
The Effect of the Antidepressant Citalopram on the Bioconcentration and Biomarker Response of Daphnia magna at Environmentally Relevant Concentrations
Abstract
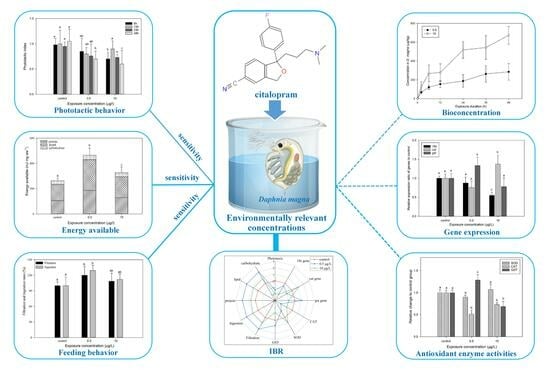
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Experimental Setup
2.3. Chemical and Biochemical Analysis
2.3.1. Extraction and Analysis of CIT
2.3.2. Available Energy Analysis
2.3.3. Antioxidant Enzyme Activity Analysis
2.4. Behavioral Analysis
2.4.1. Phototactic Behavior
2.4.2. Feeding Behavior
2.5. Gene Transcription Analysis
2.6. Integrated Biomarker Response Analysis
2.7. Data Analysis
3. Results and Discussion
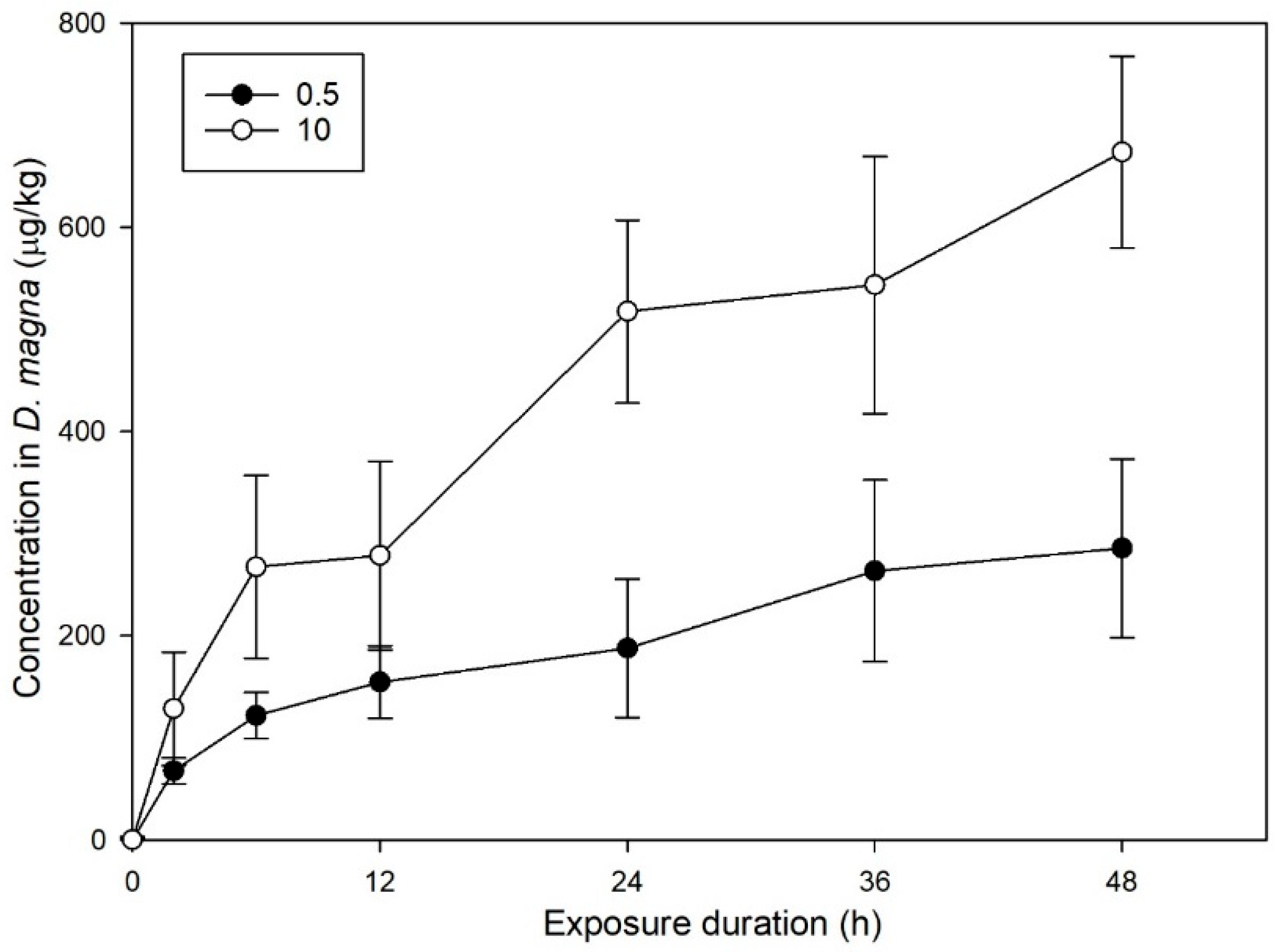
3.1. Effect of CIT on the Bioconcentration of D. magna
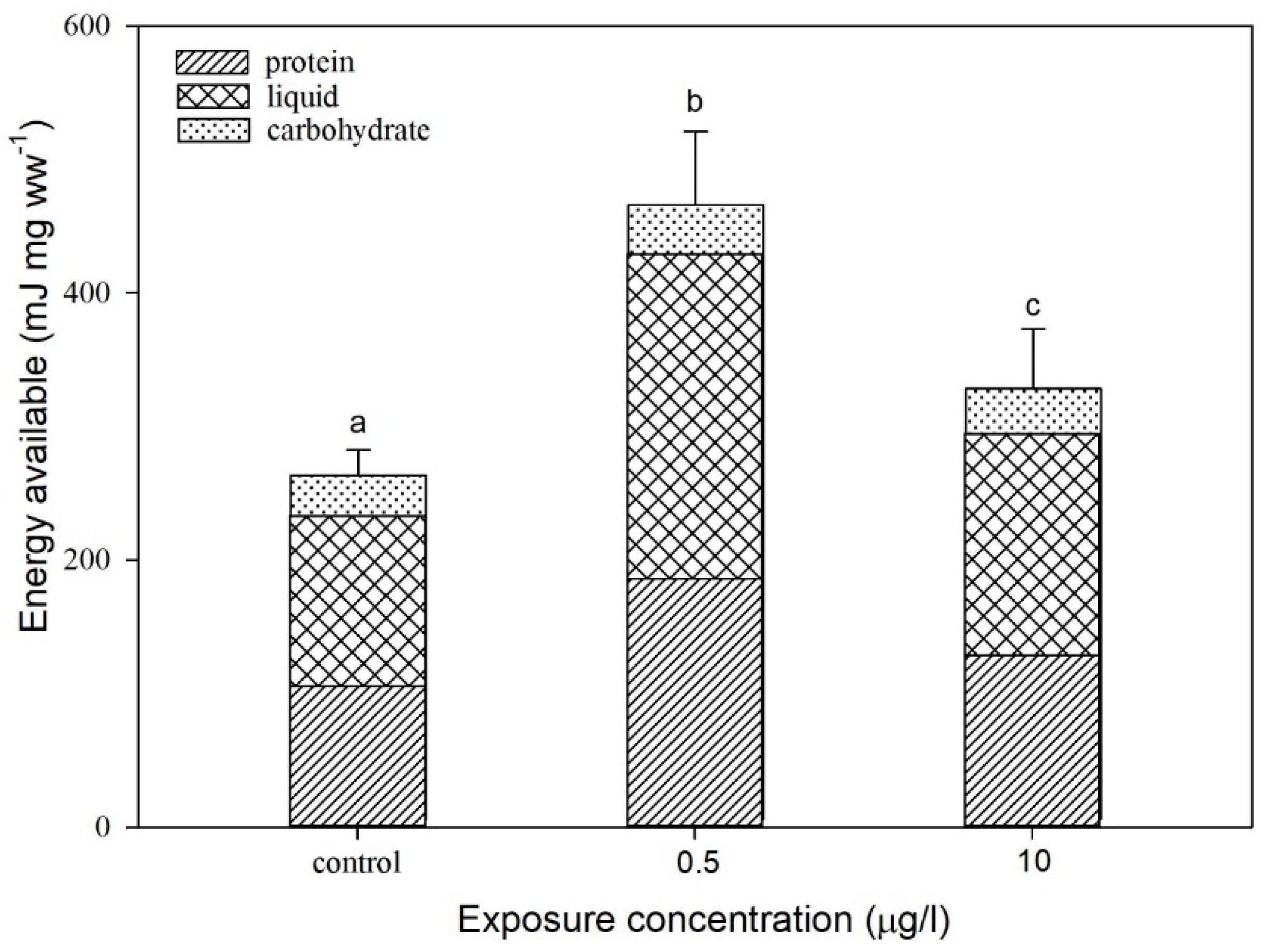
3.2. Effect of CIT on the Energy Available to D. magna
3.3. Effect of CIT on the Behavior Response of D. magna
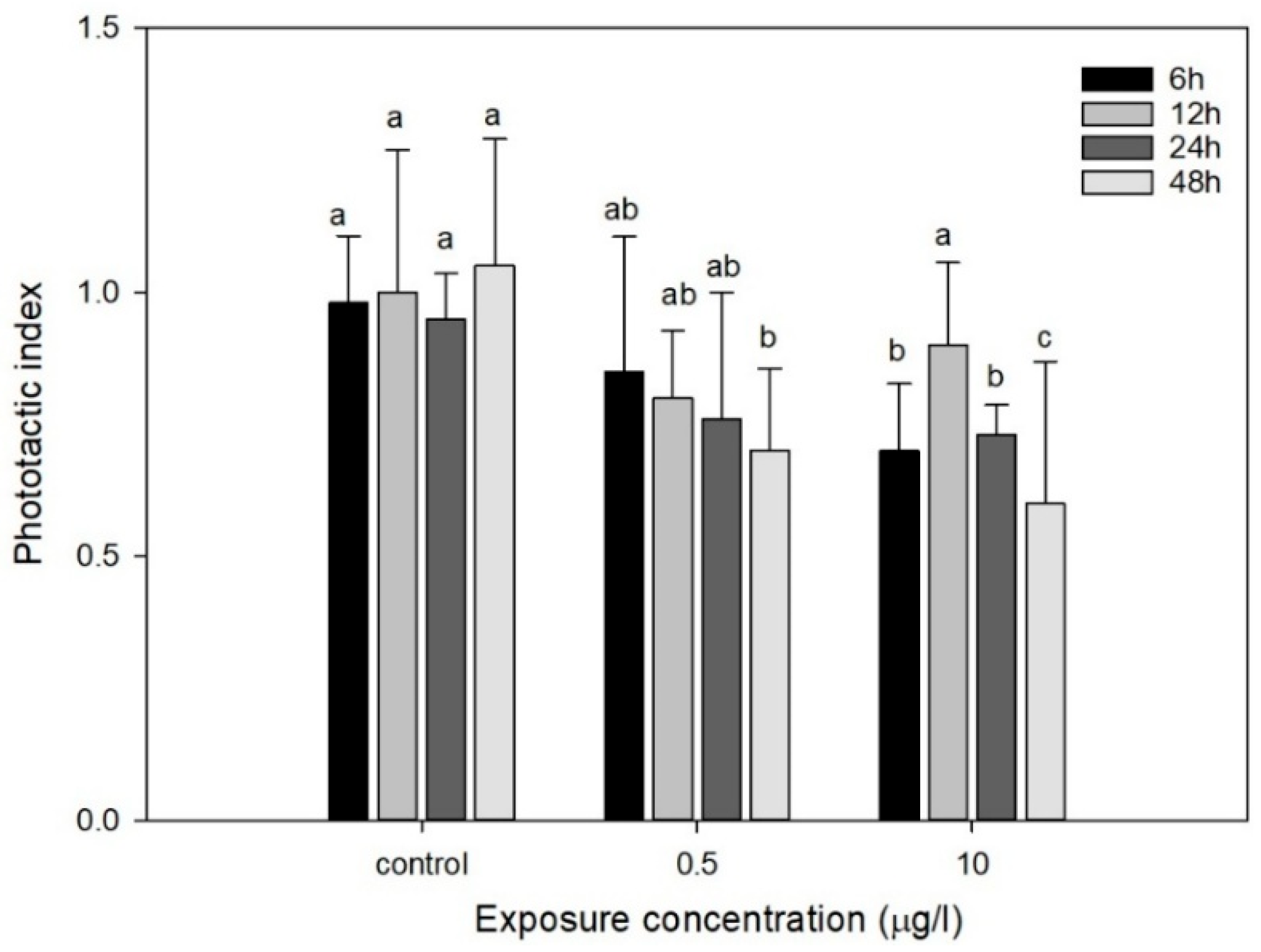
3.3.1. Phototactic Behavior
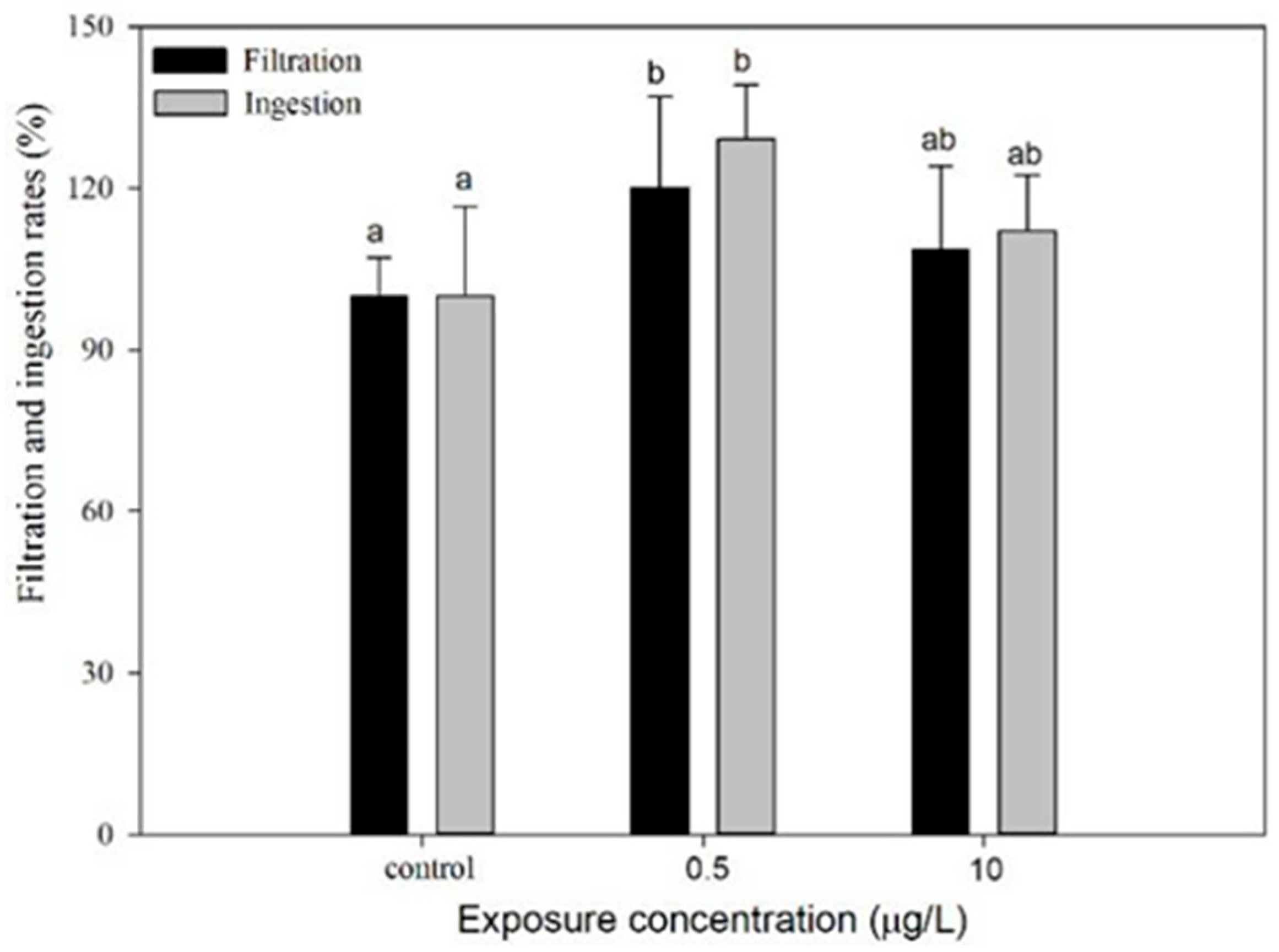
3.3.2. Feeding Behavior
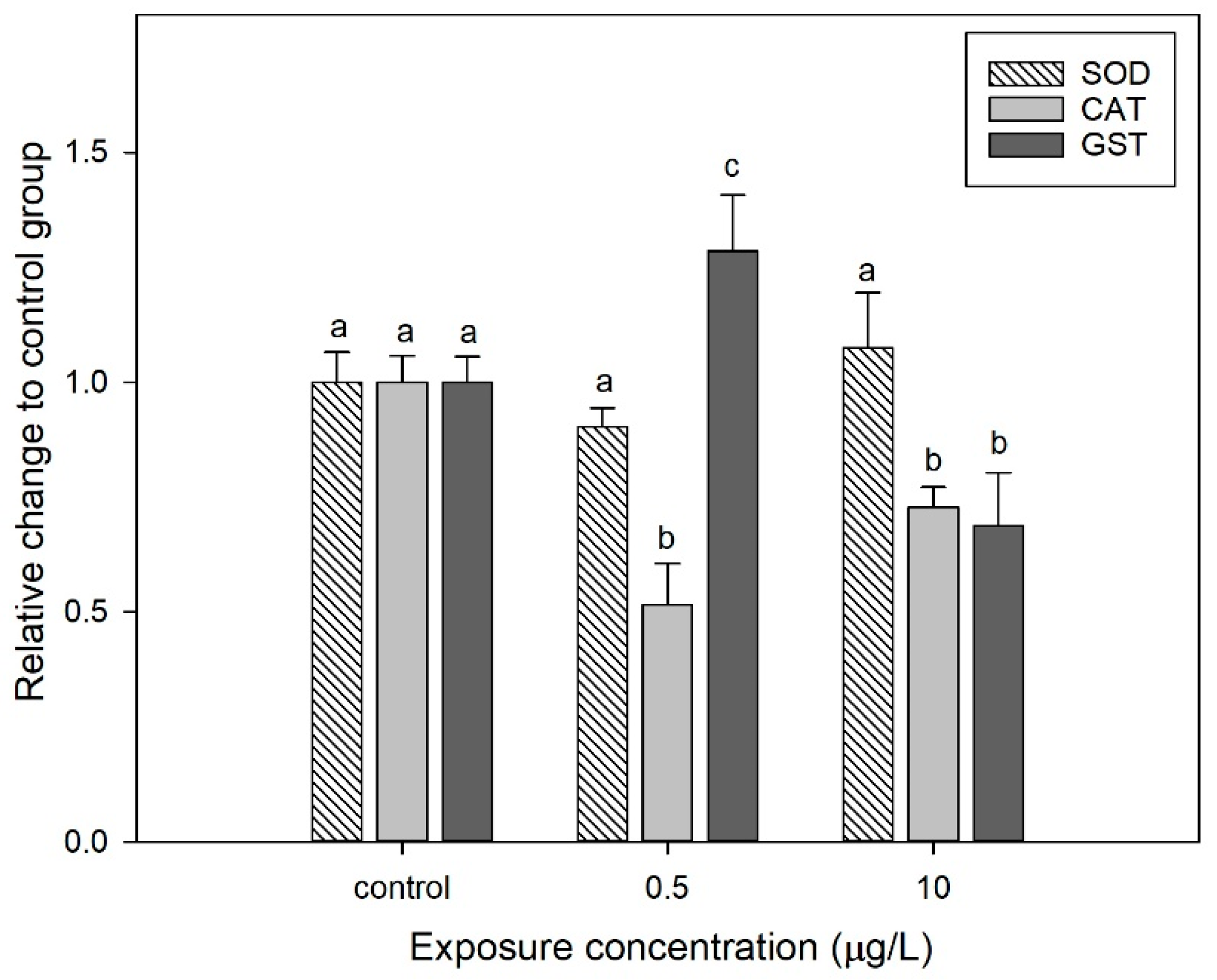
3.4. Effect of CIT on the Antioxidant Enzyme Activities of D. magna
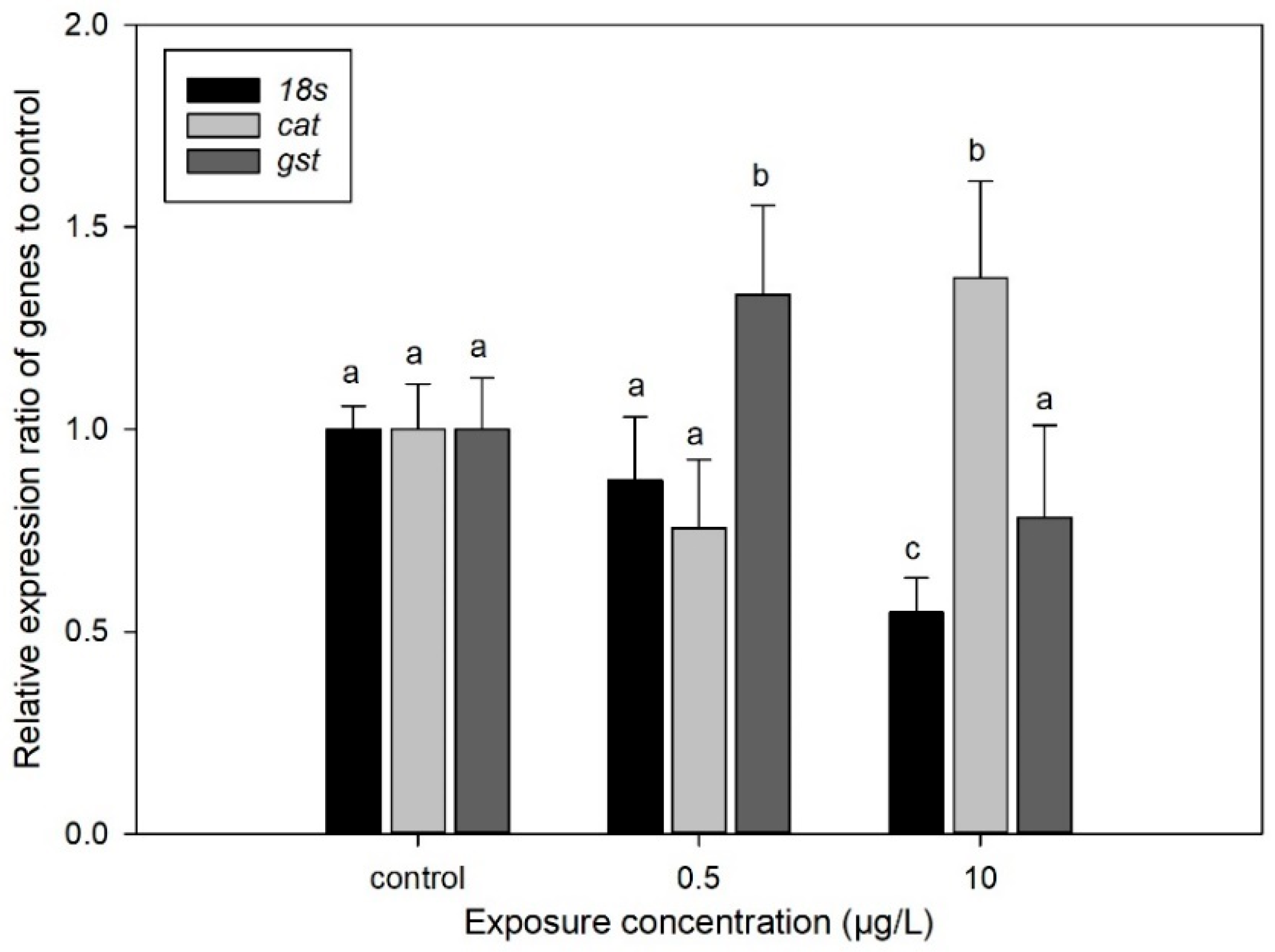
3.5. Effect of CIT on the Gene Expression of D. magna
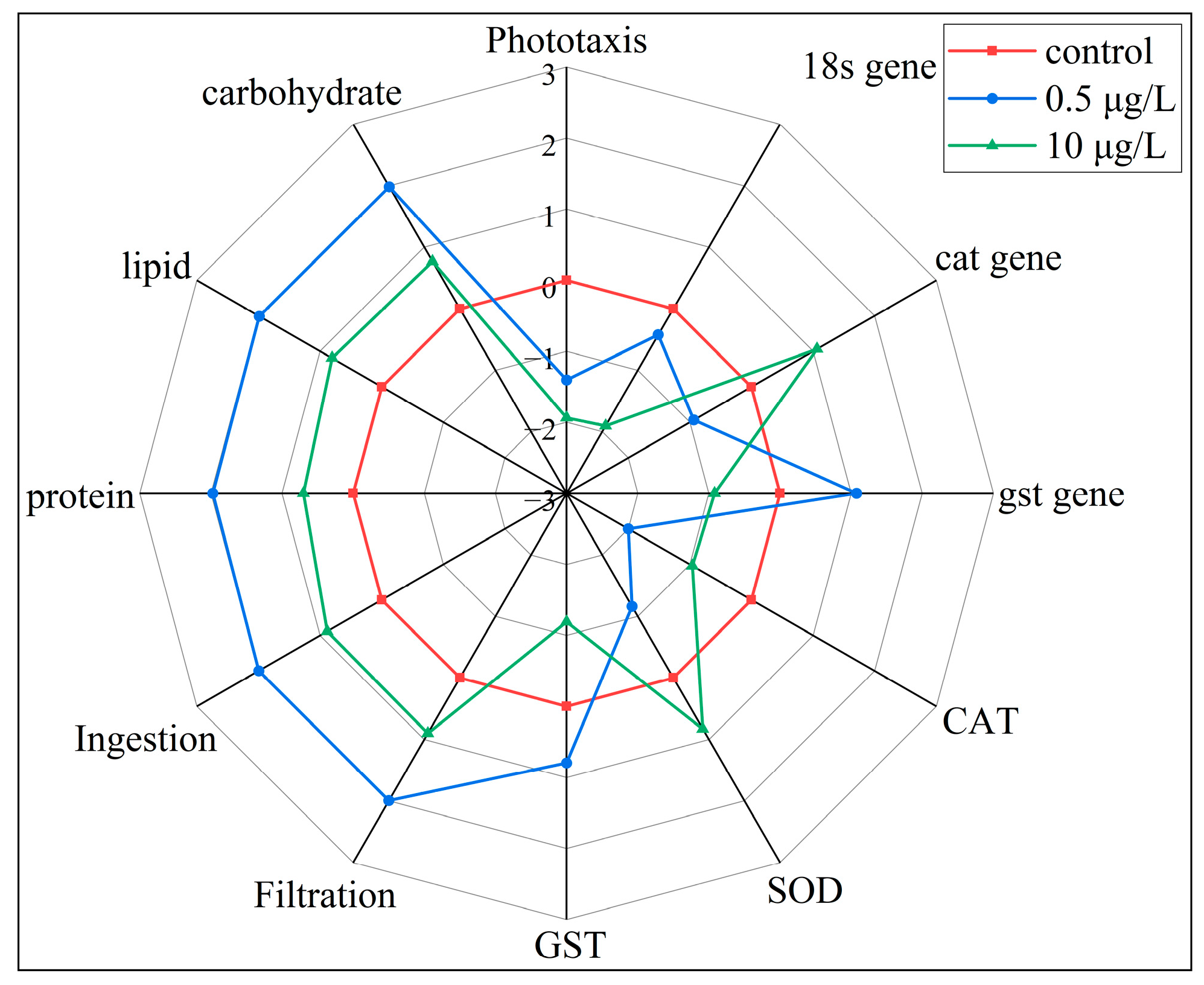
3.6. Integrated Biomarker Response
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyttel, J. Pharmacological characterization of selective serotonin reuptake inhibitors (SSRIs). Int. Clin. Psychopharmacol. 1994, 9, 19–26. [Google Scholar] [CrossRef]
- Zhu, Y.; Yu, X.; He, X.; Xue, L.; Chen, X.; Liu, R.; Yan, Z. Occurrence and risk assessment of antidepressant drugs in Nanjing water bodies. Environ. Monit. Manag. Technol. 2019, 31, 7–11. [Google Scholar]
- Jiang, M.; Wang, L.; Sheng, H. Mitochondria in depression: The dysfunction of mitochondrial energy metabolism and quality control systems. CNS Neurosci. Ther. 2024, 30, e14576. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.S.; Alonazi, M.; Al-Daihan, S.; El-Ansary, A. Prenatal SSRI exposure increases the risk of autism in rodents via aggravated oxidative stress and neurochemical changes in the brain. Metabolites 2023, 13, 310. [Google Scholar] [CrossRef]
