

Neonicotinoid-Induced Cytotoxicity: Insights into Cellular Mechanisms and Health Risks


Abstract

1. Introduction
2. The Molecular Mechanism of Neonicotinoids Inducing Cytotoxicity
2.1. Oxidative Stress Induced by Neonicotinoids
2.2. DNA Damage and Genotoxic Effects of Neonicotinoids
2.3. Neonicotinoid-Induced Cell Apoptosis: Pathways and Evidence
2.4. Endoplasmic Reticulum Stress Response Triggered by Neonicotinoid Exposure
2.5. Disruption of Calcium Homeostasis in Cells by Neonicotinoids
2.6. Immunotoxicity Mechanisms Mediated by Neonicotinoids
3. Cell Type Specific Response and Organ System Toxicity of Neonicotinoids
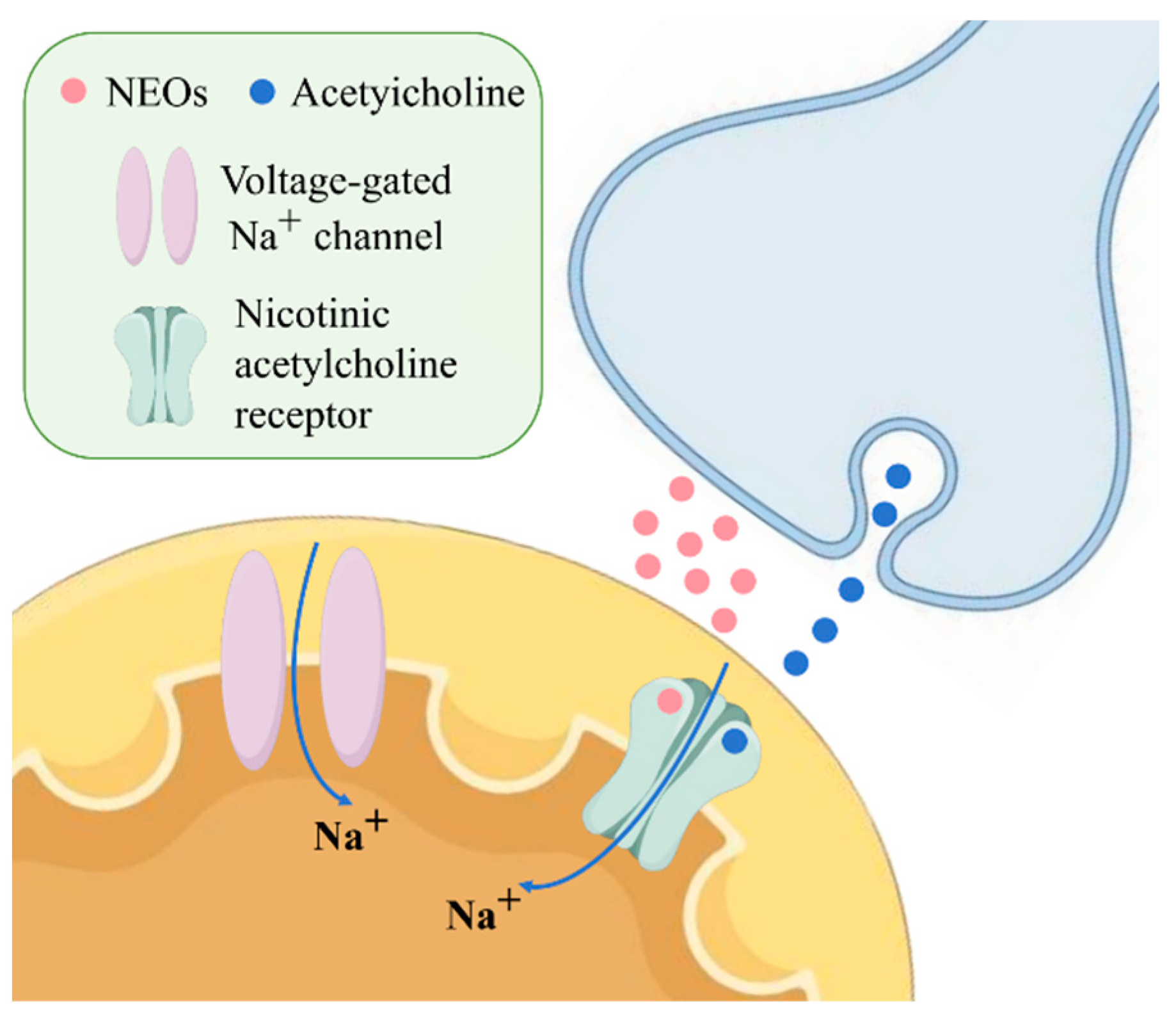
3.1. Neurotoxicity of Neonicotinoids in Neuronal Cells
3.2. Reproductive Toxicity of Neonicotinoids in Reproductive Cells and Tissues
3.3. Hepatotoxic and Nephrotoxic Effects of Neonicotinoids on Hepatocytes and Renal Tubular Cells
3.4. Immunotoxic Effects of Neonicotinoids on Immune Cell Function and Viability
3.5. Epithelial Cell Damage and Barrier Function Disruption Induced by Neonicotinoids
3.6. Toxicological Effects of Neonicotinoids on Other Cell Types and Organ Systems
4. Public Health Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Motaung, T.E. Chloronicotinyl insecticide imidacloprid: Agricultural relevance, pitfalls and emerging opportunities. Crop. Prot. 2020, 131, 105097. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhu, W.; Wang, Y.; Li, X.; Lv, J.; Luo, J.; Yang, M. Insight of neonicotinoid insecticides: Exploring exposure, mechanisms in non-target organisms, and removal technologies. Pharmacol. Res. 2024, 209, 107415. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Anadón, A.; Wu, Q.; Qiao, F.; Ares, I.; Martínez-Larrañaga, M.R.; Yuan, Z.; Martínez, M.A. Mechanism of Neonicotinoid Toxicity: Impact on Oxidative Stress and Metabolism. Annu. Rev. Pharmacol. 2018, 58, 471–507. [Google Scholar] [CrossRef] [PubMed]
- Zoltán, T.; Zsófia, K. Chronic acetamiprid exposure moderately affects the foraging behaviour of buff-tailed bumblebees (Bombus terrestris). Ethology 2023, 130, 13425. [Google Scholar] [CrossRef]
- Costas-Ferreira, C.; Faro, L.R.F. Neurotoxic effects of neonicotinoids on mamma1s what is there beyond the activation of nicotinic acety1cho1ine receptors a systematic review. Int. J. Mol. Sci. 2021, 22, 8413. [Google Scholar] [CrossRef]
- Matsuda, K.; Kanaoka, S.; Akamatsu, M.; Sattelle, D.B. Diverse actions and target-site selectivity of neonicotinoids: Structural insights. Mol. Pharmacol. 2009, 76, 1–10. [Google Scholar] [CrossRef]
- Chen, D.; Zhang, Y.; Lv, B.; Liu, Z.; Han, J.; Li, J.; Zhao, Y.; Wu, Y. Dietary exposure to neonicotinoid insecticides and health risks in the Chinese general population through two consecutive total diet studies. Environ. Int. 2020, 135, 105399. [Google Scholar] [CrossRef]
