Untargeted Metabolomics of Fermented Rice Using UHPLC Q-TOF MS/MS Reveals an Abundance of Potential Antihypertensive Compounds

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Cultures
2.2. Rice Fermentation
2.3. Determination of Angiotensin 1-Converting Enzyme Inhibitory Ability
2.4. Metabolomics Analysis
2.5. Data Analysis
2.6. Statistical Analysis
3. Results and Discussion
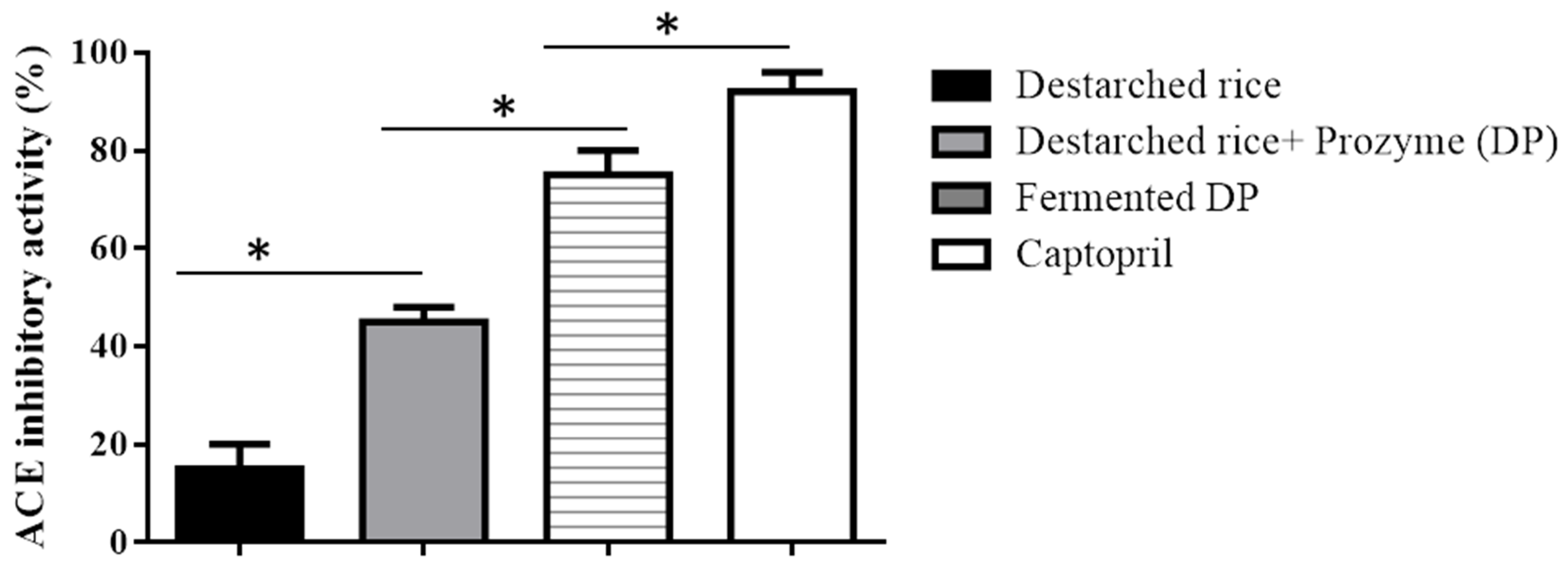
3.1. ACE Inhibitory Activity of Rice Samples
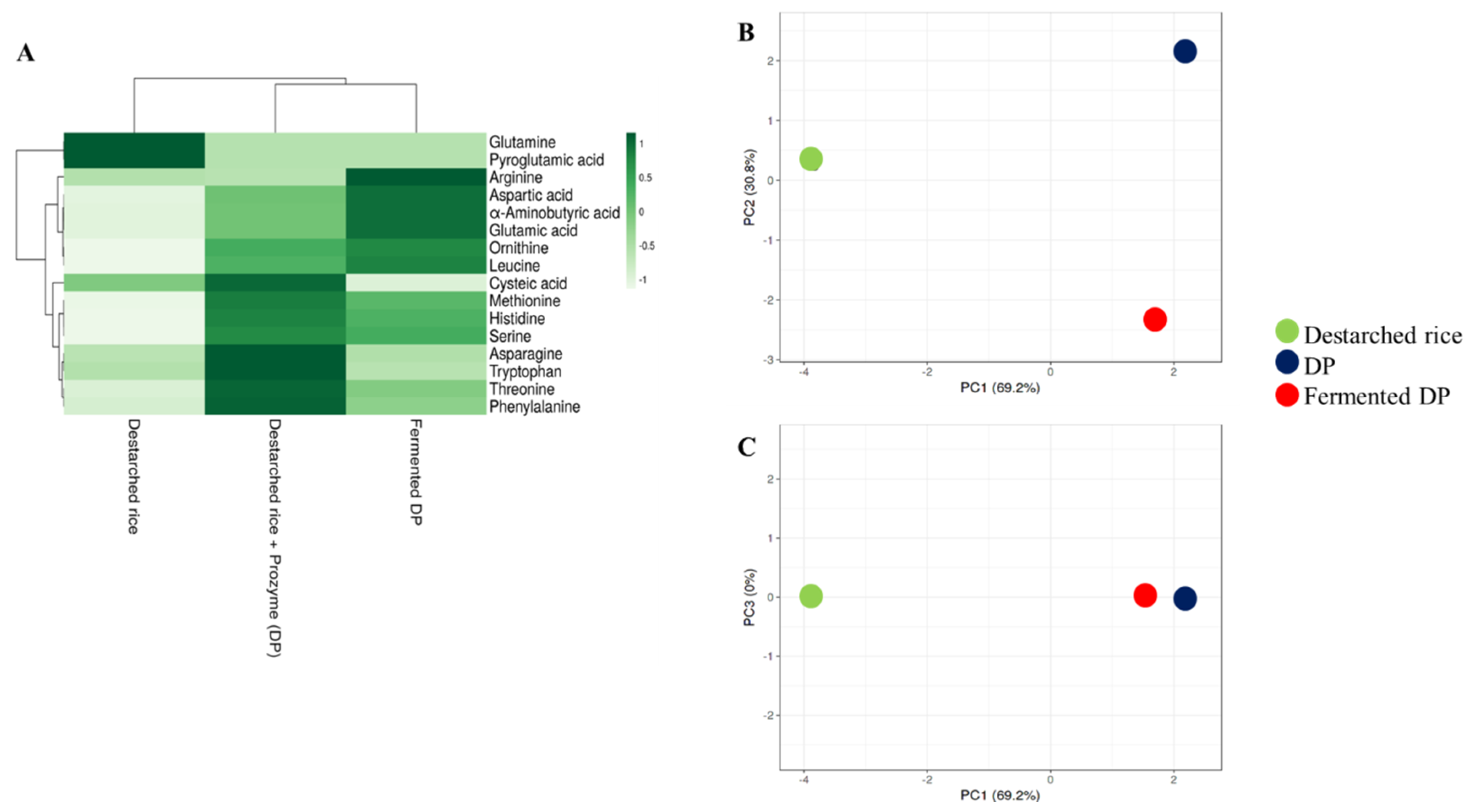
3.2. Amino Acid Levels in the Rice Samples
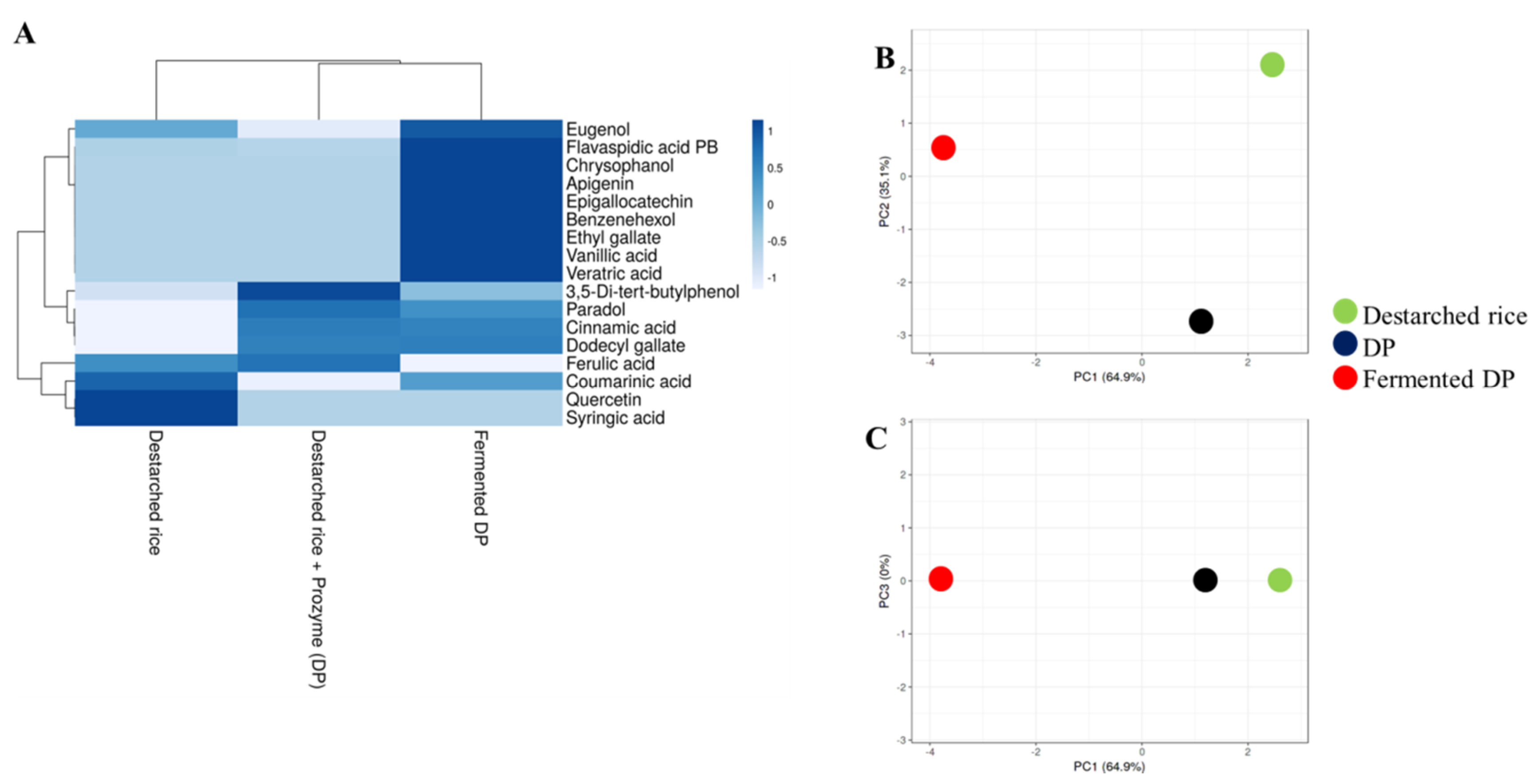
3.3. E. faecium EBD1fermentation Increases the Amount of Phenolic Compounds in Rice
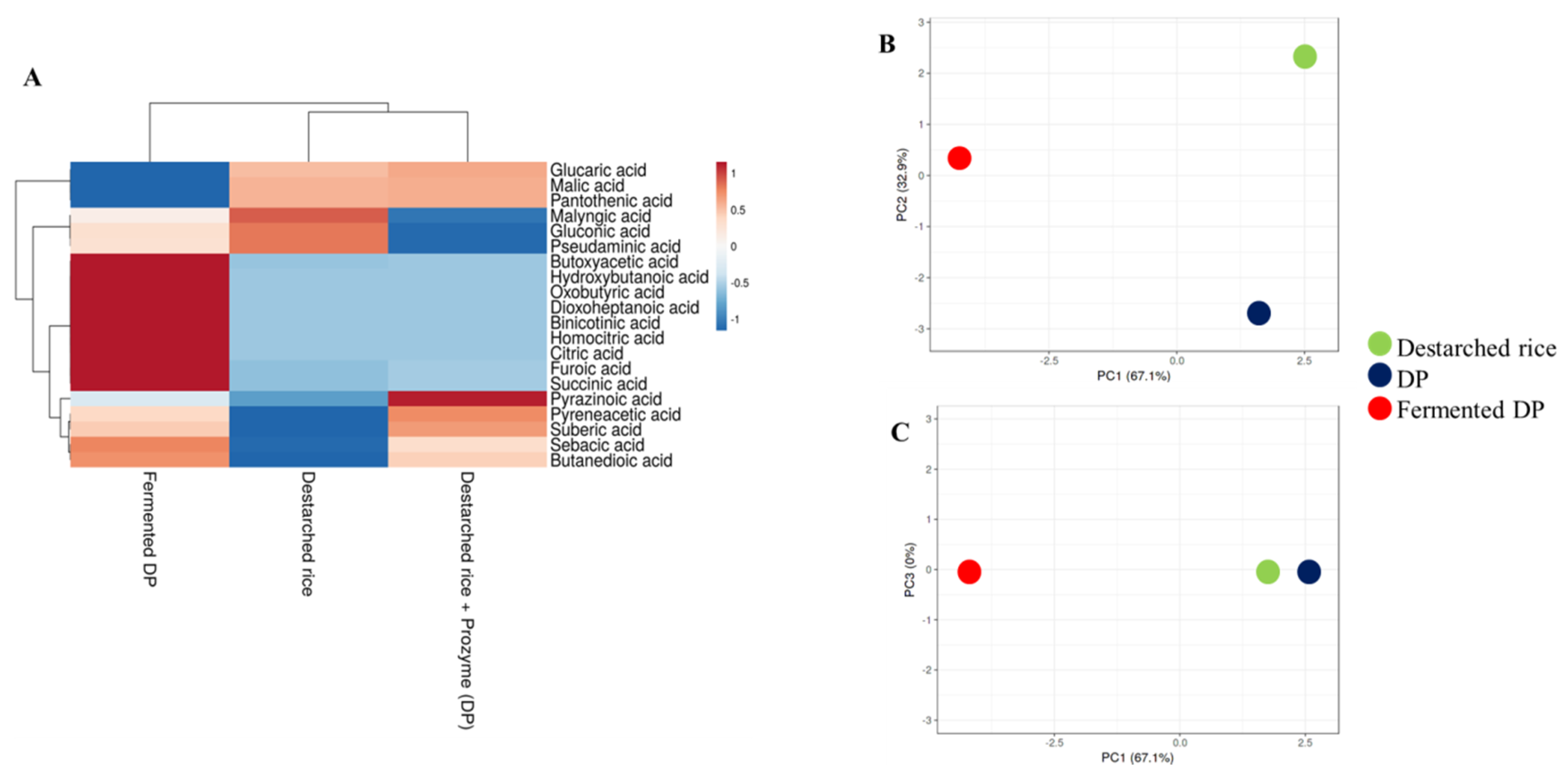
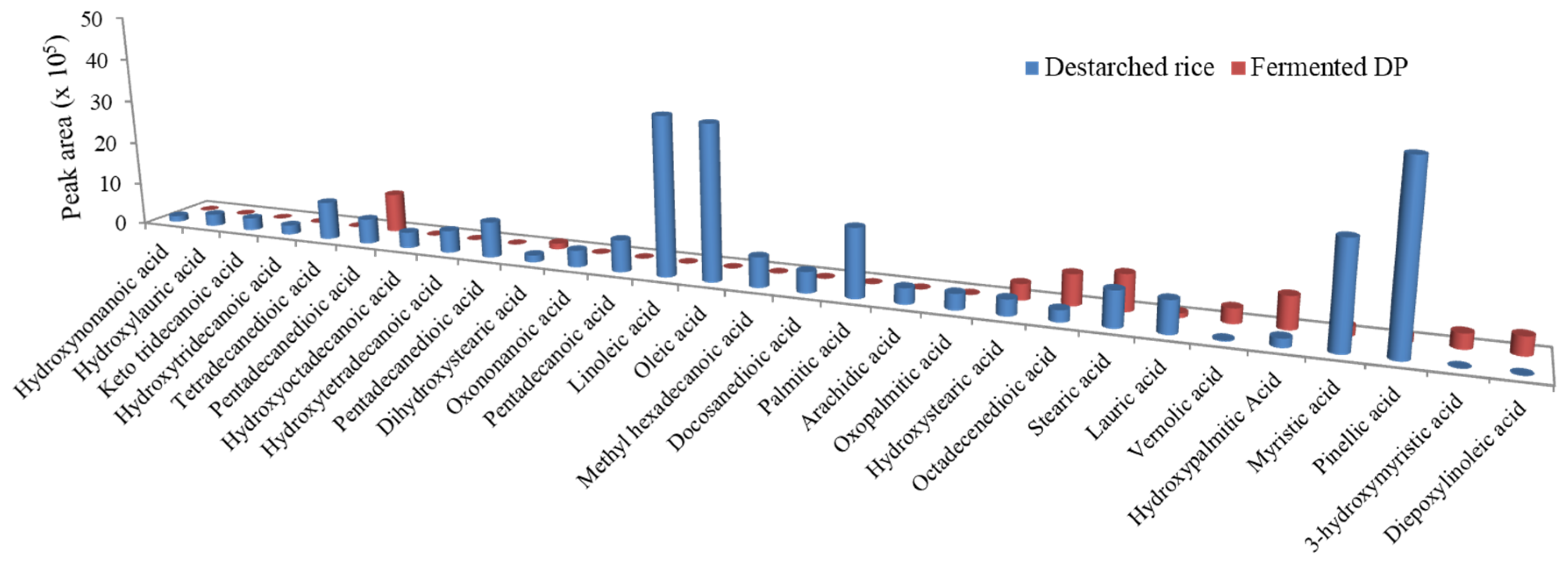
3.4. E. faecium EBD1fermentation Reduces Lipid Levels in Rice
3.5. E. faecium EBD1fermented Rice Is Enriched with Antihypertensive Peptides
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zhu, F. Anthocyanins in cereals: Composition and health effects. Food Res. Int. 2018, 109, 232–249. [Google Scholar] [CrossRef] [PubMed]
- Das, A.J.; Das, G.; Miyaji, T.; Deka, S.C. In vitro antioxidant activities of polyphenols purified from four plant species used in rice beer preparation in Assam India. Int. J. Food Prop. 2016, 19, 636–651. [Google Scholar] [CrossRef]
- Stagos, D. Antioxidant activity of polyphenolic plant extracts. Antioxidants 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Du, G.-L.; Feng, B.; Shao, H. Effects of oxidative stress on blood pressure and electrocardiogram findings in workers with occupational exposure to lead. Int. J. Med. Res. 2019, 47, 2461–2470. [Google Scholar] [CrossRef] [PubMed]
- Montezano, A.C.; Touyz, R.M. Molecular mechanisms of hypertension—Reactive oxygen species and antioxidants: A basic science update for the clinician. Can. J. Cardiol. 2012, 28, 288–295. [Google Scholar] [CrossRef] [PubMed]
- Alaaeddine, R.; Elkhatib, M.A.; Mroueh, A.; Fouad, H.; Saad, E.I.; El-Sabban, M.E.; Plane, F.; El-Yazbi, A.F. Impaired endothelium-dependent hyperpolarization underlies endothelial dysfunction during early metabolic challenge: Increased ROS generation and possible interference with NO function. J. Pharmacol. Exp. Ther. 2019, 371, 567–582. [Google Scholar] [CrossRef] [PubMed]
- Mathew, E.; Mukkadan, J. Clinical disparity of oxidative stress with blood pressure. J. Biomed. Sci. 2019, 3. [Google Scholar] [CrossRef]
- Kruger, R.; Schutte, R.; Huisman, H.; Van Rooyen, J.; Malan, N.; Fourie, C.; Louw, R.; Van der Westhuizen, F.; Van Deventer, C.; Malan, L. Associations between reactive oxygen species, blood pressure and arterial stiffness in black South Africans: The SABPA study. J. Hum. Hypertens. 2012, 26, 91–97. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current perspectives on antihypertensive probiotics. Probiotics Antimicrob. 2017, 9, 91–101. [Google Scholar] [CrossRef]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef]