- Sepúlveda-Lizcano, L.; Arenas-Villamizar, V.V.; Jaimes-Duarte, E.B.; García-Pacheco, H.; Paredes, C.S.; Bermúdez, V.; Rivera-Porras, D. Metabolic adverse effects of psychotropic drug therapy: A systematic review. Eur. J. Investig. Health Psychol. Educ. 2023, 13, 1505–1520. [Google Scholar] [CrossRef] [PubMed]
- Sengar, A.; Vijayanandan, A. Human health and ecological risk assessment of 98 pharmaceuticals and personal care products (PPCPs) detected in Indian surface and wastewaters. Sci. Total Environ. 2022, 807, 150677. [Google Scholar] [CrossRef]
- Schultz, M.M.; Furlong, E.T.; Kolpin, D.W.; Werner, S.L.; Schoenfuss, H.L.; Barber, L.B.; Blazer, V.S.; Norris, D.O.; Vajda, A.M. Antidepressant pharmaceuticals in two US effluent-impacted streams: Occurrence and fate in water and sediment, and selective uptake in fish neural tissue. Environ. Sci. Technol. 2010, 44, 1918–1925. [Google Scholar] [CrossRef]
- Mole, R.A.; Brooks, B.W. Global scanning of selective serotonin reuptake inhibitors: Occurrence, wastewater treatment and hazards in aquatic systems. Environ. Pollut. 2019, 250, 1019–1031. [Google Scholar] [CrossRef]
- Meredith-Williams, M.; Carter, L.J.; Fussell, R.; Raffaelli, D.; Ashauer, R.; Boxall, A.B.A. Uptake and depuration of pharmaceuticals in aquatic invertebrates. Environ. Pollut. 2012, 165, 250–258. [Google Scholar] [CrossRef]
- Passignat, C.; Flayac, J.; Lerebourg, R.; Minguez, L. Differential bioconcentration and sensitivity of Dreissena polymorpha and Dreissena rostriformis bugensis to the antidepressant sertraline. J. Hazard. Mater. 2025, 482, 136628. [Google Scholar] [CrossRef]
- Affinito, F.; Kordas, R.L.; Matias, M.G.; Pawar, S. Metabolic plasticity drives mismatches in physiological traits between prey and predator. Commun. Biol. 2024, 7, 653. [Google Scholar] [CrossRef] [PubMed]
- Rivetti, C.; Campos, B.; Barata, C. Low environmental levels of neuro-active pharmaceuticals alter phototactic behaviour and reproduction in Daphnia magna. Aquat. Toxicol. 2016, 170, 289–296. [Google Scholar] [CrossRef]
- Gao, J.; Zhao, S.; Li, F.; Yang, X.; Zhang, J.; Xiong, A.; Li, F. Effects of microplastics on the feeding behavior and antioxidant defense system of Daphnia magna. Res. Environ. Sci. 2021, 34, 1205–1212. [Google Scholar] [CrossRef]
- Brand, J.A.; Michelangeli, M.; Shry, S.J.; Moore, E.R.; Bose, A.P.; Cerveny, D.; Martin, J.M.; Hellström, G.; McCallum, E.S.; Holmgren, A.; et al. Pharmaceutical pollution influences river-to-sea migration in Atlantic salmon (Salmo salar). Science 2025, 388, 217–222. [Google Scholar] [CrossRef]