- Wood, T.J.; Goulson, D. The environmental risks of neonicotinoid pesticides: A review of the evidence post 2013. Environ. Sci. Pollut. Res. 2017, 24, 17285–17325. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, C.; Lu, X.; Yu, B.; Huang, Y. Advances in Research on Toxic Effects of Neonicotinoid Insecticides on Non-target Organisms. Asian J. Ecotoxicol. 2020, 15, 56–71. [Google Scholar]
- Saito, H.; Furukawa, Y.; Sasaki, T.; Kitajima, S.; Kanno, J.; Tanemura, K. Behavioral effects of adult male mice induced by low-level acetamiprid, imidacloprid, and nicotine exposure in early-life. Front. Neurosci. 2023, 17, 1239808. [Google Scholar] [CrossRef]
- Gawade, L.; Dadarkar, S.S.; Husain, R.; Gatne, M. A detailed study of developmental immunotoxicity of imidacloprid in Wistar rats. Food Chem. Toxicol. 2013, 51, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Cao, Y.; Pan, Q.; Xiao, Y.; Wang, Y.; Wang, X.; Li, X.; Li, Q.; Tang, X.; Ran, B. Neonicotinoid insecticides triggers mitochondrial bioenergetic dysfunction via manipulating ROS-calcium influx pathway in the liver. Ecotoxicol. Environ. Saf. 2021, 224, 112690. [Google Scholar] [CrossRef]
- Karaca, B.U.; Arican, Y.E.; Boran, T.; Binay, S.; Okyar, A.; Kaptan, E.; Özhan, G. Toxic effects of subchronic oral acetamiprid exposure in rats. Toxicol. Ind. Health 2019, 35, 679–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wang, Y.; Xiang, H.; Jia, H.; Zhang, Z.; Wang, X. Nephrotoxicity of Acetamiprid on Male Mice and the Rescue Role of Vitamin E. J. Anim. Vet. Adv. 2012, 11, 2721–2726. [Google Scholar]
- Cimino, A.M.; Boyles, A.L.; Thayer, K.A.; Perry, M.J. Effects of neonicotinoid pesticide exposure on human health: A systematic review. Environ. Health Perspect. 2017, 125, 155–162. [Google Scholar] [CrossRef]
- Han, W.; Tian, Y.; Shen, X. Human exposure to neonicotinoid insecticides and the evaluation of their potential toxicity: An overview. Chemosphere 2018, 192, 59–65. [Google Scholar] [CrossRef]
- Casida, J.E. Pesticide detox by design. J. Agric. Food Chem. 2018, 66, 9379–9383. [Google Scholar] [CrossRef]
- Cartereau, A.; Martin, C.; Thany, S.H. Neonicotinoid insecticides differently modulate acetycholine-induced currents on mammalian α7 nicotinic acetylcholine receptors. Br. J. Pharmacol. 2018, 175, 1987–1998. [Google Scholar] [CrossRef]
- Xu, X.; Wang, X.; Yang, Y.; Ares, I.; Martinez, M.; Lopez-Torres, B.; Martínez-Larrañaga, M.R.; Wang, X.; Anadón, A.; Martinez, M.A. Neonicotinoids: Mechanisms of systemic toxicity based on oxidative stress-mitochondrial damage. Arch. Toxicol. 2022, 96, 1493–1520. [Google Scholar] [CrossRef]
- Schwarzbacherová, V.; Wnuk, M.; Deregowska, A.; Holečková, B.; Lewinska, A. In vitro exposure to thiacloprid-based insecticide formulation promotes oxidative stress, apoptosis and genetic instability in bovine lymphocytes. Toxicol. Vitr. 2019, 61, 104654. [Google Scholar] [CrossRef]