- Dong, J.; Xu, X.; Liang, Y.; Head, R.; Bennett, L. Inhibition of angiotensin converting enzyme activity by polyphenols from tea (Camellia sinensis) and links to processing method. Food Funct. 2011, 2, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Persson, I.A.-L.; Persson, K.; Andersson, R.G. Effect of Vaccinium myrtillus and its polyphenols on angiotensin-converting enzyme activity in human endothelial cells. J. Agric. Food Chem. 2009, 57, 4626–4629. [Google Scholar] [CrossRef] [PubMed]
- Al Shukor, N.; Van Camp, J.; Gonzales, G.B.; Staljanssens, D.; Struijs, K.; Zotti, M.J.; Raes, K.; Smagghe, G. Angiotensin-converting enzyme inhibitory effects by plant phenolic compounds: A study of structure activity relationships. J. Agric. Food Chem. 2013, 61, 11832–11839. [Google Scholar] [CrossRef] [PubMed]
- Kontani, N.; Omae, R.; Kagebayashi, T.; Kaneko, K.; Yamada, Y.; Mizushige, T.; Kanamoto, R.; Ohinata, K. Characterization of Ile-His-Arg-Phe, a novel rice-derived vasorelaxing peptide with hypotensive and anorexigenic activities. Mol. Nutr. Food Res. 2014, 58, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Liu, S.; Ye, R.; Cai, G.; Ji, B.; Wu, Y. Angiotensin-I converting enzyme inhibitory tripeptides from rice protein hydrolysate: Purification and characterization. J. Funct. Foods 2013, 5, 1684–1692. [Google Scholar] [CrossRef]
- Pinciroli, M.; Aphalo, P.; Nardo, A.E.; Añón, M.C.; Quiroga, A.V. Broken rice as a potential functional ingredient with inhibitory activity of renin and angiotensin-converting enzyme. Plant Foods Hum. Nutr. 2019, 74, 405–413. [Google Scholar] [CrossRef]
- Aluko, R.E. Antihypertensive peptides from food proteins. Annu. Rev. Food Sci. Technol. 2015, 6, 235–262. [Google Scholar] [CrossRef]
- Oboh, G.; Ademiluyi, A.; Akindahunsi, A. Changes in polyphenols distribution and antioxidant activity during fermentation of some underutilized legumes. Food Sci. Technol. Int. 2009, 15, 41–46. [Google Scholar] [CrossRef]
- Rubak, Y.T.; Nuraida, L.; Iswantini, D.; Prangdimurti, E. Production of antihypertensive bioactive peptides in fermented food by lactic acid bacteria—A review. Carpath. J. Food Sci. Technol. 2019, 11, 29–44. [Google Scholar]
- Zhao, S.; Zhao, J.; Bu, D.; Sun, P.; Wang, J.; Dong, Z. Metabolomics analysis reveals large effect of roughage types on rumen microbial metabolic profile in dairy cows. Lett. Appl. Microbiol. 2014, 59, 79–85. [Google Scholar] [CrossRef]
