Fusarium Mycotoxins in Two Hulless Oat and Barley Cultivars Used for Food Purposes

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Field Trials
2.2. Preparation of Samples and Analysis of Mycotoxins by UPLC/MS/MS
2.3. Data Analyses
3. Results
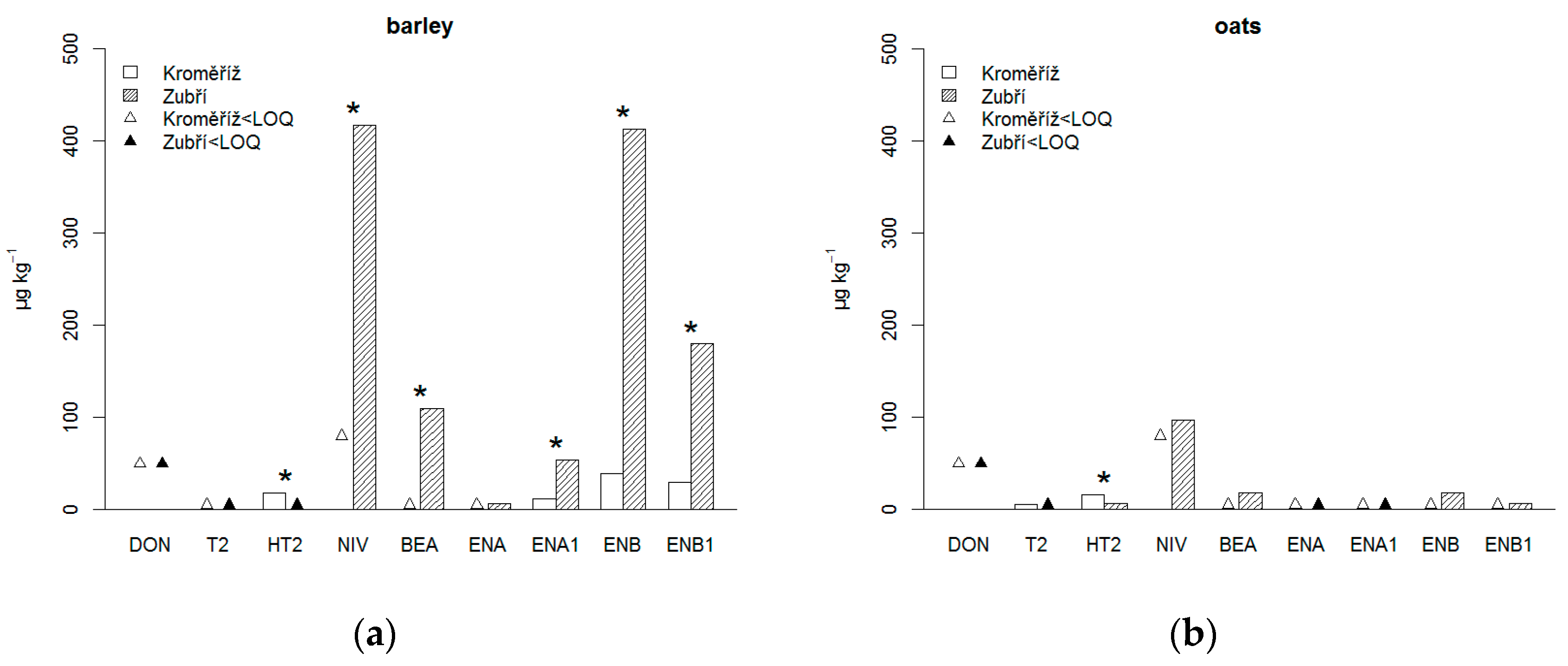
3.1. Content of Mycotoxins in Barley
3.2. Content of Mycotoxins in Oats
3.3. Comparison of Mycotoxin Occurrence in Barley and Oats
3.4. Relationship between Individual Mycotoxins
4. Discussion
4.1. Deoxynivalenol (DON) and Zearalenone (ZEA)
4.2. Fumonisins and T-2 and HT-2 Toxins
4.3. Nivalenol (NIV)
4.4. Enniatins (ENs) and Beauvericine (BEA)
4.5. The Main Differences between Hulless Barley and Oats
4.6. Factors Influencing Mycotoxin Content of Hulless Barley and Oats
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zohary, D.; Hopf, M. Domestication of Plants in the Old World: The Origin and Spread of Cultivated Plants in West Asia, Europe, and the Nile Valley, 3rd ed.; Oxford University Press: Oxford, UK, 2000; pp. 59–69. [Google Scholar]
- European Union. Commission Regulation (EC) No. 432/2012 of 16 May 2012 establishing a list of permitted health claims made on foods, other than those referring to the reduction of disease risk and to children’s development and health. Off. J. Eur. Union 2012, L 136, 1–40. [Google Scholar]
- Schöneberg, T.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Vogelgsang, S. Occurrence of Fusarium species and mycotoxins in Swiss oats—Impact of cropping factors. Eur. J. Agron. 2018, 92, 123–132. [Google Scholar]
- Clear, R.M.; Patrick, S.K.; Platford, R.G.; Desjardins, M. Occurrence and distribution of Fusarium species in barley and oat seed from Manitoba in 1993 and 1994. Can. Plant Pathol. 1996, 18, 409–414. [Google Scholar]
- Ma, H.M.; Ge, H.; Zhang, X.; Lu, W.; Yu, D.; Chen, H.; Chen, J. Resistance to Fusarium head blight and deoxynivalenol accumulation in Chinese barley. J. Phytopathol. 2009, 157, 166–171. [Google Scholar]
- Janssen, E.M.; Liu, C.; Van der Fels-Klerx, H.J. Fusarium infection and trichothecenes in barley and its comparison with wheat. World Mycotoxin J. 2018, 11, 33–46. [Google Scholar]
- Tekauz, A.; McCallum, B.; Ames, N.; Fetch, J.M. Fusarium head blight of oat—Current status in western Canada. Can. Plant Pathol. 2004, 479, 473–479. [Google Scholar]
- Gruber-Dorninger, C.; Novak, B.; Nagl, V.; Berthiller, F. Emerging Mycotoxins: Beyond Traditionally Determined Food Contaminants. J. Agric. Food Chem. 2017, 65, 7052–7070. [Google Scholar]
- European Union. Commission Regulation (EC) No 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, L 364, 5–24. [Google Scholar]