- Khidkhan, K.; Ikenaka, Y.; Ichise, T.; Nakayama, S.M.M.; Mizukawa, H.; Nomiyama, K.; Iwata, H.; Arizono, K.; Takahashi, K.; Kato, K.; et al. Interspecies differences in cytochrome P450-mediated metabolism of neonicotinoids among cats, dogs, rats, and humans. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 239, 108898. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.; Fu, L.; Wang, J.; Sun, X.; Wang, X.; Lou, J.; Zhao, M.; Wang, C.; Zhang, Q. MtDNA Copy Number in Oral Epithelial Cells Serves as a Potential Biomarker of Mitochondrial Damage by Neonicotinoid Exposure: A Cross-Sectional Study. Environ. Sci. Technol. 2023, 57, 15816–15824. [Google Scholar] [CrossRef]
- Ge, W.; Yan, S.; Wang, J.; Zhu, L.; Chen, A.; Wang, J. Oxidative stress and DNA damage induced by imidacloprid in zebrafish (Danio rerio). J. Agric. Food Chem. 2015, 63, 1856–1862. [Google Scholar] [CrossRef]
- Rymuszka, A.; Sieroslawska, A. Comparative evaluation of neonicotinoids and their metabolites-induced oxidative stress in carp primary leukocytes and CLC cells. Sci. Rep. 2024, 14, 8291. [Google Scholar] [CrossRef]
- Senyildiz, M.; Kilinc, A.; Ozden, S. Investigation of the genotoxic and cytotoxic effects of widely used neonicotinoid insecticides in HepG2 and SH-SY5Y cells. Toxicol. Ind. Health 2018, 34, 375–383. [Google Scholar] [CrossRef]
- Bianchi, M.; Paravani, E.V.; Acosta, M.G.; Odetti, L.M.; Simoniello, M.F.; Poletta, G.L. Pesticide-induced alterations in zebrafish (Danio rerio) behavior, histology, DNA damage and mRNA expression: An integrated approach. Comp. Biochem. Phys. C 2024, 280, 109895. [Google Scholar] [CrossRef] [PubMed]
- Al-Sarar, A.S.; Abobakr, Y.; Bayoumi, A.E.; Hussein, H.I. Cytotoxic and genotoxic effects of abamectin, chlorfenapyr, and imidacloprid on CHOK1 cells. Environ. Sci. Pollut. Res. 2015, 22, 17041–17052. [Google Scholar] [CrossRef] [PubMed]
- Sekeroglu, Z.A.; Sekeroglu, V.; Aydin, B.; Yedier, S.K.; Ilkun, E. Clothianidin induces DNA damage and oxidative stress in bronchial epithelial cells. Environ. Mol. Mutagen. 2020, 61, 647–655. [Google Scholar] [CrossRef]
- Baysal, M.; Atlı-Eklioğlu, Ö. Comparison of the toxicity of pure compounds and commercial formulations of imidacloprid and acetamiprid on HT-29 cells: Single and mixture exposure. Food Chem. Toxicol. 2021, 155, 112430. [Google Scholar] [CrossRef]
- Xie, Z.; Lu, G.; Zhou, R.; Ma, Y. Thiacloprid-induced hepatotoxicity in zebrafish: Activation of the extrinsic and intrinsic apoptosis pathways regulated by p53 signaling pathway. Aquat. Toxicol. 2022, 246, 106147. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, A.; Shan, T.; Dong, W.; Shi, X.; Gao, X. Cross-resistance and fitness cost analysis of resistance to thiamethoxam in Melon and Cotton Aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2020, 113, 1946–1954. [Google Scholar] [CrossRef] [PubMed]
- Duzguner, V.; Erdogan, S. Chronic exposure to imidacloprid induces inflammation and oxidative stress in the liver & central nervous system of rats. Pestic. Biochem. Phys. 2012, 104, 58–64. [Google Scholar] [CrossRef]
- Cheng, W.; Zhang, Z.; Zhang, Z.; Zhao, G.; Wang, Y. Clothianidin exposure induces cell apoptosis via mitochondrial oxidative damage. Environ. Toxicol. 2025, 40, 574–582. [Google Scholar] [CrossRef]