- Yang, D.; Yu, W.; Qu, J.; Shen, Y.; Yu, J.; Meng, R.; Tao, Z.; Chen, J.; Du, W.; Sun, H.Z.; et al. Environmentally relevant exposure to cotinine induces neurobehavioral toxicity in zebrafish (Danio rerio): A study using neurobehavioral and metabolomic approaches. Environ. Pollut. 2024, 348, 123826. [Google Scholar] [CrossRef] [PubMed]
- Sarasamma, S.; Audira, G.; Juniardi, S.; Sampurna, B.P.; Liang, S.T.; Hao, E.; Lai, Y.H.; Hsiao, C.D. Zinc Chloride Exposure Inhibits Brain Acetylcholine Levels, Produces Neurotoxic Signatures, and Diminishes Memory and Motor Activities in Adult Zebrafish. Int. J. Mol. Sci. 2018, 19, 3195. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Lu, G.; Yan, Z.; Liu, J.; Dong, H. Influence of suspended sediment characteristics on the bioaccumulation and biological effects of citalopram in Daphnia magna. Chemosphere 2018, 207, 293–302. [Google Scholar] [CrossRef]
- Hong, X.; Chen, R.; Zhang, L.; Yan, L.; Xin, J.; Li, J.; Zha, J. Long-Term Exposure to SSRI Citalopram Induces Neurotoxic Effects in Zebrafish. Environ. Sci. Technol. 2022, 56, 12380–12390. [Google Scholar] [CrossRef]
- Žabka, D.; Vojs Staňová, A.; Horáková, I.; Butor Škulcová, A.; Grabic, R.; Špalková, V.; Gál, M.; Mackuľak, T. Bioaccumulation as a method of removing psychoactive compounds from wastewater using aquatic plants. J. Chromatogr. B 2023, 1223, 123717. [Google Scholar] [CrossRef]
- Hoang, T.C.; Klaine, S.J. Influence of organism age on metal toxicity to Daphnia magna. Environ. Toxicol. Chem. 2007, 26, 1198–1204. [Google Scholar] [CrossRef]
- Yang, H.; Zhang, Z.; Liu, J.; Liu, Z.; Zhou, Z.; Feng, Q. Bioavailability of citalopram to Daphnia magna in the presence of suspended sediments with various properties. Mar. Pollut. Bull. 2022, 175, 113352. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zou, H.; Liu, Q.; Zhang, S.; Razanajatovo, R.M. Bioconcentration of the antidepressant fluoxetine and its effects on the physiological and biochemical status in Daphnia magna. Ecotoxicol. Environ. Saf. 2017, 142, 102–109. [Google Scholar] [CrossRef]
