutamate receptor 5 antagonist MPEP decreased break points for nicotine, cocaine and food in rats. Psychopharmacology 2005, 179, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Paterson, N.E.; Semenova, S.; Gasparini, F.; Markou, A. The mGluR5 antagonist MPEP decreased nicotine self-administration in rats and mice. Psychopharmacology 2003, 167, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Palmatier, M.I.; Liu, X.; Donny, E.C.; Caggiula, A.R.; Sved, A.F. Metabotropic glutamate 5 receptor (mGluR5) antagonists decrease nicotine seeking, but do not affect the reinforcement enhancing effects of nicotine. Neuropsychopharmacology 2008, 33, 2139–2147. [Google Scholar] [CrossRef][Green Version]
- Kane, J.; Hwang, Y.; Konu, O.; Loughlin, S.; Leslie, F.; Li, M. Regulation of Homer and group I metabotropic glutamate receptors by nicotine. Eur. J. Neurosci. 2005, 21, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.R.; Lawrence, A.J. The role of metabotropic glutamate receptors in addiction: Evidence from preclinical models. Pharmacol. Biochem. Behav. 2012, 100, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Dravolina, O.A.; Zakharova, E.S.; Shekunova, E.V.; Zvartau, E.E.; Danysz, W.; Bespalov, A.Y. mGlu1 receptor blockade attenuates cue-and nicotine-induced reinstatement of extinguished nicotine self-administration behavior in rats. Neuropharmacology 2007, 52, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Vansickel, A.R.; Eissenberg, T. Electronic cigarettes: Effective nicotine delivery after acute administration. Nicotine Tob. Res. 2012, 15, 267–270. [Google Scholar] [CrossRef]
- Pepper, J.; Ribisl, K.; Brewer, N. Adolescents’ interest in trying flavoured e-cigarettes. Tob. Control 2016, 25, ii62–ii66. [Google Scholar] [CrossRef]
- DeVito, E.E.; Krishnan-Sarin, S. E-cigarettes: Impact of e-liquid components and device characteristics on nicotine exposure. Curr. Neuropharmacol. 2018, 16, 438–459. [Google Scholar] [CrossRef]
- Berke, J.D.; Eichenbaum, H.B. Drug addiction and the hippocampus. Science 2001, 294, 1235. [Google Scholar] [CrossRef]
- Quintero, G.C.; treatment. Role of nucleus accumbens glutamatergic plasticity in drug addiction. Neuropsychiatr. Dis. 2013, 9, 1499. [Google Scholar] [CrossRef]
- Donny, E.; Caggiula, A.; Rowell, P.; Gharib, M.; Maldovan, V.; Booth, S.; Mielke, M.M.; Hoffman, A.; McCallum, S. Nicotine self-administration in rats: Estrous cycle effects, sex differences and nicotinic receptor binding. Psychopharmacology 2000, 151, 392–405. [Google Scholar] [CrossRef] [PubMed]
- Cheeta, S.; Irvine, E.E.; Tucci, S.; Sandhu, J.; File, S.E. In adolescence, female rats are more sensitive to the anxiolytic effect of nicotine than are male rats. Neuropsychopharmacology 2001, 25, 601–607. [Google Scholar] [CrossRef]
- Crotty Alexander, L.; Vitorino, S.; Moshensky, A.; Shin, J.; Chien, J.; Olay, J.; Nilaad, S.; Jahan, A.; Chen, Y.; Advani, I. Inhalation of JUUL Aerosols and Flavor Choice Adversely Affects Inflammatory and Metabolic States. In B107. Effects of E-Cigarettes and Their Components on Respiratory Dysfunction, Inflammation, and Repair; American Thoracic Society: New York, NY, USA, 2019; p. A4182. [Google Scholar]
- Franklin, K.B.J.; Paxinos, G. The Mouse Brain in Stereotaxic Coordinates, Compact 3rd Edition, 4th ed.; Elsevier: Amsterdam, The Netherlands, 2008; p. 1. [Google Scholar]
- Sheng, M. The postsynaptic NMDA-receptor—PSD-95 signaling complex in excitatory synapses of the brain. J. Cell Sci. 2001, 114, 1251. [Google Scholar]