- Li, J.; Li, J.; Zhai, L.; Lu, K. Co-exposure of polycarbonate microplastics aggravated the toxic effects of imidacloprid on the liver and gut microbiota in mice. Environ. Toxicol. Pharmacol. 2023, 101, 104194. [Google Scholar] [CrossRef]
- Öztaş, E.; Kara, M.; Boran, T.; Bişirir, E.; Karaman, E.F.; Kaptan, E.; Özhan, G. Cellular stress pathways are linked to acetamiprid-induced apoptosis in SH-SY5Y neural cells. Biology 2021, 10, 820. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Li, M.; Cao, X.; Li, R.; Jin, S.; Yang, H.; Xu, L.; Wang, P.; Bi, J. IRE1α-XBP1 affects the mitochondrial function of Aβ25-35-treated SH-SY5Y cells by regulating mitochondria-associated endoplasmic reticulum membranes. Front. Cell. Neurosci. 2021, 15, 614556. [Google Scholar] [CrossRef]
- El Gazzar, W.B.; Bayoumi, H.; Youssef, H.S.; Ibrahim, T.A.; Abdelfatah, R.M.; Gamil, N.M.; Iskandar, M.K.; Abdel-Kareim, A.M.; Abdelrahman, S.M.; Gebba, M.A.; et al. Role of IRE1α/XBP1/CHOP/NLRP3 signalling pathway in neonicotinoid imidacloprid-induced pancreatic dysfunction in rats and antagonism of lycopene: In vivo and molecular docking simulation approaches. Toxics 2024, 12, 445. [Google Scholar] [CrossRef]
- Liu, Y.; He, Q.; Xu, Z.; Xu, C.; Zhao, S.; Luo, Y.; Sun, X.; Qi, Z.; Wang, H. Thiamethoxam exposure induces endoplasmic reticulum stress and affects ovarian function and oocyte development in mice. J. Agric. Food Chem. 2021, 69, 1942–1952. [Google Scholar] [CrossRef]
- Lin, S.; Xiao, Y.; Li, S.; Tang, L.; Shi, H.; Hong, M.; Ding, L. Thiacloprid exposure induces oxidative stress, endoplasmic reticulum stress, and apoptosis in the liver of Mauremys reevesii. Ecol. Evol. 2025, 15, e70936. [Google Scholar] [CrossRef]
- Hirano, T.; Minagawa, S.; Furusawa, Y.; Yunoki, T.; Ikenaka, Y.; Yokoyama, T.; Hoshi, N.; Tabuchi, Y. Growth and neurite stimulating effects of the neonicotinoid pesticide clothianidin on human neuroblastoma SH-SY5Y cells. Toxicol. Appl. Pharmacol. 2019, 383, 114777. [Google Scholar] [CrossRef]
- Izuru, K.; Kiyomi, T. Exposure to neonicotinoid pesticides induces physiological disorders and affects color performance and foraging behavior in goldfish. Physiol. Rep. 2024, 12, e10138. [Google Scholar] [CrossRef]
- Costas-Ferreira, C.; Faro Lilian, R.F. Systematic Review of Calcium Channels and Intracellular Calcium Signaling: Relevance to Pesticide Neurotoxicity. Int. J. Mol. Sci. 2022, 22, 13376. [Google Scholar] [CrossRef] [PubMed]
- Torres-Sanchez, E.D.; Ortiz, G.G.; Reyes-Uribe, E.; Torres-Jasso, J.H.; Salazar-Flores, J. Effect of pesticides on phosphorylation of tau protein, and its influence on Alzheimer’s disease. World. J. Clin. Cases 2023, 11, 5628–5642. [Google Scholar] [CrossRef] [PubMed]
- Walderdorff, L.; Laval-Gilly, P.; Wechtler, L.; Bonnefoy, A.; Falla-Angel, J. Phagocytic activity of human macrophages and Drosophila hemocytes after exposure to the neonicotinoid imidacloprid. Pestic. Biochem. Phys. 2019, 160, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Badgujar, P.C.; Jain, S.K.; Singh, A.; Punia, J.S.; Gupta, R.P.; Chandratre, G.A. Immunotoxic effects of imidacloprid following 28 days of oral exposure in BALB/c mice. Environ. Toxicol. Pharmacol. 2013, 35, 408–418. [Google Scholar] [CrossRef]