- Yang, H.; Lu, G.; Yan, Z.; Liu, J.; Ma, B.; Dong, H. Biological effects of citalopram in a suspended sediment-water system on Daphnia magna. Environ. Sci. Pollut. Res. Int. 2017, 24, 21180–21190. [Google Scholar] [CrossRef]
- He, C.; Zhao, N.; Hu, L.; Tang, T.; Yang, Y.; Nie, X. Assessment of ecotoxicity effects of aspirin on non-target organism (Daphnia magna) via analysis of the responses of oxidative stress, DNA methylation-related genes expressions and life traits changes. Ecotoxicology 2023, 32, 137–149. [Google Scholar] [CrossRef]
- Bozdag, D.; van Voorthuizen, J.; Korpel, N.; Lentz, S.; Gurer-Orhan, H.; Kamstra, J.H. Dysregulation of adipogenesis and disrupted lipid metabolism by the antidepressants citalopram and sertraline. Toxicol. Appl. Pharmacol. 2024, 486, 116937. [Google Scholar] [CrossRef]
- Gómez-Canela, C.; Esquius, F.; Barata, C. The role of serotonergic signaling on phototactic and locomotor behavior in Daphnia magna. Sci. Total Environ. 2023, 856, 159042. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Xu, D.; Li, C.; Wei, S.; Guo, R.; Li, Y.; Chen, J.; Liu, Y. Combined toxicity and toxicity persistence of antidepressants citalopram and mirtazapine to zooplankton Daphnia magna. Environ. Sci. Pollut. Res. 2022, 29, 66100–66108. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Mahongnao, S.; Gupta, A.; Nanda, S. Health risk assessment for potentially toxic elements accumulation in Amaranthaceae family cultivars and their correlation with antioxidants and antinutrients. Arch. Environ. Contam. Toxicol. 2024, 87, 187–207. [Google Scholar] [CrossRef]
- Lei, P.; Yang, M.; Zhan, J.; Wang, D.; Wu, M. Pollution and ecotoxicity of antidepressant drugs in aquatic environments. J. Chem. Eng. Chin. Univ. 2017, 31, 1009–1027. [Google Scholar]
- Liu, Y.N. Toxic Effects of Lead on Growth, Reproductive Behavior, and Nutrients in Daphnia magna. Master’s Thesis, Shanxi University, Taiyuan, China, 2020. [Google Scholar]
- Yang, H.; Kong, L.; Chen, Z.; Wu, J. Effect of functional groups of polystyrene nanoplastics on the neurodevelopmental toxicity of acrylamide in the early life stage of zebrafish. Aquat. Toxicol. 2025, 278, 107177. [Google Scholar] [CrossRef]
- Sanchez, W.; Burgeot, T.; Porcher, J.M. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. Res. 2013, 20, 2721–2725. [Google Scholar] [CrossRef]