- European Union. Commission Recommendation of 27 March 2013 on the presence of T-2 and HT-2 toxin in cereals and cereal products (2013/165/EU). Off. J. Eur. Union 2013, L 91, 12–15. [Google Scholar]
- Jestoi, M. Emerging Fusarium—Mycotoxins Fusaproliferin, Beauvericin, Enniatins, and Moniliformin—A Review. Crit. Rev. Food Sci. Nutr. 2008, 48, 21–49. [Google Scholar]
- EFSA. Scientific Opinion on the risks to human and animal health related to the presence of beauvericin and enniatins in food and feed. EFSA J. 2014, 12, 3802. [Google Scholar]
- Scudamore, K.A.; Baillie, H.; Patel, S.; Edwards, S.G. Occurrence and fate of Fusarium mycotoxins during commercial processing of oats in the UK. Food Addit. Contam. Part A 2007, 24, 1374–1385. [Google Scholar]
- Berger, G.; Green, A.; Khatibi, P.; Brooks, W.; Rosso, L.; Liu, S.; Chao, S.; Griffey, C.; Schmale, D., III. Characterization of Fusarium head blight resistance and deoxynivalenol accumulation in hulled and hulless winter barley. Plant Dis. 2014, 98, 599–606. [Google Scholar]
- He, X.; Osman, M.; Helm, J.; Capettini, F.; Singh, P.K. Evaluation of Canadian barley breeding lines for Fusarium head blight resistance. Can. J. Plant Sci. 2015, 95, 923–929. [Google Scholar]
- Legzdina, L.; Buerstmayr, H. Comparison of infection with Fusarium head blight and accumulation of mycotoxins in grain of hulless and covered barley. J. Cereal Sci. 2004, 40, 61–67. [Google Scholar]
- Malachova, A.; Cerkal, R.; Ehrenbergerova, J.; Dzuman, Z.; Vaculova, K.; Hajslova, J. Fusarium mycotoxins in various barley cultivars and their transfer into malt. J. Sci. Food Agric. 2010, 90, 2495–2505. [Google Scholar]
- Yan, W.; Fregeau-Reid, J.; Rioux, S.; Pageau, D.; Xue, A.; Martin, R.; Fedak, G.; Lajeunesse, J.; Savard, M. Response of oat genotypes to Fusarium Head Blight in Eastern Canada. Crop Sci. 2010, 50, 134–142. [Google Scholar]
- Šliková, S.; Šrobárová, A.; Šudyová, V.; Polišenská, I.; Gregová, E.; Miháli, D. Response of oat cultivars to Fusarium infection with a view to their suitability for food use. Biologia 2010, 65, 609–614. [Google Scholar]
- Gagkaeva, T.; Gavrilova, O.P.; Yli-Mattila, T.; Loskutov, I.G. Sources of resistance to Fusarium head blight in VIR Oat Collection. Euphytica 2013, 195, 355–364. [Google Scholar]
- Loskutov, I.G.; Blinova, E.; Gavrilova, O.P.; Gagkaeva, T. The valuable characteristics and resistance to Fusarium disease of oat genotypes. Russ. J. Genet. 2017, 7, 290–298. [Google Scholar]
- Bolechová, M.; Benešová, K.; Běláková, S.; Čáslavský, J.; Pospíchalová, M.; Mikulíková, R. Determination of seventeen mycotoxins in barley and malt in the Czech Republic. Food Control 2015, 47, 108–113. [Google Scholar]