- Yang, Y.; Yu, Q.; Zhang, C.; Wang, X.; He, L.; Huang, Y.; Li, E.; Qin, J.; Chen, L. Acute thiamethoxam exposure induces hepatotoxicity and neurotoxicity in juvenile Chinese mitten crab (Eriocheir sinensis). Ecotoxicol. Environ. Saf. 2023, 249, 114399. [Google Scholar] [CrossRef]
- Ramirez-Candoa, L.J.; Guzmán-Vallejosa, M.S.; Aguayob, L.G.; Vera-Erazob, F.D.; Ballazc, S.J. Neurocytotoxicity of imidacloprid- and acetamiprid-based comercial insecticides over the differentiation of SH-SY5Y neuroblastoma cells. Heliyon 2023, 9, e15840. [Google Scholar] [CrossRef]
- Cheng, L.; Lu, Y.; Zhao, Z.; Hoogenboom, R.L.A.P.; Zhang, Q.; Liu, X. Assessing the combined toxicity effects of three neonicotinoid pesticide mixtures on human neuroblastoma SK-N-SH and lepidopteran Sf-9 cells. Food Chem. Toxicol. 2020, 145, 111632. [Google Scholar] [CrossRef]
- Loser, D.; Hinojosa, M.G.; Blum, J.; Schaefer, J.; Brüll, M.; Johansson, Y.; Suciu, I.; Grillberger, K.; Danker, T.; Möller, C.; et al. Functional alterations by a subgroup of neonicotinoid pesticides in human dopaminergic neurons. Arch. Toxicol. 2021, 95, 2081–2107. [Google Scholar] [CrossRef]
- Wang, X.; Qiu, J.; Xu, Y.; Pan, Y.; Chen, H.; Jia, Q.; Qian, Y. Different cellular mechanism of imidacloprid and acetamiprid by a combined targeted lipidomics and metabolomics approach in Neuro-2a cells. Toxicol. Vitr. 2022, 83, 105426. [Google Scholar] [CrossRef]
- Annabi, E.; Salem, I.B.; Abid-Essefi, S. Acetamiprid, a neonicotinoid insecticide, induced cytotoxicity and genotoxicity in PC12 cells. Toxicol. Mech. Methods 2019, 29, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, I.; Ward, L.J.; Vainikka, L.; Sultana, N.; Leanderson, P.; Flodin, U.; Li, W.; Yuan, X. Imidacloprid Induces Lysosomal Dysfunction and Cell Death in Human Astrocytes and Fibroblasts—Environmental Implication of a Clinical Case Report. Cells 2023, 12, 2772. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.I.A.; Ferreira, G.C.H.; Venter, E.A.; Botha, C.J. Cytotoxicity, morphological and ultrastructural effects induced by the neonicotinoid pesticide, imidacloprid, using a rat Leydig cell line (LC-540). Environ. Toxicol. Pharmacol. 2023, 104, 104310. [Google Scholar] [CrossRef] [PubMed]
- Nie, Z.; Niu, Y.; Zhou, W.; Kim, J.; Ock, S.A.; Cui, X. Thiamethoxam induces meiotic arrest and reduces the quality of oocytes in cattle. Toxicol. Vitr. 2019, 61, 104635. [Google Scholar] [CrossRef]
- Kapoor, U.; Srivastava, M.K.; Srivastava, L.P. Toxicological impact of technical imidacloprid on ovarian morphology, hormones and antioxidant enzymes in female rats. Food Chem. Toxicol. 2011, 49, 3086–3089. [Google Scholar] [CrossRef]
- Zuščíková, L.; Bažány, D.; Greifová, H.; Knížatová, N.; Kováčik, A.; Lukáč, N.; Jambor, T. Screening of Toxic Effects of Neonicotinoid Insecticides with a Focus on Acetamiprid: A Review. Toxics 2023, 1, 598. [Google Scholar] [CrossRef]