- Goldansaz, S.A.; Guo, A.C.; Sajed, T.; Steele, M.A.; Plastow, G.S.; Wishart, D.S. Livestock metabolomics and the livestock metabolome: A systematic review. PLoS ONE 2017, 12, e0177675. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Dong, G.; Wang, Z.; Wang, J.; Zhang, Z.; Liu, J. Rumen and plasma metabolomics profiling by UHPLC-QTOF/MS revealed metabolic alterations associated with a high-corn diet in beef steers. PLoS ONE 2018, 13, e0208031. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Pinkard, B.R.; Gorman, D.J.; Rasmussen, E.G.; Maheshwari, V.; Kramlich, J.C.; Reinhall, P.G.; Novosselov, I.V. Raman spectroscopic data from formic acid decomposition in subcritical and supercritical water. Data Brief 2020, 29, 105312. [Google Scholar] [CrossRef] [PubMed]
- Tung, L. Method of calculating molecular weight distribution function from gel permeation chromatograms. J. Appl. Polym. Sci. 1966, 10, 375–385. [Google Scholar] [CrossRef]
- Berente, B.; De la Calle García, D.; Reichenbächer, M.; Danzer, K. Method development for the determination of anthocyanins in red wines by high-performance liquid chromatography and classification of German red wines by means of multivariate statistical methods. J. Chromatogr. A 2000, 871, 95–103. [Google Scholar] [CrossRef]
- Zhou, Z.; Robards, K.; Helliwell, S.; Blanchard, C. Composition and functional properties of rice. Int. J. Food Sci. Technol. 2002, 37, 849–868. [Google Scholar] [CrossRef]
- Suwannapan, O.; Wachirattanapongmetee, K.; Thawornchinsombut, S.; Katekaew, S. Angiotensin-I-converting enzyme (ACE)-inhibitory peptides from Thai jasmine rice bran protein hydrolysates. Int. J. Food Sci. 2020, 55, 2441–2450. [Google Scholar] [CrossRef]
- Cirlini, M.; Ricci, A.; Galaverna, G.; Lazzi, C. Application of lactic acid fermentation to elderberry juice: Changes in acidic and glucidic fractions. LWT Food Sci. Technol. 2020, 118, 108779. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, Q.; Lu, A.; Liu, X.; Zhang, L.; Xu, C.; Liu, X.; Li, H.; Yang, T. Sodium butyrate suppresses angiotensin II-induced hypertension by inhibition of renal (pro)renin receptor and intrarenal renin–angiotensin system. J. Hypertens. 2017, 35, 1899–1908. [Google Scholar] [CrossRef]
- Bays, H.E.; Maccubbin, D.; Meehan, A.G.; Kuznetsova, O.; Mitchel, Y.B.; Paolini, J.F. Blood pressure-lowering effects of extended-release niacin alone and extended-release niacin/laropiprant combination: A post hoc analysis of a 24-week, placebo-controlled trial in dyslipidemic patients. Clin. Ther. 2009, 31, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Dai, T.; Chen, J.; McClements, D.J.; Hu, P.; Ye, X.; Liu, C.; Li, T. Protein–polyphenol interactions enhance the antioxidant capacity of phenolics: Analysis of rice glutelin–procyanidin dimer interactions. Food Func. 2019, 10, 765–774. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Prahalathan, P.; Raja, B. Antihypertensive and antioxidant potential of vanillic acid, a phenolic compound in L-NAME-induced hypertensive rats: A dose-dependence study. Redox Rep. 2011, 16, 208–215. [Google Scholar] [CrossRef] [PubMed]