- Campbell, H.; Choo, T.M.; Vigier, B.; Underhill, L. Mycotoxins in barley and oat samples from eastern Canada. Can. J. Plant Sci. 2000, 80, 977–980. [Google Scholar]
- Edwards, S.G. Investigation of Fusarium mycotoxins in UK barley and oat production. Project Report No.415; AHDB Cereals & Oilseeds: Stoneleigh, UK, 2007. [Google Scholar]
- Schöneberg, T.; Martin, C.; Wettstein, F.E.; Bucheli, T.D.; Mascher, F.; Bertossa, M.; Musa, T.; Keller, B.; Vogelgsang, S. Fusarium and mycotoxin spectra in Swiss barley are affected by various cropping techniques. Food Addit. Contam. Part A 2016, 33, 1608–1619. [Google Scholar]
- Gil-Serna, J.; Mateo, E.M.; Gonzalez-Jaen, M.T.; Jimenez, M.; Vazquez, C.; Patino, B. Contamination of barley seeds with Fusarium species and their toxins in Spain: An integrated approach. Food Addit. Contam. Part A 2013, 30, 372–380. [Google Scholar]
- Hietaniemi, V.; Ramo, S.; Yli-Mattila, T.; Jestoi, M.; Peltonen, S.; Kartio, M.; Sievilainen, E.; Koivisto, T.; Parikka, P. Updated survey of Fusarium species and toxins in Finnish cereal grains. Food Addit. Contam. Part A 2016, 33, 831–848. [Google Scholar]
- Beccari, G.; Prodi, A.; Tini, F.; Bonciarelli, U.; Onofri, A.; Oueslati, S.; Limayma, M.; Covarelli, L. Changes in the Fusarium Head Blight Complex of Malting Barley in a Three-Year Field Experiment in Italy. Toxins 2017, 9, 120. [Google Scholar]
- Palumbo, R.; Crisci, A.; Venâncio, A.; Cortiñas Abrahantes, J.; Dorne, J.-L.; Battilani, P.; Toscano, P. Occurrence and Co-Occurrence of Mycotoxins in Cereal-Based Feed and Food. Microorganisms 2020, 8, 74. [Google Scholar]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional barley. Food Addit. Contam. Part A 2009, 26, 1185–1190. [Google Scholar]
- Edwards, S.G. Fusarium mycotoxin content of UK organic and conventional oats. Food Addit. Contam. Part A 2009, 26, 1063–1069. [Google Scholar]
- Fredlund, E.; Gidlund, A.; Sulyok, M.; Börjesson, T.; Krska, R.; Olsen, M.; Lindblad, M. Deoxynivalenol and other selected Fusarium toxins in Swedish oats–occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 276–283. [Google Scholar]
- Hofgaard, I.S.; Aamot, H.U.; Torp, T.; Jestoi, M.; Lattanzio, V.M.T.; Klemsdal, S.S.; Waalwijk, C.; Van der Lee, T.; Brodal, G. Associations between Fusarium species and mycotoxins in oats and spring wheat from farmers’ fields in Norway over a six-year period. World Mycotoxin J. 2016, 9, 365–378. [Google Scholar]
- Nielsen, L.K.; Cook, D.J.; Edwards, S.G.; Ray, R.V. The prevalence and impact of Fusarium head blight pathogens and mycotoxins on malting barley quality in UK. Int. J. Food Microbiol. 2014, 179, 38–49. [Google Scholar]