- Babeľová, J.; Šefčíková, Z.; Čikoš, Š.; Špirková, A.; Kovaříková, V.; Koppel, J.; Makarevich, A.V.; Chrenek, P.; Fabian, D. Exposure to neonicotinoid insecticides induces embryotoxicity in mice and rabbits. Toxicology 2017, 392, 71–80. [Google Scholar] [CrossRef]
- Bal, R.; Naziroglu, M.; Turk, G.; Yilmaz, O.; Kuloglu, T.; Etem, E.; Baydas, G. Insecticide imidacloprid induces morphological and DNA damage through oxidative toxicity on the reproductive organs of developing male rats. Cell. Biochem. Funct. 2012, 30, 492–499. [Google Scholar] [CrossRef]
- Issa, S.Y.; Rahman, S.M.A.; Gaber, Y.M.; Soliman, N.A.H. Toxicological impact of Thiamethoxam on adult male rats: Histopathological, biochemical, and oxidative DNA damage assessment. Toxicol. Rep. 2025, 14, 101983. [Google Scholar] [CrossRef]
- Guimarães, A.; Bizerra, P.F.V.; Miranda, C.A.; Mingatto, F.E. Effects of imidacloprid on viability and increase of reactive oxygen and nitrogen species in HepG2 cell line. Toxicol. Mech. Methods 2021, 32, 204–212. [Google Scholar] [CrossRef]
- Li, X.; Yao, Y.; Wang, J.; Shen, Z.; Jiang, Z.; Xu, S. Eucalyptol relieves imidacloprid-induced autophagy through the miR-451/Cab39/AMPK axis in Ctenopharyngodon idellus kidney cells. Aquat. Toxicol. 2022, 249, 106204. [Google Scholar] [CrossRef]
- Vohra, P.; Khera, K.S.; Sangha, G.K. Physiological, biochemical and histological alterations induced by administration of imidacloprid in female albino rats. Pestic. Biochem. Phys. 2014, 110, 50–56. [Google Scholar] [CrossRef]
- Lohiya, A.; Kumar, V.; Punia, J.S. Imidacloprid induced oxidative stress and histopathological changes in liver of rats. Indian. J. Anim. Res. 2017, 51, 531–536. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Srivastava, M.K.; Kapoor, U.; Srivastava, L.P. A 90 days oral toxicity of imidacloprid in female rats: Morphological, biochemical and histopathological evaluations. Food Chem. Toxicol. 2010, 48, 1185–1190. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Iannaccone, M.; Ianniello, F.; Ferrara, R.; Caprio, E.; Pennacchio, F.; Capparelli, R. The neonicotinoid insecticide Clothianidin adversely affects immune signaling in a human cell line. Sci. Rep. 2017, 7, 13446. [Google Scholar] [CrossRef] [PubMed]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Appl. Biol. Sci. 2013, 110, 18466–18471. [Google Scholar] [CrossRef]
- Cestonaro, L.V.; Crestani, R.P.; Conte, F.M.; Piton, Y.V.; Schmitz, F.S.; Wyse, A.T.S.; Garcia, S.C.; Arbo, M.D. Immunomodulatory effect of imidacloprid on macrophage RAW 264.7 cells. Environ. Toxicol. Pharmacol. 2023, 101, 104190. [Google Scholar] [CrossRef]
- Costa, C.; Silvari, V.; Melchini, A.A. Genotoxicity of imidacloprid in relation to metabolic activation and composition of the commercial product. Mutat. Res. Toxicol. Environ. Mutagen. 2009, 672, 40–44. [Google Scholar] [CrossRef]