- Risso, B.; Miglioli, A.; Balbi, T.; Dumollard, R.; Canesi, L. Molecular basis for the effects of SSRIs in non-target aquatic invertebrates: A case study with Mytilus galloprovincialis early larvae. Aquat. Toxicol. 2025, 282, 107306. [Google Scholar] [CrossRef]
- Xie, Z.; Lu, G.; Yan, Z.; Liu, J.; Wang, P.; Wang, Y. Bioaccumulation and trophic transfer of pharmaceuticals in food webs from a large freshwater lake. Environ. Pollut. 2017, 222, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Wang, W.X. Accumulation kinetics and gut microenvironment responses to environmentally relevant doses of micro/nanoplastics by zooplankton Daphnia magna. Environ. Sci. Technol. 2023, 57, 5611–5620. [Google Scholar] [CrossRef] [PubMed]
- Smirnov, S.I.; Zdorovennov, R.E.; Efremova, T.V.; Palshin, N.I.; Smirnovsky, A.A.; Bogdanov, S.R.; Terzhevik, A.Y. Parameters of water column stability in a small polymictic lake in years of different weather conditions. Water Resour. 2024, 51, 299–313. [Google Scholar] [CrossRef]
- Yan, Z.; Zhao, H.; Zhu, P.; Wang, Y.; Hou, J.; Lu, G.; He, C. Polystyrene microplastics alter the trophic transfer and biotoxicity of fluoxetine in an aquatic food chain. J. Hazard. Mater. 2024, 470, 134179. [Google Scholar] [CrossRef] [PubMed]
- Guler, Y.; Ford, A.T. Antidepressants make amphipods see the light. Aquat. Toxicol. 2010, 99, 397–404. [Google Scholar] [CrossRef]
- De Coen, W.M.; Janssen, C.R.; Segner, H. The use of biomarkers in Daphnia magna toxicity testing V. In vivo alterations in the carbohydrate metabolism of Daphnia magna exposed to sublethal concentrations of mercury and lindane. Ecotoxicol. Environ. Saf. 2001, 48, 223–234. [Google Scholar] [CrossRef]
- Du, L.; Ma, D.; Jing, X.; Wang, L.; Wang, X. Effects of trichlorfon on the growth and reproductive capacity of Moina macrocopa. Acta Hydrobiol. Sin. 2014, 38, 786–790. [Google Scholar] [CrossRef]
- Han, C.; Kottapalli, A.; Boyapati, K.; Chan, S.; Jeong, Y.J. Acidity enhances the ability of 5-Aminoimidazole-4-carboxamide ribonucleotide to increase respiration and lipid metabolism in Daphnia magna. J. Korean Chem. Soc. 2019, 63, 253–259. [Google Scholar] [CrossRef]
- Yang, H.; Lu, G.; Yan, Z.; Liu, J. Influence of suspended sediment on the bioavailability of benzophenone-3: Focus on accumulation and multi-biological effects in Daphnia magna. Chemosphere 2021, 275, 129974. [Google Scholar] [CrossRef]