- Ryu, J.; Ohtsubo, K.; Izumiyama, N.; Nakamura, K.; Tanaka, T.; Yamamura, H.; Ueno, Y. The acute and chronic toxicities of nivalenol in mice. Fundam. Appl. Toxicol. 1988, 11, 38–47. [Google Scholar]
- Svingen, T.; Lund Hansen, N.; Taxvig, C.; Vinggaard, A.M.; Jensen, U.; Have Rasmussen, P. Enniatin B and beauvericin are common in Danish cereals and show high hepatotoxicity on a high-content imaging platform. Environ. Toxicol. 2017, 32, 1658–1664. [Google Scholar]
- Bryla, M.; Waskiewicz, A.; Podolska, G.; Szymczyk, K.; Jedrzejczak, R.; Damaziak, K.; Sulek, A. Occurrence of 26 Mycotoxins in the Grain of Cereals Cultivated in Poland. Toxins 2016, 8, 160. [Google Scholar]
- Jestoi, M.; Rokka, M.; Yli-Mattila, T.; Parikka, P.; Rizzo, A.; Peltonen, K. Presence and concentrations of the Fusarium-related mycotoxins beauvericin, enniatins and moniliformin in finnish grain samples. Food Addit. Contam. 2004, 21, 794–802. [Google Scholar]
- Kosiak, B.; Torp, M.; Skjerve, E.; Andersen, B. Alternaria and Fusarium in Norwegian grains of reduced quality—A matched pair sample study. Int. J. Food Microbiol. 2004, 93, 51–62. [Google Scholar]
- Uhlig, S.; Torp, M.; Heier, B.T. Beauvericin and enniatins A, A1, B and B1 in Norwegian grain: A survey. Food Chem. 2006, 94, 193–201. [Google Scholar]
- Santini, A.; Meca, G.; Uhlig, S.; Ritieni, A. Fusaproliferin, beauvericin and enniatins: Occurrence in food—A review. World Mycotoxin J. 2012, 5, 71–81. [Google Scholar]
- Covarelli, L.; Beccari, G.; Prodi, A.; Generotti, S.; Etruschi, F.; Meca, G.; Juan, C.; Manes, J. Biosynthesis of beauvericin and enniatins in vitro by wheat Fusarium species and natural grain contamination in an area of central Italy. Food Microbiol. 2015, 46, 618–626. [Google Scholar]
- Doohan, F.M.; Brennan, J.; Cooke, B.M. Influence of climatic factors on Fusarium species pathogenic to cereals. Eur. J. Plant Pathol. 2003, 109, 755–768. [Google Scholar]
- Prosperini, A.; Berrada, H.; Ruiz, M.J.; Caloni, F.; Coccini, T.; Spicer, L.J.; Perego, M.C.; Lafranconi, A. A Review of the Mycotoxin Enniatin B. Front. Public Health 2017, 5, 304. [Google Scholar]
- Logrieco, A.F.; Moretti, A. Between emerging and historical problems: An overview of the main toxigenic fungi and mycotoxin concerns in Europe. In Mycotoxins: Detection Methods, Management, Public Health and Agricultural Trade; Leslie, J., Bandyopadhyay, R., Visconti, A., Eds.; CAB International: Wallingford, UK, 2008; pp. 139–153. [Google Scholar]
- Langseth, W.; Elen, O. Differences between barley, oats and wheat in the occurrence of deoxynivalenol and other trichothecenes in Norwegian grain. J. Phytopathol. 1996, 144, 113–118. [Google Scholar]