- Peixoto-Neves, D.; Wang, Q.; Leal-Cardoso, J.H.; Rossoni, L.V.; Jaggar, J.H. Eugenol dilates mesenteric arteries and reduces systemic BP by activating endothelial cell TRPV 4 channels. Br. J. Pharmacol. 2015, 172, 3484–3494. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, M.; Raja, B. Veratric acid, a phenolic acid attenuates blood pressure and oxidative stress in L-NAME induced hypertensive rats. Eur. J. Pharmacol. 2011, 671, 87–94. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Piao, Z.H.; Sun, S.; Liu, B.; Kim, G.R.; Seok, Y.M.; Lin, M.Q.; Ryu, Y.; Choi, S.Y.; Kee, H.J. Gallic acid reduces blood pressure and attenuates oxidative stress and cardiac hypertrophy in spontaneously hypertensive rats. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef]
- Luo, D.; Xu, J.; Chen, X.; Zhu, X.; Liu, S.; Li, J.; Xu, X.; Ma, X.; Zhao, J.; Ji, X. (−)-Epigallocatechin-3-gallate (EGCG) attenuates salt-induced hypertension and renal injury in Dahl salt-sensitive rats. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Sui, H.; Yan, W. Effect of apigenin on SBP of spontaneous hypertension rats and its mechanism. J. Environ. Health 2009, 26, 112–113. [Google Scholar]
- Yusuf, M.A.; Singh, B.N.; Sudheer, S.; Kharwar, R.N.; Siddiqui, S.; Abdel-Azeem, A.M.; Fernandes Fraceto, L.; Dashora, K.; Gupta, V.K. Chrysophanol: A natural anthraquinone with multifaceted biotherapeutic potential. Biomolecules 2019, 9, 68. [Google Scholar]
- Chandrasekara, A.; Shahidi, F. Bioaccessibility and antioxidant potential of millet grain phenolics as affected by simulated in vitro digestion and microbial fermentation. J. Funct. Foods 2012, 4, 226–237. [Google Scholar] [CrossRef]
- Dey, T.B.; Kuhad, R.C. Enhanced production and extraction of phenolic compounds from wheat by solid-state fermentation with Rhizopus oryzae RCK2012. Biotech. Rep. 2014, 4, 120–127. [Google Scholar]
- Liptáková, D.; Matejčeková, Z.; Valík, L. Lactic acid bacteria and fermentation of cereals and pseudocereals. Ferment. Process. 2017, 10, 65459. [Google Scholar]
- Wang, Z.J.; Liang, C.L.; Li, G.M.; Yu, C.Y.; Yin, M. Stearic acid protects primary cultured cortical neurons against oxidative stress4. Acta Pharmacol. Sin. 2007, 28, 315–326. [Google Scholar] [CrossRef]
- Alves, N.F.B.; de Queiroz, T.M.; de Almeida Travassos, R.; Magnani, M.; de Andrade Braga, V. Acute treatment with lauric acid reduces blood pressure and oxidative stress in spontaneously hypertensive rats. Basic Clin. Pharmacol. Toxicol. 2017, 120, 348–353. [Google Scholar] [CrossRef] [PubMed]
- Antony, U.; Sripriya, G.; Chandra, T. The effect of fermentation on the primary nutrients in foxtail millet (Setaria italica). Food Chem. 1996, 56, 381–384. [Google Scholar] [CrossRef]
- Umezawa, H.; Aoyagi, T.; Ogawa, K.; Naganawa, H.; Hamada, M.; Takeuchi, T. Diprotins A and B, inhibitors of dipeptidyl aminopeptidase IV, produced by bacteria. J. Antibiot. 1984, 37, 422–425. [Google Scholar] [CrossRef]