- Lopes, D.G.; Duarte, I.A.; Antunes, M.; Fonseca, V.F. Effects of antidepressants in the reproduction of aquatic organisms: A meta-analysis. Aquat. Toxicol. 2020, 227, 105569. [Google Scholar] [CrossRef] [PubMed]
- Kooijman, S.A.L.M.; Baas, J.; Bontje, D.; Broerse, M.; van Gestel, C.A.M. Ecotoxicological applications of dynamic energy budget theory. Ecotoxicol. Model. 2009, 237–259. [Google Scholar] [CrossRef]
- Hargreaves, I.P.; Al Shahrani, M.; Wainwright, L.; O’Brien, P.J. Drug-induced mitochondrial toxicity. Drug Saf. 2016, 39, 661–674. [Google Scholar] [CrossRef]
- Vandenberg, L.N.; Colborn, T.; Hayes, T.B.; Heindel, J.J.; Jacobs, D.R., Jr.; Lee, D.-H.; Shioda, T.; Soto, A.M.; vom Saal, F.S.; Welshons, W.V.; et al. Hormones and endocrine-disrupting chemicals: Low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012, 33, 378–455. [Google Scholar] [CrossRef] [PubMed]
- Im, H.; Achar, J.C.; Shim, T.; Jung, J. Elevated temperature alleviates benzophenone-3 toxicity in Daphnia magna. Aquat. Toxicol. 2022, 242, 106047. [Google Scholar] [CrossRef]
- Fuertes, I.; Pina, B.; Barata, C. Changes in lipid pro files in Daphnia magna individuals exposed to low environmental levels of neuroactive pharmaceuticals. Sci. Total Environ. 2020, 733, 139029. [Google Scholar] [CrossRef]
- Mohanthi, S.; Sutha, J.; Gayathri, M.; Ramesh, M. Evaluation of the citalopram toxicity on early development of zebrafish: Morphological, physiological and biochemical responses. Environ. Pollut. 2024, 357, 124399. [Google Scholar] [CrossRef]
- Sanpradit, P.; Peerakietkhajorn, S. Disturbances in growth, oxidative stress, energy reserves and the expressions of related genes in Daphnia magna after exposure to ZnO under thermal stress. Sci. Total Environ. 2023, 869, 161682. [Google Scholar] [CrossRef]
- Nkoom, M.; Lu, G.; Liu, J.; Yang, H.; Dong, H. Bioconcentration of the antiepileptic drug carbamazepine and its physiological and biochemical effects on Daphnia magna. Ecotoxicol. Environ. Saf. 2019, 172, 11–18. [Google Scholar] [CrossRef]
- Liu, Q.; Deng, Z.; Chen, H.F.; Kim, M.-S.; Kim, D.-H.; Gu, L.; Lee, J.-S.; Yang, Z. Changes in induced-antipredation defense traits and transcriptome regulations of Daphnia magna in response to 5-HT1A receptor antagonist. Environ. Sci. Technol. 2024, 58, 7577–7587. [Google Scholar] [CrossRef] [PubMed]