- Martel, M.-A.; Ryan, T.J.; Bell, K.F.; Fowler, J.H.; McMahon, A.; Al-Mubarak, B.; Komiyama, N.H.; Horsburgh, K.; Kind, P.C.; Grant, S.G. The subtype of GluN2 C-terminal domain determines the response to excitotoxic insults. Neuron 2012, 74, 543–556. [Google Scholar] [CrossRef]
- Li, B.; Otsu, Y.; Murphy, T.H.; Raymond, L.A. Developmental decrease in NMDA receptor desensitization associated with shift to synapse and interaction with postsynaptic density-95. J. Neurosci. 2003, 23, 11244–11254. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.Q.; Zhu, J.; Pei, D.S.; Zong, Y.Y.; Yan, J.Z.; Hou, X.Y.; Zhang, G.Y. Overexpression of the PDZ1 domain of PSD-95 diminishes ischemic brain injury via inhibition of the GluR6· PSD-95· MLK3 pathway. J. Neurosci. Res. 2009, 87, 3626–3638. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.; Sheng, M. PDZ domain proteins of synapses. Nat. Rev. Neurosci. 2004, 5, 771–781. [Google Scholar] [CrossRef]
- Pontieri, F.E.; Tanda, G.; Orzi, F.; Di Chiara, G. Effects of nicotine on the nucleus accumbens and similarity to those of addictive drugs. Nature 1996, 382, 255–257. [Google Scholar] [CrossRef]
- Benwell, M.; Balfour, D. The effects of acute and repeated nicotine treatment on nucleus accumbens dopamine and locomotor activity. Br. J. Pharmacol. 1992, 105, 849. [Google Scholar] [CrossRef]
- Balfour, D. Neural mechanisms underlying nicotine dependence. Addiction 1994, 89, 1419–1423. [Google Scholar] [CrossRef]
- Heimer, L.; Alheid, G.F.; De Olmos, J.; Groenewegen, H.J.; Haber, S.N.; Harlan, R.E.; Zahm, D.S. The accumbens: Beyond the core-shell dichotomy. J. Neuropsychiatry Clin. Neurosci. 1997, 9, 354–381. [Google Scholar]
- Groenewegen, H.J.; Wright, C.I.; Beijer, A.V.; Voorn, P. Convergence and segregation of ventral striatal inputs and outputs. Ann. N. Y. Acad. Sci. 1999, 877, 49–63. [Google Scholar] [CrossRef]
- Balfour, D.J. Neuroplasticity within the mesoaccumbens dopamine system and its role in tobacco dependence. Curr. Drug Targets CNS Neurol. Disord. 2002, 1, 413–421. [Google Scholar] [CrossRef] [PubMed]
- Keistler, C.; Barker, J.M.; Taylor, J.R. Infralimbic prefrontal cortex interacts with nucleus accumbens shell to unmask expression of outcome-selective Pavlovian-to-instrumental transfer. Learn. Mem. 2015, 22, 509–513. [Google Scholar] [CrossRef] [PubMed]
- Augur, I.F.; Wyckoff, A.R.; Aston-Jones, G.; Kalivas, P.W.; Peters, J. Chemogenetic activation of an extinction neural circuit reduces cue-induced reinstatement of cocaine seeking. J. Neurosci. 2016, 36, 10174–10180. [Google Scholar] [CrossRef] [PubMed]
- Liechti, M.E.; Lhuillier, L.; Kaupmann, K.; Markou, A. Metabotropic glutamate 2/3 receptors in the ventral tegmental area and the nucleus accumbens shell are involved in behaviors relating to nicotine dependence. J. Neurosci. 2007, 27, 9077–9085. [Google Scholar] [CrossRef] [PubMed]
- Saal, D.; Dong, Y.; Bonci, A.; Malenka, R.C. Drugs of abuse and stress trigger a common synaptic adaptation in dopamine neurons. Neuron 2003, 37, 577–582. [Google Scholar] [CrossRef]
- D’Souza, M.S.; Markou, A. Metabotropic glutamate receptor 5 antagonist 2-methyl-6-(phenylethynyl) pyridine (MPEP) microinfusions into the nucleus accumbens shell or ventral tegmental area attenuate the reinforcing effects of nicotine in rats. Neuropharmacology 2011, 61, 1399–1405. [Google Scholar] [CrossRef]