- Lan, V.T.T.; Ito, K.; Ohno, M.; Motoyama, T.; Ito, S.; Kawarasaki, Y. Analyzing a dipeptide library to identify human dipeptidyl peptidase IV inhibitor. Food Chem. 2015, 175, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Cosenso-Martin, L.N.; Giollo-Junior, L.T.; Vilela-Martin, J.F. DPP-4 inhibitor reduces central blood pressure in a diabetic and hypertensive patient: A case report. Medicine 2015, 94. [Google Scholar] [CrossRef]
- Pacheco, B.P.; Crajoinas, R.O.; Couto, G.K.; Davel, A.P.C.; Lessa, L.M.; Rossoni, L.V.; Girardi, A.C. Dipeptidyl peptidase IV inhibition attenuates blood pressure rising in young spontaneously hypertensive rats. Int. J. Hypertens. 2011, 29, 520–528. [Google Scholar] [CrossRef]
- Zhang, Y.; Pechan, T.; Chang, S.K.C. Antioxidant and angiotensin-I converting enzyme inhibitory activities of phenolic extracts and fractions derived from three phenolic-rich legume varieties. J. Funct. Foods 2018, 42, 289–297. [Google Scholar] [CrossRef]
- Santos, M.C.; Toson, N.S.B.; Pimentel, M.C.B.; Bordignon, S.A.L.; Mendez, A.S.L.; Henriques, A.T. Polyphenols composition from leaves of Cuphea spp. and inhibitor potential, in vitro, of angiotensin I-converting enzyme (ACE). J. Ethnopharmacol. 2020, 255, 112781. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, H.; Fu, X.; Li, S.; Wei, J. A novel antioxidant and ACE inhibitory peptide from rice bran protein: Biochemical characterization and molecular docking study. LWT Food Sci. Technol. 2017, 75, 93–99. [Google Scholar] [CrossRef]
- Wang, X.; Wang, J.; Lin, Y.; Ding, Y.; Wang, Y.; Cheng, X.; Lin, Z. QSAR study on angiotensin-converting enzyme inhibitor oligopeptides based on a novel set of sequence information descriptors. J. Mol. Model. 2011, 17, 1599–1606. [Google Scholar] [CrossRef] [PubMed]
- Yano, S.; Suzuki, K.; Funatsu, G. Isolation from α-zein of thermolysin peptides with angiotensin I-converting enzyme inhibitory activity. Biosci. Biotechnol. Biochem. 1996, 60, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Cheung, H.-S.; Wang, F.-L.; Ondetti, M.A.; Sabo, E.F.; Cushman, D.W. Binding of peptide substrates and inhibitors of angiotensin-converting enzyme. Importance of the COOH-terminal dipeptide sequence. J. Biol. Chem. 1980, 255, 401–407. [Google Scholar] [PubMed]
- Zhou, P.; Yang, C.; Ren, Y.; Wang, C.; Tian, F. What are the ideal properties for functional food peptides with antihypertensive effect? A computational peptidology approach. Food Chem. 2013, 141, 2967–2973. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Ledesma, B.; Recio, I.; Ramos, M.; Amigo, L. Preparation of ovine and caprine β-lactoglobulin hydrolysates with ACE-inhibitory activity. Identification of active peptides from caprine β-lactoglobulin hydrolysed with thermolysin. Int. Dairy J. 2002, 12, 805–812. [Google Scholar] [CrossRef]