- Bernhoft, A.; Torp, M.; Clasen, P.-E.; Loes, A.-K.; Kristoffersen, A.B. Influence of agronomic and climatic factors on Fusarium infestation and mycotoxin contamination of cereals in Norway. Food Addit. Contam. Part A 2012, 29, 1129–1140. [Google Scholar]
- Kokkonen, M.; Ojala, L.; Parikka, P.; Jestoi, M. Mycotoxin production of selected Fusarium species at different culture conditions. Int. J. Food Microbiol. 2010, 143, 17–25. [Google Scholar]
- Lindblad, M.; Gidlund, A.; Sulyok, M.; Borjesson, T.; Krska, R.; Olsen, M.; Fredlund, E. Deoxynivalenol and other selected Fusarium toxins in Swedish wheat—Occurrence and correlation to specific Fusarium species. Int. J. Food Microbiol. 2013, 167, 284–291. [Google Scholar]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics, and Biology; APS Press: St. Paul, MN, USA, 2006; p. 268. [Google Scholar]
- Chelkowski, J.; Ritieni, A.; Wisniewska, H.; Mulé, G.; Logrieco, A. Occurrence of toxic hexadepsipeptides in preharvest maize ear rot infected by Fusarium poae in Poland. J. Phytopathol. 2007, 155, 8–12. [Google Scholar]
- De Nijs, M.; Rombouts, F.; Notermans, S. Fusarium molds and their mycotoxins. J. Food Saf. 1996, 16, 15–58. [Google Scholar]

{kind=link}
| Locality | Kroměříž | Zubří | ||||
|---|---|---|---|---|---|---|
| Latitude, longitude | 49°17′ N, 17°22′ E | 49°28′ N, 18°5′ E | ||||
| Altitude (m a.s.l.) | 235 | 345 | ||||
| Average annual temperature 1 | 9.2 °C | 7.5 °C | ||||
| Average total annual precipitation 1 | 576 mm | 865 mm | ||||
| Soil type | Luvic Chernozem | Gleyic Fluvisol | ||||
| Soil textural class | Silty clay loam | Sandy loam | ||||
| Year | 2015 | 2016 | 2017 | 2015 | 2016 | 2017 |
| Mean temperature 2 | 13.9 °C | 13.9 °C | 14.4 °C | 13.0 °C | 13.2 °C | 13.4 °C |
| Rainfall 2 | 184 mm | 309 mm | 248 mm | 213 mm | 468 mm | 383 mm |
| Date of sowing | 24-March | 30-March | 28-March | 14-Apr | 5-Apr | 30-March |
| Date of harvest-barley | 30-July | 27-July | 22-July | 3-August | 9-August | 24-July |
| Date of harvest-oats | 5-August | 30-July | 23-July | 3-August | 9-August | 1-August |
| p Values for Concentrations | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Crop | Factor | n | DON | T2 | HT2 | NIV | BEA | ENA | ENA1 | ENB | ENB1 |
| barley | Year a | 3 | 0.607 | 0.368 | 0.607 | 0.761 | 0.032 | 0.368 | 0.050 | 0.018 | 0.039 |
| Location b | 1 | 0.180 | n.a. | 0.028 | 0.028 | 0.028 | 0.109 | 0.043 | 0.028 | 0.028 | |
| Cultivar b | 1 | 0.655 | n.a. | 0.285 | 1.000 | 0.715 | 0.180 | 0.500 | 0.753 | 0.753 | |
| oats | Year a | 2 | n.a. | 0.202 | 0.607 | 0.135 | 0.223 | n.a. | n.a. | 0.135 | 0.135 |
| Location b | 1 | n.a. | 0.285 | 0.043 | 0.180 | 0.068 | n.a. | n.a. | 0.068 | 0.180 | |