- Byrnes, K.R.; Stoica, B.; Loane, D.J.; Riccio, A.; Davis, M.I.; Faden, A.I. Metabotropic glutamate receptor 5 activation inhibits microglial associated inflammation and neurotoxicity. Glia 2009, 57, 550–560. [Google Scholar] [CrossRef]
- Zhang, Z.-Y.; Sun, B.-L.; Liu, J.-K.; Yang, M.-F.; Li, D.-W.; Fang, J.; Zhang, S.; Yuan, Q.-L.; Huang, S.-L. Activation of mGluR5 attenuates microglial activation and neuronal apoptosis in early brain injury after experimental subarachnoid hemorrhage in rats. Neurochem. Res. 2015, 40, 1121–1132. [Google Scholar] [CrossRef]
- Alhaddad, H.; Gordon, D.M.; Bell, R.L.; Jarvis, E.E.; Kipp, Z.A.; Hinds, T.D., Jr.; Sari, Y. Chronic Ethanol Consumption Alters Glucocorticoid Receptor Isoform Expression in Stress Neurocircuits and Mesocorticolimbic Brain Regions of Alcohol-Preferring Rats. Neuroscience 2020, 437, 107–116. [Google Scholar] [CrossRef]
- Alasmari, F.; Alhaddad, H.; Wong, W.; Bell, R.L.; Sari, Y. Ampicillin/Sulbactam Treatment Modulates NMDA Receptor NR2B Subunit and Attenuates Neuroinflammation and Alcohol Intake in Male High Alcohol Drinking Rats. Biomolecules 2020, 10, 1030. [Google Scholar] [CrossRef]
- Hubert, G.W.; Paquet, M.; Smith, Y. Differential subcellular localization of mGluR1a and mGluR5 in the rat and monkey substantia nigra. J. Neurosci. 2001, 21, 1838–1847. [Google Scholar] [CrossRef]
- Shigemoto, R.; Nakanishi, S.; Mizuno, N. Distribution of the mRNA for a metabotropic glutamate receptor (mGluR1) in the central nervous system: An in situ hybridization study in adult and developing rat. J. Comp. Neurol. 1992, 322, 121–135. [Google Scholar] [CrossRef] [PubMed]
- Knackstedt, L.A.; LaRowe, S.; Mardikian, P.; Malcolm, R.; Upadhyaya, H.; Hedden, S.; Markou, A.; Kalivas, P.W. The role of cystine-glutamate exchange in nicotine dependence in rats and humans. Biol. Psychiatry 2009, 65, 841–845. [Google Scholar] [CrossRef]
- Wang, H.; Sun, X. Desensitized nicotinic receptors in brain. Brain Res. Rev. 2005, 48, 420–437. [Google Scholar] [CrossRef]
- Stefanik, M.T.; Kupchik, Y.M.; Kalivas, P.W. Optogenetic inhibition of cortical afferents in the nucleus accumbens simultaneously prevents cue-induced transient synaptic potentiation and cocaine-seeking behavior. Brain Struct. Funct. 2016, 221, 1681–1689. [Google Scholar] [CrossRef]
- Futai, K.; Kim, M.J.; Hashikawa, T.; Scheiffele, P.; Sheng, M.; Hayashi, Y. Retrograde modulation of presynaptic release probability through signaling mediated by PSD-95–neuroligin. Nat. Neurosci. 2007, 10, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, I.; Malinow, R. Postsynaptic density 95 controls AMPA receptor incorporation during long-term potentiation and experience-driven synaptic plasticity. J. Neurosci. 2004, 24, 916–927. [Google Scholar] [CrossRef] [PubMed]
- El-Husseini, A.E.; Craven, S.E.; Chetkovich, D.M.; Firestein, B.L.; Schnell, E.; Aoki, C.; Bredt, D.S. Dual palmitoylation of PSD-95 mediates its vesiculotubular sorting, postsynaptic targeting, and ion channel clustering. J. Cell Biol. 2000, 148, 159–172. [Google Scholar] [CrossRef]
- Kornau, H.-C.; Schenker, L.T.; Kennedy, M.B.; Seeburg, P.H. Domain interaction between NMDA receptor subunits and the postsynaptic density protein PSD-95. Science 1995, 269, 1737–1740. [Google Scholar] [CrossRef] [PubMed]
- Rezvani, K.; Teng, Y.; Shim, D.; De Biasi, M. Nicotine regulates multiple synaptic proteins by inhibiting proteasomal activity. J. Neurosci. 2007, 27, 10508–10519. [Google Scholar] [CrossRef] [PubMed]