- Castellano, P.; Aristoy, M.-C.; Sentandreu, M.Á.; Vignolo, G.; Toldrá, F. Peptides with angiotensin I converting enzyme (ACE) inhibitory activity generated from porcine skeletal muscle proteins by the action of meat-borne Lactobacillus. J. Proteom. 2013, 89, 183–190. [Google Scholar] [CrossRef]
- Wu, H.; He, H.-L.; Chen, X.-L.; Sun, C.-Y.; Zhang, Y.-Z.; Zhou, B.-C. Purification and identification of novel angiotensin-I-converting enzyme inhibitory peptides from shark meat hydrolysate. Process Biochem. 2008, 43, 457–461. [Google Scholar] [CrossRef]
- Ichimura, T.; Hu, J.; Aita, D.Q.; Maruyama, S. Angiotensin I-converting enzyme inhibitory activity and insulin secretion stimulative activity of fermented fish sauce. J. Biosci. Bioeng. 2003, 96, 496–499. [Google Scholar] [CrossRef]
- Byun, H.-G.; Kim, S.-K. Structure and activity of angiotensin I converting enzyme inhibitory peptides derived from Alaskan pollack skin. BMB Rep. 2002, 35, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Suetsuna, K.; Ukeda, H.; Ochi, H. Isolation and characterization of free radical scavenging activities peptides derived from casein. J. Nutr. Biochem. 2000, 11, 128–131. [Google Scholar] [CrossRef]
- Kohmura, M.; Nio, N.; Kubo, K.; Minoshima, Y.; Munekata, E.; Ariyoshi, Y. Inhibition of angiotensin-converting enzyme by synthetic peptides of human β-casein. Agric. Biol. Chem. 1989, 53, 2107–2114. [Google Scholar]
- Gu, Y.; Majumder, K.; Wu, J. QSAR-aided in silico approach in evaluation of food proteins as precursors of ACE inhibitory peptides. Food Res. Int. 2011, 44, 2465–2474. [Google Scholar] [CrossRef]
- Tauzin, J.; Miclo, L.; Gaillard, J.-L. Angiotensin-I-converting enzyme inhibitory peptides from tryptic hydrolysate of bovine αS2-casein. FEBS Lett. 2002, 531, 369–374. [Google Scholar] [CrossRef]
- Nogata, Y.; Nagamine, T.; Yanaka, M.; Ohta, H. Angiotensin I converting enzyme inhibitory peptides produced by autolysis reactions from wheat bran. J. Agric. Food Chem. 2009, 57, 6618–6622. [Google Scholar] [CrossRef]
- Escudero, E.; Sentandreu, M.A.; Arihara, K.; Toldra, F. Angiotensin I-converting enzyme inhibitory peptides generated from in vitro gastrointestinal digestion of pork meat. J. Agric. Food Chem. 2010, 58, 2895–2901. [Google Scholar] [CrossRef]
- He, R.; Malomo, S.A.; Girgih, A.T.; Ju, X.; Aluko, R.E. Glycinyl-histidinyl-serine (GHS), a novel rapeseed protein-derived peptide has blood pressure-lowering effect in spontaneously hypertensive rats. J. Agric. Food Chem. 2013, 61, 8396–8402. [Google Scholar] [CrossRef]
- Nakahara, T.; Sano, A.; Yamaguchi, H.; Sugimoto, K.; Chikata, H.; Kinoshita, E.; Uchida, R. Antihypertensive effect of peptide-enriched soy sauce-like seasoning and identification of its angiotensin I-converting enzyme inhibitory substances. J. Agric. Food Chem. 2010, 58, 821–827. [Google Scholar] [CrossRef]