- Hernandez, J.; Volland, A.; Leyshon, B.J.; Juda, M.; Ridlon, J.M.; Johnson, R.W.; Steelman, A.J. Effect of imidacloprid ingestion on immune responses to porcine reproductive and respiratory syndrome virus. Sci. Rep. 2018, 8, 11615. [Google Scholar] [CrossRef]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immune response of bumblebees (Apis mellifera L.). J. Insect. Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef]
- Rundlöf, M.; Andersson, G.K.S.; Bommarco, R.; Fries, I.; Hederström, V.; Herbertsson, L.; Jonsson, O.; Klatt, B.K.; Pedersen, T.R.; Yourstone, J.; et al. Seed coating with a neonicotinoid insecticide negatively affects wild bees. Nature 2015, 521, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Wang, X.; Li, J.; Wang, R.; Ren, F.; Pang, G.; Li, Y. Imidacloprid increases intestinal permeability by disrupting tight junctions. Ecotoxicol. Environ. Saf. 2021, 222, 112476. [Google Scholar] [CrossRef]
- Nedzvetsky, V.S.; Masiuk, D.M.; Gasso, V.Y.; Yermolenko, S.V.; Huslystyi, A.O.; Spirina, V.A. Low doses of imidacloprid induce disruption of intercellular adhesion and initiate proinflammatory changes in Caco-2 cells. Regul. Mech. Biosyst. 2021, 12, 430–437. [Google Scholar] [CrossRef]
- Tolga, Ç.; Nilüfer, Ç.; Özgür, V.; Dilek, Y.; Mümün, C. In vitro genotoxicity evaluation of acetamiprid in CaCo-2 cells using the micronucleus, comet and γH2AX foci assays. Pestic. Biochem. Phys. 2012, 104, 212–217. [Google Scholar] [CrossRef]
- Skandrani, D.; Gaubin, Y.; Beau, B. Effect of selected insecticides on growth rate and stress protein expression in cultured human A549 and SH-SY5Y cells. Toxicol. Vitr. 2006, 20, 1378–1386. [Google Scholar] [CrossRef]
- Coppola, L.; Tait, S.; Fabbrizi, E.; Perugini, M.; La Rocca, C. Comparison of the Toxicological Effects of Pesticides in Non-Tumorigenic MCF-12A and Tumorigenic MCF-7 Human Breast Cells. Int. J. Environ. Res. Public Health 2022, 19, 4453. [Google Scholar] [CrossRef]
- Abdel-Halim, K.Y.; Osman, S.R. Cytotoxicity and oxidative stress responses of imidacloprid and glyphosate in human prostate epithelial WPM-Y.1 cell line. J. Toxicol. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kim, Y.; Kim, J.; Yoon, K.S.; Clark, J.; Lee, J.; Park, Y. Imidacloprid, a neonicotinoid insecticide, potentiates adipogenesis in 3T3-L1 adipocytes. J. Agric. Food Chem. 2013, 61, 255–259. [Google Scholar] [CrossRef]
- Mesnage, R.; Biserni, M.; Genkova, D.; Wesolowski, L.; Antoniou, M.N. Evaluation of neonicotinoid insecticides for oestrogenic, thyroidogenic and adipogenic activity reveals imidacloprid causes lipid accumulation. J. Appl. Toxicol. 2018, 38, 1483–1491. [Google Scholar] [CrossRef]
- Sun, Q.; Qi, W.; Yang, J.J.; Yoon, K.S.; Clark, J.M.; Park, Y. Fipronil promotes adipogenesis via AMPKα-mediated pathway in 3T3-L1 adipocytes. Food Chem. Toxicol. 2016, 92, 217–223. [Google Scholar] [CrossRef]