- Pistillo, F.; Fasoli, F.; Moretti, M.; McClure-Begley, T.; Zoli, M.; Marks, M.J.; Gotti, C. Chronic nicotine and withdrawal affect glutamatergic but not nicotinic receptor expression in the mesocorticolimbic pathway in a region-specific manner. Pharmacol. Res. 2016, 103, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chen, H.; Steketee, J.D.; Sharp, B.M. Upregulation of ionotropic glutamate receptor subunits within specific mesocorticolimbic regions during chronic nicotine self-administration. Neuropsychopharmacology 2007, 32, 103–109. [Google Scholar] [CrossRef]
- Kalivas, P.W. Recent understanding in the mechanisms of addiction. Curr. Psychiatry Rep. 2004, 6, 347–351. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.E.; Sun, X.; Mangiavacchi, S.; Chao, S.Z. Psychomotor stimulants and neuronal plasticity. Neuropharmacology 2004, 47, 61–79. [Google Scholar] [CrossRef]
- Gould, T.J.; Davis, J.A. Associative learning, the hippocampus, and nicotine addiction. Curr. Drug Abus. Rev. 2008, 1, 9–19. [Google Scholar] [CrossRef]
- Jia, Y.; Yamazaki, Y.; Nakauchi, S.; Ito, K.I.; Sumikawa, K. Nicotine facilitates long-term potentiation induction in oriens-lacunosum moleculare cells via Ca2+ entry through non-α7 nicotinic acetylcholine receptors. Eur. J. Neurosci. 2010, 31, 463–476. [Google Scholar] [CrossRef]
- Welsby, P.; Rowan, M.; Anwyl, R. Nicotinic receptor-mediated enhancement of long-term potentiation involves activation of metabotropic glutamate receptors and ryanodine-sensitive calcium stores in the dentate gyrus. Eur. J. Neurosci. 2006, 24, 3109–3118. [Google Scholar] [CrossRef]
- Fujii, S.; Ji, Z.; Morita, N.; Sumikawa, K. Acute and chronic nicotine exposure differentially facilitate the induction of LTP. Brain Res. 1999, 846, 137–143. [Google Scholar] [CrossRef]
- Gould, T.J.; Wilkinson, D.S.; Yildirim, E.; Blendy, J.A.; Adoff, M.D. Dissociation of tolerance and nicotine withdrawal-associated deficits in contextual fear. Brain Res. 2014, 1559, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.S.; Gould, T.J. Withdrawal from chronic nicotine and subsequent sensitivity to nicotine challenge on contextual learning. Behav. Brain Res. 2013, 250, 58–61. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alhaddad, H.; Wong, W.; Sari, A.T.; Crotty Alexander, L.E.; Sari, Y. Effects of 3-Month Exposure to E-Cigarette Aerosols on Glutamatergic Receptors and Transporters in Mesolimbic Brain Regions of Female C57BL/6 Mice. Toxics 2020, 8, 95. https://doi.org/10.3390/toxics8040095
Alhaddad H, Wong W, Sari AT, Crotty Alexander LE, Sari Y. Effects of 3-Month Exposure to E-Cigarette Aerosols on Glutamatergic Receptors and Transporters in Mesolimbic Brain Regions of Female C57BL/6 Mice. Toxics. 2020; 8(4):95. https://doi.org/10.3390/toxics8040095
Chicago/Turabian StyleAlhaddad, Hasan, Woonyen Wong, Adam T. Sari, Laura E. Crotty Alexander, and Youssef Sari. 2020. "Effects of 3-Month Exposure to E-Cigarette Aerosols on Glutamatergic Receptors and Transporters in Mesolimbic Brain Regions of Female C57BL/6 Mice" Toxics 8, no. 4: 95. https://doi.org/10.3390/toxics8040095
APA StyleAlhaddad, H., Wong, W., Sari, A. T., Crotty Alexander, L. E., & Sari, Y. (2020). Effects of 3-Month Exposure to E-Cigarette Aerosols on Glutamatergic Receptors and Transporters in Mesolimbic Brain Regions of Female C57BL/6 Mice. Toxics, 8(4), 95. https://doi.org/10.3390/toxics8040095