- Del Mar Contreras, M.; Carrón, R.; Montero, M.J.; Ramos, M.; Recio, I. Novel casein-derived peptides with antihypertensive activity. Int. Dairy J. 2009, 19, 566–573. [Google Scholar] [CrossRef]
- Minervini, F.; Algaron, F.; Rizzello, C.; Fox, P.; Monnet, V.; Gobbetti, M. Angiotensin I-converting-enzyme-inhibitory and antibacterial peptides from Lactobacillus helveticus PR4 proteinase-hydrolyzed caseins of milk from six species. Appl. Environ. Microbiol. 2003, 69, 5297–5305. [Google Scholar] [CrossRef] [PubMed]
- Udenigwe, C.C.; Adebiyi, A.P.; Doyen, A.; Li, H.; Bazinet, L.; Aluko, R.E. Low molecular weight flaxseed protein-derived arginine-containing peptides reduced blood pressure of spontaneously hypertensive rats faster than amino acid form of arginine and native flaxseed protein. Food Chem. 2012, 132, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Aluko, R.E.; Nakai, S. Structural requirements of angiotensin I-converting enzyme inhibitory peptides: Quantitative structure− activity relationship study of di-and tripeptides. J. Agric. Food Chem. 2006, 54, 732–738. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time | Peak Area | Adduct/Charge | Precursor Mass | Found at Mass | Formula Finder Results | Peptides | Similar Peptides Reported in Literature* | Peptide Function | Reference |
|---|---|---|---|---|---|---|---|---|---|
| 0.93 | 221,600 | [M-H]- | 227.116 | 227.115 | C9H16N4O3 | GPG | GPG | Antihypertensive | [53] |
| 0.97 | 1,227,000 | [M-H]- | 516.206 | 516.2053 | C19H31N7O10 | QQQD | LQQQ | Antihypertensive | [54] |
| 1.31 | 506,800 | [M-H]- | 185.058 | 185.0572 | C7H10N2O4 | EG | EG | Antihypertensive | [55] |
| 3.16 | 501,500 | [M-H]- | 419.121 | 419.1206 | C18H20N4O8 | GGG | GGG | Antihypertensive | [56] |
| 8.22 | 292,200 | [M-H]- | 442.232 | 442.2311 | C19H33N5O7 | PKEA | VDKEA | Antihypertensive | [57] |
| 8.4 | 219,500 | [M-H]- | 426.237 | 426.2363 | C19H33N5O6 | GVGVP | Not found | - | - |
| 8.63 | 2,611,000 | [M-H]- | 227.105 | 227.1039 | C10H16N2O4 | EV | EV | Antihypertensive | [58] |
| 9.23 | 1,306,000 | [M-H]- | 291.1 | 291.0989 | C14H16N2O5 | EY | EY | Antihypertensive | [59] |
| 9.48 | 192,900 | [M-H]- | 544.228 | 544.2266 | C23H31N9O7 | WGGGGGG | GGG | Antihypertensive | [56] |
| 9.87 | 10,460 | [M-H]- | 293.116 | 293.1143 | C14H18N2O5 | DF | DF | Antihypertensive | [60] |
| 10.29 | 210,900 | [M-H]- | 326.209 | 326.2087 | C16H29N3O4 | VPL | VPL | DPP IV inhibitor | [46] |
| 10.6 | 1,264,000 | [M-H]- | 383.231 | 383.2305 | C18H32N4O5 | PLG | PLG | Antihypertensive | [61] |
| 11.56 | 195,100 | [M-H]- | 453.201 | 453.1996 | C20H30N4O8 | PDGA | Not found | - | - |
| 11.61 | 3,821,000 | [M-H]- | 241.121 | 241.1196 | C11H18N2O4 | EL | EL | Antioxidative | [62] |
| 11.81 | 432,500 | [M-H]- | 489.273 | 489.2725 | C25H38N4O6 | VLPY | VLPYP | Antihypertensive | [63] |
| 11.84 | 243,600 | [M-H]- | 518.227 | 518.2263 | C24H33N5O8 | DISW | ISW | Antihypertensive | [64] |