- Sevim, C.; Taghizadehghalehjoughi, A.; Kara, M. In vitro investigation of the effects of imidacloprid on AChE, LDH, and GSH Levels in the L-929 fibroblast cell line. Turk. J. Pharm. Sci. 2020, 17, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Kara, M.; Öztas, E.; Özhan, G. Acetamiprid induced cyto- and genotoxicity in AR42J pancreatic cell line. Turk. J. Pharm. Sci. 2019, 17, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Gomez, S.D.; Bustos, P.S.; Sánchez, V.G.; Ortega, M.G.; Guiñazú, N. Trophoblast toxicity of the neonicotinoid insecticide acetamiprid and an acetamiprid-based formulation. Toxicology 2020, 431, 152363. [Google Scholar] [CrossRef]
- Thompson, D.A.; Kolpin, D.W.; Hladik, M.L.; Lehmler, H.J.; Meppelink, S.M.; Poch, M.C.; Vargo, J.D.; Soupene, V.A.; Irfan, N.M.; Robinson, M.; et al. Prevalence of neonicotinoid insecticides in paired private-well tap water and human urine samples in a region of intense agriculture overlying vulnerable aquifers in eastern Iowa. Chemosphere 2023, 319, 137904. [Google Scholar] [CrossRef]
- Wang, P.; Huang, Y.; Fang, L.; Chen, M. Prenatal and childhood neonicotinoid exposure and neurodevelopment: A study in a young Taiwanese cohort. Sci. Total Environ. 2024, 946, 174232. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Yao, Y.; Huang, Z.; Huang, J.; Xu, J.; Li, X.; Bian, J.; Guo, Z.; Xiao, Q.; Lu, S. Prenatal exposure to neonicotinoids and the associations with neonatal birth outcomes and maternal health in south China. Expo. Health 2025, 17, 97–108. [Google Scholar] [CrossRef]
- Qu, Y.; Li, A.J.; Liu, X.; Lin, S.; Bloom, M.S.; Wang, X.; Li, X.; Wang, H.; Han, F.; Gao, X.; et al. Maternal serum neonicotinoids during early-mid pregnancy and congenital heart diseases in offspring: An exploratory study. Environ. Pollut. 2024, 342, 123046. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Active Ingredient | Example Formulation |
|---|---|
| Imidacloprid | 200SL (soluble concentrate), 70WG (water-dispersible granules), 25WP (Wettable Powder) |
| Thiamethoxam | 25WG, 350FS (flowable concentrate for seed treatment) |
| Acetamiprid | 20SP (soluble powder), 5SL |
| Clothianidin | 600FS |
| Thiacloprid | 480SC (suspension concentrate) |
| Dinotefuran | 20SG (soluble granules), 10WP |
| Nitenpyram | Oral tablets (mainly for veterinary use) |
| Flupyradifurone | 200SL |
| Cycloxaprid | Common in mixture formulations |
| Sulfoxaflor | 240SC, 50WG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Wang, Q. Neonicotinoid-Induced Cytotoxicity: Insights into Cellular Mechanisms and Health Risks. Toxics 2025, 13, 576. https://doi.org/10.3390/toxics13070576
Ma Y, Wang Q. Neonicotinoid-Induced Cytotoxicity: Insights into Cellular Mechanisms and Health Risks. Toxics. 2025; 13(7):576. https://doi.org/10.3390/toxics13070576
Chicago/Turabian StyleMa, Yuqing, and Qiangwei Wang. 2025. "Neonicotinoid-Induced Cytotoxicity: Insights into Cellular Mechanisms and Health Risks" Toxics 13, no. 7: 576. https://doi.org/10.3390/toxics13070576
APA StyleMa, Y., & Wang, Q. (2025). Neonicotinoid-Induced Cytotoxicity: Insights into Cellular Mechanisms and Health Risks. Toxics, 13(7), 576. https://doi.org/10.3390/toxics13070576