- Campos, B.; Rivetti, C.; Kress, T.; Barata, C.; Dircksen, H. Depressing antidepressant: Fluoxetine affects serotonin neurons causing adverse reproductive responses in Daphnia magna. Environ. Sci. Technol. 2016, 50, 6000–6007. [Google Scholar] [CrossRef] [PubMed]
- Long, E.R.; Hong, C.B.; Severn, C.G. Relationships between acute sediment toxicity in laboratory tests and abundance and diversity of benthic infauna in marine sediments: A review. Environ. Toxicol. Chem. 2001, 20, 46–60. [Google Scholar] [CrossRef]
- Politi, E.; Cutler, M.E.J.; Carvalho, L.; Rowan, J.S. A global typological approach to classify lakes based on their eutrophication risk. Aquat. Sci. 2024, 86, 52. [Google Scholar] [CrossRef]
- Chen, C.; Cui, D.; Li, J.; Ren, C.; Yang, D.; Xiang, P.; Liu, J. Organophosphorus Flame Retardant TPP-Induced Human Corneal Epithelial Cell Apoptosis through Caspase-Dependent Mitochondrial Pathway. Int. J. Mol. Sci. 2024, 25, 4155. [Google Scholar] [CrossRef]
- Osvik, R.D.; Ingebrigtsen, R.A.; Norrbin, M.F.; Andersen, J.H.; Eilertsen, H.C.; Hansen, E.H. Adding Zooplankton to the OSMAC Toolkit: Effect of Grazing Stress on the Metabolic Profile and Bioactivity of a Diatom. Mar. Drugs 2021, 19, 87. [Google Scholar] [CrossRef]
- Duan, S.; Fu, Y.; Dong, S.; Ma, Y.; Meng, H.; Guo, R.; Chen, J.; Liu, Y.; Li, Y. Psychoactive drugs citalopram and mirtazapine caused oxidative stress and damage of feeding behavior in Daphnia magna. Ecotoxicol. Environ. Saf. 2021, 230, 113147. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.D.; Hayes, J.D.; Dinkova-Kostova, A.T. Nrf2/Keap1 signaling in xenobiotic detoxification. Free Radic. Biol. Med. 2020, 157, 125–135. [Google Scholar]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic or-ganisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Oliveira, M.; Costa, P.M.; Almeida, J.R. Epigenetic silencing of detoxification genes under chemical stress. Environ. Pollut. 2020, 265, 114890. [Google Scholar]
- Venkatesan, R.; Ji, E.; Kim, S.Y. Phytochemicals that regulate neurodegenerative disease by targeting neurotrophins: A comprehensive review. BioMed Res. Int. 2015, 1, 814068. [Google Scholar] [CrossRef] [PubMed]