| 11.91 | 478,100 | [M-H]- | 395.195 | 395.1941 | C18H28N4O6 | YLS | YL | Antihypertensive | [65] |
| 11.92 | 267,600 | [M-H]- | 319.135 | 319.1335 | C13H24N2O5S | MG | MG | Antihypertensive | [66] |
| 12.12 | 249,800 | [M-H]- | 473.209 | 473.2079 | C20H34N4O7S | EMVP | MMVPI | Antihypertensive | [67] |
| 12.94 | 1,020,000 | [M-H]- | 489.237 | 489.2361 | C24H34N4O7 | GPLA | GPL | Antihypertensive | [68] |
| 13.0 | 471,200 | [M-H]- | 774.334 | 774.3331 | C44H49N5O6S | AW | AW | Antihypertensive | [69] |
| 13.3 | 187,800 | [M-H]- | 180.974 | 180.9733 | C2H2N2O8 | GG | GG | Antihypertensive | [58] |
| 13.41 | 1,627,000 | [M-H]- | 496.316 | 496.3145 | C24H43N5O6 | IVPVA | KPVAL | Antihypertensive | [70] |
| 13.42 | 1,415,000 | [M-H]- | 528.284 | 528.2836 | C27H39N5O6 | FPIV | GPFPIIV | Antihypertensive | [70] |
| 14.17 | 1,171,000 | [M-H]- | 510.331 | 510.3303 | C25H45N5O6 | GGGGG | GGG | Antihypertensive | [56] |
| 14.24 | 409,400 | [M-H]- | 482.263 | 482.2627 | C22H37N5O7 | VVPQ | VPQ | Antihypertensive | [71] |
| 15.88 | 153.2 | [M-H]- | 743.35 | 743.3477 | C33H56N6O9S2 | VMV | MV | DPP IV inhibitor | [47] |
| 17.11 | 14,500 | [M-H]- | 267.067 | 267.0646 | C8H16N2O6S | CE | Not found | - | - |
| 17.22 | 1,126,000 | [M-H]- | 310.165 | 310.1637 | C12H21N7O3 | HR | HR | DPP IV inhibitor | [47] |
| 17.8 | 360,400 | [M-H]- | 457.169 | 457.1688 | C17H34N2O8S2 | CC | CCD | Antihypertensive | [72] |
| 21.12 | 43,240 | [M-H]- | 474.264 | 474.2634 | C22H41N3O6S | ML | RML | Antihypertensive | [73] |
| 22.19 | 29,960 | [M-H]- | 473.284 | 473.2837 | C19H38N8O6 | KTAR | KTAP | Antihypertensive | [63] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daliri, E.B.-M.; Ofosu, F.K.; Chelliah, R.; Kim, J.-H.; Kim, J.-R.; Yoo, D.; Oh, D.-H. Untargeted Metabolomics of Fermented Rice Using UHPLC Q-TOF MS/MS Reveals an Abundance of Potential Antihypertensive Compounds. Foods 2020, 9, 1007. https://doi.org/10.3390/foods9081007
Daliri EB-M, Ofosu FK, Chelliah R, Kim J-H, Kim J-R, Yoo D, Oh D-H. Untargeted Metabolomics of Fermented Rice Using UHPLC Q-TOF MS/MS Reveals an Abundance of Potential Antihypertensive Compounds. Foods. 2020; 9(8):1007. https://doi.org/10.3390/foods9081007
Chicago/Turabian StyleDaliri, Eric Banan-Mwine, Fred Kwame Ofosu, Ramachandran Chelliah, Joong-Hark Kim, Jong-Rae Kim, Daesang Yoo, and Deog-Hwan Oh. 2020. "Untargeted Metabolomics of Fermented Rice Using UHPLC Q-TOF MS/MS Reveals an Abundance of Potential Antihypertensive Compounds" Foods 9, no. 8: 1007. https://doi.org/10.3390/foods9081007
APA StyleDaliri, E. B.-M., Ofosu, F. K., Chelliah, R., Kim, J.-H., Kim, J.-R., Yoo, D., & Oh, D.-H. (2020). Untargeted Metabolomics of Fermented Rice Using UHPLC Q-TOF MS/MS Reveals an Abundance of Potential Antihypertensive Compounds. Foods, 9(8), 1007. https://doi.org/10.3390/foods9081007