| Cultivar b | 1 | n.a. | 1.000 | 0.893 | n.a. | 0.655 | n.a. | n.a. | 0.180 | n.a. | |
| Mean Concentrations ± Standard Deviation and p Values | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Crop | Location | n | DON | T2 | HT2 | NIV | BEA | ENA | ENA1 | ENB | ENB1 |
| barley | 12 | 31 ± 13 | 3 ± 1 | 10 ± 9 | 239 ± 218 | 57 ± 118 | 5 ± 4 | 32 ± 32 | 226 ± 69 | 105 ± 105 | |
| oats | 12 | 25 ± 0 | 4 ± 3 | 11 ± 9 | 68 ± 77 | 10 ± 12 | 3 ± 0 | 3 ± 0 | 10 ± 5 | 4 ± 4 | |
| 0.166 | 0.113 | 0.832 | 0.022 | 0.069 | 0.079 | 0.000 | 0.000 | 0.000 | |||
| barley | KM | 6 | 36 ± 18 | 3 ± 1 | 18 ± 6 | 61 ± 51 | 4 ± 2 | 3 ± 0 | 11 ± 9 | 39 ± 31 | 30 ± 25 |
| oats | 6 | 25 ± 0 | 5 ± 3 | 16 ± 9 | 40 ± 0 | 3 ± 0 | 3 ± 0 | 3 ± 0 | 3 ± 0 | 3 ± 0 | |
| 0.378 | 0.262 | 0.810 | 0.689 | 0.378 | 0.936 | 0.066 | 0.005 | 0.020 | |||
| barley | ZB | 6 | 25 ± 0 | 3 ± 0 | 3 ± 0 | 417 ± 161 | 110 ± 154 | 7 ± 5 | 54 ± 33 | 413 ± 201 | 180 ± 101 |
| oats | 6 | 25 ± 0 | 3 ± 2 | 6 ± 6 | 96 ± 106 | 17 ± 14 | 3 ± 0 | 3 ± 0 | 18 ± 21 | 6 ± 5 | |
| 1.000 | 0.689 | 0.378 | 0.008 | 0.013 | 0.173 | 0.020 | 0.005 | 0.008 | |||
| DON | T2 | HT2 | NIV | BEA | ENA | ENA1 | ENB | |
|---|---|---|---|---|---|---|---|---|
| T2 | −0.153 | |||||||
| HT2 | 0.237 | 0.219 | ||||||
| NIV | 0.009 | −0.380 | −0.517 | |||||
| BEA | −0.122 | −0.290 | −0.495 | 0.833 | ||||
| ENA | −0.114 | −0.192 | −0.372 | 0.572 | 0.485 | |||
| ENA1 | 0.227 | −0.380 | −0.177 | 0.611 | 0.581 | 0.659 | ||
| ENB | 0.115 | −0.333 | −0.321 | 0.592 | 0.718 | 0.530 | 0.826 | |
| ENB1 | 0.208 | −0.286 | −0.206 | 0.603 | 0.695 | 0.604 | 0.914 | 0.948 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polišenská, I.; Jirsa, O.; Vaculová, K.; Pospíchalová, M.; Wawroszova, S.; Frydrych, J. Fusarium Mycotoxins in Two Hulless Oat and Barley Cultivars Used for Food Purposes. Foods 2020, 9, 1037. https://doi.org/10.3390/foods9081037
Polišenská I, Jirsa O, Vaculová K, Pospíchalová M, Wawroszova S, Frydrych J. Fusarium Mycotoxins in Two Hulless Oat and Barley Cultivars Used for Food Purposes. Foods. 2020; 9(8):1037. https://doi.org/10.3390/foods9081037
Chicago/Turabian StylePolišenská, Ivana, Ondřej Jirsa, Kateřina Vaculová, Markéta Pospíchalová, Simona Wawroszova, and Jan Frydrych. 2020. "Fusarium Mycotoxins in Two Hulless Oat and Barley Cultivars Used for Food Purposes" Foods 9, no. 8: 1037. https://doi.org/10.3390/foods9081037
APA StylePolišenská, I., Jirsa, O., Vaculová, K., Pospíchalová, M., Wawroszova, S., & Frydrych, J. (2020). Fusarium Mycotoxins in Two Hulless Oat and Barley Cultivars Used for Food Purposes. Foods, 9(8), 1037. https://doi.org/10.3390/foods9081037