- Duarte, I.A.; Reis-Santos, P.; Fran, A.S.; Cabral HFonseca, V.F. Biomarker responses to environmental contamination in estuaries: A comparative multi-taxa approach. Aquat. Toxicol. 2017, 183, 31–34. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Xia, X.; Wang, J.; Zhu, L.; Wang, J.; Zhang, F.; Ahmad, Z. Chronic toxicological effects of carbamazepine on Daphnia magna Straus: Effects on reproduction traits, body length, and intrinsic growth. Bull. Environ. Contam. Toxicol. 2019, 103, 723–728. [Google Scholar] [CrossRef]
- Cao, X.Q.; Wang, X.; Liu, B.; He, S.W.; Cao, Z.H.; Xing, S.Y.; Li, P.; Li, Z.H. Evaluation of physiological stress of grass carp chronically exposed to enrofloxacin based on IBR index. Water Air Soil Pollut. 2023, 234, 548. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Sequence | Reserve Sequence |
|---|---|---|
| β-actin | ACGATGATGTTGCGGCTTTG | CCGACAATGGAGGGGAAGAC |
| 18s | CCTGAGAAACGGCTACCACATC | CTCGGAAGAGTCCCGTATCGT |
| gst | TCTATTATCCCATCATGTTCAGCG | CCAGCAGCATACTTGTTCTGACC |
| cat | ATCGCCTTGGAACAAACTACCT | AGCTGTTCGGGAAGTAATTTGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Tan, J.; Jiang, H.; Xing, H.; Zhang, J.; Kong, D.; Chen, Z.; Kong, L. The Effect of the Antidepressant Citalopram on the Bioconcentration and Biomarker Response of Daphnia magna at Environmentally Relevant Concentrations. Toxics 2025, 13, 532. https://doi.org/10.3390/toxics13070532
Yang H, Tan J, Jiang H, Xing H, Zhang J, Kong D, Chen Z, Kong L. The Effect of the Antidepressant Citalopram on the Bioconcentration and Biomarker Response of Daphnia magna at Environmentally Relevant Concentrations. Toxics. 2025; 13(7):532. https://doi.org/10.3390/toxics13070532
Chicago/Turabian StyleYang, Haohan, Jiacheng Tan, Hanyu Jiang, Hao Xing, Jingnan Zhang, Dexin Kong, Zhuoyu Chen, and Linghui Kong. 2025. "The Effect of the Antidepressant Citalopram on the Bioconcentration and Biomarker Response of Daphnia magna at Environmentally Relevant Concentrations" Toxics 13, no. 7: 532. https://doi.org/10.3390/toxics13070532
APA StyleYang, H., Tan, J., Jiang, H., Xing, H., Zhang, J., Kong, D., Chen, Z., & Kong, L. (2025). The Effect of the Antidepressant Citalopram on the Bioconcentration and Biomarker Response of Daphnia magna at Environmentally Relevant Concentrations. Toxics, 13(7), 532. https://doi.org/10.3390/toxics13070532